Introduction
Glioblastoma (GBM) is the highest grade glioma and
is the most common malignant primary brain tumor in adults.
Although there have been advances in therapy, the median survival
time of patients with GBM is only 15 months (1–3).
Furthermore, while our understanding of the molecular and
physiological features of GBM has improved, no effective treatments
for this type of cancer have been identified (4). Therefore, there is a critical need for
new molecular targets, concepts, and approaches for treating this
devastating disease. Due to the diversity and heterogeneity of GBM
(5), targeting only a single factor
would not result in a satisfactory clinical outcome. Thus, it would
be necessary to regulate several oncogenes simultaneously.
microRNAs (miRNAs) are small, non-coding RNAs ~18–24
nucleotides in length that negatively regulate gene expression at
the post-transcriptional and/or translational level by binding
loosely complementary sequences in the 3′ untranslated regions (3′
UTRs) of target mRNAs (6). miRNAs
regulate the expression of multiple target genes and participate in
processes involved in the molecular pathology of cancer (7). Based on the concrete function and
genes targeted, miRNAs can act as oncogenes or tumor suppressors
(8,9). One example of a tumor suppressor miRNA
is miR-7, whose expression is frequently found decreased in GBM,
while its overexpression reduces cell proliferation, survival and
invasiveness in cultured glioma cells (10,11).
In contrast, miR-21 is almost invariably overexpressed in GBM and a
number of other tumor types, resulting in enhanced cell motility,
migration, and decreased apoptosis (12–14).
miRNA profiling further reveals anomalous levels of multiple miRNAs
in a wide variety of tumor types, suggesting that dysregulation of
the complex network of miRNA-mRNA interactions provides growth
advantages to tumor cells (15). In
addition, many miRNAs have been demonstrated to function as tumor
suppressors or oncogenes in glioma.
Previous studies have found that miR-708 plays
different roles in several types of cancers. One example is in
renal cancer cells, where miR-708 plays a role in inducing
apoptosis and suppressing tumorigenicity, but in lung
adenocarcinoma, increased miR-708 expression is associated with
poor prognosis (16,17). Therefore, the concrete function of
miR-708 is associated with the cell type. Due to the inconsistent
function of miR-708 in different types of cancers, studying the
concrete function of miR-708 in GBM has become increasingly of
interest to researchers, including ourselves. In our study, miR-708
in GBM cell lines was found to be decreased when compared with that
in normal brain tissue. Therefore, we hypothesized that miR-708 may
act as a tumor suppressor in GBM cell lines. In the present study,
we analyzed the function of miR-708 in human glioma cell lines. We
found that ectopic expression of miR-708 in A172 and T98G glioma
cells caused decreased proliferation, migration and invasion, with
accompanying low expression of several critical factors such as
Akt1, CCND1, MMP2, EZH2, Parp-1 and Bcl2, which play important
roles in the process of proliferation, invasion, and
anti-apoptosis. Our results identify a critical role for miR-708 in
the regulation of tumor progression in GBM, suggesting that miR-708
acts as a tumor suppressor gene for GBM.
Materials and methods
Tissue samples and cell culture
Human brain samples from epilepsy patients were
obtained from Renji Hospital, Shanghai Jiaotong University School
of Medicine, Shanghai, China. The hospital institutional review
board approved this study, and written informed consent was
obtained from all patients. Human A172, T98G, U87 and U251 GBM
cells were obtained from the ATCC (Manassas, VA, USA). The cells
were maintained in Dulbecco’s modified Eagle’s medium (DMEM)
supplemented with 10% fetal bovine serum (FBS) (both from
Gibco-BRL, Los Angeles, CA, USA) and were incubated at 37°C in 5%
CO2.
Plasmid construction and
transfection
HEK293T cells were seeded in 60-mm2
plates (4×106 cells/plate) in DMEM with 10%
heat-inactivated FBS 1 day before transfection. Cells were
transfected with 5.2 μg pGIPZ-miR-708, along with 2.36 μg psPAX2
and 0.8 μg pMD2G plasmids using Lipofectamine 2000 in DMEM. After 6
h, the transfection media were removed and replaced with DMEM
containing 10% heat-inactivated FBS without
penicillin-streptomycin. Lentiviral particles were harvested at 72
h post-transfection. The A172 and T98G cells were transduced with 3
ml additional media containing viral particles and 6 μg/ml
Polybrene. After 12 h, the conditional media were removed and
replaced with DMEM containing 10% FBS.
To construct the miR-708 overexpression
(over-miR-708) vector, 2 oligonucleotides were synthesized: forward
primer, TCGAAAGGAGCTTACAATCTAGCTGGGGTGTGCTGT
CCCCCAGCTAGATGTAAGCTCCTTTTTTT and reverse primer,
AATTAAAAAAAGGAGCTTACAATCTAGCTGG
GGGACAGCACACCCCAGCTAGATTGTAAGCTCCTT. The oligonucleotides were
annealed to be cloned into the EcoRI and XhoI sites
in the pGIPZ plasmid (Open Biosystems).
