Introduction
Bleomycin hydrolase (BLH), a member of the papain
family of cysteine proteases, is metabolically inactivated
antitumor antibiotic bleomycin (BLM) by deamination (1). It functions as a tumor suppressor gene
in hepatocellular carcinoma (2).
Accumulating data show that BLH plays multifaceted roles under
different physiological and pathological conditions, including
preparation of peptides for antigen presentation (3), involvement in the pathogenesis of
Alzheimer’s disease (4,5), detoxification of homocysteine
thiolactone (6), N-terminal
proteolysis of mutant huntingtin (7) and breakdown of deiminated filaggrin
into amino acids (8). Analysis of
the human BLH gene indicates that its features are shared by those
characteristic of housekeeping genes (9,10). For
example, BLH is expressed at high levels in human tissues except
for the kidneys and lungs (11,12).
We recently demonstrated that BLH serves as a biomarker for
predicting the efficacy of treatment of tumors with BLM (13).
The level of BLH in tumor cells affects the efficacy
of BLM (13–16). Therefore, it is necessary to
understand how BLH is regulated to facilitate the development of
new antibiotics structurally related to BLM and to demonstrate the
pathological roles in the diseases. Therefore, in the present study
we determined the stability of BLH in human tumor cells.
Materials and methods
Drugs and chemicals
BLM was purchased from Nippon Kayaku (Tokyo, Japan)
and was dissolved in phosphate-buffered saline (PBS) at 5 mg/ml and
stored at −20°C prior to use. Hydroxycamptothecin (HCPT) was
purchased from Shenzhen Wanle Pharmaceutical Co., Ltd., (Guangzhou,
China) and was dissolved in PBS at 2 mg/ml. Caspase inhibitors
Q-VD-oph and Z-DEVD-FMK were purchased from R&D Systems Inc.
(Minneapolis, MN, USA) and dissolved in dimethyl sulfoxide (DMSO)
before use. Cycloheximide (CHX) was purchased from Millipore
(Billerica, MA, USA) and dissolved in ethanol at 100 mg/ml.
Etoposide (VP-16), Z-Leu-Leu-Leu-al (MG132), and all the chemicals
except for those specifically mentioned were purchased from
Sigma-Aldrich Inc. (St. Louis, MO, USA). VP-16 and MG132 were
dissolved in DMSO at 50 mM.
Cell lines and cell culture
The HeLa human cervical cancer cell line was
purchased from the American Type Culture Collection (ATCC;
Rockville, MD, USA) and the human colon carcinoma HCT-116 cell line
was a gift from Bert Vogelstein (Johns Hopkins University,
Baltimore, MD, USA). The cell lines were cultured in Dulbecco’s
modified Eagle’s medium supplemented with 10 % fetal bovine serum
(Thermo Fisher Scientific Inc., Waltham, MA, USA). All cultures
were maintained in an incubator at 37°C in a humidified atmosphere
with 5% CO2.
Western blotting
The method was performed according to the protocol
previously published (13). The
sources of primary antibodies were as follows:
rabbit-anti-cleaved/total PARP-1 (9532), rabbit-anti-cleaved
caspase-3 (9661) and cleaved caspase-9 (9501), all from Cell
Signaling Technology, Inc. (Danvers, MA, USA). The rabbit-anti-BLH
(sc-68888), mouse-anti-His tag (sc-804) and mouse-anti-actin
(sc-1616) were from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA,
USA). All primary antibodies were diluted to 1:1,000.
Detection of apoptotic cells by Annexin
V-FITC/propidium iodide
To quantify apoptosis, cells were stained with
Annexin V and propidium iodide (PI) using an Annexin V-FITC/PI
Apoptosis kit (BD Biosciences, San Jose, CA, USA), following the
protocol provided by the manufacturer. The fluorescence intensity
was measured using a BD FACSCalibur flow cytometer.
Heterologous expression and purification
of recombinant human BLH
Human BLH gene was cloned using RT-PCR with
the forward primer 5′-CGCGGATCCATGAGCAGCT
CGGGACTGAATTCG-3′ and the reverse primer 5′-CCG CTCGAGTCACTCAGCCAAAGCTCCCATG-3′,
which contain the initiator methionine and termination codons and
the BamHI and XhoI restriction sites, respectively,
as indicated by the underlined bases. cDNA of the template was
isolated from 293T cells. The cDNA was amplified for 25 cycles as
follows: denaturing for 15 sec at 98°C, annealing for 90 sec at
60°C, and primer extension for 30 sec at 72°C. PCR was carried out
using PrimeSTAR HS DNA polymerase kit from Takara Bio Inc. The
product was cloned into the BamHI and XhoI
restriction sites of cDNA3.1 (Invitrogen) and completely sequenced
in both directions. E. coli BL21 (DE3) expressing
recombinant BLH was generated as described above, following cloning
of BLH cDNA into the BamHI and XhoI site of the
pET32a (Novagen, Darmstadt, Germany). Expression of recombinant BLH
was induced with 1 mM IPTG. Bacterial cells were sonicated and
lysates were cleared by centrifugation at 12,000 × g at 4°C for 15
min. The supernatants were used for affinity purification on a
column containing Ni-NTA-agarose. Protein concentrations of cell
lysates were determined using the Bio-Rad Protein Assay. The BLH
content was determined in comparison with BSA using SDS-PAGE.
