Introduction
Nasopharyngeal carcinoma (NPC), a malignancy of
epithelial origin, is predominantly endemic in Southern China with
an age-standardized incidence rate of 30–50 cases per 100,000
individuals/year (1). The incidence
rate for males is as high as >40 cases out of 100,000
individuals/year in the Cantonese-speaking population in Guangdong
Province, which is significantly higher than that in the other
major dialect-speaking populations who immigrated to Guangdong
Province 100 years ago from Northern China (2). Importantly, the incidence of NPC in
these populations peaks at the relatively young age of 45
years.
NPC in Southern China is closely linked to infection
of the Epstein-Barr virus (EBV) since it was found that titers of
antibodies against EBV are elevated in NPC patients, viral DNA and
proteins are present in tumor cells with a monoclonal nature, and
part of the viral genes or proteins shows transformation potential
in lymphocytes and epithelial cell lines (3,4). The
mechanism of how EBV enters into the nasopharyngeal epithelium (NE)
has not yet been conclusively elucidated; but at least two
receptors, the complement receptor type 2 (CR2) (5) and the polymeric immunoglobulin
receptor (pIgR) (6), have been
proposed. CR2 (CD21), a 145-kDa integral membrane glycoprotein, is
the human C3d and EBV receptor. The primary ligand for CR2 is C3d,
a processed form of C3 which results from proteolytic cleavage of
C3b. CR2 also serves as the EBV receptor on human B cells during
EBV infection. It was indicated that a large surface glycoprotein
of EBV, gp350/220, is the viral ligand that can bind to CR2 and
thus EBV can infect the cells. EBV can also infect recombinant
epithelial cells expressing CR2 especially when the cells are in
contact with virus-producing lymphocytes (7). In addition, it has been reported that
CR is expressed in embryonic NE cells as revealed by RT-PCR,
indicating that CR2 may be a sideway of EBV-NE cell infection
(8).
Recent studies have shown that CR2 polymorphisms may
be associated with a few immunologically mediated diseases such as
systemic lupus erythematosus (9,10) and
multiple sclerosis (11). Notably,
upregulation or downregulation of CR2 was found in different cases.
However, to date, no major susceptibility gene such as BRCA1 for
breast cancer, has been identified for NPC with significantly
increased risk, although several genes including HLA haplotypes
(12) and genes of T cell receptors
(13), cytochrome P450 2E1
(14), TLR family (15,16),
DNA repair enzymes XRCC1 and hOGG1 (17), have been reported to be associated
with the risk of NPC. The specific aim of this study was to
ascertain whether CR2 is a major NPC susceptibility gene. Our study
was conducted using candidate-gene approaches to determine the risk
associated with DNA polymorphisms among NPC patients of a
Cantonese-based population. Additionally, the basic function of the
CR2 promoter and 5′-UTR was also examined in the present study. Our
findings indicate that enhanced CR2 expression may be involved in
the oncogenesis and development of NPC.
Materials and methods
Variation screening. The CR2 sequence was
obtained from the published database of the National Center for
Biotechnology Information of USA. All the annotations in the
databases for all known exons and untranslated region of CR2 were
used. A standard polymerase chain reaction (PCR) method routinely
followed in our laboratory, as mentioned later, was employed for
all exons, 5′-untranslated region (5′-UTR) and 3′-untranslated
region (3′-UTR) of CR2. The primers for the target regions of CR2
were designed with the web-base software Primer 3.0. In the
preliminary tests, we amplified and purified the DNA samples from
24 patients with confirmed sporadic NPC in Southern China. These
samples included 48 chromosomes, providing at least a 95% confident
level to detect alleles with frequencies >5%. The PCR products
were then sequenced by ABI®3730XL Automatic Sequencer
(Applied Biosystems, Foster City, CA, USA). We used the
Polyphred/Phredphrap/Consed software package to identified
single-nucleotide polymorphism (SNP) candidates that were then
confirmed by two independent observers. These SNP positions and
individual genotypes were further confirmed using re-amplifying and
sequencing and again reversely.
