Introduction
Gallbladder carcinoma is the most common primary
malignancy of the biliary system; it is the fifth most common
malignancy of the gastrointestinal tract (GIT). It is characterized
by very high invasion and is associated with poor prognosis.
Patients with gallbladder carcinoma usually have advanced disease
at the time of diagnosis, except for a subset of patients who are
diagnosed incidentally at the time of elective cholecystectomy.
Despite advances in diagnosis and treatment of gallbladder
carcinoma, long-term survival remains dismal. Indeed, chemotherapy
and radiotherapy are ineffective as primary treatments, and
resection remains the only chance for cure. However, only a
minority of patients are candidates for resection at the time of
diagnosis. Even after curative resection, most series quote a
long-term survival of only 5–12% (1–3). Thus
alternative treatment approaches are required, for example,
intervention at the molecular level. However, the underlying
mechanisms of tumor initiation, progression and metastasis of
gallbladder carcinoma are still not fully understood.
In recent years, molecular biology studies have
found genes related to gallbladder carcinoma including
c-myc, B-cell lymphoma/leukemia-2 (Bcl-2),
P53, P16 and Survivin(4–8). The
Bcl-2 gene is one of the most important regulatory factors
in cell apoptosis. It can extend cell survival by inhibiting the
apoptotic process. Moreover, studies have found that Bcl-2
gene expression is very high in malignant tumor tissues including
those derived from gallbladder carcinoma (9–11).
These findings indicate that the Bcl-2 gene plays an
important role in the initiation and progression of gallbladder
carcinoma. Therefore, targeting the Bcl-2 gene may lead to
effective treatment for gallbladder carcinoma.
The relatively recent development of RNA
interference (RNAi) technology, which has a strong effect on
post-transcriptional gene silencing, has been widely used to target
oncogenes and inhibit cancer growth (12,13).
In the present study, we constructed eukaryotic vectors bearing
small interference RNA (siRNA) sequences targeting the Bcl-2
gene. First, the constructs were transfected into the human
gallbladder carcinoma GBC-SD cell line and stable transfectants
were selected for investigation. We observed that silencing of
Bcl-2 resulted in the growth inhibition of gallbladder carcinoma
cells and sensitization to chemical drugs, through in vitro
and in vivo experiments. Thus, we provide a basis for the
treatment of gallbladder carcinoma with Bcl-2 genetic RNAi
techniques.
Materials and methods
Cell line and culture
The human gallbladder carcinoma cell line, GBC-SD,
was maintained in the central laboratory of the Medical College of
Xi’an Jiaotong University. Cells were cultured in RPMI-1640 medium
supplemented with 10% fetal bovine serum (FBS; both from Gibco-BRL,
USA) under saturated humidity conditions at 37°C in 5%
CO2. Cells were passaged at a 1:2 ratio when the
attached cell density reached ~80 to 90%.
Animals
All animal experiments were reviewed and approved by
the Ethics Committee of the First Affiliated Hospital of the
Medical College, Xi’an Jiaotong University. BALB/c nude mice, 4–6
weeks of age (weighing 18–22 g) were provided by the laboratory
animal center of the Fourth Military Medical University and bred in
an SPF grade animal experimental center of the Xi’an Jiaotong
University at 25°C and 60–70% humidity. They were fed a standard
rodent diet and water.
Design of the specific siRNA targeting
Bcl-2
According to the Bcl-2 mRNA sequence in
GenBank and using siRNA Design Software provided by Invitrogen
(http://www.invitrogen.com), we chose a
target sequence according to the principle of siRNA target sequence
design. We then further verified the sequence through the BLAST
program from the NCBI (http://www.ncbi.nlm.nih.gov) website. The siRNA
sequences that targeted the Bcl-2 gene were as follows: 515S
sense, 5′-GATCCGCATC
GCCCTGTGGATGACTTTCAAGAGAAGTCATCCACAG
GGCGATGTTTTTTGGAAA-3′; 515A antisense, 5′-AGC
TTTTCCAAAAAACATCGCCCTGTGGATGACTTCTCT
TGAAAGTCATCCACAGGGCGATGCG-3′; ConS
sense, 5′-GATCCACTACCGTTGTTATAGGTGTTCAAGAGACA
CCTATAACAACGGTAGTTTTTTTGGAAA-3′; ConA antisense,
5′-AGCTTTTCCAAAAAAACTACCGTTGTTATAG
GTGTCTCTTGAACACCTATAACAACGGTAGTG-3′. The
515S and 515A are sense and antisense strands of DNA containing the
interference point sites. ConS and ConA are sense and antisense
strands of DNA containing the negative control. The underlined
sections in 515S and 515A, ConS and ConA are consistent or opposite
complementary sequences with the target gene mRNA and negative
sequence, respectively. GAT, GGAAA is the enzyme site, CCG is the
recognition site of RNA polymerase III, TTCAAGAGA forms the
structure of stem ring, and TTTTTT is the termination signal of
transcription. These sequences were connected and reconstructed
within the pSilencer™-EGFP vector (Department of Biochemistry and
Molecular Biology of the Fourth Military Medical University), then
transfected and integrated into the eukaryotic cell genome.
Finally, shRNAs were transcribed out from the initiation site
recognized by the RNA polymerase III.
Construction and identification of the
Bcl-2 siRNA eukaryotic expression vector
After annealing the synthetic oligonucleotides,
pSilencer™-EGFP was cut with BamHI and HindIII
enzymes (Gibco-BRL, USA). The plasmid was inoculated into
freeze-stored DH5α (Department of Biochemistry and Molecular
Biology of the Fourth Military Medical University) and then
cultivated on a plate, without antibiotics, at 37°C overnight. The
following day, monoclonal colonies were selected and inoculated in
10 ml of LB culture solution without antibiotics, and cultured at
37°C with agitation at 250 rpm overnight. The next day 1% of the
culture was transferred into 200 ml of non-antibiotic containing LB
culture solution and agitated for a further 2 h. The culture was
stopped when the solution absorbance at A260 was 0.4.