Assay of BLH activity by high performance
liquid chromatography
The metabolism of bleomycin was assessed using high
performance liquid chromatography (HPLC), which separates bleomycin
A2 and B from their inactive metabolites. Briefly,
partially-purified BLH (2 mg/ml) was incubated with 5 μg BLM in 40
μl of reaction buffer (20 mM Tris, pH 7.5), at 37°C for 1 h. The
reaction was stopped by the addition of 10 μl of methanol, the
mixture was centrifuged, and the resulting supernatant fractions
were injected into a ZORBAX Eclipse Plus C18 column (4.6 × 250 mm;
Agilent Technology Inc., Santa Clara, CA, USA). The mixtures were
eluted at 1 ml/min with 6% methanol, 6% acetonitrile and 88% acetic
acid (pH 4.2) and were detected by their absorbance at 254 nm.
Assays of BLH cleavage in vitro
Partially-purified BLH (50 ng) was incubated with
0.1 unit caspase-6, -8, -9 and-10 (BioVision, Mountain View, CA,
USA), respectively, in 20 μl reaction buffer (50 mM HEPES, pH 7.2,
0.1% CHAPS, 10 mM EDTA, 5% glycerol, 10 mM DTT, 50 mM NaCl and 100
mg/ml BSA) at 37°C for 1 h, or with 150 ng caspase-3
(Sigma-Aldrich) in 20 μl reaction buffer (10 mM HEPES, pH 7.0, 5 mM
MgCl2, 5 mM EDTA, 10% glycerol, 1 mM DTT, 50 mM NaCl and
100 mg/ml BSA) at 30°C for 1 h. The inhibition of BLH cleavage by
caspases was confirmed using 20 μM Q-VD-oph. The resulting mixtures
were separated by SDS-PAGE and detected by western blotting.
Statistical analysis
SPSS 17.0 software was used for statistical
analysis. Student’s t-test was applied for comparing the means of 2
groups and a P-value of <0.05 was considered to indicate a
statistically significant difference.
Results
Reduction of BLH levels during
apoptosis
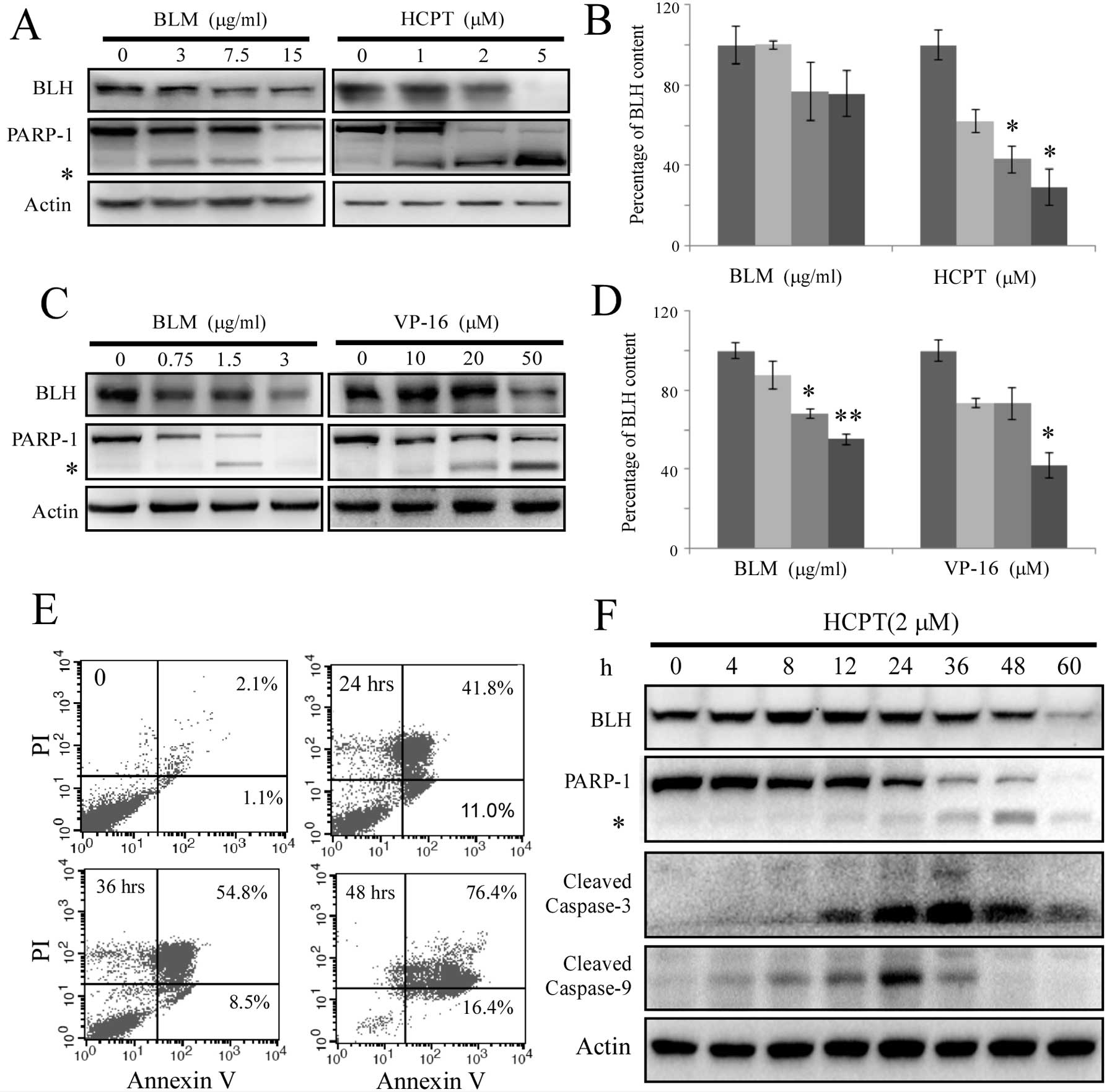
To verify whether the expression level of BLH was
affected during apoptosis, HCT-116 cells were treated for 48 h with
lethal concentrations of BLM or HCPT. The results showed that the
level of BLH was markedly reduced by treatment with high
concentrations of drug (Fig. 1A and
B). Similar results were obtained within 48 h when HeLa cells
were exposed to BLM and VP-16 (Fig. 1C
and D).