Study population. The subjects of this
case-control study consisted of 528 patients with
histopathologically confirmed NPC and 408 population controls. All
subjects were unrelated ethnic Cantonese from the
Cantonese-speaking population in Nanhai, Foshan, Shunde, Qingyuan,
Sihui and Luoding regions of Guangdong Province, China. Patients
were recruited consecutively from December 2003 to October 2004 at
the Sun Yat-sen University Cancer Center, Guangzhou, China. The
average age for all 528 patients at diagnosis of NPC was <50
years. Population controls were cancer-free individuals and
non-relatives of the patients; and they were randomly selected with
a community cancer-screening program for early detection of cancer
during the same period as the cases were enrolled. The selection
criteria for control subjects included: i) no individual history of
cancer; ii) frequency matched to NPC cases according to gender, age
(±5 years); iii) the residential region; and iv) the time period
for blood sample collection. At recruitment, informed consent was
obtained from each subject, and each participant was then
interviewed to collect detailed information on demographic
characteristics. This study was approved by the Human Ethics
Approval Committee of the Cancer Center of Sun Yat-sen University,
China.
Genotype analysis. Variations among genotypes
were determined by the PCR-based DNA direct sequencing. Genomic DNA
was extracted from the peripheral blood samples of all case and
control subjects using the DNAzol kit according to the
manufacturer’s protocol (Gibco-BRL, Life Technologies, Carlsbad,
CA, USA). The genomic DNA samples were amplified by PCR using a
GeneAmp® 9700 PCR system (Applied Biosystems). The 20-μl
PCR reaction mixture contained 20 ng DNA, 0.2 μM of each primer,
200 μM of each deoxynucleotide triphosphate, 1.5 mM of
MgCl2, 0.3 units of Taq DNA polymerase with 1X buffer
(Qiagen, Chatsworth, CA, USA). The primers used to amplify DNA
containing the 5′-UTR region were 5′-agtgggttgcgtggtcaaaa-3′ and
5′-agtggggacaatcagga cca-3′, which produce a 550-bp fragment; the
primers used to amplify DNA containing exon 10 were
5′-ataccgtccaggaaac aac-3′ and 5′-cctctttccatgatgcagtt-3′, which
produce a 606-bp fragment; the primers used to amplify DNA
containing exon 17 were 5′-ggtggactggatcaaatcag-3′ and
5′-gggcttccttttgtatag cac-3′, which produce a 420-bp fragment. The
reactions were carried out under the following conditions: an
initial melting step of 15 min at 95°C; followed by 30 cycles at
which each one consisted of 30 sec at 94°C, 30 sec at 60°C and 45
sec at 72°C, respectively; and a final elongation step of 5 min at
72°C. Automatic DNA sequencing was performed on an ABI®
3730XL Automatic Sequencer (Applied Biosystems) using direct PCR
products of the samples according to the manufacturer’s protocol.
The raw data were analyzed using Sequencing Analysis software V3.3
on a MAC operating system v9.1, Phred-Phrap-Consed software
package, DNAStar/TaqMan software package and Chromas. All
genotyping was performed blinded to the case/control status, and
blinded quality control samples were inserted to validate the
genotype. Random samples (10%) of cases and controls were sequenced
twice by different investigators to corroborate the findings, and
the reproducibility was 100%.
Preparation of the reporter constructs. The
entire 1327 bp (−1252 to +75) of the CR2 promoter was amplified
using the following primers: PPF, 5′-gatggtaccATGAAGCTTCCAG
CCAAAG-3′ and PPR, 5′-gatctcgagCGGCGGGATGCGTT
CCGAGA-3′. The sequence (−57 to +94, 151 bp) containing 5′-UTR of
CR2 was amplified using the primers PUF, 5′-gat
ggtaccTGCGCTCAGAACTAGCACGTGT-3′ and PUR, 5′-gat
ctcgagGCCACGGCCGAAGCCCCCGCG-3′. The design of the primers
incorporating the KpnI and XhoI sites at the 5′ end
(underlined primers) enabled cloning into the firefly luciferase
expression plasmid (pGL3-basic; Promega, Madison, WI, USA).