Real-time PCR
RNA was extracted from tissues or cells using TRIzol
(Invitrogen Life Technologies, Carlsbad, CA, USA). Real-time PCR
reactions were performed with TaqMan reverse transcription reagents
(Invitrogen Life Technologies) and SYBR-Green PCR Master Mix
(Applied Biosystems, Foster City, CA, USA) according to the
manufacturers’ protocols. Normalization was performed on U6 miRNA
levels. In addition, GAPDH mRNA was used as an internal control to
the normalized selected genes.
Western blotting
Western blotting was performed as previously
described (18), using primary
antibodies against Akt1, MMP2, EZH2 (1:1,000 dilution; Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA), CCND1, GAPDH, PARP-1 and
Bcl2 (1:1,000 dilution; Cell Signaling Technology, Inc., Danvers,
MA, USA), and incubated with a horseradish peroxidase-conjugated
anti-rabbit or anti-mouse antibody (1:1,000 dilution,
Sigma-Aldrich, St. Louis, MO, USA) for 2 h at room temperature.
Cell growth analysis
The cells were plated in 6-well culture plates
(5×104 cells/well), and incubated in a humidified
atmosphere of 5% CO2 at 37°C. Cell growth was monitored
by counting cell numbers at various time intervals. Three
independent experiments were carried out in triplicate.
Cell cycle analysis
The cell cycle distribution was analyzed by flow
cytometry (FCM). The cells were harvested by 0.05% trypsinization,
washed with phosphate-buffered saline (PBS), fixed with 75% ethanol
overnight at 4°C, and then incubated with RNase at 37°C for 30 min.
The nuclei were examined in a FACSCalibur flow cytometer, and DNA
histograms were analyzed by Modifit software (both from
Becton-Dickinson, Franklin Lakes, NJ, USA). The experiments were
performed in triplicate.
Apoptosis assay
The Annexin V-FITC Apoptosis Detection Kit I (Abcam,
Cambridge, MA, USA) was used to detect and qualify apoptosis by FCM
according to the manufacturer’s protocol. The cells were washed
with cold PBS, resuspended at a density of 1×106
cells/ml in 1X binding buffer, stained with FITC-labeled Annexin V
for 5 min, and immediately analyzed by the FACScan flow cytometer.
The data were analyzed by Cell Quest software
(Becton-Dickinson).
BrdU cell proliferation assay
Cell proliferation was assessed by
5-bromo-2′-deoxy-uridine (BrdU) incorporation using a Cell
Proliferation ELISA, BrdU kit (Roche Applied Science, Indianapolis,
IN, USA) according to the manufacturer’s protocol. The cells were
seeded onto a 96-well plate (1×105 cells/well) in 100 μl
culture medium and incubated at 37°C for 12, 24, 36 and 48 h. BrdU
labeling solution was added to a final concentration of 10 μM, and
the cells were incubated for an additional 2–4 h at 37°C. The
medium was removed and FixDenat (200 μl/well; Roche Applied
Science) was added to the cells and incubated for 30 min at 25°C.
The FixDenat solution was removed completely and 100 μl/well
anti-BrdU-POD working solution was added and incubated for 90 min
at 25°C. The antibody conjugate was then removed by flicking, and
the wells were washed 3 times with washing solution (200 μl/well).
Substrate solution (100 μl/well) was added and the cells were
incubated at 25°C until color development was sufficient for
photometric detection (after 12, 24, 36 and 48 h). The absorbance
(optical density) at 450 nm was measured using a microplate
reader.
Transwell invasion assay
The cells were detached and resuspended in
serum-free medium before being plated (5×105 cells/well)
into Matrigel-coated invasion chambers (Becton-Dickinson) and
allowed to invade for 16 h. The invading cells on the lower surface
of the chambers were fixed with 70% ethanol and then stained with
hematoxylin, while the remaining cells in the chambers were removed
using cotton swabs. The number of remaining cells was calculated by
counting five different fields under a microscope. The experiments
were performed in triplicate.
Statistical analysis
GraphPad Prism 5.0 was used for statistical
analysis. One-way analysis of variance (ANOVA) and Student’s
t-tests were used to analyze the significance between groups. We
considered P<0.05 to indicate statistical significance. The
means ± standard deviation of each group were calculated for all
experiments.