The presence of apoptotic cells was confirmed by
Annexin V/PI staining after HCT-116 cells were treated with 2 μM
HCPT for different times (Fig. 1E).
Approximately 53% of cells showed apoptotic features after exposure
to HCPT for 24 h. The levels of BLH and activation of the caspase
pathway were monitored after exposure to 2 μM HCPT (Fig. 1F). Upregulation of BLH expression
was detected after treatment with 2 μM HCPT for 8 h. The reduction
of BLH levels began at 24 h. Activation of caspase-3 and caspase-9
was observed within 8–12 h. The times at which BLH was degraded
were similar to those for cleavage of PARP-1. The data suggest that
the degradation of BLH was triggered by the intrinsic apoptotic
pathway.
Cleavage of BLH by caspases during
apoptosis
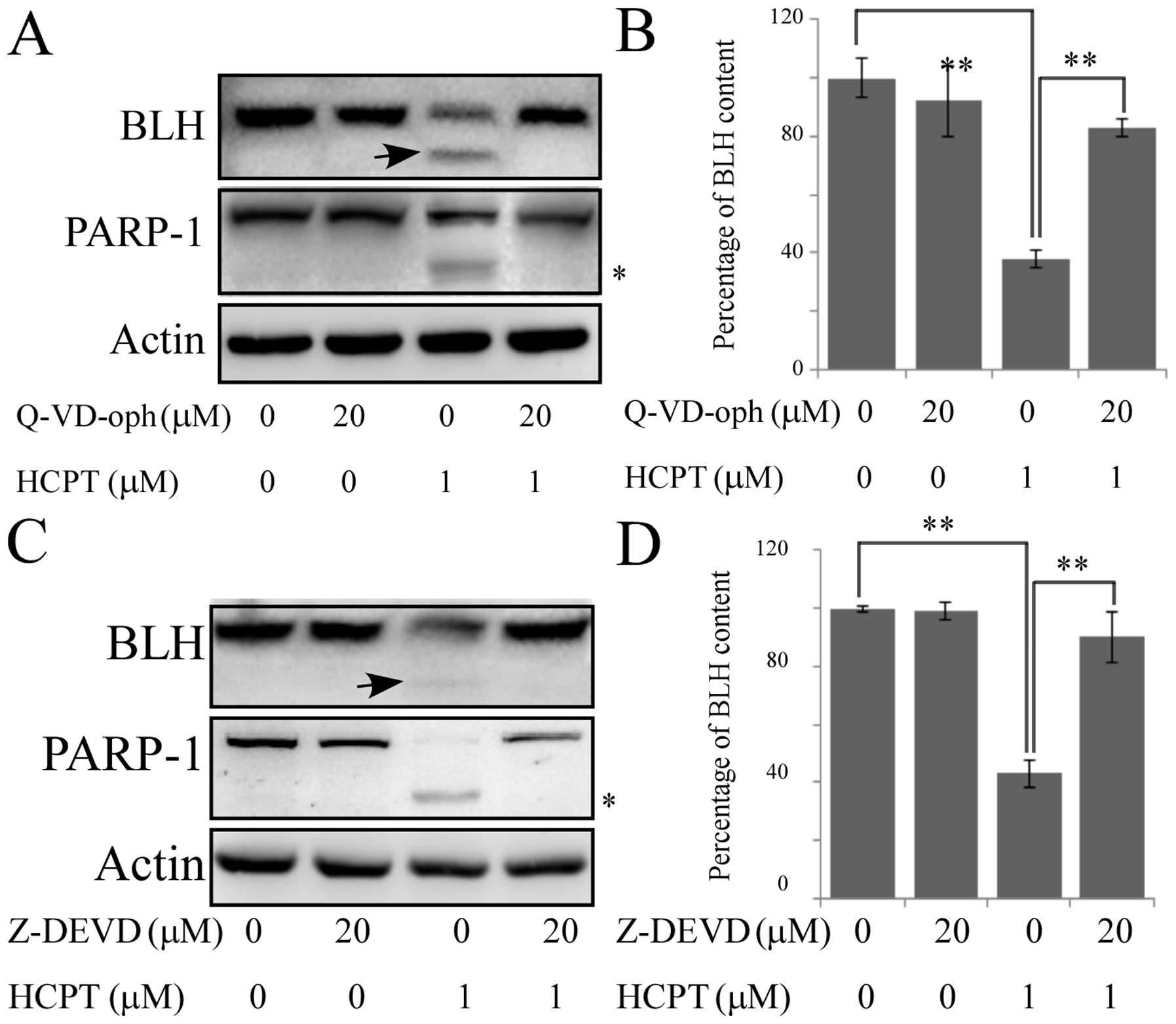
To identify the mechanism for the reduction of the
levels of BLH due to cleavage by caspase during apoptosis, the
broad-spectrum pan-caspase inhibitor Q-VD-oph was used. The
cleavage of PARP-1 and the levels of BLH were reduced after
exposure to 1 μM HCPT by treatment of HCT-116 cells with 20 μM
Q-VD-oph (Fig. 2A, lanes 3, 4 and
B), indicating that cleavage of BLH was caspase dependent. We
compared the sequence of BLH with recognition consensus of
caspases. Only 1 potential cleavage site for caspase-3 was found.
