Introduction
Worldwide, lung cancer is the most common cancer
with high incidence and mortality rates (1,2).
Non-small cell lung cancer accounts for nearly 80% of the disease,
while lung adenocarcinoma is the most common type of non-small cell
lung cancer (1). Lung
adenocarcinoma is characterized by peripheral location in the lung
and often has activating mutations in the K-ras oncogene
(3,4). Recently, several studies have
highlighted the importance of mutations of the epidermal growth
factor receptor (EGFR) and other oncogenes (5,6);
however, the causes and biology of lung adenocarcinoma are not yet
fully understood (7,8).
In cancer cells, uncontrolled cell proliferation
involves not only dysfunctions in the control of cell growth and
division but also adjustments in energy metabolism that is utilized
to support cancer cell proliferation. Several important mutant
oncogenes (e.g., TP53) have been reported to be associated
with abnormal glycolytic fueling (9,10).
Recently, deregulated cellular energy metabolism has been viewed as
a new hallmark of cancer (11). As
an indispensable cell energy factory for the survival of cells, the
mitochondrion is an important regulator of vital and lethal
functions, particularly the intrinsic pathway of apoptosis
(12). More and more data
demonstrate that the mitochondrion is a critical target for cancer
therapy (13,14).
Sirtuins are a conserved family of deacetylases and
mono-ADP-ribosyltransferases that use NAD+ as a
co-substrate (15). There are 7
members of the sirtuin family in mammals (Sirt1–Sirt7) (15). These proteins are believed to be
involved in stress response, metabolism and longevity (15). Sirtuins are widely expressed in
different tissues and are localized in different subcellular
compartments (15,16). Among the 7 members, Sirt3, Sirt4 and
Sirt5 are localized in the mitochondrion. Sirt3 is believed to be
the major deacetylase within the mitochondrion, since the
deacetylase activity of Sirt4 and Sirt5 is rather weak (17).
The role of Sirt3 in tumor biology has become a
topic of increased interest in recent years. Ashraf et
al(18) first reported that
increased levels of Sirt3 and Sirt7 transcription are associated
with node-positive breast cancer. Kim et al(19) demonstrated that gene deletion of
Sirt3 facilitates the development of breast tumors in mice,
suggesting the tumor-suppressive effect of Sirt3 in breast cancer,
which was confirmed by a later study (20). A recent report showed that Sirt3
inhibits hepatocellular carcinoma cell growth through reducing
Mdm2-mediated p53 degradation (21). However, the exact role of Sirt3 in
other tumors is still being discovered. In the present study, we
investigated the expression of Sirt3 in human lung adenocarcinoma
tissues and explored the potential role of Sirt3 in lung
adenocarcinoma.
Materials and methods
Reagents
Antibodies against Sirt3 and apoptosis-inducing
factor (AIF) were purchased from Millipore Chemicon International
(Temecula, CA, USA). Antibodies against cleaved caspase-3, bax,
bcl-2, bad, bcl-x/L, p53 and p21 were purchased from Cell Signaling
Technology, Inc. (Danvers, MA, USA). Antibody against actin was
obtained from Sigma-Aldrich, (St. Louis, MO, USA). The Annexin V +
PI kit was purchased from Promega Corporation (Madison, WI, USA).
DAPI and dichlorofluorescein diacetate (DCFH-DA) were purchased
from Invitrogen Life Technologies (Carlsbad, CA, USA). Enhanced
chemiluminescence and protease/phosphatase inhibitors were
purchased from Pierce Biotechnology, Inc. (Rockford, IL, USA).
Human lung adenocarcinoma tissue
Four pairs of lung adenocarcinoma and matched normal
adjacent tissue extracts were obtained from Chinese patients who
underwent surgical resection for diagnosis and therapy in our
hospital. Samples were obtained following informed consent
according to an established protocol approved by the Ethics
Committee of Central South University.
Cell culture
The A549 lung adenocarcinoma cell line was obtained
from ATCC (Manassas, VA, USA). Cells were cultured in Dulbecco's
modified Eagle's medium (DMEM) supplemented with 10% (v/v) fetal
bovine serum (FBS) in 95% O2 and 5% CO2.
Construction of the adenovirus expressing
Sirt3
The adenovirus expressing Sirt3 (Ad-Sirt3) or the
control adenovirus expressing GFP (Ad-GFP) were generated using the
RAPAd® CMV adenoviral expression system (Cell Biolabs,
Inc., San Diego, CA, USA) according to the manufacturer's protocol
as previously described (21).
Briefly, mouse Sirt3 cDNA was cloned into pacAd5 CMV-IRES vector.
Then, pacAd5 CMV-IRES-Sirt3 and pacAd5 9.2–100 backbone vectors
were linearized by PacI. The purified linearized DNAs were
cotransfected into 293 cells using Lipofectamine™ Plus (Invitrogen
Life Technologies). On day 8, adenovirus-containing cells and media
were harvested. Viruses were released by 3 freeze/thaw cycles and
stored at −80°C. For virus transfection, 20 μl of viral stock
solution was added into the culture medium (2 ml) for 6 h.
Cell viability assay
Cell viability was evaluated by a non-radioactive
Cell Counting Kit-8 (CCK-8) assay as described previously (21,22).
Ad-GFP- and Ad-Sirt3-transfected cells (5×103) were
seeded into 48-well plates and cultured overnight to allow
attachment. After being serum-starved for 8 h, FBS was added into
the medium. At 12, 24, 36 and 48 h, cells were incubated with 10 μl
of CCK-8 solution for 3 h at 37°C, and then the optical density at
450 nm was analyzed using a microplate reader (Tecan, Switzerland).
Experiments were performed in duplicate.
Quantitative real-time PCR analysis
Real-time PCR analysis was performed on an ABI Prism
7500 sequence detection system using PrimerScript® RT
reagent kit (Takara Bio, Inc., Shiga, Japan). The total RNA was
extracted form human tissue using TRIzol (Invitrogen Life
Technologies) and 5 μg of RNA as first-strand cDNA was used. cDNA
(100 ng) was amplified using primers as follows: Sirt3 sense,
ACAGCAACCTCCAGCAGTACGA and antisense, CGTGTAGAGCCGCAGAAGCA; β-actin
sense, GCACTCTTCCAGCCTTCCTTCC and antisense, CCGCCAGACAGCACTGTGTT.