Results
miR-708 is downregulated in GBM cell
lines
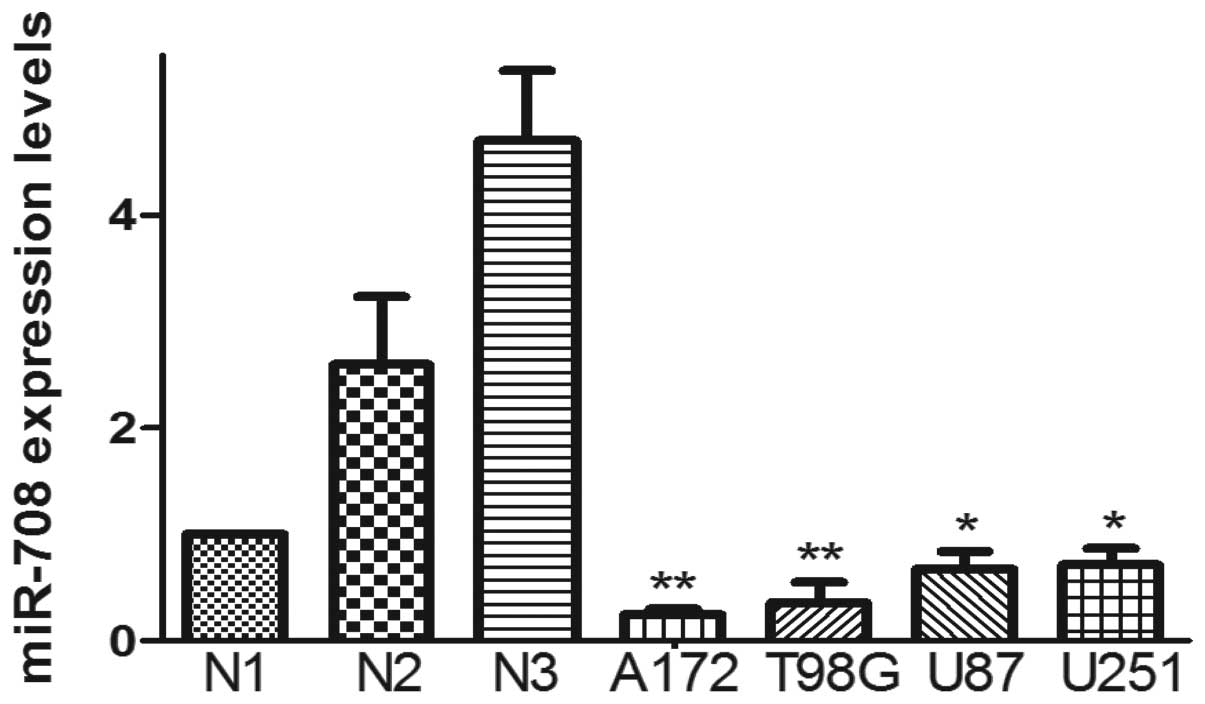
The expression of miR-708 in normal brain tissue and
GBM cell lines was quantified by real-time PCR. In the present
study, we extracted total RNA from the brain tissue of 2 epilepsy
patients and 4 GBM cell lines (U87, U251, A172, T98G). Compared to
the brain tissue, the expression of miR-708 in the 4 GBM cell lines
was low (Fig. 1).
Overexpression of miR-708 inhibits GBM
cell proliferation in vitro
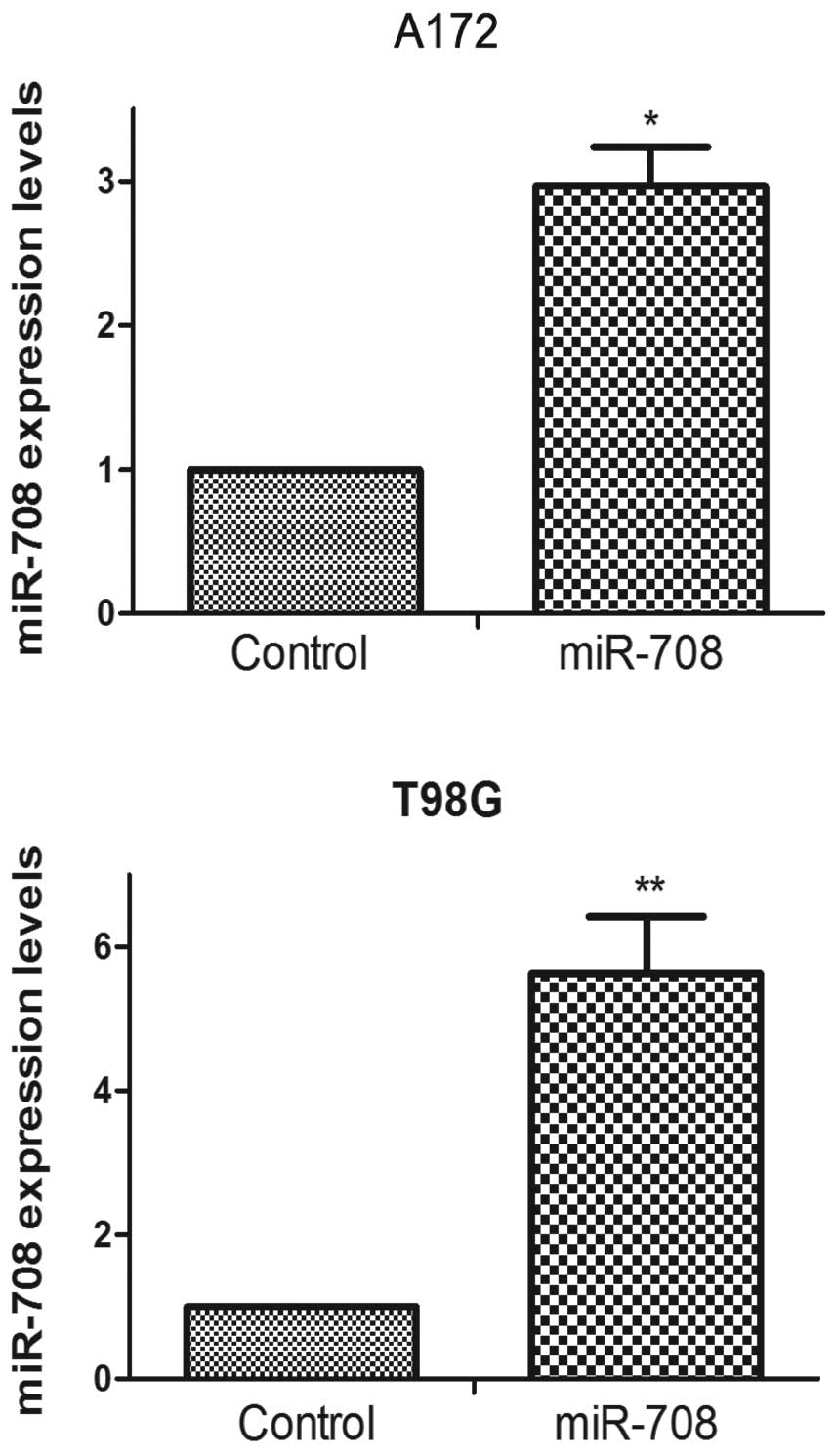
To explore the role of miR-708 in cell
proliferation, we carried out the BrdU cell proliferation and cell
counting assays for 6 days. A miR-708 oligonucleotide was used to
overexpress miR-708 in the A172 and T98G cells (Fig. 2). As shown in Fig. 3A and B, the overexpression of
miR-708 led to poor proliferative ability in the GBM cell lines
compared to the control group cells. Both the BrdU cell
proliferation assay and the cell counting assay demonstrated that
