Introduction
Cell-cell adhesion is mediated by members of the
cadherin-catenin system and among them E-cadherin and β-catenin are
important adhesion molecules involved in the viability and function
of epithelial cells as well as tissue integrity (1–10).
β-catenin, α-catenin and γ-catenin are proteins that bind to the
highly conserved intracellular cytoplasmatic tail of E-cadherin
(1,10). β-catenin, a 92-kDa protein, has been
found to be associated with the cytoplasmic portion of E-cadherin
and this association is critical for cell adhesion (4). Differential expression of β-catenin
has been reported in human cancers (7). Loss of E-cadherin-β-catenin adhesion
is an important step in the progression of many epithelial
malignancies. The function of the cadherin-catenin system in cell
adhesion as well as in intracellular signaling appears to be
regulated by multiple factors and by different molecular mechanisms
(1,3,4,9).
E-cadherin belongs to the family of cell adhesion molecules that
are Ca2+-dependent (5,6).
β-catenin regulates the function of cadherin in cell-to-cell
adhesion which is critical for the maintenance of tissue structure
and morphogenesis. The intracellular domain of E-cadherin interacts
with a variety of cytoplasmic proteins such as β-catenin, α-catenin
and α-actinin.
Interaction of α-actinin with the cadherin/catenin
cell-cell adhesion complex has been observed via α-catenin
(4). α-catenin, a 102-kDa protein,
was initially described as an E-cadherin-associated molecule
(10), but it has been shown to be
associated with other members of the cadherin family, such as
N-cadherin and P-cadherin (1,5,6,8).
It has crucial functions in the E-cadherin-mediated cell-cell
adhesion system and also as a downstream signaling molecule in the
Wnt pathway (5). γ-catenin, an
82-kDa protein, also known as the fourth armadillo repeat of
plakoglobin, is associated with high affinity binding to the
cytoplasmic domains of E-cadherin and desmosomal cadherin DSg2. It
also binds with α-catenin and N-catenin (2).
Integrins are not considered to be bona fide target
molecules for oncogenes or tumor suppressors, yet their expression
levels appear to be altered by transformation in breast cancer
cells (11,12). No characteristic integrin expression
pattern can be ascribed to all breast tumors and it is likely that
several subtypes of breast cancer may generate tumors with a
distinct expression pattern of integrins (13). Alteration of integrin expression in
the breast can be regarded as a marker of pre-malignant origin
(13). The markers used to
characterize cell lines include integrin receptors, which are cell
adhesion molecules that primarily mediate cell-matrix interactions,
being localized to focal contacts, or in the case of the α-6 β-4
integrin heterodimer to hemidesmosomes (14). High expression level of α-6 integrin
in human breast carcinomas correlates with poor prognosis (14). The prognostic value of α-6 β-4
integrin expression in breast carcinomas is influenced by laminin
production from tumor cells (11).
Cell-cell adhesion molecules including E-cadherin
are identified in adherent junctions whereas desmocollin (DSc)
glycoproteins are localized in desmosomes. Biological markers used
to differentiate between cell phenotypes also revealed components
of desmosomes such as desmocollin 1–3 (DSc). These cell adhesion
molecules are transmembrane proteins of the cadherin family that
form the adhesive core of desmosomes. Desmosomes are sites of
adhesion between adjacent cells in layers of epithelia, as well as
in certain non-epithelial tissues, and play an important role in
the maintenance of tissue architecture. DSc3 which is an important
glycoprotein and active member of the cadherin superfamily of
calcium-dependent cell-cell adhesion molecules and a principle
component of desmosomes plays a pivotal role in maintaining tissue
architecture; and therefore, loss of this component leads to a lack
of adhesion and a gain in cellular motility. DSc3 expression is
usually downregulated in breast cancer cell lines and in primary
breast tumors.
In vitro model systems have been extensively
used to gain insights into the molecular events of cancer
initiation and promotion and to identify novel
prognostic/diagnostic markers for various types of cancer. The
human breast epithelial cell line MCF-10F, spontaneously
immortalized and derived from the breast tissue of a 36-year-old
female, has the morphological characteristics of normal breast
epithelial cells (15–18). The MCF-10F cell line has been used
to detect sensitivity to both chemical carcinogens such as 7, 12,
dimethylbenz(a)anthracene (DMBA) and benzo(a)pyrene (BP) (15) and environmental carcinogens such as
ionizing radiation (16). It was
previously shown that estrogen was a prerequisite for the process
of high linear energy transfer (LET) radiation-induced
carcinogenesis (16). Several
phenotypic properties such as growth rate, anchorage-independent
growth and invasive characteristics have also been reported to be
grossly similar during the transformation process induced by
chemical carcinogens (19–25) and environmental factors, e.g.,
ionizing radiation (16). The
chemo-invasion or the ability of transformed cells to infiltrate
the basement membrane in vitro was correlated well with the
in vivo malignant characteristics. A Previous study
demonstrated that cell adhesion molecules are highly altered in
malignantly transformed cells relative to non-tumorigenic cell
lines indicating that their altered expression may support or
promote breast carcinogenic events (25). Results from our laboratory also
found that a combination of estrogen and the organophosphorous
compound parathion increased the expression of certain adhesion
molecules such as CD146 and β-catenin and the combined treatment
was found capable of altering cell proliferation and inducing
transformation of the MCF-10F cell line. To understand the
importance of cell adhesion molecules in carcinogenic events, we
aimed to examine their expression at different stages of our breast
cancer model system developed by the combined treatment of high LET
radiation and estrogen.
