Introduction
Due to the insidious nature of ovarian cancer, few
patients can be diagnosed before the cancer has spread beyond the
ovaries, making ovarian cancer the most lethal gynecological
malignancy and the fifth leading cause of cancer-related mortality
in women worldwide (1,2). Yet, our understanding of the molecular
mechanisms that govern its dissemination remains fragmental.
microRNAs (miRNAs) are small, non-coding RNAs with a
length of ~22 nucleotides, which negatively regulate gene
expression by promoting mRNA cleavage or by translational
suppression (3). miR-335 has been
found to function as an oncogene or tumor-suppressor in a wide
variety of cancers (4). It
functions as a tumor-suppressor in breast cancer and inhibits
metastasis and migration of breast cancer cells (5). Similarly, miR-335 was found to
suppress gastric cancer metastasis in vitro and in
vivo(6). Lynch et
al(7) also found that miR-335
inhibits the invasive and migratory potential of neuroblastoma
cells. Interestingly, miR-335 has been reported to be upregulated
in glioma (8), as a putative miRNA
oncogene in conferring tumorigenic features such as growth and
invasion on malignant astrocytoma cell lines in vitro and
could be potently required for differentiation of malignant glioma
cell lines (9). These findings
suggest that the roles of miR-335 vary based on the type of cancer.
It will be necessary to determine the expression status of miR-335
in each type of tumor.
Previous profiling studies have associated miR-335
with ovarian cancer (10–12). However, its expression and
biological function in ovarian cancer remain unexplored. A search
for miR-335 regulatory targets that may be involved in the
progression of ovarian cancer using online prediction algorithms
has identified Bcl-w, which harbors a putative miR-335 binding site
in its 3′-UTR. Bcl-w is an anti-apoptotic member of the Bcl-2
protein family. It is upregulated in tumors such as gastric,
colorectal, cervical and breast cancer (13–15).
It can promote cell invasion in gastric cancer (13) and inhibit apoptosis in human
colorectal cancers (16). These
results suggest that the dependency of cancer cells on
overexpressed or activated Bcl-w for various tumor-promoting
phenotypes, such as sustained survival, proliferation, invasion and
metastasis, may be different among tissues or cell types. In this
study, we demonstrated that miR-335 may act as a metastasis
suppressor in ovarian cancer by targeting Bcl-w and MMP-2.
Materials and methods
Tissue samples
Samples of human normal ovarian tissues were
obtained from patients who underwent surgery at Wuhan Union
Hospital from 2004 to 2009. The normal tissues were obtained from
patients who underwent wedge biopsy of the ovaries, or adnexectomy
due to myoma or adenomyosis. After ovary removal, the germinal
epithelium was peeled off and subjected to further analysis. After
this, 17 normal ovarian tissues were analyzed. Specimen collection
and archiving of patient data were performed upon written informed
consent and were approved by the ethics committee of the
hospital.
Cell culture and transfection
Human epithelial ovarian cancer cell lines, SKOV3,
ES2, A2780 and CAOV3, were purchased from the China Center for Type
Culture Collection (Wuhan, China) and were propagated in Dulbecco’s
modified Eagle’s medium (DMEM) (Gibco-BRL, Carlsbad, CA, USA)
supplemented with 10% fetal bovine serum (FBS) (Gibco-BRL). For
miRNA-335 overexpression, SKOV3 and ES2 cells were transfected with
miR-335 mimics or a scrambled sequence as miR-control (GenePharma
Co., Ltd., Shanghai, China). For Bcl-w overexpression, SKOV3 cells
were transfected with Bcl-w expression constructs, which had been
rendered insensitive to miR-335 by deletion of its 3′UTR
(GenePharma Co., Ltd.). All transfections were performed using
Lipofectamine™ 2000 reagent (Invitrogen Life Technologies,
Carlsbad, CA, USA) according to the manufacturer’s protocols.
RNA extraction and real-time PCR
analysis
Total RNA was extracted with TRIzol reagent
(Invitrogen Life Technologies). The expression level of miR-335 was
analyzed using the Hairpin-it™ miRNAs qPCR Quantitation kit
(GenePharma, Co. Ltd.), following the manufacturer’s instructions.