the overexpression of miR-708 in the A172 and T98G cells reduced
their proliferative ability. The cell cycle distribution of the
control and transfected cells was analyzed by FCM. In the A172 and
T98G cells, miR-708 overexpression caused G0/G1 arrest (Fig. 4). These observations suggest that
miR-708 overexpression suppresses the ability of GBM cells to
proliferate in vitro.
miR-708 overexpression suppresses the
invasive ability of GBM cells
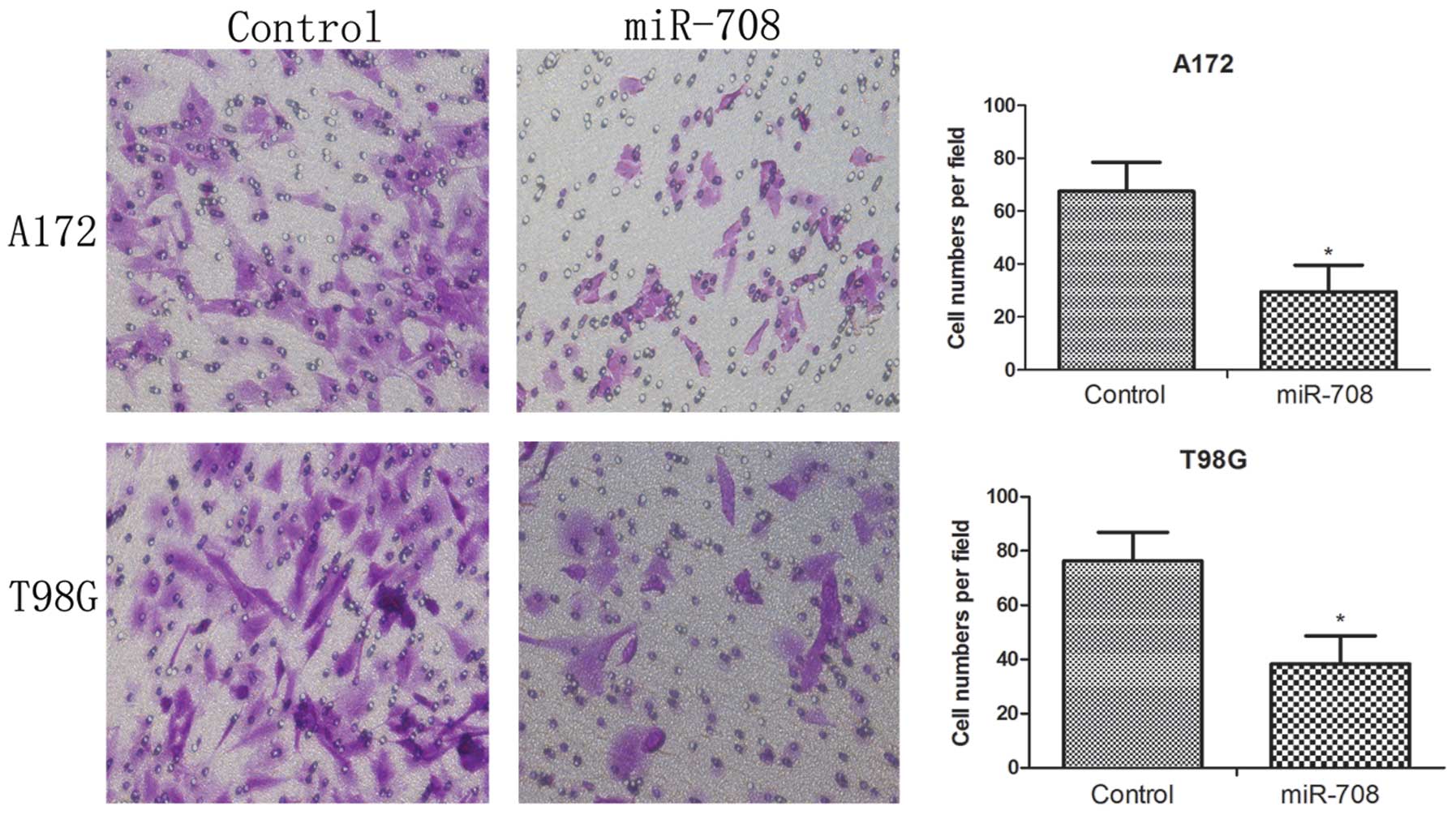
The Transwell invasion assay was used to evaluate
the impact of miR-708 expression on GBM cell invasion. Equal
numbers of A172 control, A172 miR-708 overexpression cells, T98G
control and T98G overexpression cells were placed in a Transwell
chamber. After 16 h, the cells that passed through the membrane
were counted under a microscope. As shown in Fig. 5, the over-miR-708 cells exhibited
poor invasive ability (P<0.05). These data suggest that miR-708
overexpression suppresses the invasive ability of GBM cells.
