Introduction
Plant-derived active constituents and their
semi-synthetic or synthetic analogs have served as one of the major
sources of anticancer drugs. An analysis of current
chemotherapeutic agents and their sources indicates that nearly 60%
of approved anticancer drugs are derived from natural products
(1). Ginseng, such as American
ginseng (Panax quinquefolius L.) and Asian ginseng (Panax
ginseng C.A. Meyer), is the root of different Panax
species (Araliaceae) and is one of the most commonly used
traditional medicines. The saponins of ginseng (also known as
ginsenosides) are its major active components and have been shown
to possess anti-inflammatory, antitumor, and neuroprotective
activities (2,3). Two types of ginsenosides in ginseng,
protopanaxatriol (PTS) and protopanaxadiol (PDS) (2,4) have
been shown to exert anticancer properties (5–9). After
oral administration of PDS ginsenosides (e.g., Rg3) to mice, PDS is
metabolically converted to protopanaxadiol (PPD) and Compound K
(CK) by intestinal bacteria (10,11).
Compound K can significantly inhibit the PMA-induced MMP-9
secretion and protein expression via suppressing the DNA-binding
and transcriptional activities of AP-1, which is the downstream
factor of p38 MAPK, ERK and JNK (12). Thus, it is of importance to
understand the anticancer effects and possible mechanisms
associated with ginseng derivatives.
We previously investigated the cancer
chemopreventive activities of American ginseng root extracts (AGE
and S-AGE), fractions (S2h) and pure ginsenoside Rg3 on human
colorectal cancer cells (13).
Ginsenoside Rg3 was shown to exert antiproliferative effects on
HCT116 cells in vitro and to inhibit tumor growth in a nude
mouse xenograft tumor model (14).
Furthermore, we conducted a microarray expression profiling
analysis and found that the expression levels of 76 genes, such as
A kinase (PRKA) anchor protein 8 (AKAP8L) and phosphatidylinositol
transfer protein α (PITPNA), were differentially regulated after
the treatment of HCT116 cells with S2h (American ginseng extract)
or ginsenoside Rg3 (13). As one of
the most important metabolites of the ginseng herb, PPD and its
derivates have therapeutic potential for inhibiting the growth and
invasiveness of tumors. However, the molecular mechanisms
underlying the anticancer activity of PPD remain to be fully
elucidated.
The present study investigated the anticancer
effects of PPD and its mode of action in human cancer cells. We
found that PPD inhibited growth and induced cell cycle arrest in
HCT116 cells. Furthermore, PPD inhibited the xenograft tumor growth
in athymic nude mice. The xenograft tumor size was significantly
reduced following treatment with PPD for up to 3 weeks.
Furthermore, PPD inhibited the expression of PITPNA while
upregulating AKAP8L expression in HCT116 cells. Pathway-specific
reporter assays indicated that PPD effectively inhibited the NF-κB,
JNK and MAPK/ERK signaling pathways. Thus, our results suggest that
PPD may exert its anticancer activity on colon cancer cells through
targeting major signaling pathways, such as NF-κB, JNK and
MAPK/ERK.
Materials and methods
Chemicals and drug preparations
PPD was kindly provided by Professor Ping Li of
China Pharmaceutical University (Nanjing, China) with a purity
>95% as confirmed by HPLC (4,15). PPD
was dissolved in dimethyl sulfoxide (DMSO) (15 mM stock solution).
For in vivo treatment, PPD was dissolved in PEG. Unless
otherwise indicated, all chemicals were obtained from Fisher
Scientific (Pittsburgh, PA, USA) or Sigma-Aldrich (St. Louis, MO,
USA).
Cell culture
Human colorectal cancer lines (HCT116 and SW480),
breast cancer cell lines (MDA-MB-468 and MDA-MB-231), prostate
cancer cell lines (PC3 and DU145), osteosarcoma cell lines (MG63
and 143B) and HEK-293 cells were purchased from the American Type
Culture Collection (ATCC, Manassas, VA, USA) and grown in
Dulbecco’s modified Eagle’s medium (DMEM) (Invitrogen Life
Technologies, Carlsbad, CA, USA) supplemented with 10% fetal bovine
serum (FBS; HyClone Laboratories, Logan, UT, USA) and 50 units
penicillin/streptomycin in 5% CO2 at 37°C.
MTT proliferation assay
A modified MTT assay was used to examine the cell
growth inhibitory effect of ginsenosides on cell proliferation as
previously described (16). Cells
were seeded in 96-well plates (1×104 cells/well, 50–70%
density). Ginsenosides were added to the cells at various
concentrations and incubation was carried out for 48 h. Fifteen
microliters of dye solution was added to each well and incubated
for an additional 4 h. One hundred microliters/well
solubilization/stop solution was added to stop the reaction and to
solubilize the formazan crystals in a humidified atmosphere
overnight. Absorbance at 570 nm was determined using a 96-well
microplate reader.
Crystal violet assay
HCT116 cells were treated with the indicated
concentrations of drugs. At the endpoints, the cell culture medium
was carefully removed. The wells were gently washed with
phosphate-buffered saline (PBS) at room temperature. The medium was
aspirated and cells were stained with 0.5% crystal violet formalin
solution at room temperature for 20–30 min. After staining, the
cells were washed with tap water and air-dried at room temperature
(17,18).
Flow cytometry and cell cycle
analysis
Flow cytometry was carried out to quantitatively
detect the cell cycle distribution (19). Cells were plated into 6-well plates
for drug treatments. At 24, 48 and 72 h post treatment, cells were
harvested, washed with PBS, fixed in cold methanol overnight at 4°C
and stained with 50 ng/ml propidium iodide (PI) by incubation at
4°C for 15 min. The stained cells were analyzed by flow
cytometry.
RNA isolation and semi-quantitative
reverse transcription-polymerase chain reaction (RT-PCR)
analysis
Total RNA was isolated using TRIzol reagents and
used to generate cDNA templates by RT reaction with hexamer and
SuperScript® II RT (both from Invitrogen Life
Technologies). The first strand cDNA products were further diluted
10-fold and used as PCR templates. Semi-quantitative RT-PCR was
carried out as described (20).