Distinct DNA from blood samples, each containing a different
genotype of 24T and C, were used as templates to obtain the
promoter and 5′-UTR sequence for cloning. The PCR conditions
consisted of 0.2 μM of each primer, 200 μM of each deoxynucleotide
triphosphate, 1.5 mM of MgCl2, 0.3 units of Taq DNA
polymerase with 1X buffer (Qiagen) in a final volume of 20 μl. PCR
was performed at 94°C for 2 min, followed by 25 cycles at 94°C for
30 sec, 55°C for 1 min and 72°C for 1 min. The final extension step
was at 72°C for 5 min. The pure PCR products were digested with
KpnI/XhoI and then were cloned into the
KpnI/XhoI site of pGL3-basic plasmids. We
discriminated them by sequencing, and named them PWILD and PMUT for
the variants of the promoter constructs, and UWILD and UMUT for the
variants of the 5′-UTR construct. Recombinant DNA cloning was
performed according to standard methodologies (18). Plasmid DNA for the transfections was
prepared using the Qiagen Plasmid Maxi kit (Qiagen, Hilden,
Germany) according to the manufacturer’s instructions. All
luciferase constructs were verified by sequencing.
Cell-based gene expression and luciferase
assay. Human NPC C666 cells were grown in RPMI-1640 medium
(Gibco Laboratories, Grand Island, NY, USA) containing 10%
heat-inactivated fetal bovine serum (FBS) and 1% L-glutamine
(Sigma, Chemical Co., St. Louis, MO, USA) in a humidified chamber
maintained at 37°C containing 5% CO2. Cells in a 24-well
plate were grown to 70% confluence prior to the transfections. Each
construct (1.5 μg) was cotransfected with pRL-EF (25 ng) as an
internal control using 2 μl of Lipofectamine 2000 (Invitrogen,
Carlsbad, CA, USA). The control plasmid contained the pGL3-control,
rather than the construct. This control was cotransformed with the
pRL-EF plasmid as a positive control for luciferase expression.
Forty-eight and sixty hours later, the cells were harvested and
assayed for firefly luciferase and Renilla luciferase
activities using Dual Luciferase Reporter Assay system (Promega).
The relative firefly luciferase activities were normalized against
the Renilla luciferase activities.
Statistical analysis. For each polymorphism,
deviation of the genotype frequencies in the control subjects from
those expected under Hardy-Weinberg equilibrium was assessed using
the standard Chi-square test. The distributions of genotypes
between case patients and control subjects were compared using the
Monte Carlo approach (19) (data
not shown). We estimated the cancer risk associated with the
alleles, genotypes as odds ratios (ORs) and 95% confidence
intervals (CIs) with adjustment particularly for age using SPSS
software (version 11.0). All statistical tests were two-sided, and
the probability level <0.05 was used as the criterion of
significant statistical difference. In in vitro experiments,
the transfection and luciferase assays were performed in
triplicate. The data were analyzed using the ANOVA test.
Statistical significant differences were considered for P<0.05,
and data points represent the means ± SD.
Results
Study sample and variation screening. The
blood DNA samples from 528 sporadic NPC cases and from 408 controls
were screened for possible SNPs of CR2. No significant statistical
differences in the age (P=0.0795) and gender (P=0.6266)
distributions were observed between the NPC patients and controls,
suggesting that the frequency matching was adequate (Table I). In order to identify SNPs with a
>5% frequency in the NPC cases, we used a relatively small
sample containing 24 NPC cases. We sequenced all the coding exons,
the promoter region and the exon-intron boundary regions of CR2 for
detection of possible alterations. We detected 3 SNPs in CR2
including 1 in exon 1 (24 T>C, rs3813946), 1 in exon 10 (18650
G/A, rs1048971) and 1 in IVS17 (25775 T/A, rs17258996), all of the
SNPs have already been registered in the NCBI database. Since 24
T>C in 5′-UTR and 25775 T/A in IVS17 were fully linked as a
haplotype of all the study population, and therefore had the same
frequency totally different from that in NCBI, only 24 T>C was
assessed in the following analysis.
 | Table IDistribution of characteristics of the
study subjects. |
Table I
Distribution of characteristics of the
study subjects.