U6 was used as the reference gene for normalization.
For determination of Bcl-w expression levels,
reverse transcription was run using RevertAid™ First Strand cDNA
synthesis kit (MBI Fermentas, Flamborough, ON, Canada) according to
the supplied protocol. The 25 μl polymerase chain reaction (PCR)
mixture included 2X SYBR-Green qPCR master mix (Toyobo, Japan), 2.5
μl of cDNA template and 0.3 μM of each primer. Primers for Bcl-w
were as follows: forward, 5′-GAGCCATATAGTTCCTTGGGA-3′ and reverse,
5′-TAGAATAAGTGGGGAGTGGGA-3; and for β-actin, forward,
5′-GCCAACACAGTGCTGTCTGG-3′ and reverse,
5′-GCTCAGGAGGAGCAATGATCTTG-3′. β-actin was used as a reference gene
for normalization. The expression of each gene was defined from the
threshold cycle (Ct), and relative expression levels were
calculated using the 2−ΔΔCT method. All reactions were
run in triplicate.
Cell proliferation analysis
Cell proliferation was measured by detecting
incorporation of 5-ethynyl-2′-deoxyuridine (EdU) during DNA
synthesis using Clik-iT® EdU Imaging kit (Invitrogen
Life Technologies) with minor modifications to the manufacturer’s
protocol. Briefly, cells that had been propagated in a 96-well
plate for 48 h were exposed to 10 μM of EdU for 4 h. After EdU
exposure, cells were fixed with 4% formaldehyde and treated with
0.5% Triton X-100. Proliferative cells were visualized using
Click-iT® reaction cocktail. Subsequently, cells were
counterstained with Hoechst 33342. All assays were carried out in
triplicate.
Migration and invasion assays
To evaluate cell migration and invasion, Transwell
cell migration and Matrigel invasion assays were carried out
according to a previously reported protocol (17). The results are represented as the
means of the cell numbers of 10 randomly selected visual fields
(x200 magnification). All assays were performed in triplicate. In
corroboration with the Transwell cell migration and Matrigel
invasion assays, a wound-healing assay was carried out according to
a previously reported protocol (18). Percent migration was calculated by
measuring the length and width of the cell-free area. The width was
measured at five points along the scratch area and then averaged to
obtain an accurate representation of the entire scratch area.
Percent migration was determined using the following formula: [Δ
area/area (day 0)] × 100.
F-actin immunofluorescence
SKOV3 cells were transfected with miR-335 mimics or
the miR-control, as described above, and plated onto uncoated glass
coverslips in a 12-well plate. The cells were fixed with 4%
formaldehyde, permeabilized in 0.1% Triton X-100 and reacted with
phallotoxins (Invitrogen Life Technologies) for 60 min.
Subsequently, cell nuclei were counterstained with Hoechst 33342.
Images were acquired using an 1X71 digital camera (Olympus, Tokyo,
Japan).
Bioinformatics
For miRNA target gene prediction, we used TargetScan
release 5.1 online software (http://www.targetscan.org/) and PicTac (http://pictar.mdc-berlin.de/).
Protein extraction and western blot
analysis
Total cell extracts were resolved on SDS
polyacrylamide gels and blotted onto a Hybond PVDF membrane (GE
Healthcare, UK). The following antibodies were used: anti-Bcl-w
(1:500 dilution; Cell Signaling Technology, Inc., Beverly, MA,
USA), anti-β-actin (1:1,000 dilution; Santa Cruz Biotechnology,
Inc., Santa Cruz, CA, USA), anti-MMP-2 (1:1,000 dilution; Vector
Laboratories, Burlingame, CA, USA). The antigen-antibody complexes
were visualized using horseradish peroxidase-conjugated secondary
antibody (1:5,000 dilution; Santa Cruz Biotechnology, Inc.).
Statistical analysis
Data are presented as means ± SD from at least 3
separate experiments. Unless otherwise noted, one-way ANOVA was
used for comparisons between groups. All statistical analyses were
carried out with SPSS 12.0 computer software (SPSS, Inc., Chicago,
IL, USA). P<0.05 was considered to indicate a statistically
significant result.