miR-708 overexpression induces apoptosis
of GBM cells
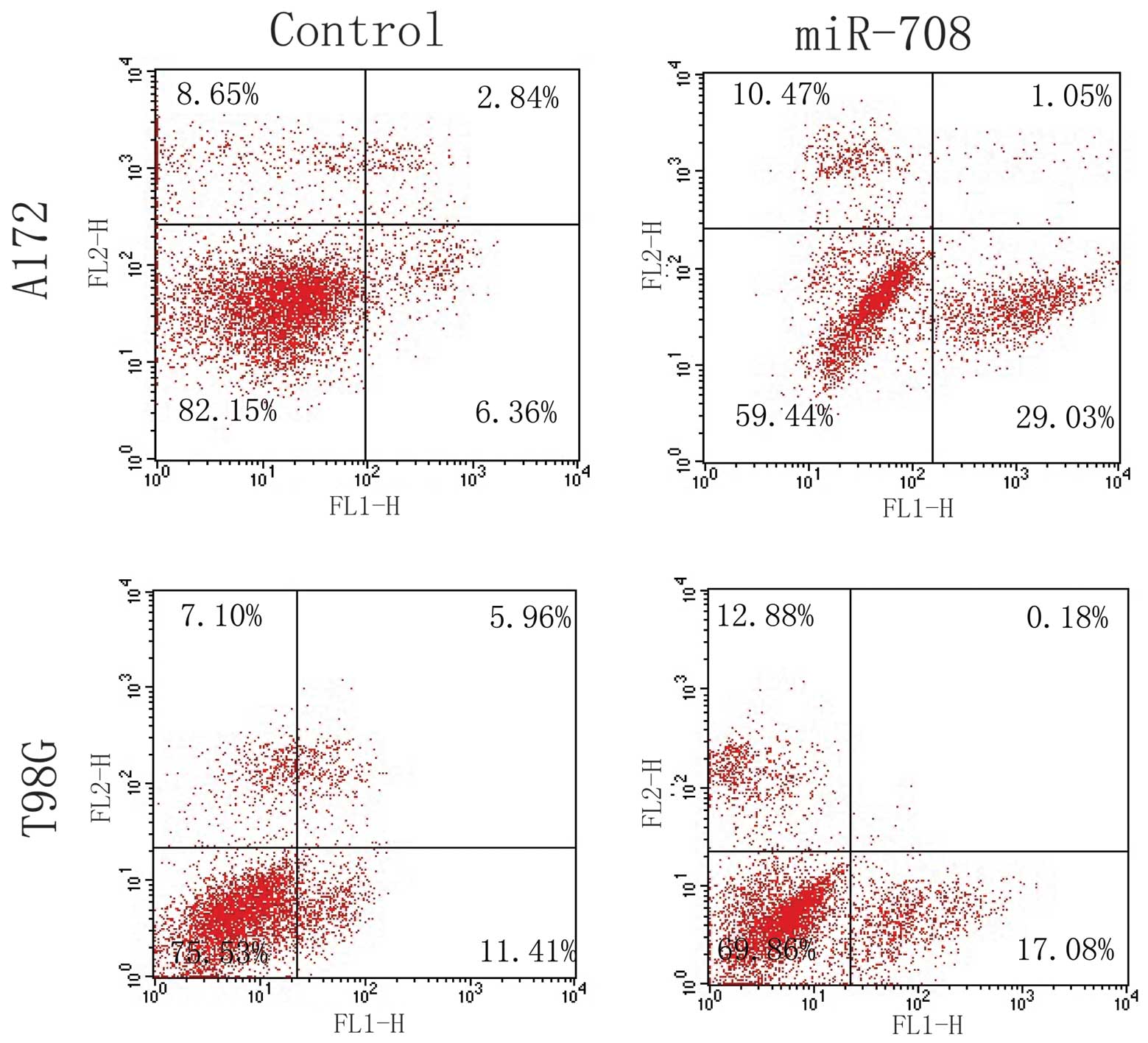
We used FCM to assess the effect of miR-708
expression on GBM cell apoptosis. As shown in Fig. 6, statistically significant increases
in Annexin V+ apoptotic cells were observed in the
over-miR-708 groups. This suggests that miR-708 induces GBM cell
apoptosis.