Materials and methods
Cell lines
MCF-10F cells were grown in DMEM/F-12 (1:1) medium
supplemented with antibiotics [100 U/ml penicillin, 100 μg/ml
streptomycin, 2.5 μg/ml amphotericin B (all from Life Technologies,
Grand Island, NY, USA)] and 10 μg/ml and 5% equine serum
(Biofluids, Rockville, MD, USA), 0.5 μg/ml hydrocortisone (Sigma,
St. Louis, MO, USA) and 0.02 μg/ml epidermal growth factor
(Collaborative Research, Bedford, MA, USA) were added (15–25).
An in vitro experimental breast cancer model (Alpha model)
developed by exposure of the immortalized human breast epithelial
cell line, MCF-10F, to low doses of high LET α particle radiation
(150 keV/μm) and subsequent growth in the presence or absence of
17β-estradiol was used in this study. This model consisted of human
breast epithelial cells at different stages of transformation: i) a
control cell line (MCF-10F); ii) MCF-l0F continually treated with
estradiol at 10−8 M (E or Estrogen) (Sigma-Aldrich)
named Estrogen cell line; iii) a malignant cell line (Alpha3); and
iv) a malignant and tumorigenic cell line (Alpha5) and the Tumor2
cell line derived from cells originating from a tumor after
injection of Alpha5 cells in nude mice.
Protein expression by
immunocytochemistry
Exponentially growing cells were plated on a glass
chamber slide (Nunc Inc., Naperville, IL, USA) as previously
described (16) at a density of
1×104 cells/ml of growth medium. Three independent
biological experiments were performed. The following primary
antibodies were used for detecting the protein expression:
β-catenin (E-5) (Sc-7963), α-catenin E (G-11) (Sc-9988) and
γ-catenin (H-1) (Sc-5415); E-cadherin (N-20) (Sc-1500) and integrin
β3 (N-20) (Sc-6627) (all from Santa Cruz Biotechnology, Santa Cruz,
CA, USA). Rhodamine conjugated secondary antibody was from Jackson
ImmunoResearch Laboratories (West Grove, PA, USA). All the
antibodies were used at a 1:500 dilution from the original stock
concentrations. Slides were mounted with coverslips using
Vectashield mounting medium (Vector Laboratories, Burlingame, CA,
USA). The cells were examined using a Zeiss Axiovert 100 TV
microscope (Carl Zeiss, Thornwood, NY, USA) with a 40× 11.3 NA
objective lens equipped with a laser scanning confocal attachment
(LSM 410 Carl Zeiss). The staining intensity of cells was
quantified as previously described (16,21).
Composite fluorescence images were generated and collected by
excitation at 488 nm using an argon/krypton laser (488 nm) as
previously described (16). A
semi-quantitative estimation based on the relative staining
intensity of protein expression was determined for the parental,
non-tumorigenic and tumorigenic cell lines. The number of
immunoreactive cells (30 cells/field) was counted in 5 randomly
selected microscopy fields per sample. Standard errors of the mean
are shown in the representative figures. Statistical analysis was
carried out with the F-test (randomized block) and comparisons
between groups with the Bonferroni-t-test with significance at a
P-value of <0.05.
For evaluation of protein expression by
immunoperoxidase staining, exponentially growing cell lines were
plated on a glass chamber slide (Nunc Inc.), at a density of
1×104 cells/ml of medium and allowed to grow for 2–3
days until 70% confluence (21).
The cells were fixed with buffered paraformaldehyde at room
temperature, incubated with 1% H2O2 in
methanol to block endogenous peroxidase and again washed twice with
buffer solution. Subsequently, cell cultures were then covered with
normal horse serum for 30 min at room temperature and incubated
with the anti-rabbit monoclonal antibody (Santa Cruz Biotechnology)
at a 1:500 dilution overnight at 4°C and then incubated for 45 min
with diluted biotinylated secondary antibody solution (Vector
Laboratories) and Vectastin Elite ABC reagent (Vector Laboratories)
was used. The experiments were repeated thrice in cells with
identical passages in vitro. The number of immunoreactive
cells (50 cells/fields) was counted in several randomly selected
microscopic fields (x400) per sample; 10 fields were counted for
each cell line.
Fluorescence-labeled probe preparation
for the microarray analysis
The poly(A) mRNA from normal, radiation- and
estrogen-treated breast cell lines were isolated using
QIA-Direct-mRNA Isolation kit (Qiagen, Valencia, CA, USA).
Fluorescence-labeled cDNA was prepared from 1 μg of each of these
poly(A) mRNAs by using oligo dT-primed polymerization and
Superscript II reverse transcriptase kit (Life Technologies) in the
presence of either Cy3- or Cy5-labeled dCTP following the standard
procedure as described (http://cmgm.stanford.edu/pbrown/protocols.html). The
appropriate Cy3- and Cy5-labeled probes were pooled and hybridized
to the microarray on glass coverslips for 16 h at 65°C and then
washed with high stringency for analysis.