The bacterial culture was then placed on ice for 30 min, followed
by centrifugation at 5,000 rpm for 10 min at 4°C. The supernatant
was then removed and the cells were retrieved. The precipitate was
resuspended in TSS under aseptic conditions; it was then frozen at
−70°C until use. The recycling products and annealing products were
connected at 4°C overnight. The connection was added to 100 μl of
competent cells and shaken slightly, incubated on ice for 30 min,
then placed in a 42°C water bath for 90 sec, followed by cooling
for 2–3 min. Two hundred microliters of preheated no penicillin
(Amp) LB medium was then added to each tube, then agitated slowly
at 140–150 rpm for 45 min at 37°C to promote bacterial recovery.
The transformed competent cells were smeared onto an agar plate
that contained antibiotic until the liquid was completely absorbed.
Then, the plate was inverted and cultured for 12–16 h at 37°C until
the colonies appeared. Six colonies were randomly selected from
each group. The plasmid was extracted with the ‘small kit’ from
Shanghai HuaShun Company, according to the instructions.
Three microliters of extracted plasmid DNA was cut
with the BamHI and HindIII enzyme and cultured for 3
h at 37°C. The digested products were separated on a 1% agarose gel
by electrophoresis and then photographed under a UV
transilluminator. The positive clones identified by enzyme
digestion were sent to Beijing Augct DNA-Syn Biotechnology Co.,
Ltd. for sequence testing. Recombinant plasmids were named
pSilencer™-EGFP sh515 (experimental group) and pSilencer™-EGFP
shCon (negative control group).
Transfection of GBC-SD cells with Bcl-2
siRNA
GBC-SD cells in the logarithmic growth phase were
inoculated into a 6-well plate at a concentration of
2×105 cells/well. Transfection by a liposome-mediated
DNA method using Lipofectamine 2000 (Invitrogen, Gaithersburg, USA)
was performed 24–48 h when the culture confluence reached 80% in
accordance with the instruction. The experiment was divided into
two groups transfected with either pSilencer™-EGFP sh515
(experimental group) or pSilencer™-EGFP shCon (negative control
group).
The culture solution of transiently transfected
GBC-SD cells was replaced every 2–3 days. The cells began to die
after two days when treated with G418, whose screening
concentration was 400 μg/ml. Positive clones were observed in 2
weeks. Thereafter, the cloned cells did not appear to die in the
presence of G418 (400 μg/ml), but their growth was slow. A visible
cloning cell was formed in ~6–8 weeks. The monoclone was selected
for further expansion in culture. The experiment was divided into
the experimental group and the negative control group.
Reverse transcription-polymerase chain
reaction (RT-PCR)
GBC-SD human gallbladder carcinoma cells were
transiently transfected with pSilencer™-EGFP vector and the
negative control empty vector, and the cells of each group were
collected 12 h later for total RNA extraction using TRIzol reagent
(Invitrogen). Eight microliters of RNA was used to synthesize cDNA
and then subjected to PCR amplification. PCR primers were
synthesized by Beijing Augct DNA-Syn Biotechnology Co., Ltd. The
Bcl-2 primer sequences were as follows: upstream primer
5′-CTGGGAGAACAGGGTACGATAA-3′, downstream primer
5′-AGCCAGGAGAAATCAAACA GAG-3′, resulting in a PCR product of 210
bp. β-actin was amplified as a control using the following primer
sequences: upstream 5′-TGCGCAGAAAACAAGATGATT-3′ and downstream
5′-TGGGGGACAAAAAGGGGGAAGG-3′, resulting in a PCR product of 450 bp.
The PCR conditions were: one cycle of denaturing at 94°C for 30 sec
(first cycle was 94°C for 4 min), annealing at 60°C for 30 sec,
extension at 72°C for 30 sec, followed by 25 cycles. β-actin cDNA
was amplified at the same time as an internal standard control. The
PCR products were loaded onto a 1% agarose gel for electrophoresis.
The stably transfected GBC-SD cells were also tested for knockdown
of Bcl-2 with RT-PCR.
Western blotting
The GBC-SD cells were transfected with recombinant
plasmid vectors of the two groups and harvested 12 h later. The
total cell protein was extracted with the RIPA total protein kit
(Santa Cruz Biotechnology, Inc., USA) and quantified with the ABC
protein quantification method. The sample was separated by SDS-PAGE
(12%) and transferred onto a nitrocellulose membrane. The western
blot analysis method was used to detect Bcl-2 protein
expression using rabbit anti-human Bcl-2 polyclonal antibody
(AB1720; Chemicon, USA). Detection was performed with an enhanced
chemiluminescence agent. The expression of β-actin was tested as an
internal standard control. The stably transfected GBC-SD cells were
also tested for Bcl-2 protein expression using the same method.
Cell growth and proliferation assay
GBC-SD cells were transfected transiently with the
vector of each group and digested 12 h later. The cells were
inoculated into a 96-well plate at a concentration of
5×103 cells/well. A concentration of 10 μmol/l
Ponasterone A was added to each well as an inducer. A further 20 μl
of freshly prepared MTT (0.5 mg/ml) (Sigma, USA) was added to
different wells at 12, 24, 36, 48 and 60 h later, respectively. The
cells were cultured at 37°C for 4 h, then MTT liquid was removed
and 150 μl DMSO (Sigma, USA) was added to each well. Optical
density (OD) readings were obtained at 490 nm, from which the
inhibition rate of tumor cell growth was calculated using the
formula: Inhibition rate = (average absorbance (A) value of control
group - average A value of experimental group)/average A value of
control group × 100%.
Flow cytometric apoptosis assay
Apoptotic cells were determined using the Annexin
V/fluorescein isothiocyanate (FITC) Apoptosis Detection kit
(Jingmei Biotech Co., Shenzhen, China) and an EPICS XL-MCL flow
cytometer (Becton-Dickinson) according to the manufacturer’s
instructions. Briefly, GBC-SD cells, and cells stably transfected
with either GBC-SD-RNAi pSilencer™-EGFP shCon or GBC-SD-RNAi
pSilencer™-EGFP sh515 were collected and single cell suspensions
(1×106 cells) were prepared. Briefly, 1×106
cells were stained with Annexin V/FITC for 30 min at 4°C in the
dark and then stained with propidium iodide for 10 min before flow
cytometric analysis.