The mRNA levels of housekeeping gene β-actin were used as
control.
Immunochemistry
Immunochemistry was performed as described
previously (23,24). Briefly, the A549 cells were seeded
onto coverslips and transfected with Ad-GFP or Ad-Sirt3 for 6 h. At
24 h after transfection, cells were washed twice with PBS and then
fixed with buffer containing 4% paraformaldehyde and 0.1% Triton
X-100 at room temperature for 20 min, followed by incubation with
an antibody against AIF at 37°C for 2 h and incubation with Alexa
Fluor 555-conjugated secondary antibody. DAPI was used to stain the
nucleus.
Immunoblotting
Immunoblotting analyses of cell extracts were
performed as described previously (22,25).
Human tissues or cells were lysed with RIPA buffer with protease
inhibitor/protein phosphatase inhibitors. Samples were subjected to
10% SDS-PAGE, and transferred onto PVDF membranes at 100 V for 1–2
h. After being blocked in blocking buffer for 4 h, the membrane was
incubated with a specific primary antibody and then followed by the
HRP-labeled secondary antibody, and investigation using the
enhanced chemiluminescence system (Pierce Biotechnology, Inc.,
Rockford, IL, USA).
Annexin V + PI staining assay
The Annexin V + PI assay for apoptosis was performed
using flow cytometry as described previously (23). Briefly, Ad-GFP- and
Ad-Sirt3-transfected cells were trypsinized, washed in ice-cold PBS
and re-suspended in 1 ml of the supplied buffer (1×106
cells/ml). A 100 μl sample (1×105 cells) was incubated
with 5 μl FITC-conjugated Annexin V and 5 μl PI for 30 min at 25°C
in the dark. Then, cells were analyzed immediately using a BD
FACSCalibur (BD Biosciences, Franklin Lakes, NJ, USA). The
apoptotic cells were estimated as the percentage of cells that
stained positive for FITC-Annexin V while remaining impermeable to
PI (Annexin V+/PI−).
Statistical analysis
Data are expressed as means ± SEM. Differences were
evaluated by a two-tailed Student's t-test or ANOVA followed by
Tukey's post hoc test. Statistical significance was set at
P<0.05.
Results
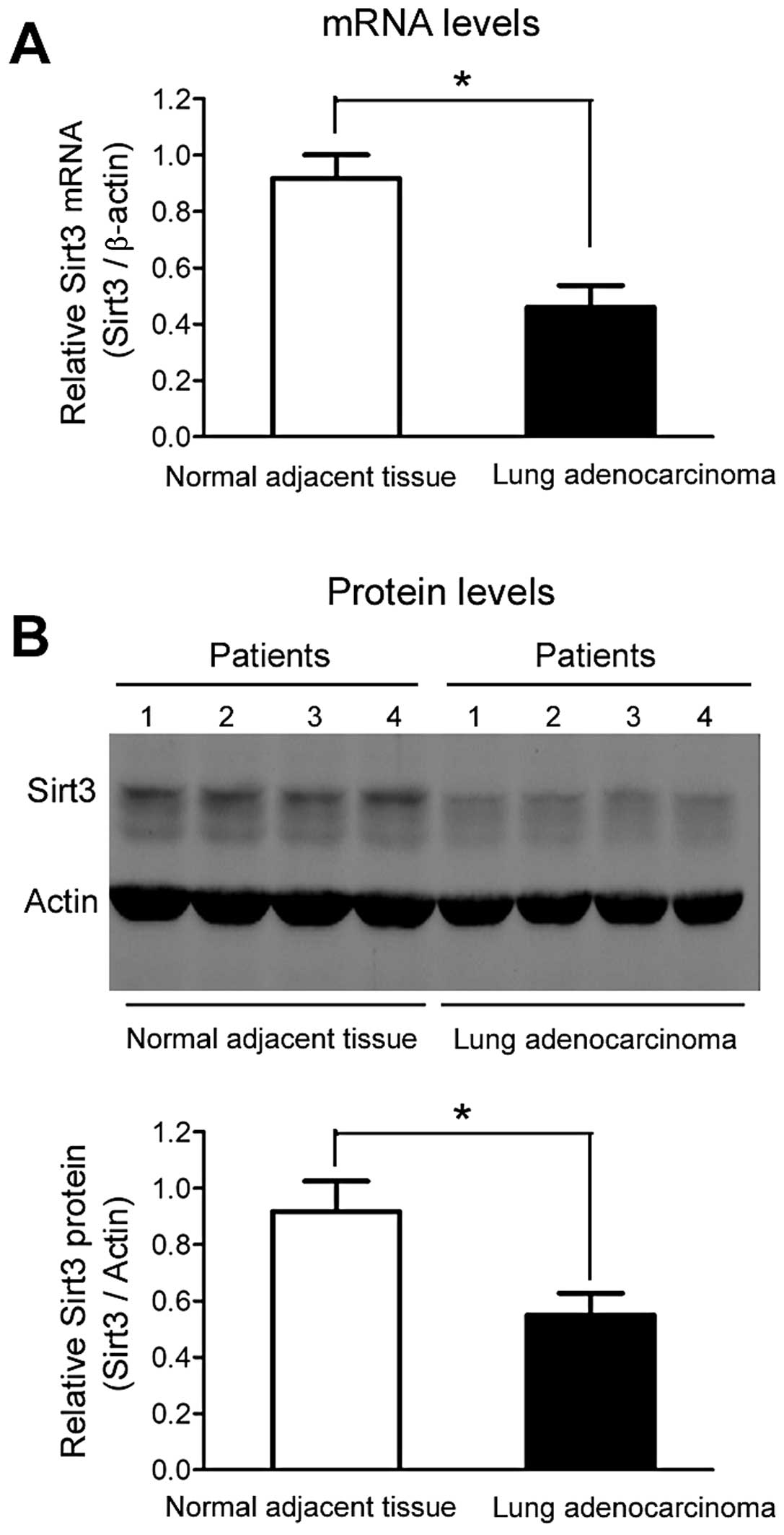
Sirt3 is downregulated in human lung
adenocarcinoma tissue
As shown in Fig. 1A,
Sirt3 mRNA levels were significantly downregulated (~50–60%) in
human lung adenocarcinoma tissue when compared with normal adjacent
tissue. Similarly, a marked downregulation of Sirt3 protein levels
in human lung adenocarcinoma tissue was observed (Fig. 1B).