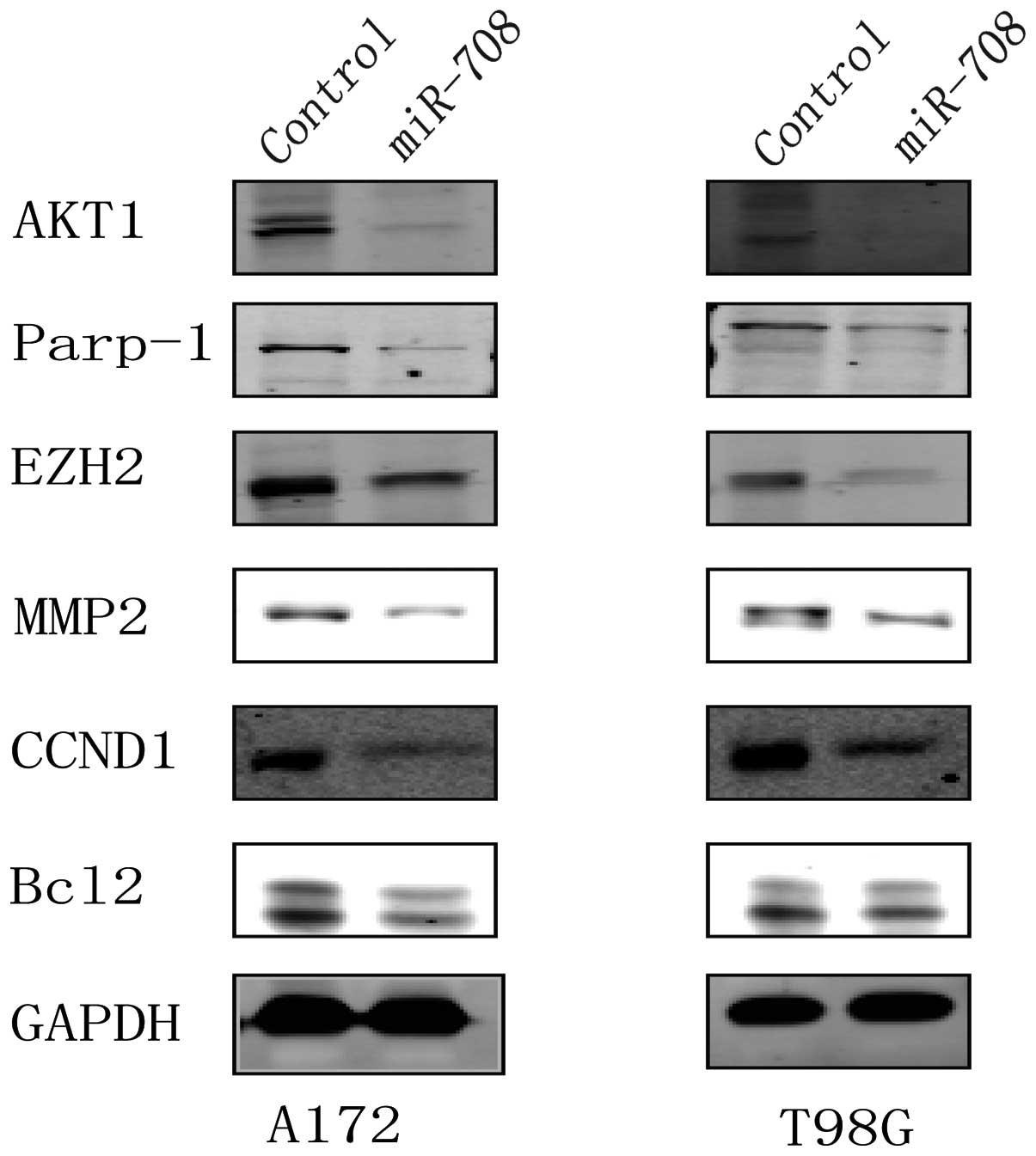
miR-708 overexpression regulates the
expression of components of proliferation, invasion and apoptosis
factors
To investigate the potential downstream effectors of
miR-708, we examined several signaling molecules that contribute to
the proliferation, invasion and anti-apoptosis ability of various
types of cancers. These included Akt1, CCND1, MMP2, EZH2, Parp-1
and Bcl2. For example, Akt1 and CCND1 play an important role in the
proliferation of cancer cells (19,20);
EZH2 and MMP2 contribute to the invasion of GBM cells (21–23);
and Parp-1 and Bcl2 act as anti-apoptosis factors for several types
of cells (24,25). As shown in Fig. 7, the expression of Akt1, CCND1,
MMP2, EZH2, Parp-1 and Bcl2 in the over-miR-708 cells was reduced
when compared to the levels in the control group cells.
Discussion
In the present study, we identified an important
tumor-suppressive miRNA, miR-708, that plays a critical role in the
process of GBM. Compared to the brain tissue from epilepsy
patients, the GBM cell lines A172, T98G, U87 and U251 contained low
amounts of miR-708. The ectopic expression of miR-708 decreased the
proliferation and invasion of GBM cells and induced apoptosis in
the cells. In other words, the expression level of miR-708 may
negatively regulate glioma malignancy. These data suggest that
miR-708 plays a critical role in the glioma malignant phenotype and
it may function as an antitumor factor in glioma cells. Several
studies have reported on the dysregulation of various miRNAs in
GBM, such as let-7, miR-21, miR-221/222 and miR-10b (11,13,26–28).