Briefly, PCR primers were designed using the Primer3 program to
amplify the human genes of interest (product sizes 150–180 bp) as
follows: GAPDH forward, 5′-CAACGAATTTGGCTACAGCA-3 and reverse,
5′-AGGGGAGATTCAGTGTGGTG-3′; PITPNA forward,
5′-CGTCCTACCCCCATGTTG-3′ and reverse, 5′-ACTGGGCAGCGTCTGTTC-3′; and
AKAP8L forward, 5′-GCAGGCAGGCAAGAAGAG-3′ and reverse,
5′-TGGCCATCTCGTCCTCAT-3′. A touchdown cycling program was carried
out as follows: 94°C for 2 min for 1 cycle, 92°C for 20 sec, 68°C
for 30 sec, and 72°C for 12 cycles with a decrease of 18°C per
cycle and then at 92°C for 20 sec, 57°C for 30 sec, and 72°C for 20
sec for 20 to 25 cycles depending on the abundance of a given gene.
The specificity of PCR products was confirmed by resolving PCR
products on 1.5% agarose gels. All samples were normalized with the
internal control GAPDH.
Xenograft tumor model and xenogen
bioluminescence imaging
The HCT116-Luc cell line, which stably expresses
firefly luciferase, was generated as previously described (19,21).
Animal use and care were carried out according to the protocol
guidelines approved by the Institutional Animal Care and Use
Committee. Athymic nude mice (female, 4–6 weeks old, ~20 g body
weight, n=5/group; Harlan SD, Indianapolis, IN, USA) were used.
HCT116-Luc cells were harvested and resuspended in PBS to a final
density of 2×107 cells/ml. Cells (1×106) were
injected subcutaneously into the flanks of the mice. At 1 week post
injection, PPD was administered (30 mg/kg) through an i.p.
injection once every 2 days for 3 week.
For whole body bioluminescence imaging, animals were
anesthetized with isoflurane attached to a nose-cone mask within
the Xenogen IVIS 200 imaging system after implantion for 1 week.
Mice were injected (i.p.) with D-Luciferin sodium salt (Gold
BioTechnology, St. Louis, MO, USA) at 100 mg/kg in 0.1 ml sterile
PBS. The pseudo-images were obtained by superimposing the emitted
light over the grayscale images of the animal. Quantitative
analysis was carried out with Xenogen’s Living Image V2.50.1
software as described (13,19). Animals were sacrificed after 3
weeks, and tumor samples were retrieved for histologic
examination.
Histologic evaluation and
immunohistochemical staining
Retrieved tumor tissues were fixed in 10% formalin
and embedded in paraffin. Serial sections of the embedded specimens
were stained with hematoxylin and eosin. Paraffin-embedded sections
were deparaffinized and then rehydrated in a graduated manner. The
deparaffinized samples were subjected to antigen retrieval and
fixation. Slides were blocked and probed with an antiproliferating
cell nuclear antigen (PCNA) antibody (Santa Cruz Biotechnology,
Inc., Santa Cruz, CA, USA), followed by incubation with the
anti-mouse IgG-biotin secondary antibody. Finally, sections were
incubated with HRP-streptavidin and visualized by
3,3′-diaminobenzidine staining (22).
Construction of pathway-specific Gaussia
luciferase reporters, establishment of HCT116-GLuc reporter lines,
and Gaussia luciferase assay
Promoters responsive to the following signaling
pathways, including MAPK/ERK, MAPK/JNK, Wnt, Notch, cell
cycle/pRb-E2F, NF-κB, Myc/Max, hypoxia (namely Elk-1/SRF, AP-1,
TCF/LEF, RBP-Jκ, E2F/DP1, NF-κB and hypoxia-inducible factor-1)
were cloned into our homemade pBGLuc retroviral vector. All
subcloned promoter fragments were verified by DNA sequencing.
Stable HCT116-GLuc reporter lines were established
using the retroviral transduction approach as previously described
(14,19,21).
Gaussia luciferase activity was determined using the Gaussia
luciferase assay kit (New England Biolabs). Briefly, HCT116-GLuc
cell lines were seeded in 24-well culture plates and treated with 0
or 10 μM PPD. After 24 h, cell culture medium was subjected to
Gaussia luciferase assay. Each assay condition was conducted in
triplicate.
Statistical analysis
The in vitro experiments were performed in
triplicate. Data are expressed as the means ± standard error (SE).
Statistical significances between vehicle treatment vs.
drug-treatment were determined by one-way ANOVA and the Student’s
t-test. A value of p<0.05 was considered to indicate a
statistically significant result.