Results
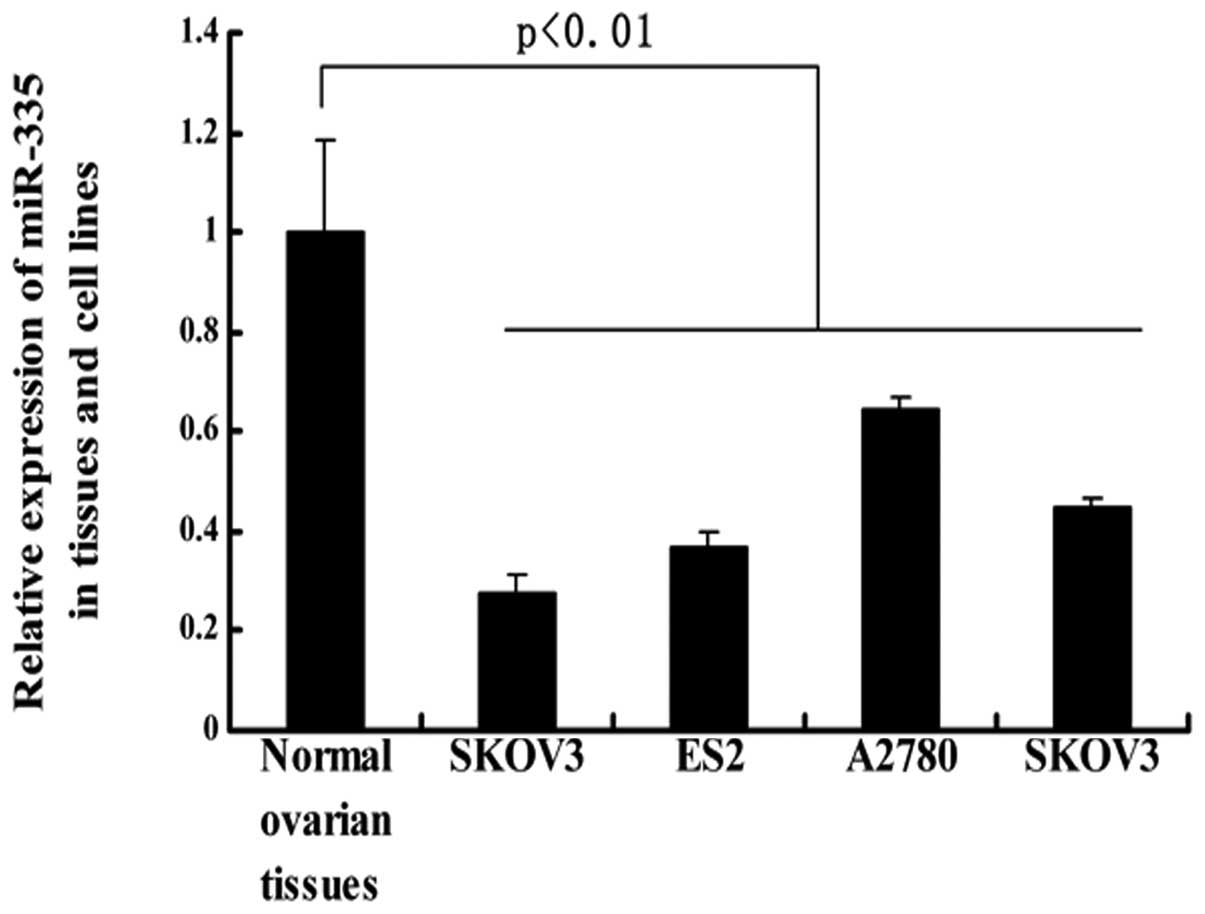
miR-335 is downregulated in ovarian
cancer cell lines
To determine whether miR-335 plays a role in the
tumorigenesis of epithelial ovarian cancer, we examined its
expression in 17 normal ovarian tissue samples using stem-loop
quantitative reverse transcription-polymerase chain reaction
(RT-PCR). We found that miR-335 expression was reduced in ovarian
cancer cell lines in comparison with normal ovarian epithelium
(Fig. 1).