Affymetrix HG-U133A Plus 2.0 GeneChip
microarray gene expression analysis
The breast cancer model (Alpha model) consisting of:
i) MCF-10F, ii) Estrogen, (iii) Alpha3, iv) Alpha5 and v) Tumor2
cell lines was used to analyze gene expression by the Affymetrix
U133A oligonucleotide microarray (Affymetrix, Santa Clara, CA, USA)
which contains 14,500 genes. Arrays were quantitatively analyzed
for gene expression using the Affymetrix GeneChip Operating
software (GCOS) with a dual global scaling option in the Genes@Work
software platform of the discovery algorithm SPLASH (structural
pattern localization analysis by sequential histograms) with a
false discovery rate of 0.05 (26).
Results
Previous morphological research (17) indicated that the parental MCF-10 and
Estrogen cell lines did not exhibit any of the features that
characterize malignant cells (anchorage-independent growth in soft
agar, invasion and tumor growth in nude mice). In contrast to
MCF-10F, the Alpha3 cell line formed colonies in soft agar and
invaded but failed to form tumors in immunosuppressed mice.
However, the Alpha5 cell line induced mammary gland tumors in
animals after cell injection. The cell line derived from such
tumors was named Tumor2. In the present study, all of these
malignant and non-malignant cell lines were used to analyze the