Results
PPD inhibits the proliferative activity
of human cancer cells in vitro
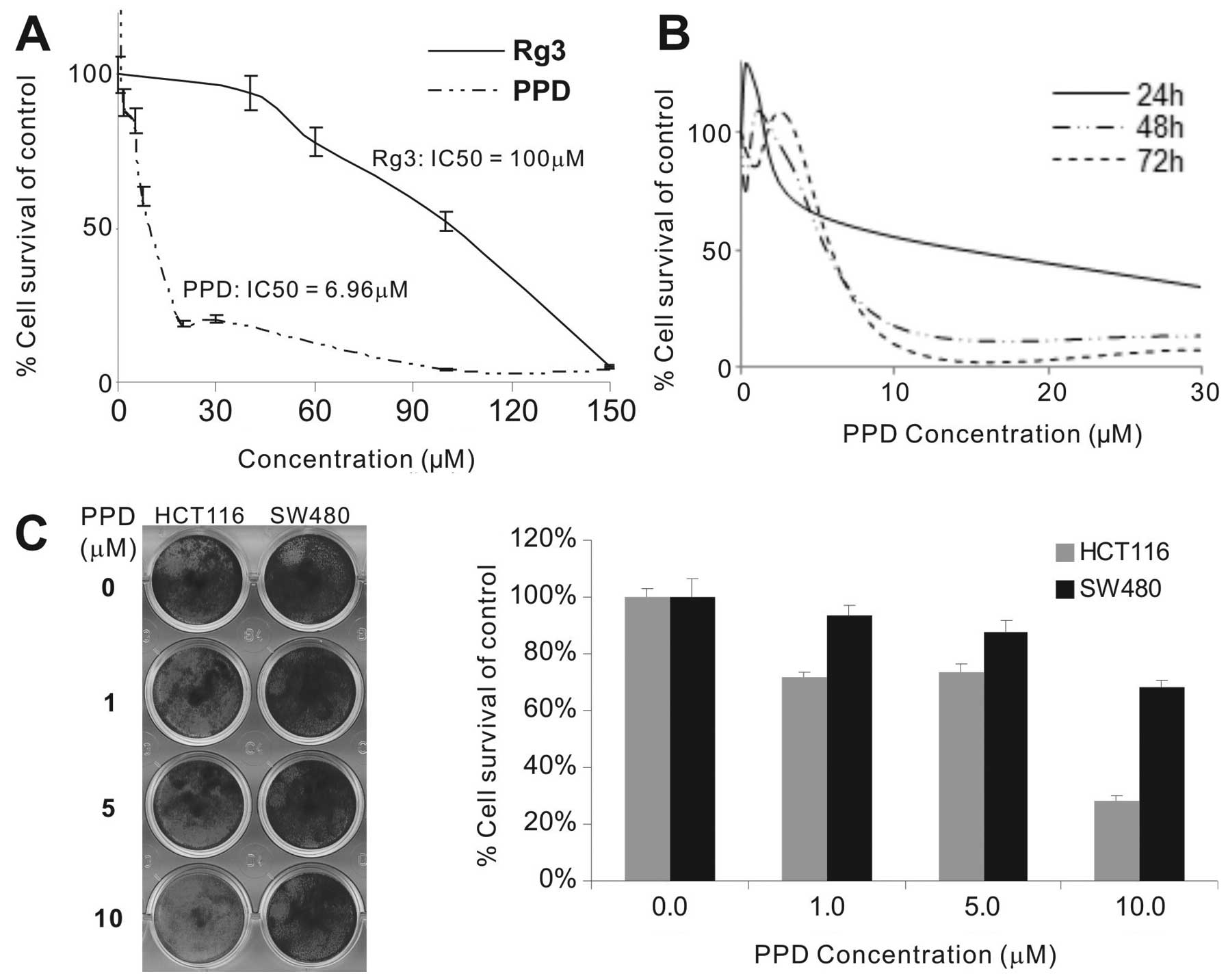
The effect of PPD and ginsenoside Rg3 on the
proliferation of HCT116 cells was evaluated by MTT assay. The
IC50 of PPD was significantly lower than that of Rg3 in
the HCT116 cells (Fig. 1A). At
10–30 μM, PPD exhibited strong anti-proliferation effects after 48
and 72 h of treatment (Fig. 1B). We
also investigated whether PPD inhibits the viability of other human
cancer cell lines. Treatment of different types of cancer cells
with different dosages of PPD for 48 h significantly suppressed the
cell proliferation of the tested cancer cell lines: human colon
cancer (HCT116 and SW480), breast cancer (MDA-MB-468 and
MDA-MB-231), prostate cancer (PC3 and DU145) and osteosarcoma cell
lines (MG63 and 143B) (data not shown). The IC50 value
for PPD in these cancer cell lines was 4.69 μM for HCT116, 8.99 μM
for SW480, 7.64 μM for MDA-MB-468, 4.49 μM for MDA-MB-231, 1.40 μM
for PC3, 4.71 μM for DU145, 5.17 μM for MG63, and 8.36 μM for 143B,
respectively. Crystal violet staining assay revealed that 10 μM PPD
had anti-proliferation effects on the HCT116 cells (cell viability
<30%) although PPD was less effective on the SW480 cells
(Fig. 1C), which was consistent
with the differences in their IC50 values.
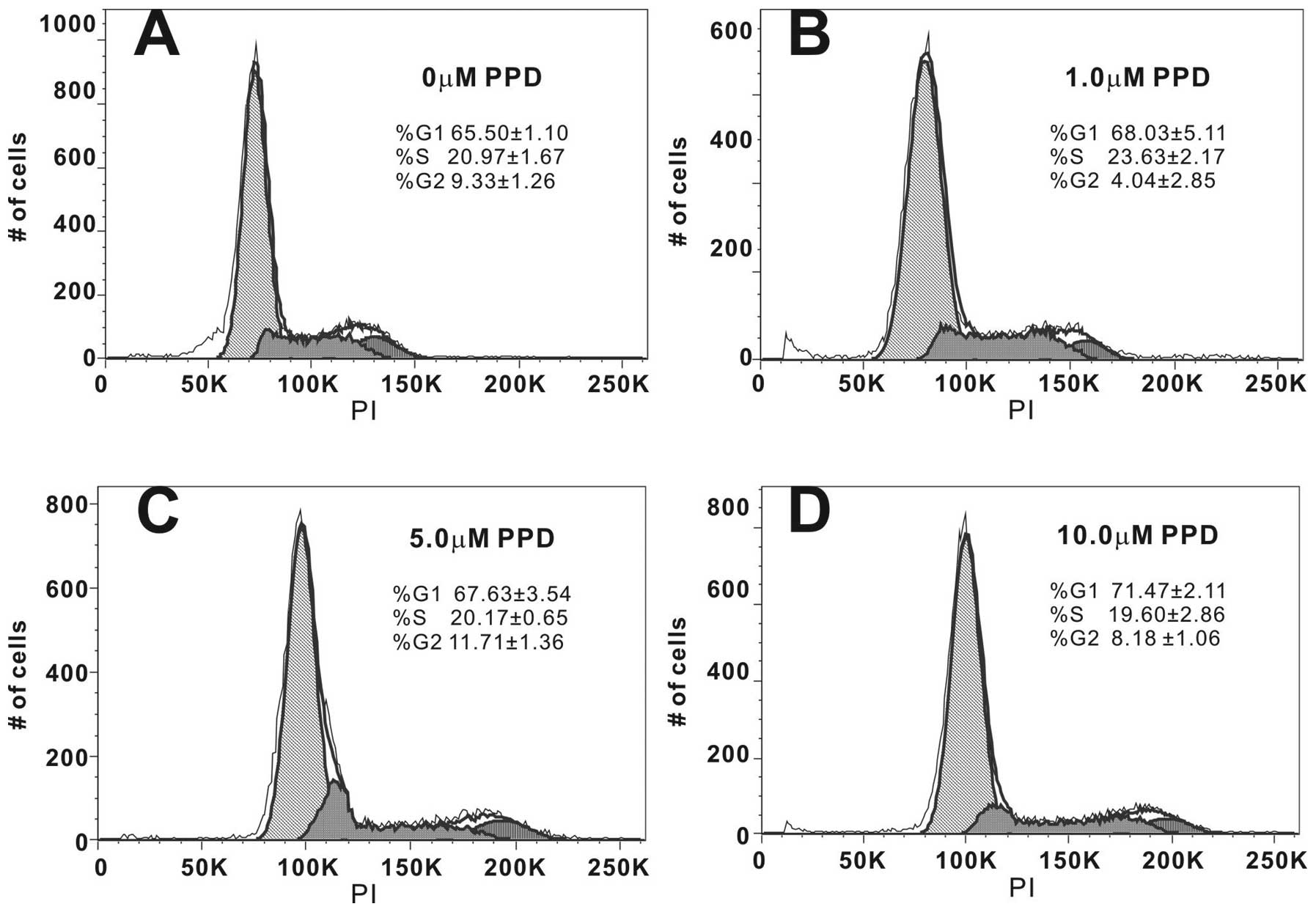
PPD restricts the proliferating cancer
cells in the G1/S phases of the cell cycle
In order to better understand the mechanism behind