Chemotherapy drug sensitivity assay
The number of living cells was counted by the trypan
blue staining method and used to adjust the number of cells to
5×105/ml. The cells were inoculated into a 96-well plate
in a volume of 100 μl/well and cultured for 24 h in an incubator
with 5% CO2 and saturated humidity at 37°C. A volume of
25 μl of different drugs was added into the experimental wells: 1
μg/ml 5-fluorouracil (5-FU; Tianjin Jinyao Amino Acid Co., Ltd.),
0.3 μg/ml mitomycin C (MMC; Hebei QiYuan Pharmaceuticals Corp.),
0.04 μg/ml adriamycin (ADM; Zhejiang HaiZheng Pharmaceutical Co.,
Ltd.) and 0.3 μg/ml cisplatin (DDP; Shandong Luoxin Pharmaceutical
Co., Ltd.) (drug concentration was subjected to 1/10 of blood
concentration), while the same amount of PBS was added into the
control wells. The medium was removed after cultivation for 48 h,
and 25 μl of freshly prepared MTT solution (2 mg/ml) was added into
each well. The supernatant was removed after cultivation for 6 h at
37°C, and 150 μl of DMSO was added to each well. OD readings of
each well were measured at 570 nm when the purple crystals were
dissolved completely, with 630 nm as a reference wavelength. The
inhibition rate was calculated using the following formula:
Inhibition rate (%) = (1 - average OD value of the experimental
well/average OD value of the control well) × 100.
In vivo tumorigenicity assay
In vivo tumorigenicity assay
Twelve nude mice, 4–6 weeks of age, were randomly
divided into the Bcl-2 siRNA experimental and control groups
(n=6). For the experimental group, Bcl-2 siRNA stably
transfected GBC-SD suspensions of 6×106 cells in 0.2 ml
were subcutaneously injected into the left flank of nude mice. For
the control group, GBC-SD suspensions alone of 6×106
cells in 0.2 ml were injected into the left flank of nude mice.
Gene therapy studies
The human gallbladder carcinoma xenograft nude mouse
model was generated. Briefly, 18 BALB/c nude mice, 4–6 weeks of
age, were injected with a total number of 6×106 GBC-SD
cells/mouse into the right flank. The 18 mice were randomly divided
into three groups: pSilencer™-EGFP sh515 group (experimental
group), pSilencer™-EGFP shCon group (empty vector negative control
group) and the normal control group. Next, 10 μg recombinant DNA
plasmid pSilencer™-EGFP sh515 and pSilencer™-EGFP shCon (negative
control) were each mixed with 30 μl Lipofectamine 2000 liposome
(Invitrogen), and then they were injected into multiple sites of
peritumoral tissue of the mice every 2 days for a total of five
injections.
Observation of xenograft tumor
growth
The general condition of the nude mice was observed
every day, while tumor size was measured every 4 days. The tumor
size was calculated according to the formula: V = πa2b/6
(a, tumor short diameter; b, tumor long diameter) and the growth
rate was calculated according to the formula: Average growth rate =
mean tumor volume (mm)3/host with tumor time (days), and
tumor growth curves were drawn. Mice were sacrificed after 6 weeks,