It localizes to Asp39 in the N-terminus of BLH (DLLD), a caspase-3
specific consensus recognition sequence (17). The reduction in the level of BLH was
significantly inhibited by the caspase-3-specific inhibitor
Z-DEVD-FMK (Fig. 2C and D),
consistent with the predicted presence of a caspase-3 recognition
site. Similar results were also observed during apoptosis of HeLa
cells induced by BLM and VP-16 (data not shown).
Hydrolysis of BLM by recombinant BLH
The findings described above that degradation of BLH
is caspase-dependent prompted us to further characterize the
cleavage of BLH in vitro. Human recombinant BLH was
expressed in E. coli BL21 and partially purified using a
Ni-NTA-agarose column. The concentration of the resultant
recombinant protein was estimated to be ~2 mg/ml. The activity of
recombinant BLH was confirmed by its ability to hydrolyze BLM, and
the resultant reaction mixtures were analyzed using reversed phase
chromatography monitored at 254 nm. The results showed that the
recombinant BLH converted BLM-A2 and B2 to its known metabolites,
and the hydrolysis of BLM-B2 was faster than that of BLM-A2, with
most B2 hydrolyzed at 20 min and only approximately two-thirds of
A2 at 60 min (Fig. 3).
Cleavage of BLH by caspase-3 and
caspase-9 in vitro
We incubated recombinant BLH in vitro with
caspase-3, -6, -8, -9 and -10, respectively. Recombinant BLH was
only cleaved detectably by caspase-3 and caspase-9. When the
recombinant BLH was incubated with purified caspase-3 at 37°C, both
its degradation product and cleaved band were detected over a
period of 12 h (Fig. 4A). The
caspase-3-specific inhibitor Z-DEVD-FMK inhibited degradation of
BLH (Fig. 4B). The small amount of
BLH cleavage by caspase-9 was also detected using an anti-His Tag
antibody but not by an anti-BLH antibody (Fig. 4C). This cleavage reaction was
inhibited by Q-VD-oph (Fig.
4D).
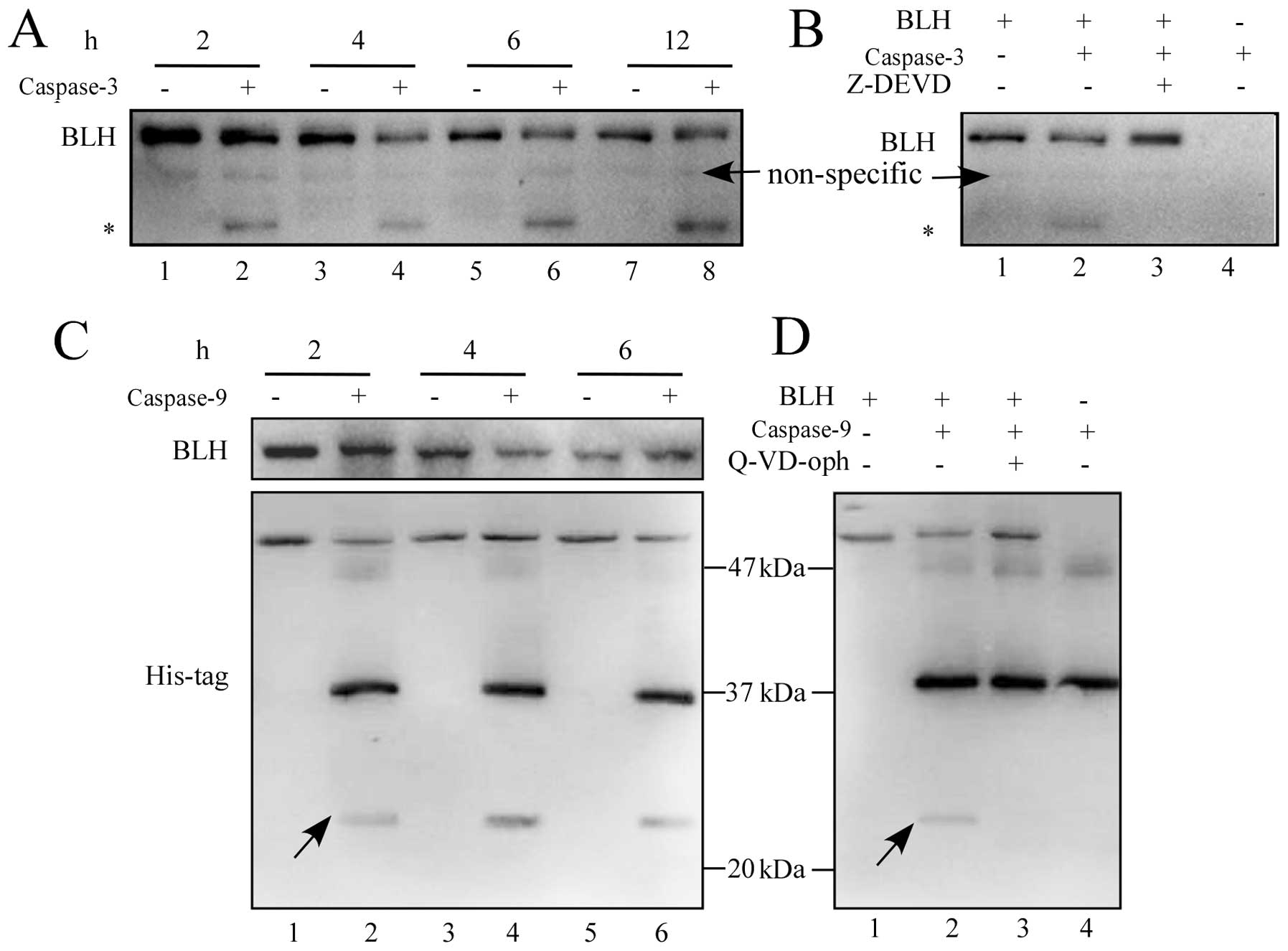 | Figure 4Cleavage of BLH by caspase-3 and
caspase-9 in vitro. (A) Recombinant BLH (50 ng) was