PPD-mediated inhibition of cell proliferation, we analyzed the
distribution of PPD-treated cancer cells in different phases of the
cell cycle by flow cytometry following treatment of cells with
different concentrations of PPD for 48 h. We found that PPD caused
a dose-dependent cell accumulation in the G1/S phase (Fig. 2). For example, treatment of HCT116
cells with 10 μM PPD led to an 86 to 91% increase in cells in the
G1+S phase (Fig. 2A vs. D), leading
to fewer cells progressing to the G2 phase. These results indicate
that cell cycle progression was significantly blocked in the G1/S
phase when cells were treated with PPD.
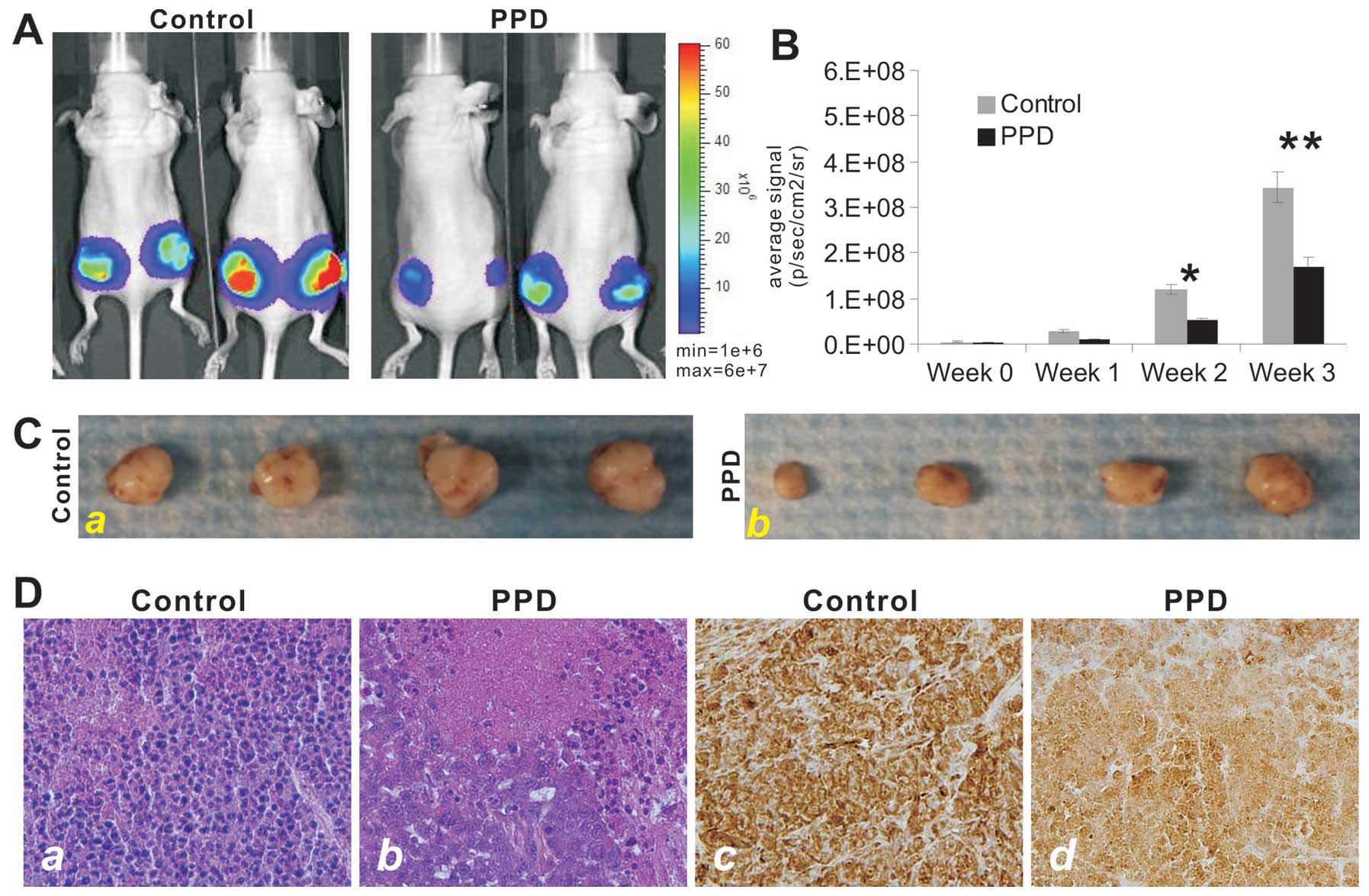
PPD effectively inhibits tumor growth in
vivo
We further tested the antitumor activity of PPD in a
xenograft tumor model of human colon cancer. The firefly
luciferase-tagged HCT116 cells were subcutaneously injected in mice
to form xenograft tumors. At one week, the tumor-bearing athymic
nude mice were i.p. administered PPD at 30 mg/kg once every 2 days
for up to 3 weeks. The tumor growth was monitored by using whole
body Xenogen imaging (Fig. 3A). PPD
was shown to significantly inhibit xenograft tumor growth at 2
weeks after treatment (Fig. 3B). At
the endpoint (3 months after treatment), the xenograft tumors were
retrieved and were shown to be smaller in the PPD treatment group
(Fig. 3C). Histologically, the
PPD-treated xenograft tumors exhibited significant necrosis
(Fig. 3D-a vs. -b).