| Cases (n=528) | Controls (n=408) | |
|---|
| Characteristics | n (%) | n (%) | P-valuea |
|---|
| Gender |
| Male | 388 (73.48) | 294 (72.05) | 0.6266 |
| Female | 140 (26.51) | 114 (27.94) | |
| Age (years) |
| ≤45 | 375 (71.02) | 267 (65.44) | 0.0795 |
| >45 | 153 (28.98) | 141 (34.56) | |
| Mean age | 42±18 | 43±10 | |
Association of individual SNPs with the risk of
NPC. We focused on 2 SNPs and detected their frequency in all
of the subjects including the 528 NPC cases and 408 controls. When
comparing the allelic frequency between the cases and controls, SNP
24 T>C was found to have a significant difference among the
cases and controls (P=0.0221, Table
II). After categorizing the cases into 2 age groups of ≤45 and
>45 years, the minor allele C frequency was 12.4% of cases in
the ≤45-year age group, whereas it was 6.9% in the controls. The
allelic frequencies in the cases were significantly different from
those of the controls (P=0.0034, Table III). The odds ratio (OR=1.81) also
showed a higher risk of NPC in individuals carrying the minor
alleles. The distributions of the other SNP 18650 G/A was not
different between the cases and controls (Tables II and III).
| Table IICR2 genotype frequencies of selected
SNPs and their contributions to the risk of NPC. |
Table II
CR2 genotype frequencies of selected
SNPs and their contributions to the risk of NPC.
| Cases (n=528) | Controls (n=408) | | |
|---|
| Genotype | n (%) | n (%) | P-value | OR (95% CI) |
|---|
| 24 T/C |
| TT | 415 (78.6) | 343 (84.1) | | |
| TC | 101 (19.1) | 60 (14.7) | C−/C+a | |
| CC | 12 (2.3) | 5 (1.2) | 0.0345 | 1.44 (1.03–2.01) |
| Allele C
frequency | 0.118 | 0.086 | 0.0221 | 1.43 (1.05–1.95) |
| 18650 G/A |
| GG | 376 (71.2) | 309 (75.7) | | |
| GA | 117 (22.2) | 72 (17.6) | A−/A+b | |
| AA | 35 (6.6) | 27 (6.7) | 0.1214 | 1.26
(0.94–1.69) |
| Allele A
frequency | 0.177 | 0.154 | 0.1924 | 1.18
(0.92–1.51) |
| Table IIICR2 genotype frequencies of selected
SNPs stratified by age and their associations with risks of
NPC. |
Table III
CR2 genotype frequencies of selected
SNPs stratified by age and their associations with risks of