incubated with purified caspase-3 (50 ng) at 30°C for 2, 4, 6 and
12 h and the reaction mixtures were subjected to western blotting
using 12.5% SDS-PAGE gel and an anti-BLH antibody (lanes 1–8). (B)
Cleavage of BLH by caspase-3 was specifically inhibited by 20 μM
Z-DEVD-FMK. Lane 1, BLH alone; lane 2, BLH plus caspase-3 for 2 h;
lane 3, BLH plus caspase-3 and Z-DEVD-FMK; lane 4, 50 ng caspase-3
alone. (C) Recombinant BLH (50 ng) was incubated at 37°C with
purified caspase-9 (1 unit) for 2, 4 and 6 h, and the reaction
mixtures were subjected to western blotting using a 12.5% SDS-PAGE
gel and anti-His Tag antibody (lanes 1–6). (D) Cleavage of BLH by
caspase-9 was inhibited by 20 μM Q-VD-oph. Lane 1, BLH alone; lane
2, BLH plus caspase-9 for 2 h; lane 3, BLH plus caspase-9 and
Q-VD-oph; lane 4, 1 unit caspase-9 alone. The asterisk (*)
indicates a fragment of BLH. One representative result from 3
independent experiments is shown. |
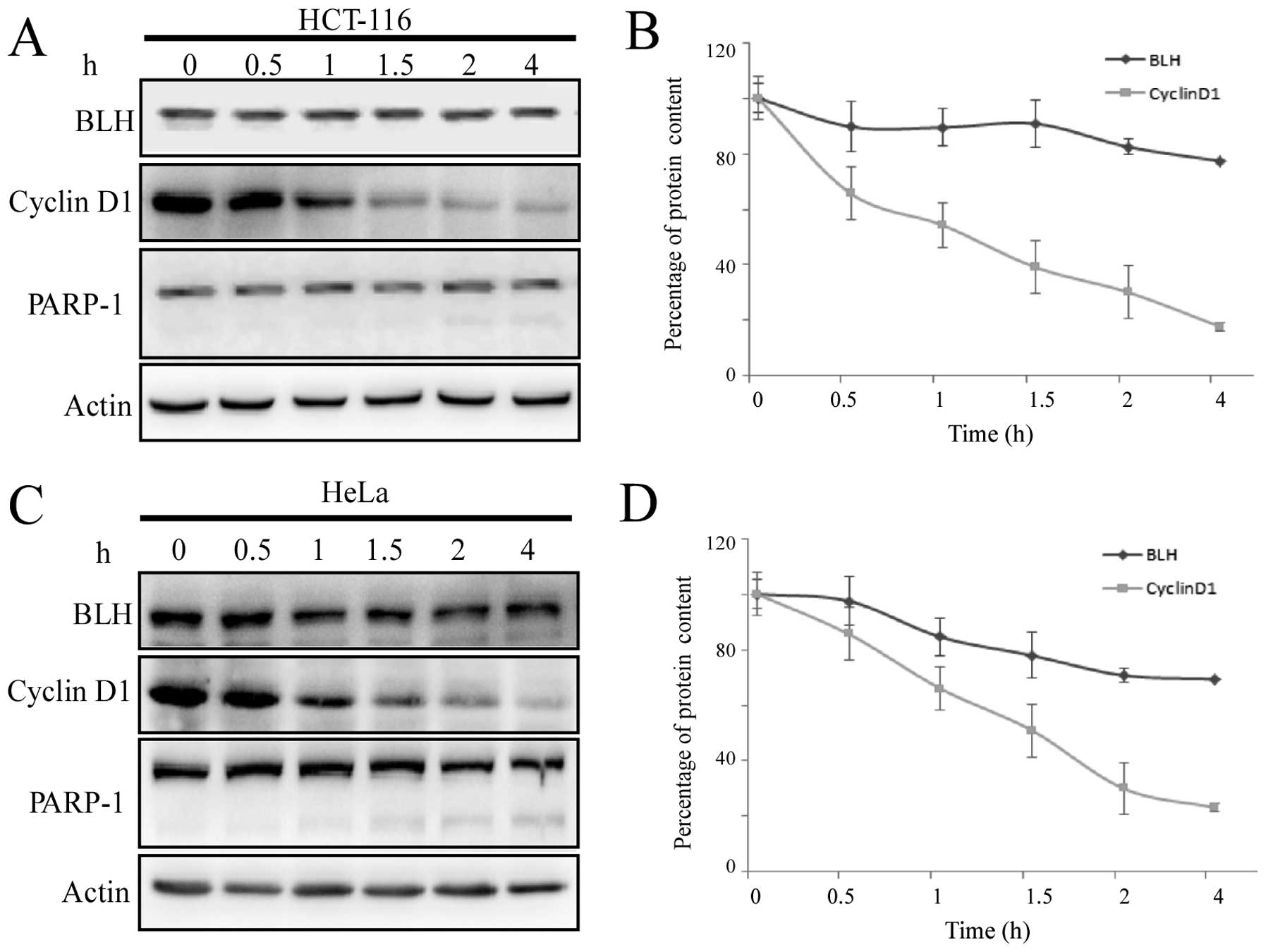
Lower rate of BLH degradation at normal
culture conditions
To determine whether the degradation of BLH is
involved in the ubiquitin-proteasome pathway, de novo
protein synthesis was blocked with the protein synthesis inhibitor
CHX under normal culture conditions. The stability of short-lived
cyclin D1 was monitored for comparison. Treatment of HCT-116 cells
with 20 μg/ml CHX resulted in a time-dependent reduction of the
cyclin D1 levels, and only 50% of its initial level remained after
exposure to CHX for 1 h. By contrast, the level of BLH remained
constant initially and was then reduced by ~20% after treatment
with CHX for 4 h (Fig. 5A and B). A
small amount of the cleaved PARP fragment was observed at 4 h,
suggesting induction of apoptosis by CHX. This prevented us from
further calculations of the half-life of BLH degradation using this
method. These same results were obtained using HeLa cells (Fig. 5C and D).
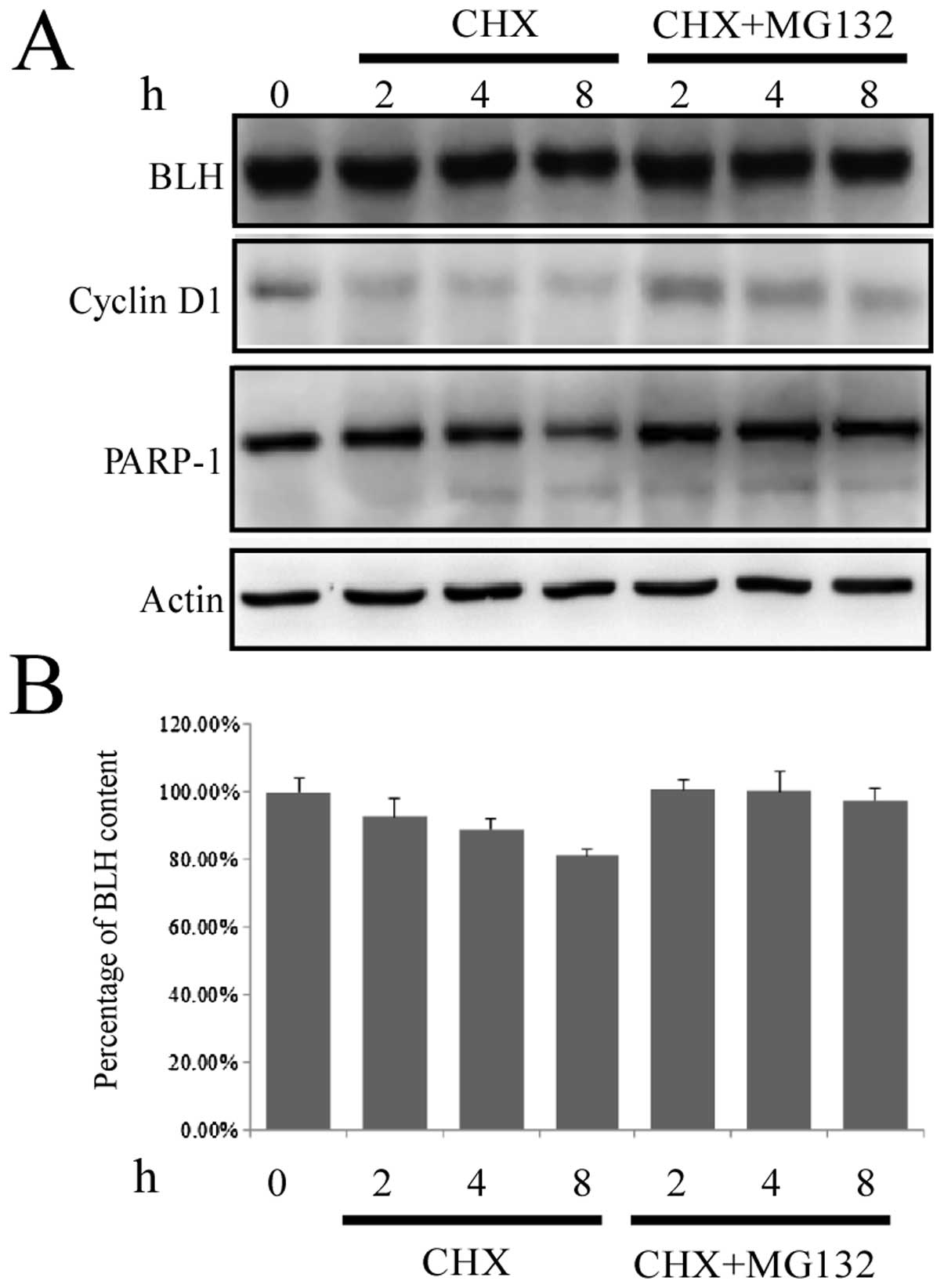
To verify whether BLH is degraded via the
ubiquitin-proteasome pathway, HCT-116 cells were treated with CHX