expression of adhesion molecules in breast carcinogenesis.
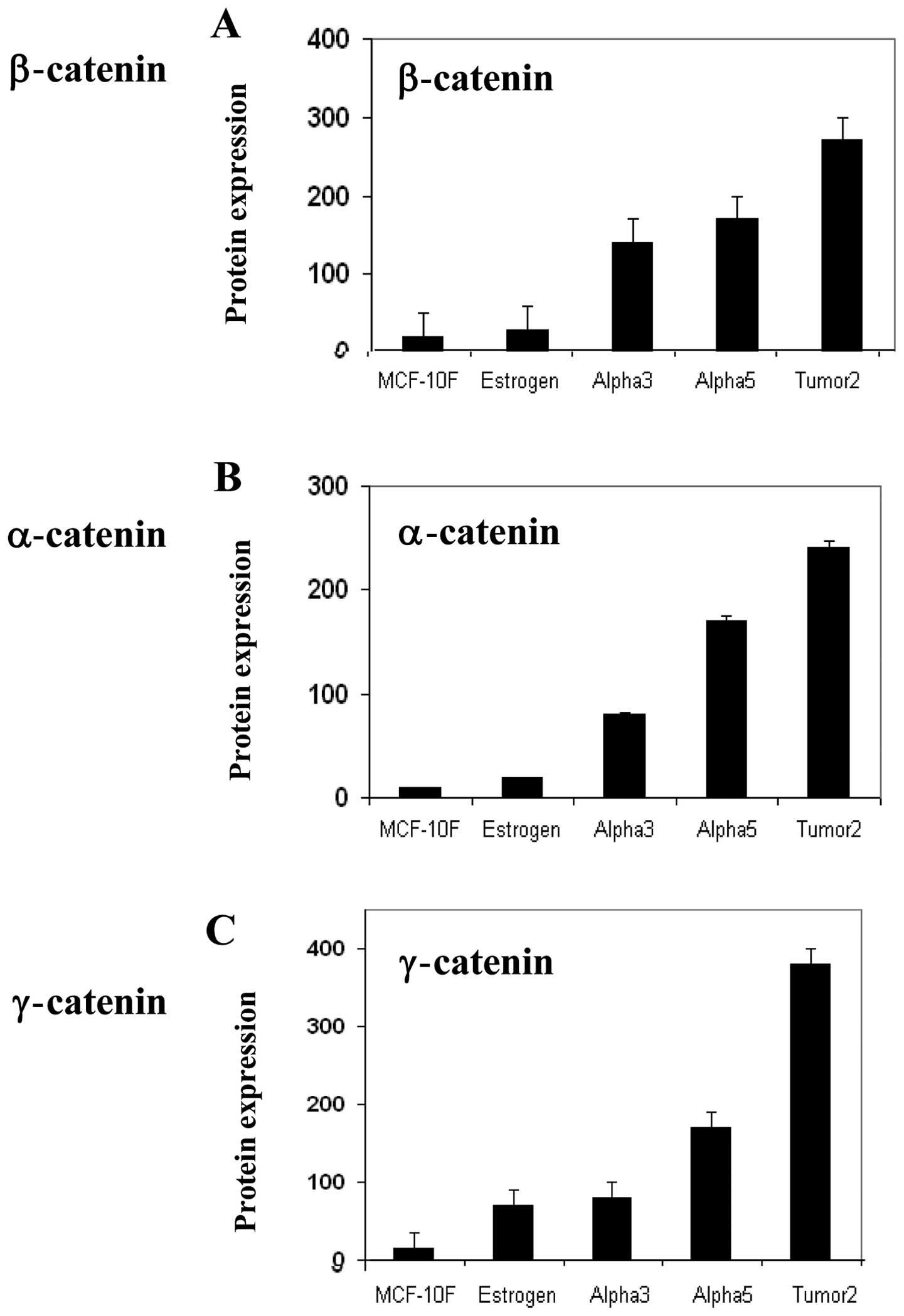
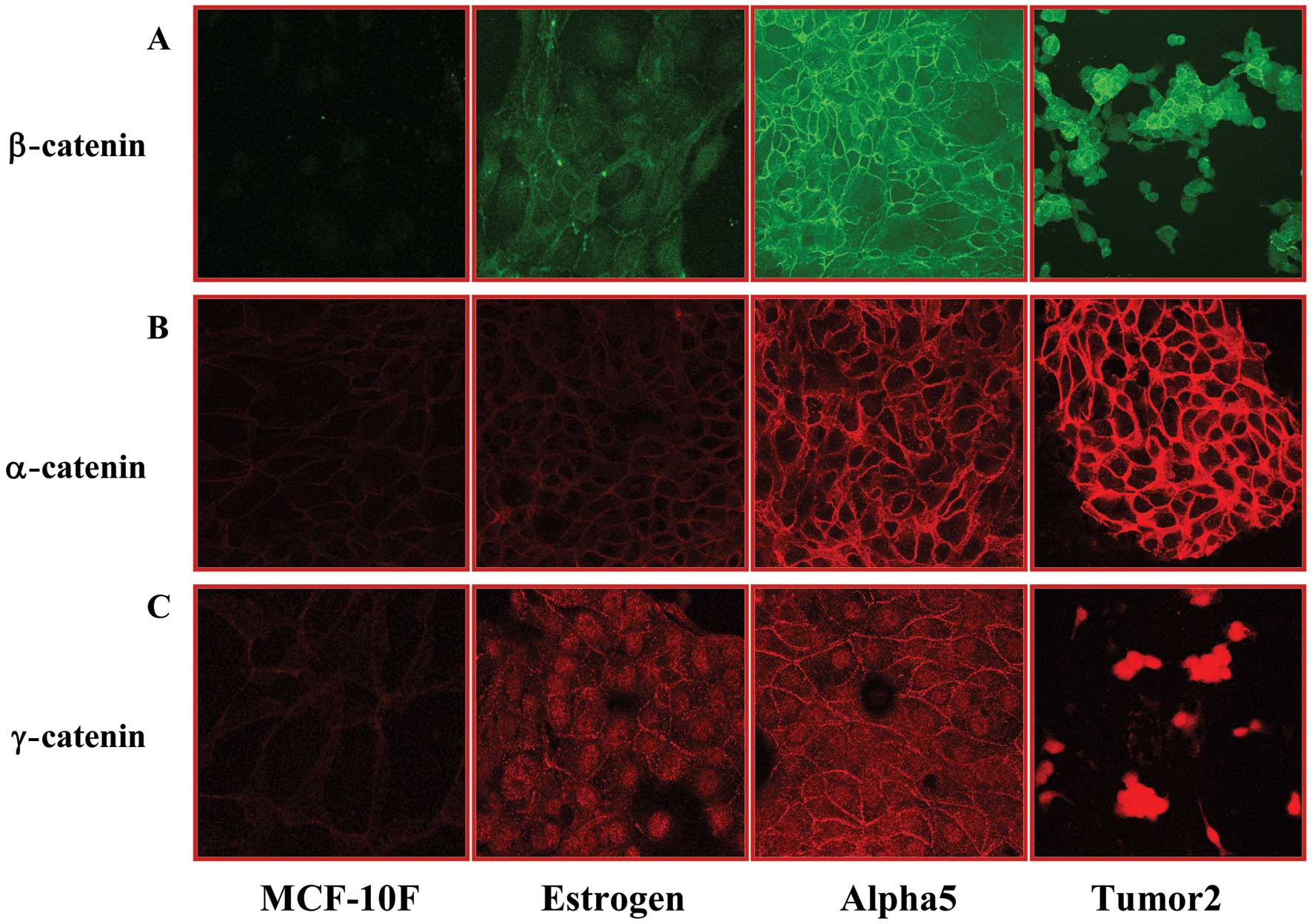
The immunofluorescence data obtained concerning the
relative expression of different adhesion molecules in the MCF-10F,
Estrogen, Alpha3, Alpha5 and Tumor2 cell lines are shown in
Fig. 1. α-catenin, β-catenin and
γ-catenin expression was significantly (P<0.05) higher in the
Alpha5 and Tumor2 cell lines when compared to the levels in the
MCF-10F, Estrogen and Alpha3 cell lines. Representative images of
immunoperoxidase and fluorescence staining are presented in
Fig. 2. Fig. 3A and C show the average values of
the levels of E-cadherin and integrin β-6 protein expression in the
MCF-10F, Estrogen, Alpha3, Alpha5 and Tumor2 cell lines as
determined by immunoperoxidase staining Fig. 3B and D. Such expression was
significantly (P<0.05) higher in the Alpha5 and Tumor2 cell
lines than that in the MCF-10F, Estrogen and Alpha3 cell lines.
However, the Tumor2 cell line had reduced expression levels of
E-cadherin and integrin β-6 protein when compared to that in the
Alpha5 cell line, but was higher than that in the Alpha3, Estrogen
and MCF-10F cell lines. The levels of these proteins were similar
in the parental MCF-10F and Estrogen cell lines.