NPC.
| Cases (n=375) | Controls
(n=267) | | |
|---|
| Genotype | n (%) | n (%) | P-value | OR (95% CI) |
|---|
| 24 T/C |
| Age ≤45 years |
| TT | 294 (78.4) | 233 (87.3) | | |
| TC | 73 (19.5) | 31 (11.6) | C−/C+a | |
| CC | 8 (2.1) | 3 (1.1) | 0.0039 | |
| Allele C
frequency | 0.124 | 0.069 | 0.0034 | 1.81
(1.21–2.70) |
| 18650 G/A |
| Age >45
years |
| GG | 124 (81.0) | 118 (83.7) | | |
| GA | 24 (15.7) | 16 (11.3) | A−/A+b | |
| AA | 5 (3.3) | 7 (5.0) | 0.5531 | |
| Allele A
frequency | 0.111 | 0.106 | 0.8541 | 1.05
(0.62–1.77) |
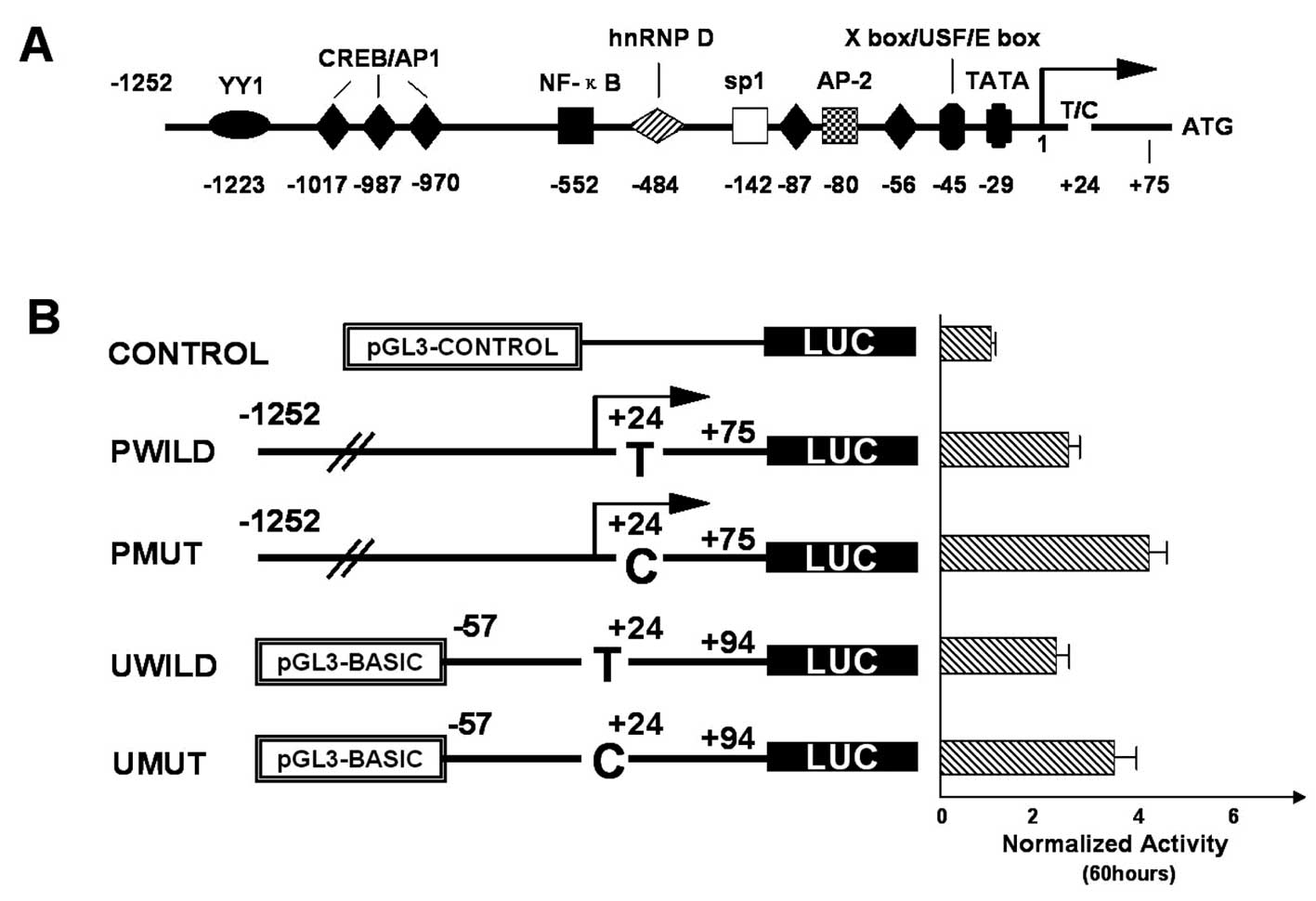
Comparison of luciferase activity among the
constructs. CR2 5′-UTR variants used in the present study were
obtained by PCR and were cloned into the pGL3/LUC basic or promoter
vectors (Fig. 1) as described in
Materials and methods. To compare 5′-UTR-mediated gene expression
by each of the four constructs, the C666 cells were transiently
transfected with the recombinant luciferase vectors (PMUT or PWILD,
UMUT or UWILD constructs, pGL3-control and pEF-RL). The
Renilla-normalized luciferase activities of the P group
(PMUT and PWILD) and U group (UMUT and UWILD) constructs were as
follows: 48 h after transfection, the reporter gene activity of the
MUT construct was higher than that of the WILD construct
(P<0.05) either in the P or U group. Moreover, 60 h after
transfection, the normalized luciferase activity was markedly
different, with the PMUT/UMUT construct increasing the reporter
gene activity by ~40–50% compared to that of the PWILD/UWILD
construct (P<0.01; Fig. 1).
Discussion
Genetic background plays an important role in the
development of NPC, which is an endemic multifactorial genetic
disease resulting from gene-environment-EBV interaction. Genetic
susceptibility plays a critical role in determining the individual
risk of NPC (20). We are in the
process of systematically searching for genetic susceptibility
genes for NPC. Besides those genes involved in carcinogen
metabolism, such as CYP2E1, genes regulating immune response
against microbial infection are also our priority (14).