and the tumors were removed and weighed. The liver, lung, spleen,
kidney and other organs were also removed. All were fixed in 10%
(volume fraction) formaldehyde solution, paraffin embedded, and
then cut into 4- to 5-μm sections for histological study. The
sections from the tumor were stained with Bcl-2 by
immunohistochemistry or with H&E staining, in order to
investigate tumor metastasis and side effects.
Statistical analysis
The data are expressed as mean ± standard deviation
(SD). Statistical significance was determined using the
χ2 test and Student’s t-test. A value of P<0.05 was
considered to indicate a statistically significant difference. SPSS
13.0 for Windows (SPSS, Inc., Chicago, IL, USA) was used for
statistical analyses. All statistical tests were two-tailed.
Results
DNA sequencing and identification of the
Bcl-2 siRNA recombinant plasmid vector
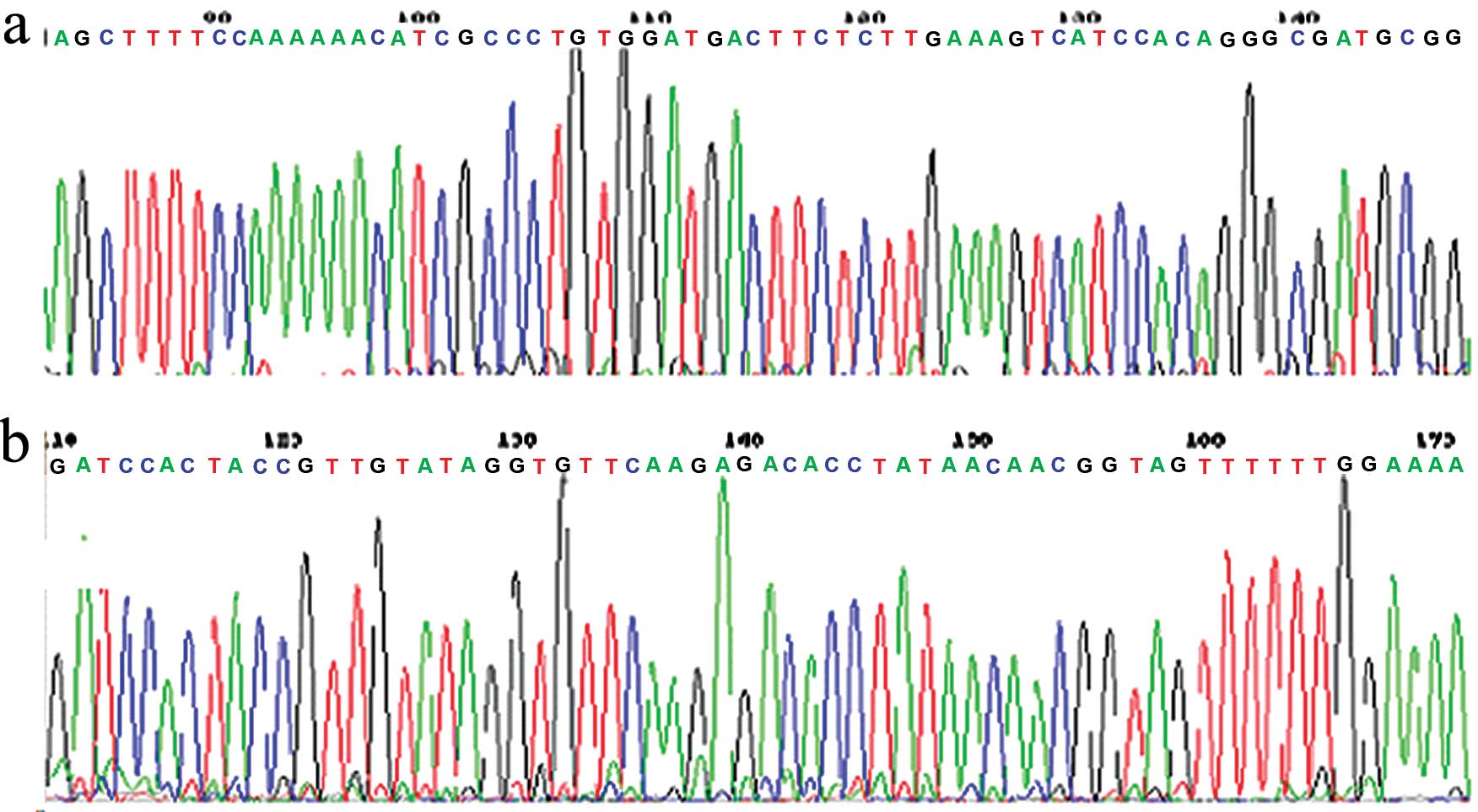
The results of DNA sequencing, from Beijing Augct
DNA-SYN Biotechnology Co., Ltd., showed that the sequence of the
vector was identical to the designed sequence, indicating that the
recombinant plasmid vector was constructed successfully (Fig. 1). The recombinant plasmid which
contained the shRNA coding sequence was named pSilencer™-EGFP sh515
(experimental group) and the empty vector was referred to
pSilencer™-EGFP shCon (negative control group). Both plasmids were
amplified for future study.
In vitro studies
Effect of Bcl-2 siRNA transfection
into GBC-SD cells
The expression of each plasmid was confirmed by
fluorescence microscopy 12 h after transfection of cells. Ten
fields of view were chosen in each group (experimental and
control), from which the number of green fluorescent-positive cells
and the total number of cells were calculated. The transfection
efficiency was calculated in accordance with the formula:
Transfection efficiency = (number of green fluorescent-positive
cells/total number of cells) × 100%. Experiments were repeated
three times. The result showed that there was no fluorescence
signal in pSilencer™-EGFP shCon (negative control group) after its
transient transfection into human gallbladder carcinoma GBC-SD
cells, but the transfection rate of pSilencer™-EGFP sh515
(experimental group) varied between 45 and 70% (Fig. 2a). After stable transfection with
pSilencer™-EGFP into GBC-SD cells, as before, the fluorescence test
showed no fluorescent signals in the negative control group, but
there was readily detectable fluorescence in the experimental
groups for which the transfection rate was nearly 100%. This
confirmed that the GBC-SD cells had been stably transfected with
pSilencer™-EGFP (Fig. 2b).
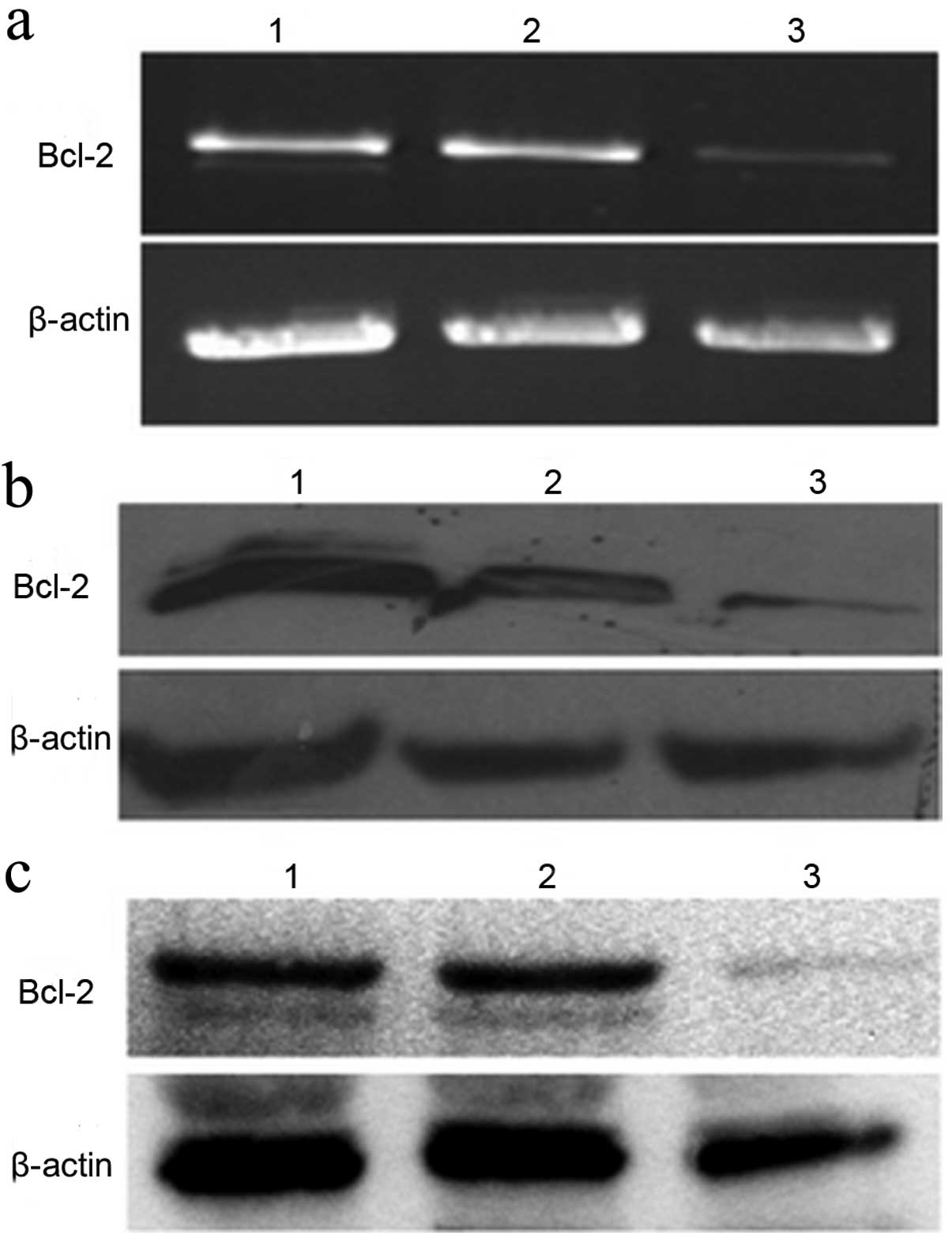
Bcl-2 siRNA inhibits Bcl-2 mRNA
expression
Twelve hours after transient transfection of GBC-SD
cells with the recombined pSilencer™-EGFP vector, or the negative
control empty vector, we observed significant inhibition of
Bcl-2 mRNA expression in the pSilencer™-EGFP sh515
experimental group when compared with those of the blank control
and pSilencer™-EGFP shCon groups (Fig.