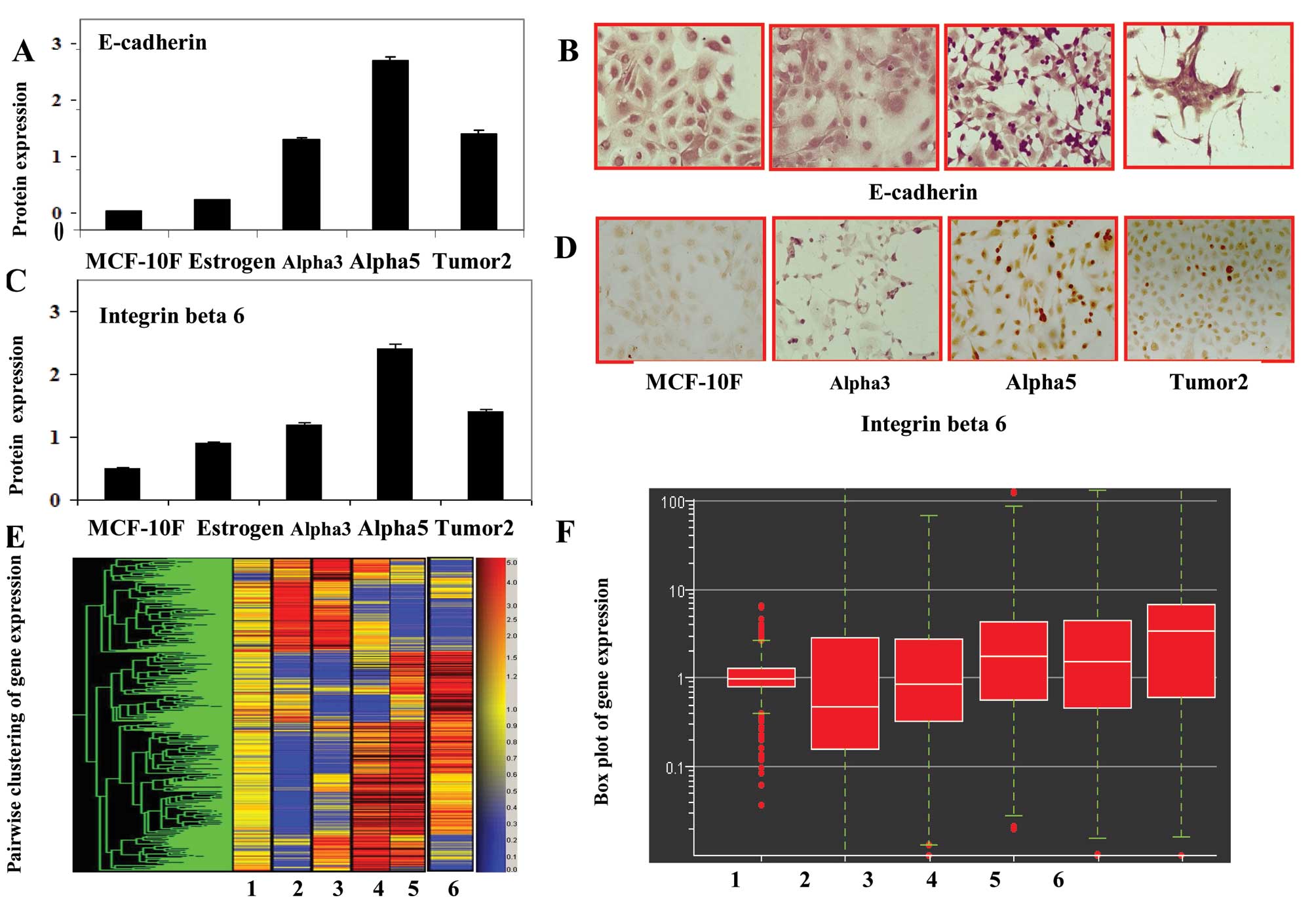 | Figure 3Bars in the histograms represent the
average (A) E-cadherin and (C) integrin β 6 protein expression in
the MCF-10F; Estrogen, Alpha3, Alpha5 and Tumor2 cell lines as
determined by peroxidase staining. Representative images of (B)
E-cadherin and (D) integrin β 6 protein expression in the MCF-10F,
Alpha3, Alpha5 and Tumor2 cell lines as stained by peroxidase
techniques. The primary antibodies used were mouse monoclonal
antibody. (E) Cluster-dendrogram of gene expression and (F)
fold-change gene expression indicated by a scatter plot of the
following pairwise comparative studies of the cell lines: 1,
MCF-10F/E; 2, MCF-10F/Alpha3; 3, E/Alpha5; 4, Alpha3/Alpha5; 5,
Alpha 3/Tumor2 and 6, Alpha5/Tumor2. |
Genes that were found to be differentially expressed
between the cell lines of the established Alpha model were studied.
Cluster-dendrogram and fold changes in gene expression of the cell
line model are shown in Fig. 3E and
F. Histogram plots showed differential expression of
E-cadherin, integrin β-6 and DSc3 genes in the cell lines as
detected by gene chip array. Results of the pairwise comparison of
the cell lines examined for the expression of the above-mentioned
genes are shown in Table I. The
following pairs of cell lines were analyzed: MCF-10F/Estrogen (E)
(Fig. 3F); MCF-10F/Alpha3;
Estrogen/Alpha5; Alpha3/Alpha5; Alpha5/Tumor2; and Alpha 3/Tumor2.