EBV is a ubiquitous herpes virus that infects more
than 90% of the human population and establishes a life-long viral
persistence in the host. EBV has been consistently identified as an
important risk factor for NPC, with a dose-response relationship
between EBV antibodies and NPC risk. The single clonally derived
viral genome can be found in all endemic NPC cells (21). Altered cell signaling is the
molecular basis for EBV infection-induced aberrant cell
proliferation. The EBV DNA may persist for its lifetime in an
episomal form in the host carrier cells. The virus-encoded protein
products or the virus-transcribed non-coding regulatory dsRNAs can
activate the transcription of otherwise silenced cellular genes,
which leads to the synthesis of enzymes capable of promoting viral
and cellular DNA replication. Thus, these proteins or/and
regulatory dsRNAs block apoptosis and drive host cells toward
division and immortalization. Particularly at later stages of
oncogenesis, the viral-encoded proteins and the viral-transcribed
non-coding regulatory dsRNAs, inducing false signaling and
activation of the proliferation pathways, bring the previously
infected cells into a full transformation burst (22).
CR2 may be a good candidate susceptibility genes for
NPC due to its function of regulating the immune response and
mediating EBV infection. Chronic EBV infection that may lead to
tumorigenesis in nasopharyngeal epithelium cells is mediated
through recognition of EBV stimuli by CR2 and pIgR receptors.
Improper regulation or compromised function of CR2 may contribute
to NPC. Our study represents the first comprehensive evaluation of
an association between sequence variants in the 5′-UTR of CR2 and
NPC. Variations in gene 5′-UTR can lead to altered gene expression
levels, and potential disequilibrium of the normal cellular
machinery. In the present study, we identified DNA variants in the
5′-UTR region of CR2 that may account for variability of CR2
transcription regulation among individuals with NPC, namely genetic
variations due to mutations in the CR2 5′-UTR. The variant 24C
conferred a nearly 2-fold increase in the risk for developing NPC
(OR=1.81; 95% CI, 1.21–2.70) in the investigated population.
Although the observed 2-fold increase in risk is modest, our
finding is intriguing as genes in multiple pathways alter the risk
for NPC, and each individual gene likely contributes only a modest
risk. This phenomenon is also observed in other complex diseases
(23). It is interesting that the
risk associated with the mutation 24C genotype was more pronounced
in younger (45 years or younger) individuals. These results suggest
that the 24C substitution in the 5′-UTR of exon 1 in CR2 may act as
a genetic susceptibility factor for NPC.
Initiation of translation is one of the most
important steps that may influence the level of gene expression,
and 5′-UTR sequences may greatly contribute to this step. In fact,
recent studies have shown that 5′-UTR plays an important role in
the regulation of gene expression (24) in a variety of organisms (microbes,
plants and animals). The 5′-UTR-mediated regulation has been
suggested to modulate gene expression through both stimulatory
(25) and inhibitory mechanisms
(26), including influencing RNA
transcription (27),
post-transcriptional modification of RNA (secondary structure and
mRNA stability) (28) and
alteration of translational efficiency (29). Indeed, a recent study demonstrated
that 5′-UTR plays an important role in post-transcriptional
modification and/or translation. The 5′-UTR has also been shown to
affect translation efficiency through a cap-independent internal
ribosome entry site (IRES)-mediated mechanism (30). Although this novel mechanism of the
translation initiation was first discovered in picornaviral RNAs,
increasing evidence indicates that certain eukaryotic mRNAs also
apply a similar mechanism. The IRES-mediated translation initiation
requires specific sequences that can form a Y-shaped secondary
structure and a short stem loop near the start of the AUG codon
(31). Approximately 6.4% of human
5′-UTR sequences contain IRES and these leader sequences are
usually >200 bp in length. However, it is currently unknown
whether shorter leader sequences (with <200 bp), such as the
hSP-A 5′-UTR variants under study, are involved in IRES-mediated
translational mechanism.
It has been shown that the activity of the CR2
promoter is induced in IM-9 B cells through the protein kinase A-
and protein kinase C-signaling pathways, and by anti-CD40 Ab and
IL-4 (32). The transcription
factors that were subsequently identified to play a role in this
induced expression include AP-1, CREB and an X box/E box-binding
protein. In addition, the protein kinase A and protein kinase
C-responsive heterogeneous nuclear ribonucleoprotein D (hnRNP
D)3 was found to specifically bind to a novel element in
the CR2 promoter. It was reported that truncation of promoter
sequences from −1252 to −57 did not have a significant effect on
promoter activity. Thus, the −57/+75 construct displayed activity
similar to that of the −1252/+75 full promoter. These findings
clearly suggest that critical positive promoter elements are
present in the −57/+75 region, which may contain potential
regulatory elements such as CREB/AP-1 half-site, X box and E box.