Immunohistochemical staining with a PCNA antibody revealed that
xenograft tumor cells treated with PPD exhibited a marked decrease
in cell proliferation (Fig. 3D-a vs.
-b). Thus, the in vivo results suggest that PPD may be
developed into an efficacious anticancer agent.
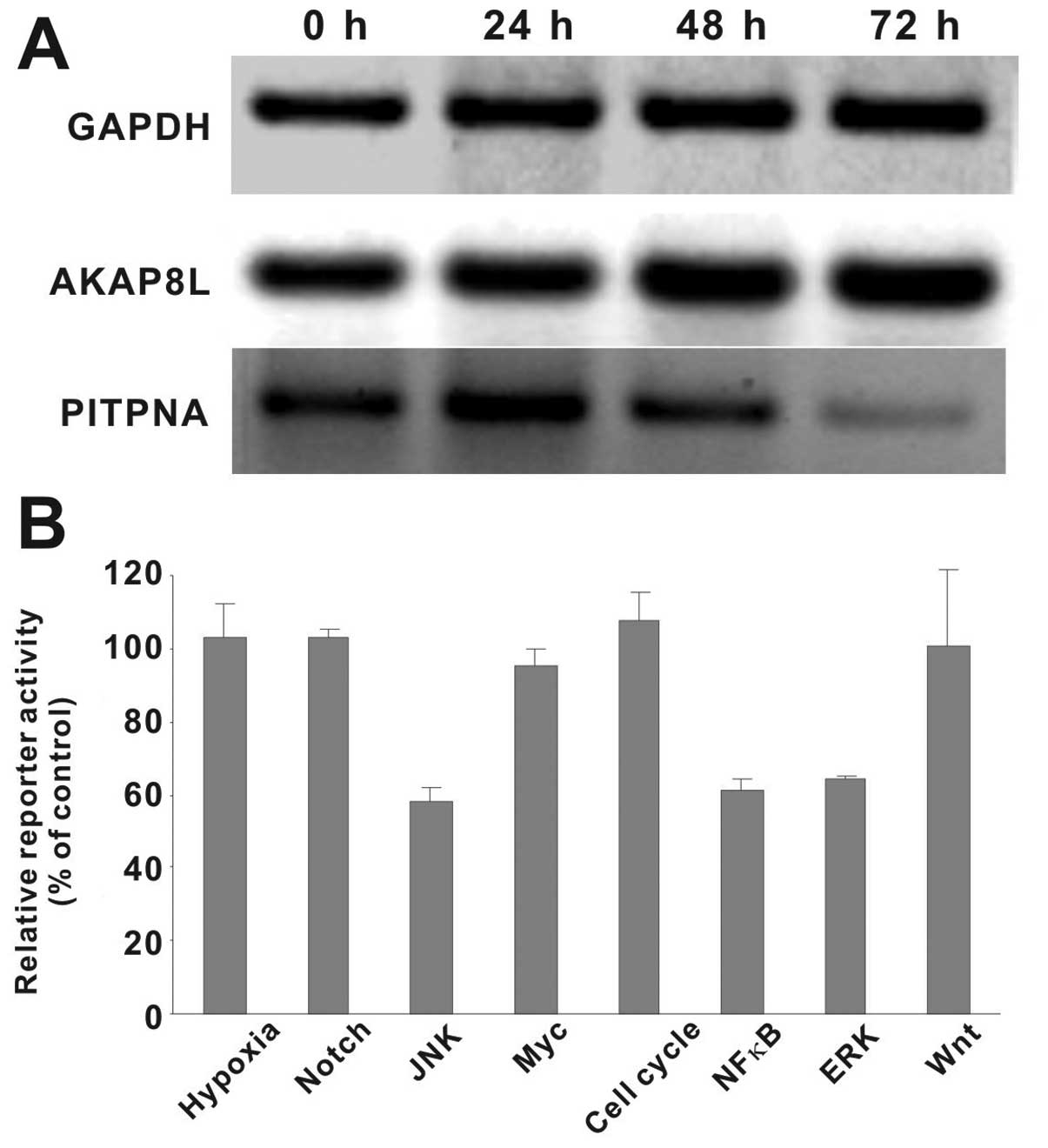
Rg3 targets AKAP8L and PITPNA may be
involved in the antitumor effect of PPD
We previously found that the expression levels of
AKAP8L and PITPNA were significantly altered following treatment
with S2h (American ginseng extract) or ginsenoside Rg3 in HCT116
cells (13). As one of the main
metabolites of S2h and the aglycon of ginsenoside Rg3, PPD was
shown to affect the expression levels of AKAP8L and PITPNA in the
PPD-treated HCT116 cells (Fig. 4A).
Specifically, PPD treatment slightly upregulated AKAP8L expression
while significantly inhibiting PITPNA expression in a
time-dependent fashion. These results suggest that upregulation of
AKAP8L and/or downregulation of PITPNA may play an important role
in mediating the anticancer activities conferred by ginsenoside
derivatives, such as PPD. However, the exact mechanism behind their
roles in the anticancer action of PPD needs to be thoroughly
investigated.