Results indicated that the pair-wise comparison did not reveal any
alteration in E-cadherin expression between MCF-10F/Estrogen (E)
cell lines whereas there was almost a 19- and 7-fold alteration in
MCF-10F/Alpha3 and MCF-10F/Alpha5 combinations. Similarly, between
Alpha3/Alpha5, there was a 6-fold change in E-cadherin gene
expression. Comparison of Alpha3/Tumor2 and Alpha5/Tumor2 cell
lines revealed 14- and 3-fold changes in expression, respectively.
Comparison between MCF-10F/Alpha3, Alpha3/Alpha5, and Alpha5/Tumor2
showed ~4-, 5- and 3-fold changes, respectively in integrin β-6
gene expression. However, no significant changes in expression were
observed in other pairwise combinations. Finally, pairwise
comparison of gene expression between MCF-10F/Alpha3, Alpha3/Alpha5
and Alpha5/Tumor2 showed changes of 3- to 4-fold whereas comparison
of the Alpha3/Tumor2 cell lines revealed a 13-fold change in
expression for the DSc3 gene. MCF-10F/Estrogen (E) and
MCF-10F/Alpha5 combinations showed no significant alterations in
gene expression.
 | Table IFold-change and pair-wise analysis of
differentially expressed genes a breast cancer model. |
Table I
Fold-change and pair-wise analysis of
differentially expressed genes a breast cancer model.
| Genes |
|---|
|
|
|---|
| E-cadherin | Integrin β-6 | DSc3 |
|---|
|
|
|
|
|---|
| Cell lines | Fold-change | Regulation | Fold-change | Regulation | Fold-change | Regulation |
|---|
| MCF-10F/Estrogen | 1.3 | ↑ | −1.0 | ↓ | −1.4 | ↓ |
| MCF-10F/Alpha3 | −19.9 | ↓ | 3.6 | ↑ | −3.2 | ↓ |
| Estrogen/Alpha5 | −7.3 | ↓ | −1.2 | ↓ | 1.6 | ↑ |
| Alpha3/Alpha5 | 5.6 | ↑ | −4.7 | ↓ | 3.6 | ↑ |
| Alpha3/Tumor2 | 2.6 | ↑ | 3.1 | ↑ | 3.6 | ↑ |
| Alpha5/Tumor2 | 14.3 | ↑ | −1.5 | ↓ | 13.1 | ↑ |
Discussion
Our previous study indicated that the combined
treatment of ionizing radiation and estrogen yielded different
stages in a malignantly transformed breast cancer cell model
system, which we termed the Alpha model system (17). Utilizing this model system, altered
expression of different cell adhesion molecules was detected in the
parental, non-tumorigenic and malignantly transformed cell lines
originally derived from the parental MCF-10A cell line. Results of
this study indicated that some of the cell adhesion molecules may
have prognostic/diagnostic significance for breast
carcinogenesis.
Expression levels of α-catenin, β-catenin and
γ-catenin were significantly greater in the Alpha5 and Tumor2 cell
lines when compared to these levels in the MCF-10F, Estrogen and
Alpha3 cell lines. β-catenin appears to be a critical component of
a complex signal transduction pathway that regulates the central
process of cellular proliferation and differentiation. The
intracellular β-catenin level was found to be regulated by its
association with the adenomatous polyposis coli tumor-suppressor
protein and GSK-3-β (10).
Consistent with our study, a previous study also showed an
increased level of β-catenin in human bronchial epithelial cells
transformed by treatment with the tobacco-specific nitrosamine,
4-(methylnitrosamino)-1-(3-pyridyl)-1-butatone (27). Other studies reported that catenins,
particularly when expressed in the cytoplasm with the E-cadherin
complex, are sensitive prognostic markers for invasive breast
cancer (3,7) since the E-cadherin/α-catenin complex
is capable of modulating cell-cell and cell-matrix adhesive
properties (3,7,11). In
support of this finding, the E-cadherin/α-catenin complex has also
been shown to modulate cell-cell and cell-matrix adhesive
properties of invasive colon carcinoma cells (3). The E-cadherin-catenin complex is also
the target of many growth factors and hormone-dependent signaling
pathways that regulate its function and expression (24). E-cadherin expression correlates with
poor survival in breast carcinoma even though it is strongly
expressed in both luminal and myoepithelial cells (28). Adherent junctions and desmosomes are
characteristic of epithelial cells. In the present study, we found
that E-cadherin protein expression was higher in the malignantly
transformed Alpha5 cell line than that in the parental MCF-10F cell
line. However, reduced E-cadherin expression was found in the
xenograft derived Tumor2 cell line when compared to that in Alpha5
although the precise reason for this is not clear. Both E-cadherin
and integrin β-6 proteins were overexpressed in Alpha5 when
compared to the protein levels in the MCF-10F, Estrogen and Alpha3
cell lines as determined by immunoperoxidase staining. Integrin β-6
and the DSc3 genes were expressed at higher levels in both the
Alpha3 and Alpha5 cell lines. Gene expression analysis identified
several of the adhesion molecules that were differentially
expressed in carcinogenesis. Reduced E-cadherin expression similar
to that observed in the Tumor 2 cell line probably indicates the
loss of the epithelial phenotype. Reduced E-cadherin is common in
many breast carcinomas and it is frequently lost in infiltrating
lobular carcinomas which otherwise clearly exhibit an epithelial
phenotype (29).