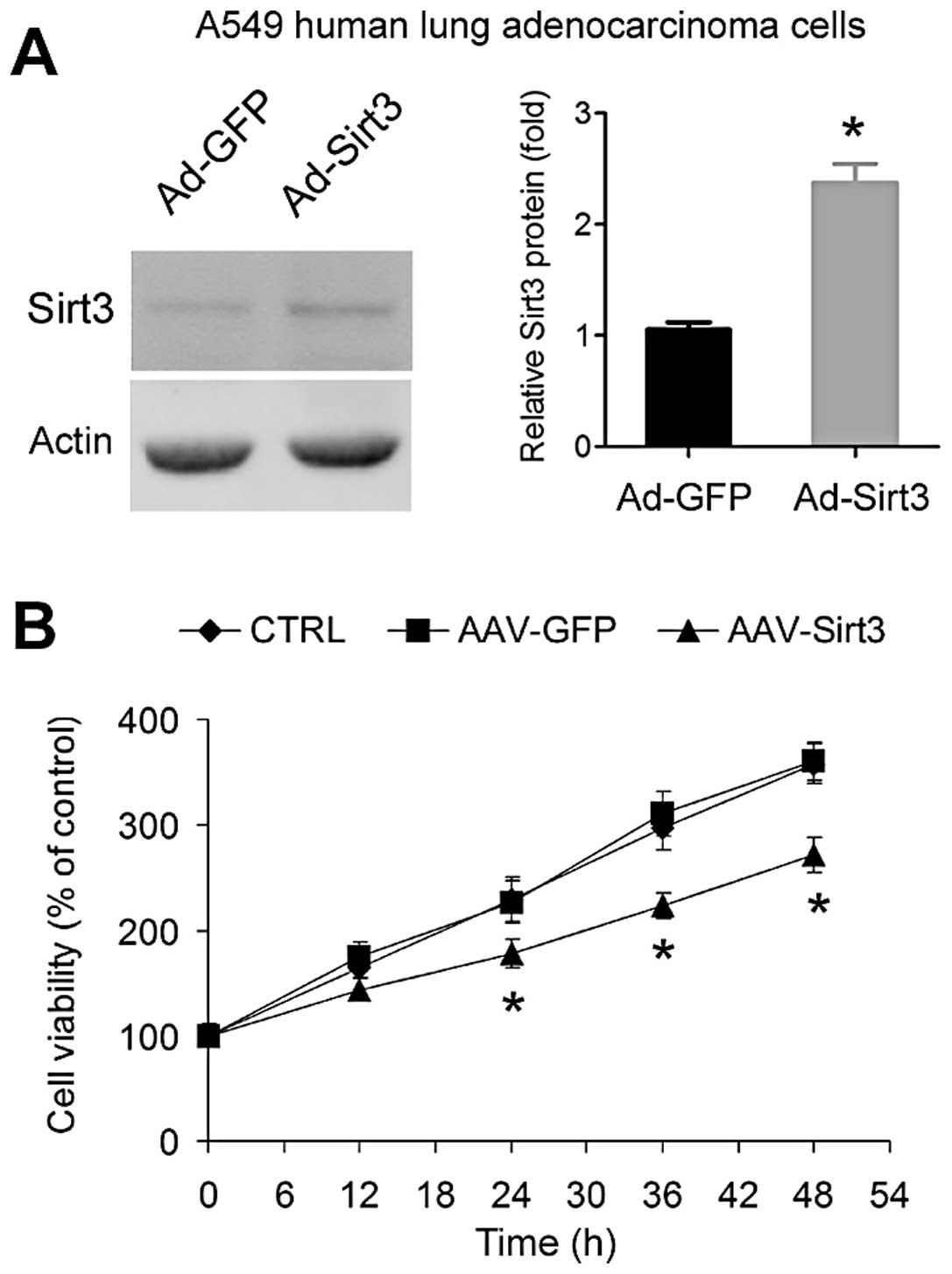
Overexpression of Sirt3 inhibits lung
adenocarcinoma cell growth
We next tested whether overexpression of Sirt3
influences A549 lung adenocarcinoma cell growth in vitro.
Overexpression of Sirt3 using adenovirus (Fig. 2A) significantly inhibited the growth
of A549 cells (Fig. 2B).