PPD targets MAPK and NF-κB signaling
pathways in human colon cancer
We sought to further investigate the mechanistic
basis underlying the anticancer activity of PPD. Although the above
results indicate that Rg3 targets AKAP8L and PITPNA may play an
important role in the mode of anticancer action of PPD, it is known
that cancer development usually hijacks multiple cellular signaling
pathways (23,24). Thus, we tested the effect of PPD on
8 major cancer-related signaling pathways, including MAPK/ERK,
MAPK/JNK, Wnt, Notch, cell cycle/pRb-E2F, NF-κB, Myc/Max and
hypoxia. When HCT116 cells containing the pathway-speciifc
reporters were treated with 10 μM PPD, we found that the relative
reporter activities for NF-κB, MAPK/JNK and MAPK/ERK pathways were
significantly inihibited (Fig. 4B).
The other 5 pathways, noticeably Myc/Max and Wnt, were not
significantly affected by PPD in HCT116 cells. Thus, these results
suggest that PPD may exert its anticancer activity at least in part
by targeting the ERK, JNK and/or NF-κB signaling pathways although
the exact mechanism needs to be fully elucidated.
Discussion
In the present study, we demonstrated the
effectiveness of PPD, a metabolite of ginseng saponin, against
multiple tumor types in human cancer cell culture and animal
models. PPD inhibited human cancer cell growth in 8 types of human
cancer cells and these results were consistent with other reports
on the effects of PPD on human cancer cell lines (25–29).
Previous studies have shown that PPD induced cell cycle arrest in
the G0–G1 phase in human hepatocellular carcinoma SMMC7721 cells
(30) or in the G1 phase in U937
human monocytic leukemia cells (31). Similar variability was observed in
the HCT116 colon cancer cells.
Ginseng is one of the most widely used medicinal
plants and remains a top selling natural product globally. The
major bioactive constituents in ginseng are ginsenosides, a group
of triterpene glycosides (2,4).
Several natural ginseng saponins have been shown to exhibit high
potency against multiple tumor types in cell culture and animal
models (32). Kim et
al(33) studied 11 ginsenosides
and determined that Rg3 and Rh2 inhibited the proliferation of
prostate cancer cells. Iishi et al(34) used a rat AOM-induced tumor model to
determine the effects of Rg3 in inhibiting the cell proliferation
of colon cancer cells. Jia et al(35) reported that Rh2 inhibited
proliferation, induced apoptosis in cancer cell lines, and
sensitized drug-resistant breast cancer cells to paclitaxel. Rk1
and Rg5 were also found to inhibit the proliferation of human
hepatocellular carcinoma cells (36,37).
Notably, Compound K
[(20-O-(β-D-glucopyranosyl)-20(S)-protopanaxadiol)], one of
the most important intestinal metabolites isolated from ginseng PDS
saponins, can also induce apoptotic cell death concurrent with cell
cycle arrest in the G0–G1 phase in SMMC7721 cells (30) and G1 phase arrest in U937 cells
(31). Compound K was also found to
inhibit the cell viability and induce apoptosis of human gastric
carcinoma cells via the Bid-mediated mitochondrial pathway
(38). Moreover, Compound K
significantly inhibited PMA-induced MMP-9 secretion and protein
expression by suppressing DNA-binding and transcriptional
activities of AP-1, which is the downstream factor of p38 MAPK, ERK
and JNK (12). Similar to many
other herbal medicines, ginseng is usually taken orally. In this
form its bioavailability is low due to incomplete absorption
(39). To date, the
biotransformation of ginsenosides to their metabolites by
intestinal bacteria has been reported. Some of the metabolites,
such as Compound K and PPD, have shown various bioactivities
including cancer chemoprevention (15). Nonetheless, the anticancer
mechanisms of these ginseng metabolites are largely unknown.
The low in vivo toxicity of PPD suggests that
this compound or its derivatives may have potential for clinical
applications in cancer chemotherapy (2). However, PPD is a highly hydrophobic
molecule with limited water solubility and has low in vivo
uptake. In our in vivo studies, we used PEG and PEG400 to
improve the solubility of PPD. Our results clearly demonstrated
that this formulation could circumvent the limited solubility
and/or bioavailability of PPD.
The glycoside of PPD was shown to induce apoptosis
of human prostate cancer cells via inhibition of the NF-κB
(40,41), JNK and ERK pathways (12). PPD may induce apoptosis by
decreasing caspase-3 activity (25,42),
MMP secretion (26) or the ER
stress pathway (27). However, it
remains unclear how PPD targets cancer-related signaling pathways.
In the present study, we used a cell-based, unbiased,
pathway-specific analysis and identified three major pathways that
may be targeted by PPD. Thus, future investigation should be
directed towards how these pathways are inhibited. These lines of
investigations are critical for the potential clinical use of PPD
as an anticancer agent.
In conclusion, we demonstrated that PPD effectively
inhibits cell proliferation and tumor growth of human cancer both
in vitro and in vivo. We demonstrated that the
anticancer activity of PPD in colon cancer cells is at least in
part due to the downregulation of multiple signaling pathways,
noticeably MAPK/ERK, MAPK/JNK and NF-κB. Although further
investigations are required to dissect the underlying mechanisms,
these results illustrate the potential clinical applications for
PPD alone or in combination with other anticancer agents.
Acknowledgements
The reported study was supported in part by research
grants from the NIH (P01 AT004418 to C.-Z.W., W.D., C.-S.Y. and
T.-C.H.; and AT005362 to C.-Z.W.) and the Natural Science
Foundation of China (NSFC grant no. 81102852 to J.-L.G. and grant
no. 81228024 to J.-L.G. and T.-C.H.).
References
|
1
|
Cragg GM, Newman DJ and Snader KM: Natural
products in drug discovery and development. J Nat Prod. 60:52–60.
1997. View Article : Google Scholar
|
|
2
|
Wang CZ and Yuan CS: Potential role of
ginseng in the treatment of colorectal cancer. Am J Chin Med.