However, these findings do not exclude the possibility that
promoter elements upstream of −57 play a role in regulating CR2
promoter activity.
Since the SNP 24T>C is located in the −57/+75
region of 5′-UTR, we investigated its significance on gene
expression in the present study. The results of transient
transfection indicated that the 5′-UTR of CR2 differentially
influences the expression of the reporter gene in C666 cells. The
activity of PMUT and UMUT was significantly increased compared with
the WILD constructs, indicating that the sequences within the CR2
5′-UTR, and specifically those surrounding the 24 position,
regulate CR2 promoter activity and the variation in this region may
lead to increases in CR2 expression. The higher levels of CR2 may
be a risk of NPC susceptibility, since upregulated CR2 expression
may enhance the EBV infection through either B cells or epithelial
cells thus mediating the development and oncogenesis of NPC.
Nevertheless, we do not exclude the possibility that other
mechanisms may also contribute to the increase in CR2 levels during
NPC onset.
The SNP 24T>C was also reported as a susceptible
factor of systemic lupus erythematosus (9,10). By
constructing −315/+75 promoter region into the luciferase report
system and the transfection constructs into the Raji B
lymphoblastoid cell line, expression of the major 24 T allele
resulted in a 2-fold increase in transcriptional activity compared
with expression of the minor C allele. The differences between the
above study and ours may be due to the result of multiple factors,
including alterations in different cell lines and different cloned
regions of the promoter.
Acknowledgements
This study was supported by the National Natural
Science Foundation (30000141 and 81173616), the National Key Basic
Research Projects (973 Projects, G19980510) and the National
Science and Technology Project (2002BA711A03) of China. We thank
Lizhen Chen and Qisheng Feng for providing the clinical data and we
also thank Ruhua Zhang for the DNA sequencing.
References
|
1
|
Yu MC and Yuan JM: Epidemiology of
nasopharyngeal carcinoma. Semin Cancer Biol. 12:421–429. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chan TC, Teo ML and Johnson J:
Nasopharyngeal carcinoma. Ann Oncol. 13:1007–1015. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang XS, Wang HH, Hu LF, et al: V-val
subtype of Epstein-Barr virus nuclear antigen 1 preferentially
exists in biopsies of nasopharyngeal carcinoma. Cancer Lett.
211:11–18. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hu C, Wei W, Chen X, et al: A global view
of the oncogenic landscape in nasopharyngeal carcinoma: an
integrated analysis at the genetic and expression levels. PLoS One.
7:e410552012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fujisaku A, Harley JB, Frank MB, Gruner
BA, Frazier B and Holers VM: Genomic organization and polymorphisms
of the human C3d/Epstein-Barr virus receptor. J Biol Chem.
264:2118–2125. 1989.PubMed/NCBI
|
|
6
|
Hirunsatit R, Kongruttanachok N,
Shotelersuk K, et al: Polymeric immunoglobulin receptor
polymorphisms and risk of nasopharyngeal cancer. BMC Genet.
4:32003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zheng Y, Zhang W, Ye Q, et al: Inhibition
of Epstein-Barr virus infection by lactoferrin. J Innate Immun.
4:387–398. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shao X, He Z, Chen Z and Yao K: Expression
of an Epstein-Barr-virus receptor and Epstein-Barr-virus-dependent
transformation of human nasopharyngeal epithelial cells. Int J
Cancer. 71:750–755. 1997. View Article : Google Scholar
|
|
9
|
Wu H, Boackle SA, Hanvivadhanakul P, et
al: Association of a common complement receptor 2 haplotype with
increased risk of systemic lupus erythematosus. Proc Natl Acad Sci
USA. 104:3961–3966. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cruickshank MN, Karimi M, Mason RL, et al:
Transcriptional effects of a lupus-associated polymorphism in the
5′ untranslated region (UTR) of human complement receptor 2
(CR2/CD21). Mol Immunol. 52:165–173. 2012.PubMed/NCBI
|
|
11
|
Simon K, Yang X, Munger K, et al:
Variation in the Epstein-Barr virus receptor, CR2, and risk of
multiple sclerosis. Mult Scler. 13:947–948. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ghandri N, Gabbouj S, Farhat K, et al:
Association of HLA-G polymorphisms with nasopharyngeal carcinoma
risk and clinical outcome. Hum Immunol. 72:150–158. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen Y and Chan SH: Polymorphism of T-cell
receptor genes in nasopharyngeal carcinoma. Int J Cancer.