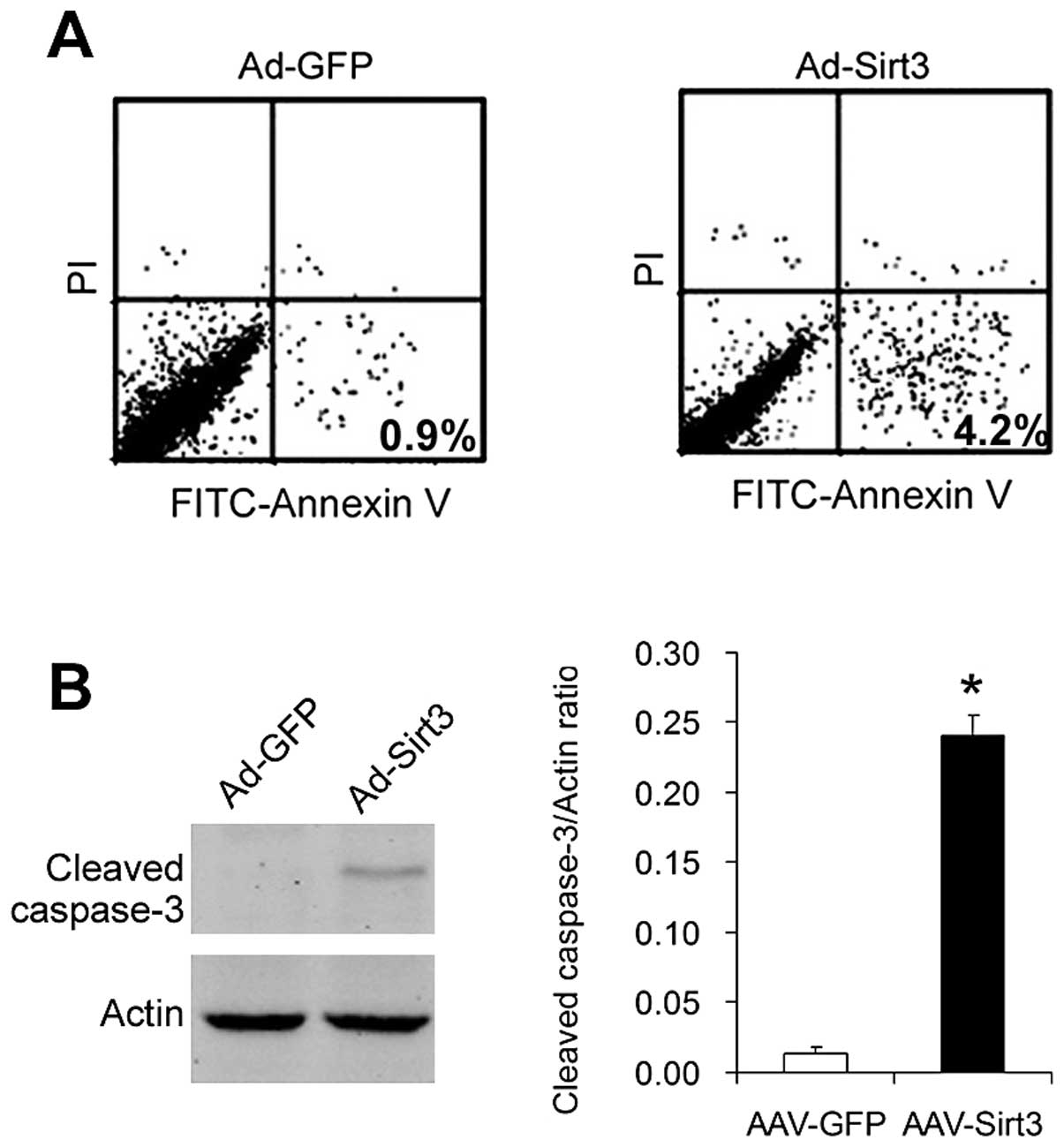
Overexpression of Sirt3 induces apoptosis
in lung adenocarcinoma cells
We studied the effects of overexpression of Sirt3 on
the apoptosis of lung adenocarcinoma cells. First, we analyzed
Ad-GFP- and Ad-Sirt3-transfected cells with the FITC-Annexin V + PI
staining assay. As shown in Fig.
3A, the ratio of apoptotic cells (Annexin
V+/PI−) was ~0.9% in the control
Ad-GFP-transfected cells, whereas the apoptotic ratio increased to
4.2% in Ad-Sirt3-transfected cells.
Then, we analyzed the levels of cleaved caspase-3, a
key mediator and marker of apoptosis. As shown in Fig. 3B, cleaved caspase-3 expression was
detected in the cells overexpressing Sirt3 (Ad-Sirt3-transfected
cells) but not in the control cells (Ad-GFP-transfected cells).