36:1019–1028. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang CZ, Aung HH, Zhang B, Sun S, Li XL,
He H, Xie JT, He TC, Du W and Yuan CS: Chemopreventive effects of
heat-processed Panax quinquefolius root on human breast
cancer cells. Anticancer Res. 28:2545–2551. 2008.PubMed/NCBI
|
|
4
|
Qi LW, Wang CZ and Yuan CS: Isolation and
analysis of ginseng: advances and challenges. Nat Prod Rep.
28:467–495. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cho SH, Chung KS, Choi JH, Kim DH and Lee
KT: Compound K, a metabolite of ginseng saponin, induces apoptosis
via caspase-8-dependent pathway in HL-60 human leukemia cells. BMC
Cancer. 9:4492009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
King MM and Murphy LL: Role of cyclin
inhibitor protein p21 in the inhibition of HCT116 human colon
cancer cell proliferation by American ginseng (Panax
quinquefolius) and its constituents. Phytomedicine. 17:261–268.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yoo JH, Kwon HC, Kim YJ, Park JH and Yang
HO: KG-135, enriched with selected ginsenosides, inhibits the
proliferation of human prostate cancer cells in culture and
inhibits xenograft growth in athymic mice. Cancer Lett. 289:99–110.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Du GJ, Wang CZ, Zhang ZY, Wen XD, Somogyi
J, Calway T, He TC, Du W and Yuan CS: Caspase-mediated
pro-apoptotic interaction of panaxadiol and irinotecan in human
colorectal cancer cells. J Pharm Pharmacol. 64:727–734. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Du GJ, Wang CZ, Qi LW, Zhang ZY, Calway T,
He TC, Du W and Yuan CS: The synergistic apoptotic interaction of
panaxadiol and epigallocatechin gallate in human colorectal cancer
cells. Phytother Res. 27:272–277. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hasegawa H, Sung JH, Matsumiya S and
Uchiyama M: Main ginseng saponin metabolites formed by intestinal
bacteria. Planta Med. 62:453–457. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xie JT, Wang CZ, Zhang B, Mehendale SR, Li
XL, Sun S, Han AH, Du W, He TC and Yuan CS: In vitro and in vivo
anticancer effects of American ginseng berry: exploring
representative compounds. Biol Pharm Bull. 32:1552–1558. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jung SH, Woo MS, Kim SY, Kim WK, Hyun JW,
Kim EJ, Kim DH and Kim HS: Ginseng saponin metabolite suppresses
phorbol ester-induced matrix metalloproteinase-9 expression through
inhibition of activator protein-1 and mitogen-activated protein
kinase signaling pathways in human astroglioma cells. Int J Cancer.
118:490–497. 2006. View Article : Google Scholar
|
|
13
|
Luo X, Wang CZ, Chen J, Song WX, Luo J,
Tang N, He BC, Kang Q, Wang Y, Du W, He TC and Yuan CS:
Characterization of gene expression regulated by American ginseng
and ginsenoside Rg3 in human colorectal cancer cells. Int J Oncol.
32:975–983. 2008.PubMed/NCBI
|
|
14
|
He BC, Gao JL, Luo X, Luo J, Shen J, Wang
L, Zhou Q, Wang YT, Luu HH, Haydon RC, Wang CZ, Du W, Yuan CS, He
TC and Zhang BQ: Ginsenoside Rg3 inhibits colorectal tumor growth
through the down-regulation of Wnt/β-catenin signaling. Int J
Oncol. 38:437–445. 2011.PubMed/NCBI
|
|
15
|
Wang HY, Qi LW, Wang CZ and Li P:
Bioactivity enhancement of herbal supplements by intestinal
microbiota focusing on ginsenosides. Am J Chin Med. 39:1103–1115.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gao JL, He TC, Li YB and Wang YT: A
traditional Chinese medicine formulation consisting of Rhizoma
corydalis and Rhizoma curcumae exerts synergistic
anti-tumor activity. Oncol Rep. 22:1077–1083. 2009.PubMed/NCBI
|
|
17
|
He BC, Chen L, Zuo GW, Zhang W, Bi Y,
Huang J, Wang Y, Jiang W, Luo Q, Shi Q, Zhang BQ, Liu B, Lei X, Luo
J, Luo X, Wagner ER, Kim SH, He CJ, Hu Y, Shen J, Zhou Q, Rastegar
F, Deng ZL, Luu HH, He TC and Haydon RC: Synergistic antitumor
effect of the activated PPARgamma and retinoid receptors on human
osteosarcoma. Clin Cancer Res. 16:2235–2245. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
He BC, Gao JL, Zhang BQ, Luo Q, Shi Q, Kim
SH, Huang E, Gao Y, Yang K, Wagner ER, Wang L, Tang N, Luo J, Liu
X, Li M, Bi Y, Shen J, Luther G, Hu N, Zhou Q, Luu HH, Haydon RC,
Zhao Y and He TC: Tetrandrine inhibits Wnt/β-catenin signaling and
suppresses tumor growth of human colorectal cancer. Mol Pharmacol.
79:211–219. 2011.