3a).
Bcl-2 siRNA inhibits Bcl-2 protein
expression
Twelve hours after transient transfection of GBC-SD
cells with pSilencer™-EGFP sh515 or pSilencer™-EGFP shCon, protein
lysates were collected and tested by western blot analysis for
Bcl-2 expression. The results showed that Bcl-2
protein expression in the pSilencer™-EGFP sh515 experimental group
was significantly reduced when compared with that of the
pSilencer™-EGFP shCon negative control group or the non-transfected
(blank) group, consistent with RT-PCR results (Fig. 3b). Similar results were also
obtained using protein lysates prepared from the stably transfected
GBC-SD cells (Fig. 3c).
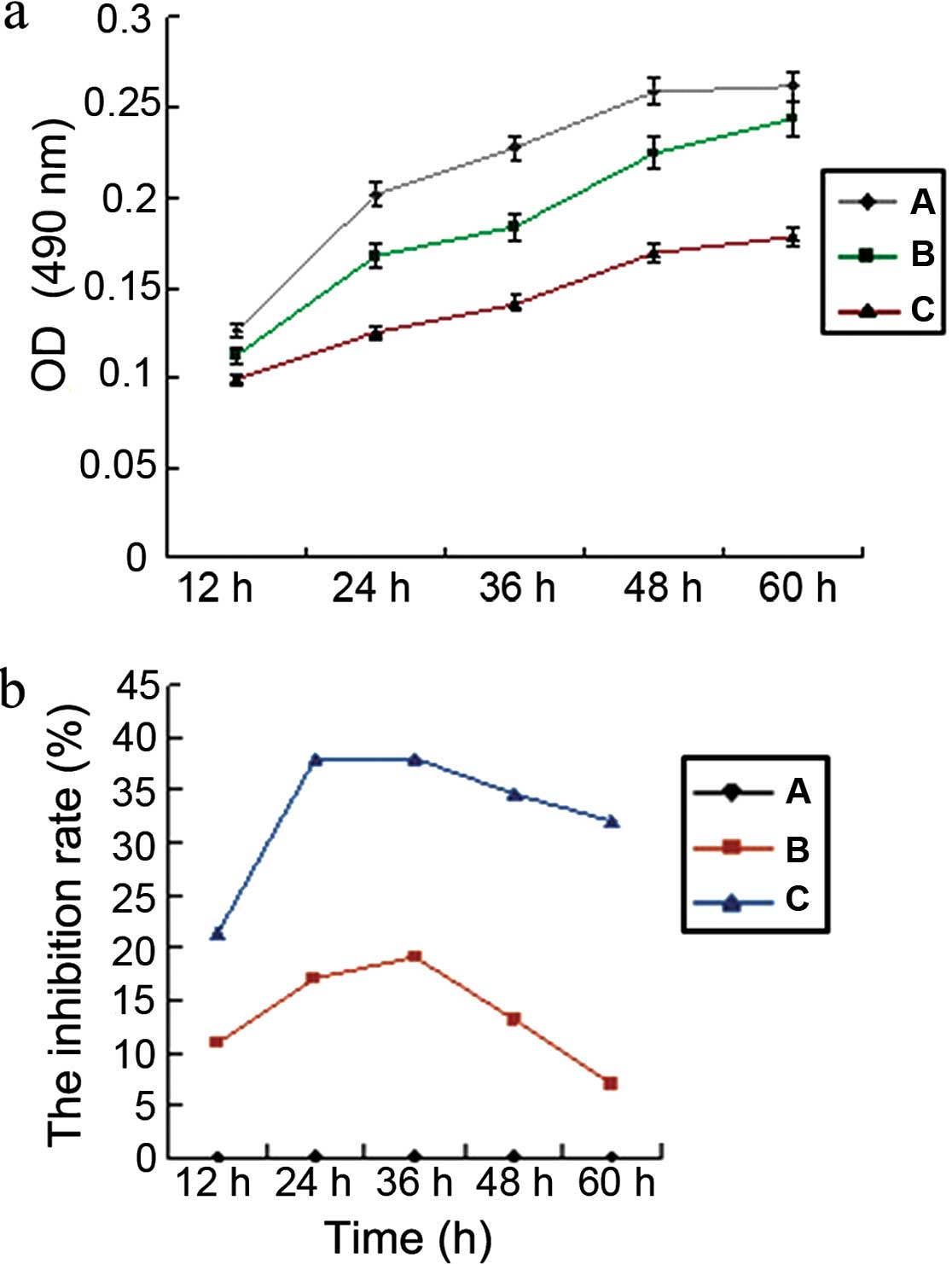
Bcl-2 siRNA inhibits cell growth and
proliferation in GBC-SD cells
Twelve hours after transfecting GBC-SD cells with
Bcl-2 siRNA, the activity of cells was detected every 12 h
using the MTT method. Data analysis revealed that cell
proliferation decreased significantly (P<0.05) after silencing
of Bcl-2, as determined by comparing the cell activity of
the pSilencer™-EGFP sh515 experimental group to either the negative
control group (pSilencer™-EGFP shCon) or the non-transfected blank
control group. There was no significant difference in the cellular
activity between the two latter groups (P>0.05) (Fig. 4). The results showed that
pSilencer™-EGFP sh515 silenced the Bcl-2 gene in the GBC-SD
human gallbladder carcinoma cells effectively and inhibited the
growth and proliferation of the tumor cells.