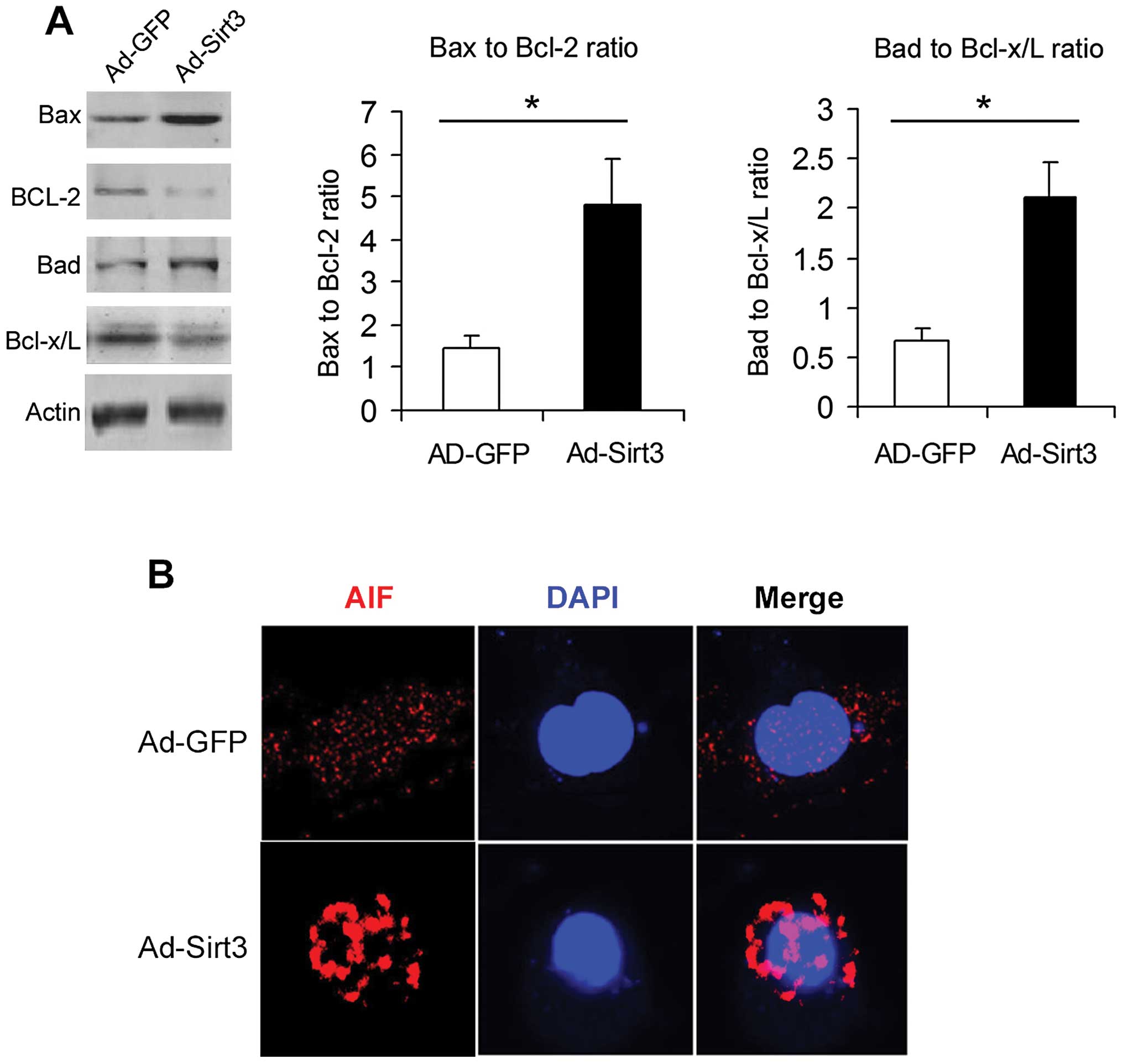
Overexpression of Sirt3 modulates
apoptotic-related proteins in lung adenocarcinoma cells
The Bcl-2 family proteins and AIF are important
determinants regulating cellular apoptosis. Overexpression of Sirt3
increased the bax-bcl-2 ratio and bad-bcl-x/L ratio (Fig. 4A). Moreover, immunofluorescence
analysis (Fig. 4B) showed that
Sirt3 overexpression induced AIF nuclear translocation.
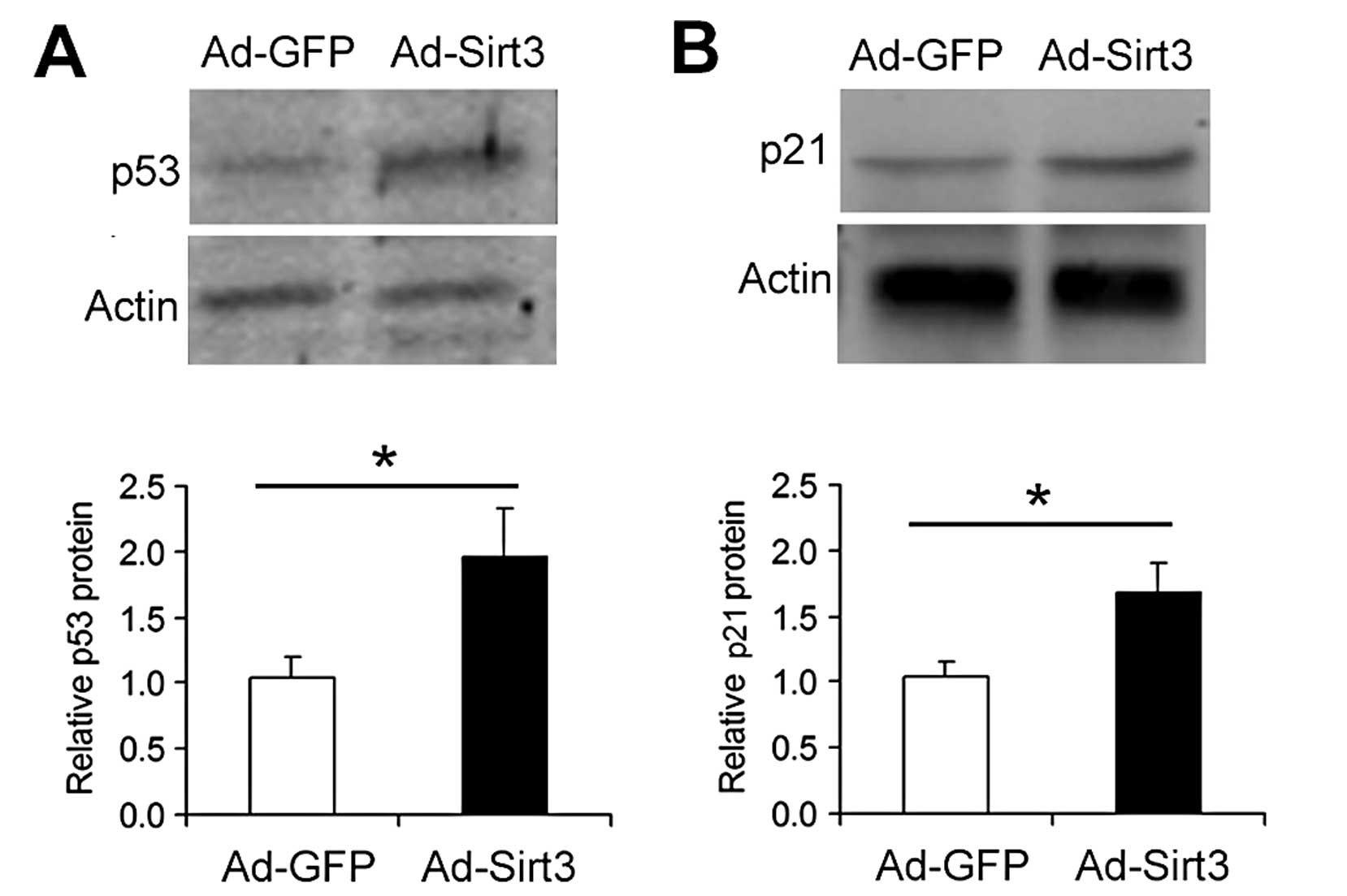
Overexpression of Sirt3 upregulates the
p53 signaling pathway in lung adenocarcinoma cells
Next, we studied the influence of Sirt3
overexpression on p53 signaling in A549 cells. Compared with the
Ad-GFP-transfected cells, Ad-Sirt3-transfected cells exhibited
increased protein levels of p53 (Fig.