|
|
19
|
Su Y, Wagner ER, Luo Q, Huang J, Chen L,
He BC, Zuo GW, Shi Q, Zhang BQ, Zhu G, Bi Y, Luo J, Luo X, Kim SH,
Shen J, Rastegar F, Huang E, Gao Y, Gao JL, Yang K, Wietholt C, Li
M, Qin J, Haydon RC, He TC and Luu HH: Insulin-like growth factor
binding protein 5 suppresses tumor growth and metastasis of human
osteosarcoma. Oncogene. 30:3907–3917. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen L, Jiang W, Huang J, He BC, Zuo GW,
Zhang W, Luo Q, Shi Q, Zhang BQ, Wagner ER, Luo J, Tang M, Wietholt
C, Luo X, Bi Y, Su Y, Liu B, Kim SH, He CJ, Hu Y, Shen J, Rastegar
F, Huang E, Gao Y, Gao JL, Zhou JZ, Reid RR, Luu HH, Haydon RC, He
TC and Deng ZL: Insulin-like growth factor 2 (IGF-2) potentiates
BMP-9-induced osteogenic differentiation and bone formation. J Bone
Miner Res. 25:2447–2459. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Su Y, Luo X, He BC, Wang Y, Chen L, Zuo
GW, Liu B, Bi Y, Huang J, Zhu GH, He Y, Kang Q, Luo J, Shen J, Chen
J, Jin X, Haydon RC, He TC and Luu HH: Establishment and
characterization of a new highly metastatic human osteosarcoma cell
line. Clin Exp Metastasis. 26:599–610. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Luu HH, Zhou L, Haydon RC, Deyrup AT,
Montag AG, Huo D, Heck R, Heizmann CW, Peabody TD, Simon MA and He
TC: Increased expression of S100A6 is associated with decreased
metastasis and inhibition of cell migration and anchorage
independent growth in human osteosarcoma. Cancer Lett. 229:135–148.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Vogelstein B and Kinzler KW: Cancer genes
and the pathways they control. Nat Med. 10:789–799. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: the next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Park EJ, Zhao YZ, Kim J and Sohn DH: A
ginsenoside metabolite,
20-O-beta-D-glucopyranosyl-20(S)-protopanaxadiol, triggers
apoptosis in activated rat hepatic stellate cells via caspase-3
activation. Planta Med. 72:1250–1253. 2006.PubMed/NCBI
|
|
26
|
Li G and Wang Z, Sun Y, Liu K and Wang Z:
Ginsenoside 20(S)-protopanaxadiol inhibits the proliferation and
invasion of human fibrosarcoma HT1080 cells. Basic Clin Pharmacol
Toxicol. 98:588–592. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhu GY, Li YW, Tse AK, Hau DK, Leung CH,
Yu ZL and Fong WF: 20(S)-Protopanaxadiol, a metabolite of
ginsenosides, induced cell apoptosis through endoplasmic reticulum
stress in human hepatocarcinoma HepG2 cells. Eur J Pharmacol.
668:88–98. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lee JY, Shin JW, Chun KS, Park KK, Chung
WY, Bang YJ, Sung JH and Surh YJ: Antitumor promotional effects of
a novel intestinal bacterial metabolite (IH-901) derived from the
protopanaxadiol-type ginsenosides in mouse skin. Carcinogenesis.
26:359–367. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li N, Huang Y, Xiao W, Liu J, Li X and Li
P: Synthesis and cytotoxic activities of new fatty acid esters of
20(S)-protopanaxadiol. Z Naturforsch C. 66:199–204. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ming YL, Song G, Chen LH, Zheng ZZ, Chen
ZY, Ouyang GL and Tong QX: Anti-proliferation and apoptosis induced
by a novel intestinal metabolite of ginseng saponin in human
hepatocellular carcinoma cells. Cell Biol Int. 31:1265–1273. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kang KA, Kim YW, Kim SU, Chae S, Koh YS,
Kim HS, Choo MK, Kim DH and Hyun JW: G1 phase arrest of the cell
cycle by a ginseng metabolite, compound K, in U937 human monocytic
leukemia cells. Arch Pharm Res. 28:685–690. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang W, Zhao Y, Rayburn ER, Hill DL, Wang
H and Zhang R: In vitro anti-cancer activity and structure-activity
relationships of natural products isolated from fruits of Panax
ginseng. Cancer Chemother Pharmacol. 59:589–601. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kim HS, Lee EH, Ko SR, Choi KJ, Park JH
and Im DS: Effects of ginsenosides Rg3 and Rh2 on the proliferation
of prostate cancer cells. Arch Pharm Res. 27:429–435. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Iishi H, Tatsuta M, Baba M, Uehara H,
Nakaizumi A, Shinkai K, Akedo H, Funai H, Ishiguro S and Kitagawa
I: Inhibition by ginsenoside Rg3 of bombesin-enhanced peritoneal
metastasis of intestinal adenocarcinomas induced by azoxymethane in
Wistar rats. Clin Exp Metastasis. 15:603–611. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jia WW, Bu X, Philips D, Yan H, Liu G,
Chen X, Bush JA and Li G: Rh2, a compound extracted from ginseng,
hypersensitizes multidrug-resistant tumor cells to chemotherapy.
Can J Physiol Pharmacol. 82:431–437. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kim YJ, Kwon HC, Ko H, Park JH, Kim HY,
Yoo JH and Yang HO: Anti-tumor activity of the ginsenoside Rk1 in
human hepatocellular carcinoma cells through inhibition of
telomerase activity and induction of apoptosis. Biol Pharm Bull.
31:826–830. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lee KY, Lee YH, Kim SI, Park JH and Lee
SK: Ginsenoside-Rg5 suppresses cyclin E-dependent protein kinase
activity via up-regulating p21Cip/WAF1and
down-regulating cyclin E in SK-HEP-1 cells. Anticancer Res.
17:1067–1072. 1997.PubMed/NCBI
|
|
38
|
Hu C, Song G, Zhang B, Liu Z, Chen R,
Zhang H and Hu T: Intestinal metabolite compound K of panaxoside
inhibits the growth of gastric carcinoma by augmenting apoptosis
via Bid-mediated mitochondrial pathway. J Cell Mol Med. 16:96–106.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Qi LW, Wang CZ and Yuan CS: American
ginseng: potential structure-function relationship in cancer
chemoprevention. Biochem Pharmacol. 80:947–954. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kim SM, Lee SY, Cho JS, Son SM, Choi SS,
Yun YP, Yoo HS, Yoon do Y, Oh KW, Han SB and Hong JT: Combination
of ginsenoside Rg3 with docetaxel enhances the susceptibility of
prostate cancer cells via inhibition of NF-kappaB. Eur J Pharmacol.
631:1–9. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Choo MK, Sakurai H, Kim DH and Saiki I: A
ginseng saponin metabolite suppresses tumor necrosis
factor-α-promoted metastasis by suppressing nuclear factor-κB
signaling in murine colon cancer cells. Oncol Rep. 19:595–600.
2008.PubMed/NCBI
|
|
42
|
Popovich DG and Kitts DD: Mechanistic
studies on protopanaxadiol, Rh2, and ginseng (Panax
quinquefolius) extract induced cytotoxicity in intestinal
Caco-2 cells. J Biochem Mol Toxicol. 18:143–149. 2004. View Article : Google Scholar : PubMed/NCBI
|