In previous studies, cell adhesion was also analyzed
by β-catenin protein expression and was also found higher in cells
treated with parathion alone and estrogen combined with parathion
in comparison to control and estrogen-treated cells. The function
of the cadherin-catenin system in cell adhesion as well as
intracellular signaling appears to be subjected to multifactorial
control by a variety of different mechanisms. β-catenin had a
similar reaction in the presence of parathion alone and combined
with estrogen in comparison to the control and in the presence of
estrogen. However, it seems that estrogens did not play a role in
this pesticide-induced model mediated by the cadherin-catenin
complex since both substances had equal effect. This complex seems
to initiate signaling events implicated in differentiation and
growth control. Other studies have indicated that the
E-cadherin-catenin complex is the target of many growth factors and
hormone-dependent signaling pathways which regulate its function
and expression (25,30). Other authors have found that
catenin, particularly when it is expressed in the cytoplasm, seems
to be a very sensitive prognostic marker with the E-cadherin
complex in invasive breast cancer (1,3,9,10).
It may have invasive capabilities since a possible role of the
E-cadherin/α-catenin complex in modulating cell-cell and
cell-matrix adhesive properties of invasive colon carcinoma cells
has been reported (25). There is
evidence that changes in the shape of the epithelial surface are
features of the cell transformation of epithelial cells. Our
results suggest that aberrant expression of β-catenin may be
involved in tumor metastasis.
Several genes involved in adhesion function such as
E cadherin, integrin β-6, and Dsc3 were found through gene
expression microarray studies. E-cadherin was not significantly
different when the MCF-10F and Estrogen cell lines were compared.
Differential gene expression has been reported in the literature
between luminal and myoepithelial cells when DSc2 and DSc3 were
compared with the DSc3 gene expression solely found in
myoepithelial population (30).
Thus, the expression profile of genes, particularly in conjunction
with other markers, can help to distinguish between luminal and
myoepithelial cells. In the present study, there was a decrease in
DSc3 gene expression. Integrin signaling is a well known
requirement for the complex process of metastasis which seems to
occur through a series of steps that involve local tissue invasion,
intravasation and survival in colonization and circulation
(14,30). The α-6 β-4 integrin complex is
strongly expressed in myoepithelial cells but not in luminal
epithelium. Expression of desmosomal cadherins is largely confined
to epithelial cells. Integrin β-6 protein expression was
significantly greater in Alpha5 than in the MCF-10F and estrogen
cell lines while the Tumor2 cell line exhibited a decreased protein
expression when compared with Alpha5. The α-6 β-4 integrin complex
is strongly expressed by myoepithelial cells but not in luminal
epithelium (13,30).
Results of this study suggest that environmental
agents (e.g., ionizing radiation) in the presence of estrogen can
drastically affect human breast cell adhesion phenomena thereby
promoting or supporting the molecular events of the process of
cellular transformation. Future studies are required to verify
whether the altered expression of cell adhesion molecules precedes
or accompanies the cellular transformation process. Elucidation of
the precise role of cell adhesion molecules in carcinogenic events
may be helpful to assess their prognostic/diagnostic significance
for breast carcinogenesis in clinical settings.
Acknowledgements
The support provided by FONDECYT #1120006 (G.M.C.)
and MINEDUC Universidad de Tarapacá (G.M.C.) is greatly
appreciated. We also thank Dr Manikandan Jayapal and Dd Praksah
Hande of the National University of Singapore for the analysis of
the Affymetrix microarray data.
References
|
1
|
Pierceall WE, Woodard AS, Morrow JS, Rimm
D and Fearon ER: Frequent alterations in E-cadherin and alpha- and
beta-catenin expression in human breast cancer cell lines.
Oncogene. 11:1319–1326. 1995.PubMed/NCBI
|
|
2
|
Ozawa M, Terada H and Pedraza C: The
fourth armadillo repeat of plakoglobin (gamma-catenin) is required
for its high affinity binding to the cytoplasmic domains of
E-cadherin and desmosomal cadherin Dsg2, and the tumor suppressor
APC protein. J Biochem. 118:1077–1082. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Breen E, Steele G Jr and Mercurio AM: Role
of the E-cadherin/alpha-catenin complex in modulating cell-cell and
cell-matrix adhesive properties of invasive colon carcinoma cells.
Ann Surg Oncol. 2:378–385. 1995. View Article : Google Scholar
|
|
4
|
Knudsen KA, Soler AP, Johnson KR and
Wheelock MJ: Interaction of α-actinin with the cadherin/catenin
cell-cell adhesion complex via α-catenin. J Cell Biol. 130:67–77.
1995.
|
|
5
|
Sacco PA, McGranahan TM, Wheelock MJ and
Johnson KR: Identification of plakoglobin domains required for
association with N-cadherin and α-catenin. J Biol Chem.
25:20201–20206. 1995.PubMed/NCBI
|
|
6
|
Wahl JK, Sacco PA, McGranahan-Sadler TM,
Sauppé LM, Wheelock MJ and Johnson KR: Plakoglobin domains that
define its association with the desmosomal cadherins and the
classical cadherins: identification of unique and shared domains.
Cell Sci. 109:1143–1154. 1996.PubMed/NCBI
|
|
7
|
Takayama T, Shiozaki H, Shibamoto S, Oka
H, Kimura Y, Tamura S, Inoue M, Monden T, Ito F and Monden M:
Beta-catenin expression in human cancers. Am J Pathol. 148:39–46.