However, this is the first report that implicates miR-708 in the
process of GBM.
Our studies indicate that miR-708 modulates the
expression profiles of multiple genes involved in cell
proliferation, invasion and apoptosis. Moreover, these genes are
aberrantly expressed in cancer. Akt1 and CCND1 play a critical role
in the regulation of cell proliferation, particularly Akt1, which
can increase cell proliferation through the Akt1 pathway (19,20).
CCND1 can also affect cell proliferation through regulation of the
cell cycle (29,30). Dysregulation of CCND1 can also be
correlated with the aggressive behavior of a tumor. Lebeau et
al(30) reported that CCND1
regulates glioma cell proliferation by controlling progression to
the G1 phase of the cell cycle. In addition, our data showed that
miR-708 overexpression induced G0/G1 arrest, subsequently reducing
the proliferative ability of GBM cells. The effects of Akt1 and
CCND1 provide strong evidence for miR-708 as a tumor suppressor
that regulates cell proliferation in GBM cells.
Targeting the invasive ability of cells is an
important treatment strategy for different types of tumors
(31), including GBM. Aberrant
expression of many genes, which act as promoters of cell invasion,
are observed in cancer. MMP2 is one of the key enzymes involved in
the degradation of the extracellular matrix, and its high
expression is associated with tumor cell invasion (23,32).
In addition, EZH2 plays a role in increasing the invasive ability
of several tumor cell types (33,34).
Our data showed that MMP2 and EZH2 were reduced when miR-708 was
overexpressed in GBM cells. The regulation of MMP2 and EZH2 also
provides support for miR-708 as a tumor suppressor that targets
cell invasion.
Apoptosis is a well-orchestrated cellular mechanism
that balances cell proliferation and cell death (35). The ability to evade apoptosis is a
hallmark of tumorigenesis. As prior studies have demonstrated that
Parp-1 and Bcl2 are anti-apoptotic proteins, the increased
expression of these genes could contribute to tumor progression
(24,25). In our study, the expression of
Parp-1 and Bcl2 in the over-miR-708 GBM cells was low in contrast
to the control group cells. These data add additional credibility
to the tumor suppressor role of miR-708 in GBM.
In addition, Saini et al(16) reported that miR-708 plays a
tumor-suppressive role in renal cancer cells, while Jang et
al(17) confirmed that miR-708
acts as an oncogene contributing to tumor growth and disease
progression by directly downregulating TMEM88 in lung cancer. This
phenomenon is plausible because of the biodiversity of tumors,
indicating that the concrete function of miR-708 depends on the
cell type.
In conclusion, we demonstrated that the
overexpression of miR-708 in the human GBM cell lines A172 and T98G
significantly affected cell proliferation, invasion and apoptosis.
Our data suggest that these biological effects may be related to
the modulated expression of Akt1, CCND1, MMP2, EZH2, Parp-1 and
Bcl2. However, the exact mechanism of miR-708 regulation in these
genes requires further study. The effect of GBM cell proliferation,
invasion, and apoptosis by miR-708 suggests that it may act as a
tumor suppressor in human GBM. Furthermore, miR-708 may be an
attractive target for therapeutic intervention in GBM.
Acknowledgements
This study was supported by a project for building
hospital clinical key disciplines (no. RJ.4101307) and a grant from
the Shanghai government (no. 0952nm03900).
References
|
1
|
Stupp R, Mason WP, van den Bent MJ, et al:
Radiotherapy plus concomitant and adjuvant temozolomide for
glioblastoma. N Engl J Med. 352:987–996. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chin L and Gray JW: Translating insights
from the cancer genome into clinical practice. Nature. 452:553–563.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wiedemeyer R, Brennan C, Heffernan TP, et
al: Feedback circuit among INK4 tumor suppressors constrains human
glioblastoma development. Cancer Cell. 13:355–364. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dunn GP, Rinne ML, Wykosky J, et al:
Emerging insights into the molecular and cellular basis of
glioblastoma. Genes Dev. 26:756–784. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bonavia R, Inda MM, Cavenee WK and Furnari
FB: Heterogeneity maintenance in glioblastoma: a social network.
Cancer Res. 71:4055–4060. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bartel DP: MicroRNAs: genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sassen S, Miska EA and Caldas C: MicroRNA:
implications for cancer. Virchows Arch. 452:1–10. 2008. View Article : Google Scholar
|
|
8
|
Shenouda SK and Alahari SK: MicroRNA
function in cancer: oncogene or a tumor suppressor? Cancer
Metastasis Rev. 28:369–378. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen CZ: MicroRNAs as oncogenes and tumor
suppressors. N Engl J Med. 353:1768–1771. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lee ST, Chu K, Oh HJ, et al: Let-7
microRNA inhibits the proliferation of human glioblastoma cells. J
Neurooncol. 102:19–24. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kefas B, Godlewski J, Comeau L, et al:
microRNA-7 inhibits the epidermal growth factor receptor and the
Akt pathway and is down-regulated in glioblastoma. Cancer Res.