plus 50 μM MG132, a specific inhibitor of the ubiquitin-proteasome
pathway. The degradation of cyclin D1 was completely suppressed,
and degradation of BLH was also inhibited (Fig. 6), indicating that BLH is degraded
via the ubiquitin-proteasome pathway.
Discussion
In the present study, we investigated the
degradation of BLH under different conditions. To the best of our
knowledge, this is the first report showing that degradation of BLH
is caspase-dependent during apoptosis and that BLH can be cleaved
by caspase-3 and caspase-9 in vitro. In apoptotic pathways,
caspases act as signal transducers, directing the cell toward death
by cleaving numerous substrates (18). We identified a caspase-3 recognition
site at Asp39 in the N-terminus of BLH (DLLD), but no site for
caspase-9. This could be explained by the requirement for a
tertiary structure, as caspases can cleave at ‘non-canonical’
sites, such as the viral caspase inhibitor p35 bound to caspase-8
(19).
Our present results are consistent with data of
others showing that the half-life of BLH is greater than 6 h
(20). However, we could not
determine the half-life of BLH precisely, as CHX treatment led to
apoptotic degradation of BLH. Protein stability plays a role in the
abundance of cellular proteins, and several housekeeping proteins
tend to have stable mRNAs and proteins to improve energy efficiency
(21). The function of BLH is
evolutionarily conserved from yeasts to humans and the yeast
homologue of BLH metabolizes BLM in mammalian cells (22). The features of BLH are
characteristic of housekeeping genes (9). The relationship between stability of
BLH and its function is being investigated in our laboratory.
BLH can hydrolyze the bleomycin to generate the
inactive deamino form. The mice with knockout of BLH gene are very
sensitive to bleomycin (15). The
patients with homozygous variant G/G of BLH gene SNP (A1450G) are
associated with reduced survival in testicular germ cell cancer
(16). The expression of BLH