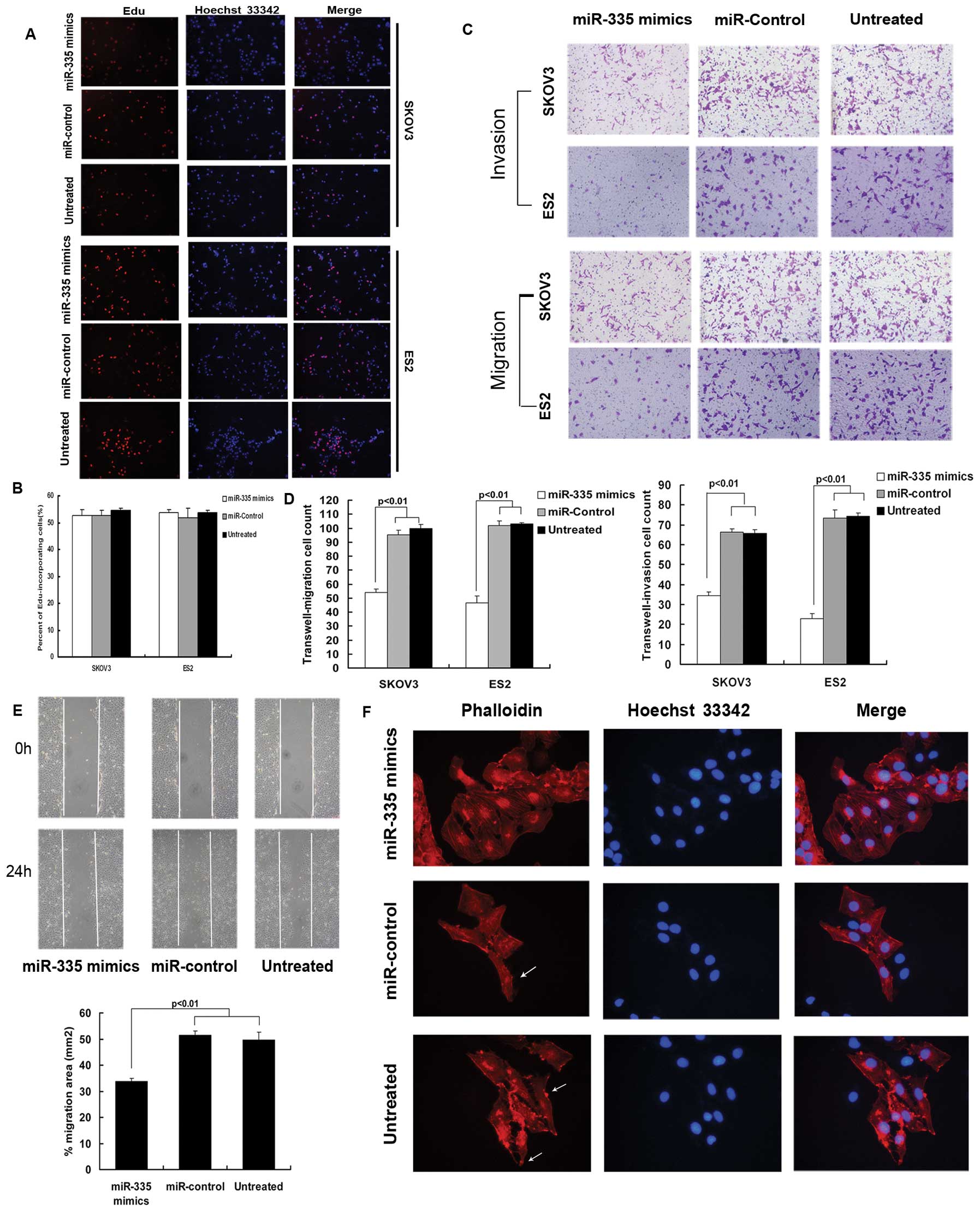
miR-335 suppresses metastasis-relevant
traits in vitro
Given that miR-335 is downregulated in ovarian
cancer cell lines, we proceeded to explore its impact on relevant
biological behavior of ovarian cancer. Since the expression of
miR-335 was most significantly reduced in the SKOV3 and ES2 cell
lines, we chose these for further study. Thus, we overexpressed
miR-335 in SKOV3 and ES2 cell lines by transiently transfecting
them with hsa-miR-335 mimics. Ectopic miR-335 did not affect cell
proliferation in vitro (Fig. 2A
and B), but did reduce the migratory and invasive ability of
SKOV3 and ES2 cells (Fig. 2C and
D). The effect of miR-335 overexpression on cell migration was
further supported by the wound-healing assay (Fig. 2E). Moreover, under a florescence
microscope, numerous F-actin bundles were clearly visible in SKOV3
cells transfected with the miR-335 mimics, while control cells
contained markedly fewer, finer and shorter bundles and exhibited
an elongated morphology with lamellipodia (Fig. 2F), suggesting miR-335 exerts
anti-migratory activity in ovarian cancer cells at least partly by
inhibiting actin polymerization.