5A) and p21 (Fig. 5B), a
p53-downstream factor.
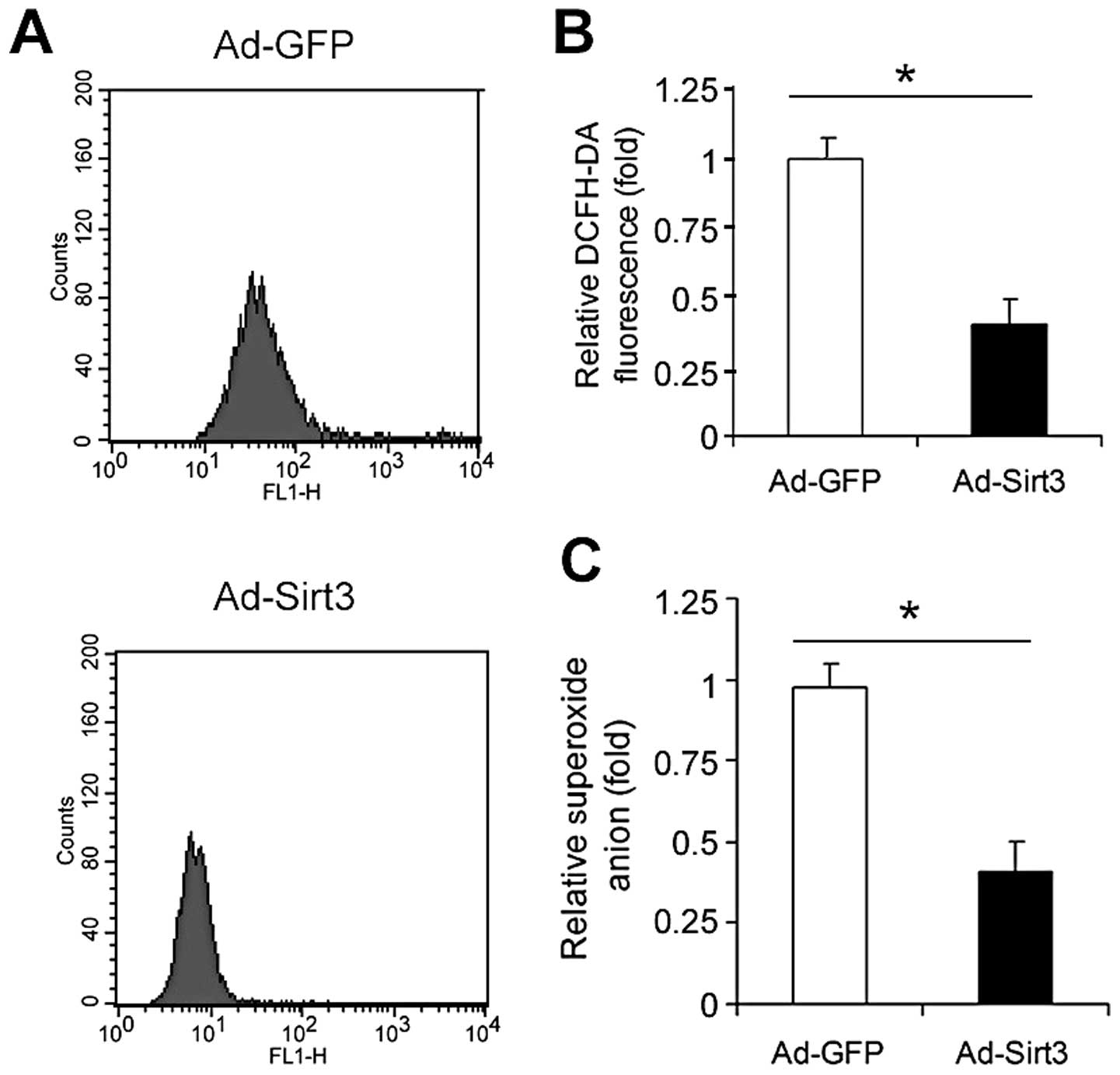
Overexpression of Sirt3 decreases
oxidative stress in lung adenocarcinoma cells
Finally, we studied the influence of Sirt3
overexpression on oxidative stress in A549 cells. The reactive
oxygen species (ROS) generation measured by the DCFH-DA assay
(Fig. 6A) revealed that
overexpression of Sirt3 significantly decreased the total ROS level
in A549 cells (Fig. 6B). Moreover,
overexpression of Sirt3 attenuated the superoxide anion level in
A549 cells (Fig. 6B).
Discussion
In the present study, we initially demonstrated that
Sirt3 was markedly downregulated in lung adenocarcinoma tissue when
compared with that in the normal adjacent tissue. Using
adenovirus-mediated overexpression, we found that Sirt3
overexpression inhibited the growth of A549 lung adenocarcinoma
cells. Further analyses, including Annexin V + PI assay, cleaved
caspase-3 immunoblotting, bax-bcl-2 ratio, bad-bcl-x/L ratio and
AIF translocation, showed that Sirt3 overexpression promoted
apoptosis in A549 lung adenocarcinoma cells. Furthermore, Sirt3
overexpression downregulated ROS and superoxide anion levels in
A549 lung adenocarcinoma cells. These data clearly suggest that
Sirt3 is a tumor suppressor through the induction of apoptosis in
lung adenocarcinoma cells.
The first important finding of our study is that
Sirt3 was downregulated in human lung adenocarcinoma tissue.