increased after exposure to low concentrations of bleomycin
(13). All data indicate that the
BLH level plays an important roles in bleomycin action. The present
results provide new evidence for the regulation of BLH level that
may aid in the translational development of new bleomycin-related
antibiotics.
Loss of BLH leads to astrogliosis and behavioral
changes in mice (23), suggesting
its importance in the functions of neurons. Although it has not
been confirmed that genetic polymorphisms of BLH are
involved in the genesis of Alzheimer’s disease, regulation of the
secretion of amyloid precursor protein by BLH plays a function
(5). Accumulating data show that
apoptosis of neurons is involved in the genesis of Alzheimer’s
disease (24). Furthermore, the
levels of BLH activity are reduced in patients with Alzheimer’s
disease (4). This finding might be
explained by the degradation of BLH during the apoptosis of neurons
by a mechanism similar to that demonstrated here.
In summary, the degradation of BLH can occur through
the caspase-3 and ubiquitin-proteasome pathways. The results
presented here facilitate further studies of BLH functions under
different physiological and pathological conditions.
Acknowledgements
The present study was supported by grants from the
National Scientific Foundation of China (81273553) and the National
S&T Major Special Project on Major New Drug Innovation
(2012ZX09301002-001-015).
References
|
1
|
Chen J and Stubbe J: Bleomycins: towards
better therapeutics. Nat Rev Cancer. 5:102–112. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Okamura Y, Nomoto S, Hayashi M, et al:
Identification of the bleomycin hydrolase gene as a methylated
tumor suppressor gene in hepatocellular carcinoma using a novel
triple-combination array method. Cancer Lett. 312:150–157. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Towne CF, York IA, Watkin LB, Lazo JS and
Rock KL: Analysis of the role of bleomycin hydrolase in antigen
presentation and the generation of CD8 T cell responses. J Immunol.