miR-335 suppresses the invasiveness of
ovarian cancer by targeting Bcl-w and Bcl-w effector, MMP-2
For miRNA target gene prediction, we used TargetScan
release 5.1 online software (http://www.targetscan.org/, Whitehead Institute for
Biomedical Research, Cambridge, MA, USA). Among a total of 146
genes that was potentially targeted by miR-335, we found that Bcl-w
may contribute to the metastasis of ovarian cancer. To establish
that miR-335 targets Bcl-w in ovarian cancer, we overexpressed
miR-335 in SKOV3 and ES2 cells. We found that ectopic miR-335
significantly repressed Bcl-w protein levels while no difference
was detected between the miR-control and untreated cells. Moreover,
a concomitant repression of the Bcl-w effector, MMP-2, was also
observed (Fig. 3A and B). These
observations demonstrated that miR-335 targets Bcl-w in ovarian
cancer. In spite of these findings, no reduction in Bcl-w was
observed at the mRNA level when miR-335 was modulated in SKOV3 and
ES2 cells (Fig. 3C and D).
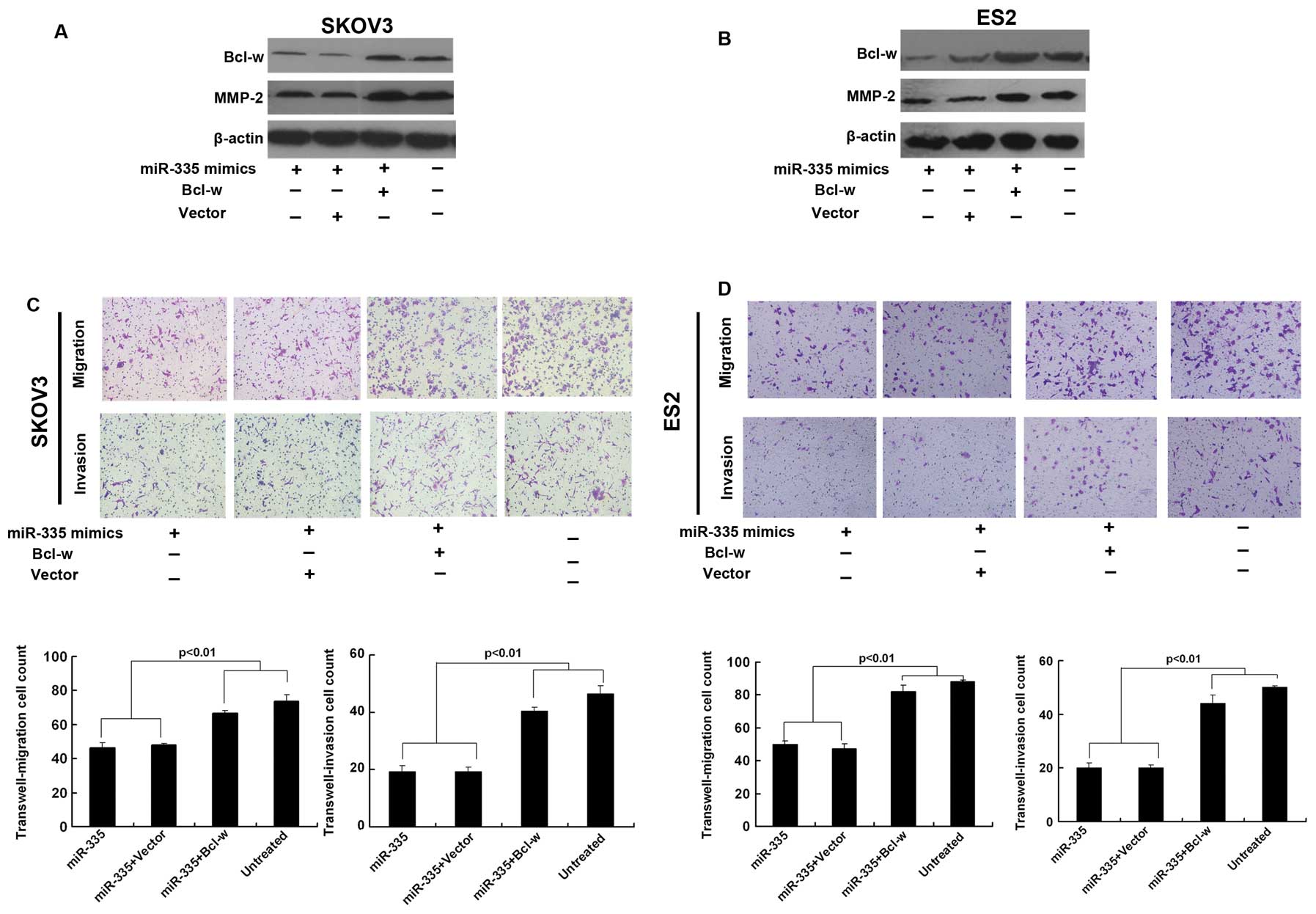
Ectopic Bcl-w reverses miR-335-mediated
invasion defects
To establish that the ability of miR-335 to impede
cell invasion is mainly mediated by the downregulation of Bcl-w, we
concomitantly transfected SKOV3 and ES2 cells with 335 mimics and
Bcl-w expression constructs. In these cells, ectopic Bcl-w reversed
the miR-335-imposed invasion defects (Fig. 4C and D). Under the same conditions,
we observed a significant increase in MMP-2 (Fig. 4A and B).
Discussion
miRNAs can modulate a wide variety of biological
processes. In the present report, we demonstrated that a single