68:3566–3572. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Papagiannakopoulos T, Shapiro A and Kosik
KS: MicroRNA-21 targets a network of key tumor-suppressive pathways
in glioblastoma cells. Cancer Res. 68:8164–8172. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chan JA, Krichevsky AM and Kosik KS:
MicroRNA-21 is an antiapoptotic factor in human glioblastoma cells.
Cancer Res. 65:6029–6033. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gabriely G, Wurdinger T, Kesari S, et al:
MicroRNA 21 promotes glioma invasion by targeting matrix
metalloproteinase regulators. Mol Cell Biol. 28:5369–5380. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lu J, Getz G, Miska EA, et al: MicroRNA
expression profiles classify human cancers. Nature. 435:834–838.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Saini S, Yamamura S, Majid S, et al:
MicroRNA-708 induces apoptosis and suppresses tumorigenicity in
renal cancer cells. Cancer Res. 71:6208–6219. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jang JS, Jeon HS, Sun Z, et al: Increased
miR-708 expression in NSCLC and its association with poor survival
in lung adenocarcinoma from never smokers. Clin Cancer Res.
18:3658–3667. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Su CH, Zhao R, Zhang F, et al: 14–3–3sigma
exerts tumor-suppressor activity mediated by regulation of COP1
stability. Cancer Res. 71:884–894. 2011.
|
|
19
|
Vivanco I and Sawyers CL: The
phosphatidylinositol 3-kinase AKT pathway in human cancer. Nat Rev
Cancer. 2:489–501. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sommer G, Dittmann J, Kuehnert J, et al:
The RNA-binding protein La contributes to cell proliferation and
CCND1 expression. Oncogene. 30:434–444. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Suvà ML, Riggi N, Janiszewska M, et al:
EZH2 is essential for glioblastoma cancer stem cell maintenance.
Cancer Res. 69:9211–9218. 2009.PubMed/NCBI
|
|
22
|
Orzan F, Pellegatta S, Poliani PL, et al:
Enhancer of Zeste 2 (EZH2) is up-regulated in malignant gliomas and
in glioma stem-like cells. Neuropathol Appl Neurobiol. 37:381–394.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Luo Y, Liang F and Zhang ZY: PRL1 promotes
cell migration and invasion by increasing MMP2 and MMP9 expression
through Src and ERK1/2 pathways. Biochemistry. 48:1838–1846. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sugo N, Niimi N, Aratani Y, Masutani M,
Suzuki H and Koyama H: Decreased PARP-1 levels accelerate embryonic
lethality but attenuate neuronal apoptosis in DNA polymerase
β-deficient mice. Biochem Biophys Res Commun. 354:656–661.
2007.PubMed/NCBI
|
|
25
|
Gross A, McDonnell JM and Korsmeyer SJ:
BCL-2 family members and the mitochondria in apoptosis. Genes Dev.
13:1899–1911. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang C, Zhang J, Hao J, et al: High level
of miR-221/222 confers increased cell invasion and poor prognosis
in glioma. J Transl Med. 10:1192012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang J, Han L, Ge Y, et al: miR-221/222
promote malignant progression of glioma through activation of the
Akt pathway. Int J Oncol. 36:913–920. 2010.PubMed/NCBI
|
|
28
|
Guessous F, Alvarado-Velez M,
Marcinkiewicz L, et al: Oncogenic effects of miR-10b in
glioblastoma stem cells. J Neurooncol. 112:153–163. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jain M, Kumar S, Upadhyay R, et al:
Influence of apoptosis (BCL2, FAS), cell cycle (CCND1) and growth
factor (EGF, EGFR) genetic polymorphisms on survival outcome: an
exploratory study in squamous cell esophageal cancer. Cancer Biol
Ther. 6:1553–1558. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lebeau A, Unholzer A, Amann G, et al:
EGFR, HER-2/neu, cyclin D1, p21 and p53 in correlation to cell
proliferation and steroid hormone receptor status in ductal
carcinoma in situ of the breast. Breast Cancer Res Treat.
79:187–198. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Livant DL: Targeting invasion induction as
a therapeutic strategy for the treatment of cancer. Curr Cancer
Drug Targets. 5:489–503. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Westermarck J and Kähäri VM: Regulation of
matrix metalloproteinase expression in tumor invasion. FASEB J.
13:781–792. 1999.PubMed/NCBI
|
|
33
|
Ren G, Baritaki S, Marathe H, et al:
Polycomb protein EZH2 regulates tumor invasion via the
transcriptional repression of the metastasis suppressor RKIP in
breast and prostate cancer. Cancer Res. 72:3091–3104. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Rao ZY, Cai MY, Yang GF, et al: EZH2
supports ovarian carcinoma cell invasion and/or metastasis via
regulation of TGF-β1 and is a predictor of outcome in ovarian
carcinoma patients. Carcinogenesis. 31:1576–1583. 2010.
|
|
35
|
Subramanian S and Steer CJ: MicroRNAs as
gatekeepers of apoptosis. J Cell Physiol. 223:289–298.
2010.PubMed/NCBI
|