Bcl-2 siRNA induces GBC-SD cell
apoptosis
To further study the effect of Bcl-2 siRNA on
GBC-SD cell apoptosis, cells were stained with Annexin V-FITC and
propidium iodide. As shown in Fig.
5, the apoptotic percentage of GBC-SD/Bcl-2 siRNA cells
was 30.83±4.2%, which was significantly higher than that of the
GBC-SD/Bcl-2 negative control (4.3±1.3%) and GBC-SD cells
(3.6±1.1%) (P<0.05). This implies that inhibition of
Bcl-2 is able to induce apoptosis in gallbladder cancer
GBC-SD cells.
Bcl-2 siRNA increases sensitivity of
GBC-SD cells to chemotherapy drugs
Four types of chemotherapy drugs were diluted into
the following concentrations: 5-FU 1 μg/ml; MMC 0.3 μg/ml; ADM 0.04
μg/ml; DDP 0.3 μg/ml, and their inhibitory rates on cells with and
without silencing of Bcl-2 were calculated. The results
showed that the sensitivity of gallbladder carcinoma cells to the 4
drugs was increased to different levels after stable transfection
with Bcl-2 siRNA stably. However, only the results of the
5-FU group were of statistical significance (P<0.05), while
there were no significant differences between the other groups
(P>0.05) (Fig. 6).
In vivo studies
RNAi targeting Bcl-2 inhibits
tumorigenicity in vivo
Xenograft tumor growth
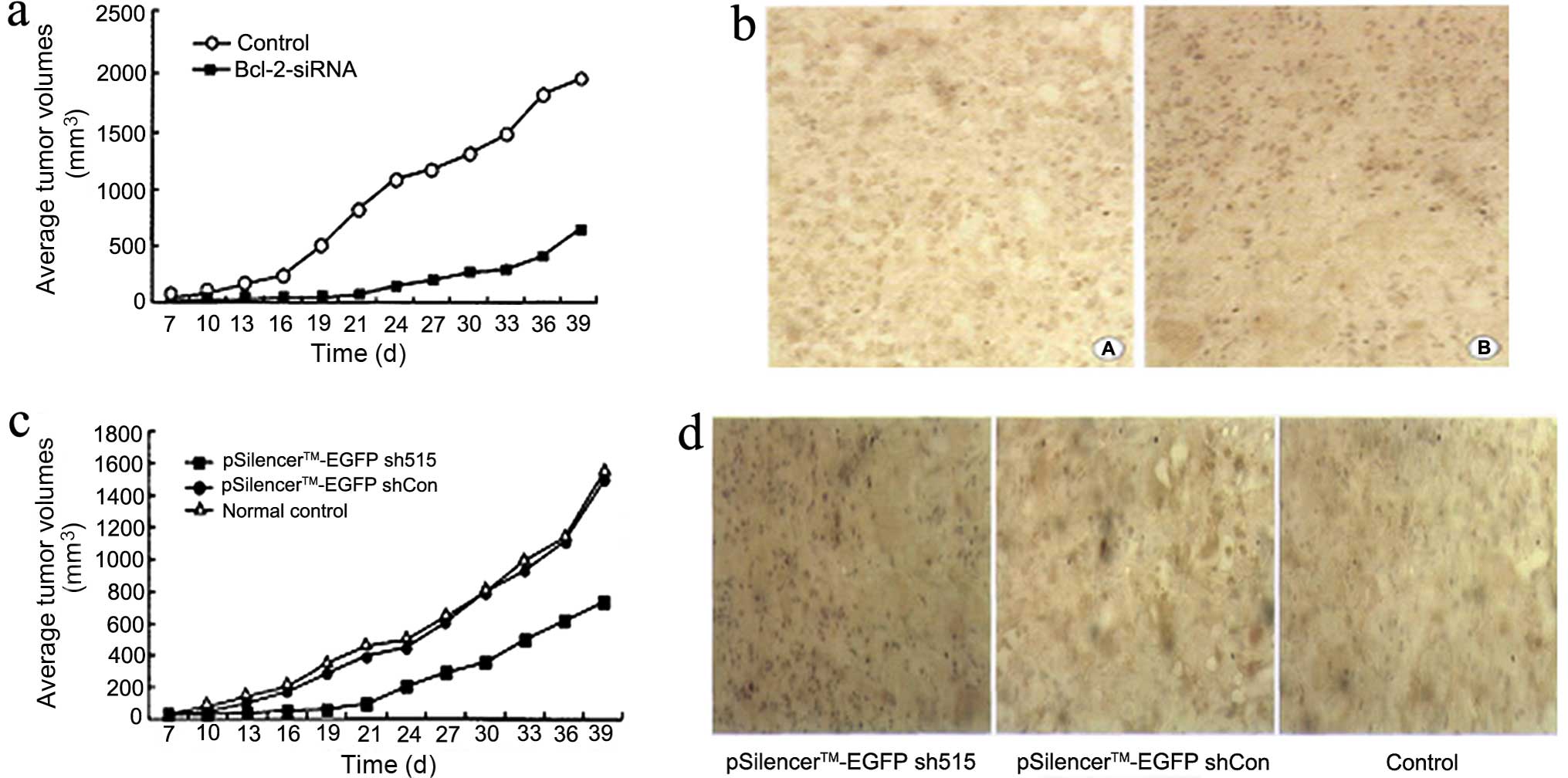
The tumorigenicity of the control group and
Bcl-2 siRNA experimental group were 100 and 60%,
respectively. The average volume of tumors was 1914.6±125.0 and
629.7±78.9 mm3 in the control and experimental group,
respectively. The average growth rate was 45.58 and 14.99% in the
control and experimental group, respectively. The average weight of
tumors was 2.24±0.33 and 0.77±0.12 g, in the control and
experimental group, respectively. Thus, the average volume, average
growth rate and average weight of tumors in the control group were
significantly greater than those of the Bcl-2 siRNA group
(P<0.05) (Fig 7a).