1996.
|
|
8
|
Salomon D, Sacco PA, Roy SG, Simcha I,
Johnson KR, Wheelock MJ and Ben-Ze'ev A: Regulation of beta-catenin
levels and localization by over-expression of plakoglobin and
inhibition of the ubiquitin-proteasome system. J Cell Biol.
139:1325–1335. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lim SC and Lee MS: Significance of
E-cadherin/β-catenin complex and cyclin D1 in breast cancer. Oncol
Rep. 9:915–928. 2002.
|
|
10
|
Nakopoulou L, Gakiopoulou-Givalou H,
Karayiannakis AJ, Giannopoulou I, Keramopoulos A, Davaris P and
Pignatelli M: Abnormal α-catenin expression in invasive breast
cancer correlates with poor patient survival. Histopathology.
40:536–546. 2002.
|
|
11
|
Tagliabue E, Ghirelli C, Squicciarini P,
Aiello P, Colnaghi MI and Menard S: Prognostic value of alpha 6
beta 4 integrin expression in breast carcinomas is affected by
laminin production from tumour cells. Clin Cancer Res. 4:407–410.
1998.PubMed/NCBI
|
|
12
|
Stingl J, Eaves CJ, Kuusk U and Emerman
JT: Phenotypic and functional characterization in vitro of a
multipotent epithelial cell present in the normal adult human
breast. Differentiation. 63:201–213. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jones C, Nonni AV, Fulford L, Merrett S,
Chaggar R, Eusebi V and Lakhani SR: CGH analysis of ductal
carcinoma of the breast with basaloid/myoepithelial cell
differentiation. Br J Cancer. 85:422–427. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Friedrichs K, Ruiz P, Franke F, Gille I,
Terpe HJ and Imhof BA: High expression level of α-6 integrin in
human breast-carcinoma is correlated with reduced survival. Cancer
Res. 55:901–906. 1995.
|
|
15
|
Calaf GM and Russo J: Transformation of
human breast epithelial cells by chemical carcinogens.
Carcinogenesis. 14:483–492. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Calaf GM and Hei TK: Establishment of a
radiation- and estrogen-induced breast cancer. Carcinogenesis.
21:769–776. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Soule HD, Maloney TM, Wolman SR, Peterson
WD Jr, Brenz R, McGrath CM, Russo J, Pauley RJ, Jones RF and Brooks
SC: Isolation and characterization of a spontaneously immortalized
human breast epithelial cell line MCF-10. Cancer Res. 50:6075–6086.
1990.PubMed/NCBI
|
|
18
|
Calaf GM, Russo J and Alvarado ME:
Morphological phenotypes in neoplastic progression of benz (alpha)
pyrene-treated breast epithelial cells. J Submicrosc Cytol Pathol.
32:535–545. 2000.PubMed/NCBI
|
|
19
|
Calaf GM and Hei TK: Oncoprotein
expressions in human breast epithelial cells transformed by
high-LET radiation. Int J Radiat Biol. 77:31–40. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Calaf G, Alvarado M and Hei TK: Beta
catenin is associated with breast cancer progression in
vitro. Int J Oncol. 26:913–921. 2005.PubMed/NCBI
|
|
21
|
Calaf G, Roy D and Hei TK: Immunochemical
analysis of protein in breast epithelial cells transformed by
estrogens and high linear energy transfer (LET) radiation.
Histochem Cell Biol. 124:261–274. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Calaf G, Alvarado ME and Hei TK:
Oncoprotein expression and morphological phenotypes of human breast
epithelial cells transformed by c-Ha-ras oncogene. Oncol Rep.
14:885–893. 2005.PubMed/NCBI
|
|
23
|
Calaf GM and Roy D: Gene expression
signature of parathion-transformed human breast epithelial cells.
Int J Mol Med. 19:741–750. 2007.PubMed/NCBI
|
|
24
|
Calaf GM and Roy D: Gene and protein
expressions induced by 17 beta estradiol and parathion in cultured
breast epithelial cells. Mol Med. 13:255–265. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Calaf GM and Roy D: Cell adhesion proteins
altered by 17β estradiol and parathion in breast epithelial cells.
Oncol Rep. 19:165–169. 2008.
|
|
26
|
Califano A: SPLASH: Structural pattern
localization analysis by sequential histograms. Bioinformatics.
16:341–357. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhou H, Calaf GM and Hei TK: Malignant
transformation of human bronchial epithelial cells with the
tobacco-specific nitrosamine,
4-(methylnitrosamino)-1-(3-pyridyl)-1-butatone. Int J Cancer.
106:821–826. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Peralta Soler A, Knudsen KA, Salazar H,
Han AC and Keshgegian AA: P-cadherin in breast carcinoma indicates
poor survival. Cancer. 86:1263–1272. 1999.PubMed/NCBI
|
|
29
|
Moll R, Mitze M, Frixen UH and Birchmeier
W: Differential loss of E-cadherin expression in infiltrating
ductal and lobular breast carcinoma. Am J Pathol. 143:1731–1742.
1993.PubMed/NCBI
|
|
30
|
Gordon LA, Mulligan KT, Maxwell-Jones H,
Adams M, Walker RA and Jones JL: Breast cell invasive potential
relates to the myoepithelial phenotype. Int J Cancer. 106:8–16.
2003. View Article : Google Scholar : PubMed/NCBI
|