human miRNA, miR-335, is endowed with the ability to repress the
pro-metastatic target, Bcl-w, and thereby to inhibit invasion in
ovarian cancer.
To date, only a limited number of miRNAs with
pro-metastatic or anti-metastatic functions have been identified in
ovarian cancer (19–23), miR-34 has been reported to be
endowed with the ability to mitigate invasion (19). miR-21 has been categorized as a
pro-metastatic gene (22). However,
its specific contributions to metastasis are not easily discerned
as they have significant effects on cell proliferation at the same
time. In contrast, miR-335 obstructs invasion without exerting
confounding influences on cell proliferation. As such, it may aptly
be categorized as an ‘exclusive invasion suppressor’.
Bcl-w is an anti-apoptotic member of the Bcl-2
protein family. It suppresses apoptosis by direct interaction with
pro-apoptotic members and by blocking their apoptotic activities
(24–27), and previous evidence suggests that
the expression of Bcl-w is associated with infiltrative morphotypes
(14). Recently, reporter gene
assays have validated its role as an effector of miR-335 (6). Although mutations in the
anti-apoptotic Bcl-2 family members (Bcl-xL, Mcl-1, Bcl-w and A1)
have not been identified as a cause of tumors, high expression of
these proteins can contribute to carcinogenesis in cooperation with
other proto-oncogenes (28). Bcl-w
can promote cell invasion by activating the PI3K-Akt-Sp1-MMP-2
pathway (13) and blocking the
invasion suppression action of Bax (29). In our research, we demonstrated that
miR-335 supressed the invasiveness of ovarian cancer by targeting
Bcl-w and Bcl-w effector, MMP-2.