Acetylation was found in >20% of mitochondrial proteins,
including many longevity regulators and metabolism enzymes in a
large-scale proteomics analysis (26), implicating the wide influence of
acetylation modification among mitochondrial proteins. As
acetylation modification is mainly mediated by sirtuins and Sirt3
is the major deacetylase of sirtuins in mitochondria (17), Sirt3 is believed to be a potent
regulator in mitochondria. Furthermore, the energy status dictates
the status of mitochondrial protein acetylation, suggesting that
Sirt3-mediated mitochondrial acetylation may be a critical
regulatory mechanism underlying the adaptive response to energy
stress. In line with the potential role of Sirt3 in mitochondrial
biology, metabolically active tissues with high oxidative capacity,
such as skeletal muscle, liver, brain, kidney and adipose tissue,
express Sirt3 abundantly (27). The
downregulation of Sirt3 in lung adenocarcinoma may lead to enhanced
acetylation status and ROS generation in mitochondria, which are
tightly associated with higher cancer risk (28,29).
Previously, significant reduction in the Sirt3 copy
number was found in human breast cancer (30) and hepatocellular carcinoma (21,31).
Moreover, loss of Sirt3 in vitro led to tumorigenesis
(19), and Sirt3 levels were
decreased in human breast cancer (19,20).
In breast cancer cells, Finley et al(20) demonstrated that SIRT3 mediates
metabolic reprogramming by destabilizing hypoxia inducible factor
1α (HIF-1α), a transcription factor that controls glycolytic gene
expression. Murine tumors lacking Sirt3 exhibit abnormally high
levels of ROS that directly induce genomic instability and cellular
metabolic reprogramming (20). Bell
et al(32) also showed that
Sirt3 suppressed tumor growth via inhibition of HIF-1α. All these
data suggest the inhibitory effects of Sirt3 in tumor. However, we
also noted a contradictory finding. Sirt3 was upregulated in oral
cancer and was required to protect oral cancer from stress-mediated
cell death by various stimuli (33). In our studies, we found that Sirt3
was downregulated in human lung adenocarcinoma tissue, supporting
the tumor-suppressive role of Sirt3 in lung adenocarcinoma.
Subsequent functional analysis also showed that Sirt3
overexpression inhibited A549 lung adenocarcinoma cell
proliferation.
The inhibitory effect of Sirt3 overexpression on
lung adenocarcinoma cell growth was very obvious in vitro,
which prompted us to study apoptosis. Using Annexin V + PI assay
and cleaved caspase-3 immunoblotting, we confirmed the
apoptosis-inducing effects of Sirt3 in A549 lung adenocarcinoma
cells. After revealing the pro-apoptotic feature of Sirt3, we
observed that Sirt3 overexpression was modulated by apoptotic
signaling pathways (bax, bcl-2, bad, bcl-x/L and AIF). Bcl-2 family
proteins are known to play a pivotal role in the induction of
mitochondrial caspase activation and in the regulation of apoptosis
(34). AIF is a flavoprotein that
is normally confined to the mitochondrial intermembrane space, yet
translocates to the nucleus to induce peripheral chromatin
condensation and triggers large-scale DNA degradation to fragments
of ~50 kbp (35). In our study,
overexpression of Sirt3 increased bax-bcl-2 and bad/bcl-x/L ratios
and induced AIF nuclear translocation. We postulated that these
changes may contribute to the apoptotic-inducing effects of Sirt3
in A549 lung adenocarcinoma cells.
Finally, we provide evidence showing that Sirt3
overexpression was associated with an upregulated p53 protein level
and a decreased ROS level in lung adenocarcinoma cells. Sirt1,
another member of the sirtuin family, has been found to be able to
directly deacetylate p53 and increase p53 protein level (36,37).
Recently, a study demonstrated that Sirt3 also enhanced the p53
protein level in hepatocellular carcinoma cells (21). In agreement with these studies, we
observed that Sirt3 overexpression upregulated p53 and its
downstream factor p21 in lung adenocarcinoma cells. Giving that p53
is a well-known tumor suppressor (38), we hypothesized that p53 upregulation
is quite likely to contribute to the tumor-suppressive effect of
Sirt3 in lung adenocarcinoma cells.
Collectively, we demonstrated that the expression of
Sirt3 was decreased in human lung adenocarcinoma tissue.
Overexpression of Sirt3 exhibited an obvious antitumor effect in
the A549 lung adenocarcinoma cell line through induction of
apoptosis. Our finding concerning the regulation of lung
adenocarcinoma cell growth by Sirt3 may provide an important focus
for the further understanding of lung adenocarcinoma and novel
therapeutic interventions.
Acknowledgements
This study was supported by grants from the ‘New
Teachers Fund for Doctor Stations’, Ministry of Education of China
(no. 20100162120059) and the Fundamental Research Funds for the
Central Universities (no. 2011QNZT175).
References
|
1
|
Herbst RS, Heymach JV and Lippman SM: Lung
cancer. N Engl J Med. 359:1367–1380. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jemal A, Simard EP, Dorell C, et al:
Annual report to the nation on the status of cancer, 1975–2009,
featuring the burden and trends in human papillomavirus
(HPV)-associated cancers and HPV vaccination coverage levels. J
Natl Cancer Inst. 105:175–201. 2013.
|
|
3
|
Gazdar AF: The molecular and cellular
basis of human lung cancer. Anticancer Res. 14:261–267.
1994.PubMed/NCBI
|
|
4
|
Graziano SL, Gamble GP, Newman NB, et al:
Prognostic significance of K-ras codon 12 mutations in
patients with resected stage I and II non-small-cell lung cancer. J
Clin Oncol. 17:668–675. 1999.
|
|
5
|
Ding L, Getz G, Wheeler DA, et al: Somatic
mutations affect key pathways in lung adenocarcinoma. Nature.
455:1069–1075. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Weir BA, Woo MS, Getz G, et al:
Characterizing the cancer genome in lung adenocarcinoma. Nature.