Histopathological changes
H&E staining of different organs from nude mice
showed necrosis of the tumors while the other organs were normal.
Bcl-2 immunohistochemical staining showed that the control
group expression was stronger with a positive expression rate of
50.4±1.3%. The Bcl-2 staining in the experimental group was
weaker than that in the control group with a positive expression
rate of 28.2±2.1%. This difference was significant between the two
groups (P<0.05) (Fig. 7b).
RNAi targeting Bcl-2 inhibits tumor
growth in vivo
Growth of xenograft tumor
The average volume, average growth rate and average
weight of experimental group tumors were significantly lower than
those determined for the empty vector negative control and normal
control groups (P<0.05). There was no significant difference
when the empty vector negative control group and the normal control
group were compared (P>0.05) (Fig.
7c).
Histopathological changes
H&E staining of different organs from nude mice
showed necrosis of the tumors while other organs were normal.
Bcl-2 immunohistochemical staining results showed that the
Bcl-2-positive expression rate in tumors of the empty vector
negative control group and the control group was 51.2±2.3 and
53.0±1.7%, respectively; there was no significant difference
between these two groups (P>0.05). The Bcl-2 staining in
the experimental group was the weakest with a positive expression
rate of 34.5±2.8%, which was significantly lower than the other two
groups (P<0.05) (Fig. 7d).
Discussion
In recent years, molecular biology research has
showed that primary gallbladder carcinoma results from the
different effects of polygenes, as well as genetic and
environmental carcinogenic factors (14,15).
The Bcl-2 gene is one of the most important regulatory
factors in cell apoptosis which plays an important role in the
initiation and progression of gallbladder carcinoma. The
Bcl-2 gene was first discovered from chromosome fragment
sites in follicle type human non-Hodgkin’s B lymphoma by Tsujimoto
et al(16), thus it was
called Bcl-2. Bcl-2 protein, one of the Bcl-2 protein
family members, is an important gene which inhibits cell apoptosis.
At present, it is believed that the Bcl-2 gene inhibits cell
apoptosis through blocking many pro-apoptotic factors. The normal
Bcl-2 gene, which has three exons and two promoters and is
~230 kb, is located in chromosome 18q21.3. There is an intron of
~225 kb between exons 1 and 2, and the length of the intron between
exons 2 and 3 is ~370 kb. The Bcl-2 gene acts at the end of
the apoptotic pathway. Its actions include blocking digestion of
DNA by endonuclease, influencing DNA repair, blocking apoptosis
proteins induced by DNA damage or directly acting on these proteins
to render them ineffective. In addition, Bcl-2 protein coded
by the gene can inhibit normal programmed cell death and stimulate
the occurrence of tumors with overgrowth. Research has shown that
the Bcl-2 gene is highly expressed in a variety of malignant
tumors. Indeed, the Bcl-2 gene is highly expressed and the
Bcl-2 protein positive expression rate is ~23.4–51.7% in
gallbladder carcinoma (5). The
expression in gallbladder carcinoma is much higher than that in
gallbladder adenoma and the Bcl-2 gene expression level in poorly
differentiated gallbladder carcinoma is higher than that in
well-differentiated gallbladder carcinoma. The Bcl-2 protein
expression level in the early stage of gallbladder carcinoma is
higher than that in the progressive stage. The apoptosis index (AI)
and ratio of AI/proliferation index (MI) are higher in
Bcl-2-negative gallbladder carcinoma compared with positive
tumors. This suggests that the Bcl-2 gene, which is a
prospective target gene, plays an important role in the initiation
and progression of gallbladder carcinoma.
RNAi technology is a type of post transcriptional
gene silencing phenomenon, that uses sequence-specific small
molecule RNA to identify target mRNA, and then cleaves in the
specific site via the protein complex RISC. This leads to specific
mRNA degradation and eventual blockage of target gene expression.
Its characteristics include high efficiency, specificity, ease and
rapidity. It can block target genes, similarly to the effect of
gene knockout (17). This
technology has been used widely and has promoted research on gene
function. RNAi technology has already been applied to a variety of
malignant tumor gene therapies and has obtained better results in
diseases such as chronic lymphoma and breast cancers (18,19).
However, it is still used in the experimental stages of gallbladder
carcinoma therapy.
In our study, after successfully constructing the
Bcl-2 RNAi vector, we introduced it into human gallbladder
GBC-SD cells through both transient and stable transfection. The
mRNA and protein expression levels of the Bcl-2 gene were
tested by RT-PCR and western blot analysis. The results showed that
the vector significantly inhibited Bcl-2 mRNA transcription
and protein translation.
The growth and proliferation of gallbladder
carcinoma cells transfected with Bcl-2 siRNA were tested by
MTT method. The results showed that GBC-SD cell proliferative
ability was significantly decreased after Bcl-2 gene
silencing. The inhibition of cell growth was time-dependent.
Indeed, although the growth of GBC-SD cells was inhibited at each
tested time, the strongest inhibitory effects were observed at 24
and 36 h after transfection, with inhibition rates of 38.12±0.48
and 38.08±0.75, respectively. The inhibition rate began to decrease
after 48 and 60 h with inhibition rates of 34.67±0.63 and
31.98±0.52, respectively.