56:830–833. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jia WH, Pan QH, Qin HD, et al: A
case-control and a family-based association study revealing an
association between CYP2E1 polymorphisms and nasopharyngeal
carcinoma risk in Cantonese. Carcinogenesis. 30:2031–2036. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
He JF, Jia WH, Fan Q, et al: Genetic
polymorphisms of TLR3 are associated with nasopharyngeal carcinoma
risk in Cantonese population. BMC Cancer. 7:1942007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhou XX, Jia WH, Shen GP, et al: Sequence
variants in toll-like receptor 10 are associated with
nasopharyngeal carcinoma risk. Cancer Epidemiol Biomarkers Prev.
15:862–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cho EY, Hildesheim A, Chen CJ, et al:
Nasopharyngeal carcinoma and genetic polymorphisms of DNA repair
enzymes XRCC1 and hOGG1. Cancer Epidemiol Biomarkers Prev.
12:1100–1104. 2003.PubMed/NCBI
|
|
18
|
Wang G, Guo X and Floros J: Human SP-A
3′-UTR variants mediate differential gene expression in basal
levels and in response to dexamethasone. Am J Physiol Lung Cell Mol
Physiol. 284:L738–L748. 2003.
|
|
19
|
Sham PC and Curtis D: Monte Carlo tests
for associations between disease and alleles at highly polymorphic
loci. Ann Hum Genet. 59:97–105. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ng WT, Yau TK, Yung RW, et al: Screening
for family members of patients with nasopharyngeal carcinoma. Int J
Cancer. 113:998–1001. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cheung F, Pang SW, Hioe F, et al:
Nasopharyngeal carcinoma in situ: two cases of an emerging
diagnostic entity. Cancer. 83:1069–1073. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rajcani J and Kudelova M: Gamma
herpesviruses: pathogenesis of infection and cell signaling. Folia
Microbiol. 48:291–318. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lohmueller KE, Pearce CL, Pike M, et al:
Meta-analysis of genetic association studies supports a
contribution of common variants to susceptibility to common
disease. Nat Genet. 33:177–182. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang G, Guo X and Floros J: Differences in
the translation efficiency and mRNA stability mediated by 5′-UTR
splice variants of human SP-A1 and SP-A2 genes. Am J Physiol Lung
Cell Mol Physiol. 289:497–508. 2005.
|
|
25
|
Shalev A, Blair PJ, Hoffmann SC, et al: A
proinsulin gene splice variant with increased translation
efficiency is expressed in human pancreatic islets. Endocrinology.
143:2541–2547. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kos M, Denger S, Reid G, et al: Upstream
open reading frames regulate the translation of the multiple mRNA
variants of the estrogen receptor alpha. J Biol Chem.
277:37131–37138. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hua XJ, Van de Cotte B, Van Montagu M, et
al: The 5′untranslated region of the At-P5R gene is involved in
both transcriptional and post-transcriptional regulation. Plant J.
26:157–169. 2001.
|
|
28
|
Kebaara B, Nazarenus T, Taylor R, et al:
The Upf-dependent decay of wild-type PPR1 mRNA depends on its
5′-UTR and first 92 ORF nucleotides. Nucleic Acids Res.
31:3157–3165. 2003.PubMed/NCBI
|
|
29
|
Zou Z, Eibl C and Koop HU: The stem-loop
region of the tobacco psbA 5′UTR is an important determinant of
mRNA stability and translation efficiency. Mol Genet Genomics.
269:340–349. 2003.
|
|
30
|
Vagner S, Galy B and Pyronnet S:
Irresistible IRES. Attracting the translation machinery to internal
ribosome entry sites. EMBO Rep. 2:893–898. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Rubtsova MP, Sizova DV, Dmitriev SE, et
al: Distinctive properties of the 5′-untranslated region of human
hsp70 mRNA. J Biol Chem. 278:22350–22356. 2003.
|
|
32
|
Vereshchagina LA, Tolnay M and Tsokos GC:
Multiple transcription factors regulate the inducible expression of
the human complement receptor 2 promoter. J Immunol. 166:6156–6163.
2001. View Article : Google Scholar : PubMed/NCBI
|