450:893–898. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Molina JR, Yang P, Cassivi SD, Schild SE
and Adjei AA: Non-small cell lung cancer: epidemiology, risk
factors, treatment, and survivorship. Mayo Clin Proc. 83:584–594.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mitsudomi T and Yatabe Y: Epidermal growth
factor receptor in relation to tumor development: EGFR gene and
cancer. FEBS J. 277:301–308. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
DeBerardinis RJ, Lum JJ, Hatzivassiliou G
and Thompson CB: The biology of cancer: metabolic reprogramming
fuels cell growth and proliferation. Cell Metab. 7:11–20. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jones RG and Thompson CB: Tumor
suppressors and cell metabolism: a recipe for cancer growth. Genes
Dev. 23:537–548. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: the next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kroemer G, Galluzzi L and Brenner C:
Mitochondrial membrane permeabilization in cell death. Physiol Rev.
87:99–163. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fulda S, Galluzzi L and Kroemer G:
Targeting mitochondria for cancer therapy. Nat Rev Drug Discov.
9:447–464. 2010. View
Article : Google Scholar
|
|
14
|
Mathupala SP, Ko YH and Pedersen PL: The
pivotal roles of mitochondria in cancer: Warburg and beyond and
encouraging prospects for effective therapies. Biochim Biophys
Acta. 1797:1225–1230. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Haigis MC and Guarente LP: Mammalian
sirtuins - emerging roles in physiology, aging, and calorie
restriction. Genes Dev. 20:2913–2921. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Milne JC and Denu JM: The Sirtuin family:
therapeutic targets to treat diseases of aging. Curr Opin Chem
Biol. 12:11–17. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lombard DB, Alt FW, Cheng HL, et al:
Mammalian Sir2 homolog SIRT3 regulates global mitochondrial lysine
acetylation. Mol Cell Biol. 27:8807–8814. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ashraf N, Zino S, Macintyre A, et al:
Altered sirtuin expression is associated with node-positive breast
cancer. Br J Cancer. 95:1056–1061. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kim HS, Patel K, Muldoon-Jacobs K, et al:
SIRT3 is a mitochondria-localized tumor suppressor required for
maintenance of mitochondrial integrity and metabolism during
stress. Cancer Cell. 17:41–52. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Finley LW, Carracedo A, Lee J, et al:
SIRT3 opposes reprogramming of cancer cell metabolism through HIF1α
destabilization. Cancer Cell. 19:416–428. 2011.PubMed/NCBI
|
|
21
|
Zhang YY and Zhou LM: Sirt3 inhibits
hepatocellular carcinoma cell growth through reducing Mdm2-mediated
p53 degradation. Biochem Biophys Res Commun. 423:26–31. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang P, Xu TY, Guan YF, Su DF, Fan GR and
Miao CY: Perivascular adipose tissue-derived visfatin is a vascular
smooth muscle cell growth factor: role of nicotinamide
mononucleotide. Cardiovasc Res. 81:370–380. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang P, Xu TY, Guan YF, et al:
Nicotinamide phosphoribosyltransferase protects against ischemic
stroke through SIRT1-dependent adenosine monophosphate-activated
kinase pathway. Ann Neurol. 69:360–374. 2011. View Article : Google Scholar
|
|
24
|
Wang P, Zhang RY, Song J, et al: Loss of
AMP-activated protein kinase-α2 impairs the insulin-sensitizing
effect of calorie restriction in skeletal muscle. Diabetes.
61:1051–1061. 2012.
|
|
25
|
Wang P, Guan YF, Du H, Zhai QW, Su DF and
Miao CY: Induction of autophagy contributes to the neuroprotection
of nicotinamide phosphoribosyltransferase in cerebral ischemia.
Autophagy. 8:77–87. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kim SC, Sprung R, Chen Y, et al: Substrate
and functional diversity of lysine acetylation revealed by a
proteomics survey. Mol Cell. 23:607–618. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Shi T, Wang F, Stieren E and Tong Q:
SIRT3, a mitochondrial sirtuin deacetylase, regulates mitochondrial
function and thermogenesis in brown adipocytes. J Biol Chem.
280:13560–13567. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Arif M, Senapati P, Shandilya J and Kundu
TK: Protein lysine acetylation in cellular function and its role in
cancer manifestation. Biochim Biophys Acta. 1799:702–716. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Benhar M, Engelberg D and Levitzki A: ROS,
stress-activated kinases and stress signaling in cancer. EMBO Rep.
3:420–425. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chin SF, Teschendorff AE, Marioni JC, et
al: High-resolution aCGH and expression profiling identifies a
novel genomic subtype of ER negative breast cancer. Genome Biol.
8:R2152007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang CZ, Liu L, Cai M, et al: Low SIRT3
expression correlates with poor differentiation and unfavorable
prognosis in primary hepatocellular carcinoma. PLoS One.
7:e517032012. View Article : Google Scholar
|
|
32
|
Bell EL, Emerling BM, Ricoult SJ and
Guarente L: SirT3 suppresses hypoxia inducible factor 1α and tumor
growth by inhibiting mitochondrial ROS production. Oncogene.
30:2986–2996. 2011.PubMed/NCBI
|
|
33
|
Alhazzazi TY, Kamarajan P, Joo N, et al:
Sirtuin-3 (SIRT3), a novel potential therapeutic target for oral
cancer. Cancer. 117:1670–1678. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Youle RJ and Strasser A: The BCL-2 protein
family: opposing activities that mediate cell death. Nat Rev Mol
Cell Biol. 9:47–59. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
35
|
Susin SA, Lorenzo HK, Zamzami N, et al:
Molecular characterization of mitochondrial apoptosis-inducing
factor. Nature. 397:441–446. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Langley E, Pearson M, Faretta M, et al:
Human SIR2 deacetylates p53 and antagonizes PML/p53-induced
cellular senescence. EMBO J. 21:2383–2396. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Cheng HL, Mostoslavsky R, Saito S, et al:
Developmental defects and p53 hyperacetylation in Sir2 homolog
(SIRT1)-deficient mice. Proc Natl Acad Sci USA. 100:10794–10799.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Greenblatt MS, Bennett WP, Hollstein M and
Harris CC: Mutations in the p53 tumor suppressor gene: clues to
cancer etiology and molecular pathogenesis. Cancer Res.
54:4855–4878. 1994.PubMed/NCBI
|