Cell apoptosis was analyzed by flow cytometry. The
results showed that the apoptotic percentage of GBC-SD/Bcl-2
siRNA cells was 30.83±4.2%, which was significantly higher than
that of the GBC-SD/Bcl-2 negative control (4.3±1.3%) and
GBC-SD cells (3.6±1.1%) (P<0.05). This confirms that the
Bcl-2 protein is an important gene which inhibits cell
apoptosis and that inhibition of Bcl-2 is able to induce
apoptosis in gallbladder cancer GBC-SD cells.
We further detected the sensitivity of Bcl-2
siRNA transfected gallbladder carcinoma cells to 4 types of
commonly used chemotherapy drugs (5-FU, MMC, ADM and DDP). The
results showed that the sensitivity of gallbladder carcinoma cells
to the 4 drugs increased to different extents after Bcl-2
gene silencing, but only the results of the 5-FU group were
statistically significant (P<0.05). There were no significant
differences between the other groups. Thus, these data provide
strong evidence that gene therapy targeting Bcl-2 in
gallbladder carcinoma can enhance chemotherapeutic efficiency.
A subcutaneous gallbladder carcinoma xenograft nude
mouse model was designed for the tumorigenicity assay. The results
showed that the tumorigenicity of the control group was 100%,
indicating highly malignant and oncogenic abilities of gallbladder
carcinoma. In contrast, the results showed that after transfecting
GBC-SD cells with Bcl-2 siRNA, oncogenic abilities
significantly decreased, and the average volume, average growth
rate, and average weight of the experimental group tumors were all
significantly lower than those of the control group. The results of
Bcl-2 siRNA therapy showed that local injection with the
recombinant plasmid of the Bcl-2 siRNA vector inhibited the
growth of GBC-SD cell tumor xenografts in nude mice. In order to
overcome the disadvantage of siRNA degradation in vivo, we
injected it at multiple sites. However, although we found that
siRNA expressed by the plasmid vector inhibited the growth of
tumors in nude mice when compared with the empty plasmid vector,
the tumor growth rate was still high. Although it decreased
Bcl-2 expression, the degree of reduction was limited; tumor
cells still expressed Bcl-2 at a high level. In this
experiment, whether or not Bcl-2 expression is altered due
to the length of transfection time was not tested. Other apoptotic
proteins were not tested, this is an additional limitation of our
study. Therefore, further studies are needed to make siRNA
transfection more efficient, possibly through the exploration of
more appropriate transfection vectors, transfection approaches and
transfection timings.
Acknowledgements
The present study was supported by Science and
Technology Fund of Shaanxi Province (no. 2008K09-05) and the
National Natural Science Foundation of China (no. 30971340).
References
|
1
|
Lai CH and Lau WY: Gallbladder cancer - a
comprehensive review. Surgeon. 6:101–110. 2008. View Article : Google Scholar
|
|
2
|
Zhu AX, Hong TS, Hezel AF and Kooby DA:
Current management of gallbladder carcinoma. Oncologist.
15:168–181. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Miller J and Jarnagin WR: Gallbladder
carcinoma. Eur J Surg Oncol. 34:306–312. 2008. View Article : Google Scholar
|
|
4
|
Pelengaris S, Khan M and Evan GI:
Suppression of Myc-induced apoptosis in beta cells exposes multiple
oncogenic properties of Myc and triggers carcinogenic progression.
Cell. 109:321–334. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mikami T, Yanagisawa N, Baba H, Koike M
and Okayasu I: Association of Bcl-2 protein expression with
gallbladder carcinoma differentiation and progression and its
relation to apoptosis. Cancer. 85:318–325. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Quan ZW, Wu K, Wang J, Shi W, Zhang Z and
Merrell RC: Association of p53, p16, and vascular endothelial
growth factor protein expressions with the prognosis and metastasis
of gallbladder cancer. J Am Coll Surg. 193:380–383. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Goldin RD and Roa JC: Gallbladder cancer:
a morphological and molecular update. Histopathology. 55:218–229.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shaffer EA: Gallbladder cancer: the
basics. Gastroenterol Hepatol. 4:737–741. 2008.PubMed/NCBI
|
|
9
|
Weyhenmeyer B, Murphy AC, Prehn JH and
Murphy BM: Targeting the anti-apoptotic Bcl-2 family members for
the treatment of cancer. Exp Oncol. 34:192–199. 2012.PubMed/NCBI
|
|
10
|
García-Sáez AJ: The secrets of the Bcl-2
family. Cell Death Differ. 19:1733–1740. 2012.
|
|
11
|
Sasatomi E, Tokunaga O and Miyazaki K:
Spontaneous apoptosis in gallbladder carcinoma. Relationships with
clinicopathologic factors, expression of E-cadherin, bcl-2
protooncogene, and p53 oncosuppressor gene. Cancer. 78:2101–2110.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Meister G and Tuschl T: Mechanisms of gene
silencing by double-stranded RNA. Nature. 431:343–349. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fjose A, Ellingsen S, Wargelius A and Seo
HC: RNA interference: mechanisms and applications. Biotechnol Annu
Rev. 7:31–57. 2001. View Article : Google Scholar
|
|
14
|
Maurya SK, Tewari M, Mishra RR and Shukla
HS: Genetic aberrations in gallbladder cancer. Surg Oncol.
21:37–43. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kuroki T, Tajima Y, Matsuo K and Kanematsu
T: Genetic alterations in gallbladder carcinoma. Surg Today.
35:101–105. 2005. View Article : Google Scholar
|
|
16
|
Tsujimoto Y, Finger LR, Yunis J, Nowell PC
and Croce CM: Cloning of the chromosome breakpoint of neoplastic B
cells with the t(14;18) chromosome translocation. Science.
226:1097–1099. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mello CC and Conte D Jr: Revealing the
world of RNA interference. Nature. 431:338–342. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tyner J and Druker BJ: RNAi screen for
therapeutic target in leukemia. Cell Cycle. 8:21442009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lima RT, Martins LM, Guimarães JE, Sambade
C and Vasconcelos MH: Specific downregulation of bcl-2 and xIAP by
RNAi enhances the effects of chemotherapeutic agents in MCF-7 human
breast cancer cells. Cancer Gene Ther. 11:309–316. 2004. View Article : Google Scholar : PubMed/NCBI
|