178:6923–6930. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Suszynska J, Tisonczyk J, Lee HG, Smith MA
and Jakubowski H: Reduced homocysteine-thiolactonase activity in
Alzheimer’s disease. J Alzheimers Dis. 19:1177–1183.
2010.PubMed/NCBI
|
|
5
|
Lefterov IM, Koldamova RP and Lazo JS:
Human bleomycin hydrolase regulates the secretion of amyloid
precursor protein. FASEB J. 14:1837–1847. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zimny J, Sikora M, Guranowski A and
Jakubowski H: Protective mechanisms against homocysteine toxicity:
the role of bleomycin hydrolase. J Biol Chem. 281:22485–22492.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ratovitski T, Chighladze E, Waldron E,
Hirschhorn RR and Ross CA: Cysteine proteases bleomycin hydrolase
and cathepsin Z mediate N-terminal proteolysis and toxicity of
mutant huntingtin. J Biol Chem. 286:12578–12589. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kamata Y, Taniguchi A, Yamamoto M, Nomura
J, Ishihara K, Takahara H, Hibino T and Takeda A: Neutral cysteine
protease bleomycin hydrolase is essential for the breakdown of
deiminated filaggrin into amino acids. J Biol Chem.
284:12829–12836. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ferrando AA, Pendás AM, Llano E, Velasco
G, Lidereau R and López-Otín C: Gene characterization, promoter
analysis, and chromosomal localization of human bleomycin
hydrolase. J Biol Chem. 272:33298–33304. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ferrando AA, Velasco G, Campo E and
Lopez-Otin C: Cloning and expression analysis of human bleomycin
hydrolase, a cysteine proteinase involved in chemotherapy
resistance. Cancer Res. 56:1746–1750. 1996.PubMed/NCBI
|
|
11
|
Brömme D, Rossi AB, Smeekens SP, Anderson
DC and Payan DG: Human bleomycin hydrolase: molecular cloning,
sequencing, functional expression, and enzymatic characterization.
Biochemistry. 35:6706–6714. 1996.PubMed/NCBI
|
|
12
|
Montoya SE, Ferrell RE and Lazo JS:
Genomic structure and genetic mapping of the human neutral cysteine
protease bleomycin hydrolase. Cancer Res. 57:4191–4195.
1997.PubMed/NCBI
|
|
13
|
Chen J, Chen Y and He Q: Action of
bleomycin is affected by bleomycin hydrolase but not by caveolin-1.
Int J Oncol. 41:2245–2252. 2012.PubMed/NCBI
|
|
14
|
Lazo JS, Boland CJ and Schwartz PE:
Bleomycin hydrolase activity and cytotoxicity in human tumors.
Cancer Res. 42:4026–4031. 1982.PubMed/NCBI
|
|
15
|
Schwartz DR, Homanics GE, Hoyt DG, Klein
E, Abernethy J and Lazo JS: The neutral cysteine protease bleomycin
hydrolase is essential for epidermal integrity and bleomycin
resistance. Proc Natl Acad Sci USA. 96:4680–4685. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
De Haas EC, Zwart N, Meijer C, et al:
Variation in bleomycin hydrolase gene is associated with reduced
survival after chemotherapy for testicular germ cell cancer. J Clin
Oncol. 26:1817–1823. 2008.PubMed/NCBI
|
|
17
|
Timmer JC and Salvesen GS: Caspase
substrates. Cell Death Differ. 214:66–72. 2007. View Article : Google Scholar
|
|
18
|
Crawford ED and Wells JA: Caspase
substrates and cellular remodeling. Annu Rev Biochem. 80:1055–1087.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xu G, Cirilli M, Huang Y, Rich RL, Myszka
DG and Wu H: Covalent inhibition revealed by the crystal structure
of the caspase-8/p35 complex. Nature. 410:494–497. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Koldamova RP, Lefterov IM, DiSabella MT
and Lazo JS: An evolutionarily conserved cysteine protease, human
bleomycin hydrolase, binds to the human homologue of
ubiquitin-conjugating enzyme 9. Mol Pharmacol. 54:954–961.
1998.PubMed/NCBI
|
|
21
|
Schwanhäusser B, Busse D, Li N, Dittmar G,
Schuchhardt J, Wolf J, et al: Global quantification of mammalian
gene expression control. Nature. 473:337–342. 2011.
|
|
22
|
Pei Z, Calmels TP, Creutz CE and Sebti SM:
Yeast cysteine proteinase gene ycp1 induces resistance to bleomycin
in mammalian cells. Mol Pharmacol. 48:676–681. 1995.PubMed/NCBI
|
|
23
|
Montoya SE, Thiels E, Card JP and Lazo JS:
Astrogliosis and behavioral changes in mice lacking the neutral
cysteine protease bleomycin hydrolase. Neuroscience. 146:890–900.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chaitanya GV, Steven AJ and Babu PP:
PARP-1 cleavage fragments: signatures of cell-death proteases in
neurodegeneration. Cell Commun Signal. 8:312010. View Article : Google Scholar : PubMed/NCBI
|