In spite of this, we found that miR-335-mediated
defects in invasion could not be fully mitigated by ectopic Bcl-w,
and that miR-335 affected actin polymerization, an important step
in EMT. Thus, the possibility that miR-335-mediated defects in
invasion are achieved by recruiting a cohort of effectors cannot
formally be excluded, again highlighting the pleiotropic action of
miRNAs.
miRNAs effect gene silencing via both translational
repression or mRNA degradation pathways. When an miRNA and its
cognate mRNA interact with perfect complementarity, the target mRNA
is directly cleaved, resulting in the reduction in the abundance of
the target mRNA. However, in animals, miRNAs generally interact
with target mRNAs with only partial complementarity. This mode of
recognition results in translational repression of the target mRNA
with much smaller effects on the level of target mRNA abundance
(6,12,15,30).
In our study, it was found that ectopic miR-335 significantly
reduced Bcl-w expression at the protein level. However, no
corresponding decrease in Bcl-w expression was observed at the mRNA
level. In fact, such a finding is not unprecedented (19) and is consistent with the notion that
mammalian cells favor translational repression.
In summary, our results suggest that decreased
miR-335 facilitates ovarian cancer invasion via the Bcl-w pathway.
To our knowledge, this represents the first observation of reduced
miR-335 expression in human ovarian cancer. Moreover, as metastases
are responsible for patient mortality in ovarian cancer, the
ability of miR-335 to impede invasion may prove to be clinically
useful.
Acknowledgements
We thank the Department of Obstetrics and
Gynecology, Union Hospital, Wuhan, Hubei, China, for providing the
tissue samples. This study was supported by the Scientific Research
Foundation for the Returned Overseas Chinese Scholars, the State
Education Ministry.
References
|
1
|
Jemal A, Tiwari RC, Murray T, et al:
Cancer statistics, 2004. CA Cancer J Clin. 54:8–29. 2004.
View Article : Google Scholar
|
|
2
|
Moss C and Kaye SB: Ovarian cancer:
progress and continuing controversies in management. Eur J Cancer.
38:1701–1707. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Esquela-Kerscher A and Slack FJ: Oncomirs
- microRNAs with a role in cancer. Nat Rev Cancer. 6:259–269. 2006.
View Article : Google Scholar
|
|
4
|
Kong YW, Ferland-McCollough D, Jackson TJ
and Bushell M: microRNAs in cancer management. Lancet Oncol.
13:e249–e258. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tavazoie SF, Alarcón C, Oskarsson T, et
al: Endogenous human microRNAs that suppress breast cancer
metastasis. Nature. 451:147–152. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xu Y, Zhao F, Wang Z, et al: microRNA-335
acts as a metastasis suppressor in gastric cancer by targeting
Bcl-w and specificity protein 1. Oncogene. 31:1398–1407. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lynch J, Fay J, Meehan M, et al: miRNA-335
suppresses neuroblastoma cell invasiveness by direct targeting of
multiple genes from the non-canonical TGF-β signalling pathway.
Carcinogenesis. 33:976–985. 2012.PubMed/NCBI
|
|
8
|
Schmitz KJ, Helwig J, Bertram S, et al:
Differential expression of microRNA-675, microRNA-139–3p and
microRNA-335 in benign and malignant adrenocortical tumours. J Clin
Pathol. 64:529–535. 2011.PubMed/NCBI
|
|
9
|
Shu M, Zhou Y, Zhu W, et al: microRNA 335
is required for differentiation of malignant glioma cells induced
by activation of cAMP/protein kinase A pathway. Mol Pharmacol.
81:292–298. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dahiya N, Sherman-Baust CA, Wang TL, et
al: microRNA expression and identification of putative miRNA
targets in ovarian cancer. PLoS One. 3:e24362008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sorrentino A, Liu CG, Addario A, Peschle
C, Scambia G and Ferlini C: Role of microRNAs in drug-resistant
ovarian cancer cells. Gynecol Oncol. 111:478–486. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wyman SK, Parkin RK, Mitchell PS, et al:
Repertoire of microRNAs in epithelial ovarian cancer as determined
by next generation sequencing of small RNA cDNA libraries. PLoS
One. 4:e53112009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bae IH, Park MJ, Yoon SH, et al: Bcl-w
promotes gastric cancer cell invasion by inducing matrix
metalloproteinase-2 expression via phosphoinositide 3-kinase, Akt,
and Sp1. Cancer Res. 66:4991–4995. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lee HW, Lee SS, Lee SJ and Um HD: Bcl-w is
expressed in a majority of infiltrative gastric adenocarcinomas and
suppresses the cancer cell death by blocking stress-activated
protein kinase/c-Jun NH2-terminal kinase activation.
Cancer Res. 63:1093–1100. 2003.PubMed/NCBI
|
|
15
|
Wilson JW, Nostro MC, Balzi M, et al:
Bcl-w expression in colorectal adenocarcinoma. Br J Cancer.
82:178–185. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kitamura S, Kondo S, Shinomura Y, et al:
Met/HGF receptor modulates bcl-w expression and inhibits apoptosis
in human colorectal cancers. Br J Cancer. 83:668–673. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lü MH, Li CZ, Hu CJ, et al: microRNA-27b
suppresses mouse MSC migration to the liver by targeting SDF-1α in
vitro. Biochem Biophys Res Commun. 421:389–395. 2012.PubMed/NCBI
|
|
18
|
Moore LD, Isayeva T, Siegal GP and
Ponnazhagan S: Silencing of transforming growth factor-β1 in situ
by RNA interference for breast cancer: implications for
proliferation and migration in vitro and metastasis in vivo. Clin
Cancer Res. 14:4961–4970. 2008.
|
|
19
|
Corney DC, Hwang CI, Matoso A, et al:
Frequent downregulation of miR-34 family in human ovarian cancers.
Clin Cancer Res. 16:1119–1128. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fan X, Liu Y, Jiang J, et al: miR-20a
promotes proliferation and invasion by targeting APP in human
ovarian cancer cells. Acta Biochim Biophys Sin. 42:318–324. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li J, Liang S, Yu H, Zhang J, Ma D and Lu
X: An inhibitory effect of miR-22 on cell migration and invasion in
ovarian cancer. Gynecol Oncol. 119:543–548. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lou Y, Yang X, Wang F, Cui Z and Huang Y:
MicroRNA-21 promotes the cell proliferation, invasion and migration
abilities in ovarian epithelial carcinomas through inhibiting the
expression of PTEN protein. Int J Mol Med. 26:819–827.
2010.PubMed/NCBI
|
|
23
|
Tsuda N, Kawano K, Efferson CL and
Ioannides CG: Synthetic microRNA and double-stranded RNA targeting
the 3′-untranslated region of HER-2/neu mRNA inhibit HER-2 protein
expression in ovarian cancer cells. Int J Oncol. 27:1299–1306.
2005.
|
|
24
|
Antonsson B: Bax and other pro-apoptotic
Bcl-2 family ‘killer-proteins’ and their victim the mitochondrion.
Cell Tissue Res. 306:347–361. 2001.
|
|
25
|
Bae IH, Yoon SH, Lee SB, Park JK, Ho JN
and Um HD: Signaling components involved in Bcl-w-induced migration
of gastric cancer cells. Cancer Lett. 277:22–28. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gross A, McDonnell JM and Korsmeyer SJ:
BCL-2 family members and the mitochondria in apoptosis. Genes Dev.
13:1899–1911. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Uren RT, Dewson G, Chen L, et al:
Mitochondrial permeabilization relies on BH3 ligands engaging
multiple prosurvival Bcl-2 relatives, not Bak. J Cell Biol.
177:277–287. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhivotovsky B and Orrenius S:
Carcinogenesis and apoptosis: paradigms and paradoxes.
Carcinogenesis. 27:1939–1945. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kim EM, Kim J, Park JK, et al: Bcl-w
promotes cell invasion by blocking the invasion-suppressing action
of Bax. Cell Signal. 24:1163–1172. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wightman B, Ha I and Ruvkun G:
Posttranscriptional regulation of the heterochronic gene lin-14 by
lin-4 mediates temporal pattern formation in C. elegans.
Cell. 75:855–862. 1993. View Article : Google Scholar : PubMed/NCBI
|