Introduction
Natural products and their derivatives have
historically been invaluable as sources of chemopreventive and
therapeutic agents (1). Recently,
more attention has been paid to bioactive natural products of
endophytic fungi, isolated from higher plants (2). Endophytic fungi are important
components of microbial biodiversity. Biological diversity means
chemical diversity implying unique structural features and
pronounced biological activities as lead structures for novel drugs
(3).
Apoptotic cell death is a self-destructing process
under strict control and plays an important role in tissue
homeostasis. Impairment of the apoptotic signaling pathway leads to
insufficient or excessive cell death which results in the
progression of human cancers (4).
Thus, a chemical compound which promotes cell apoptosis and
inhibits cancer cell proliferation is considered to be a potential
anticancer drug (5).
The p53 family members are mainly involved in
apoptosis induction and cell cycle arrest under cellular stress.
The p53 family consists of 3 members: p53, p63 and p73. Tumor
suppressor p53 is a transcription factor that maintains genome
stability and normal cell growth (6,7). The
tumor suppressor p53 is activated in response to stress such as DNA
damage and oncogene activation. p53 can promote cell apoptosis and
thus enhance the chemosensitivity to anticancer agents (8,9).
Unlike p53, p73 is rarely mutated in human cancers. Due to high
homology to p53, the p53-related protein p73 is capable of
transactivating p53 target genes and also plays an important role
in the regulation of chemosensitivity (10,11).
The p53 family members are an attractive pharmaceutical target for
cancer therapy. Many compounds have been discovered which can
induce the activation of p53 and the death of tumor cells (12–15).
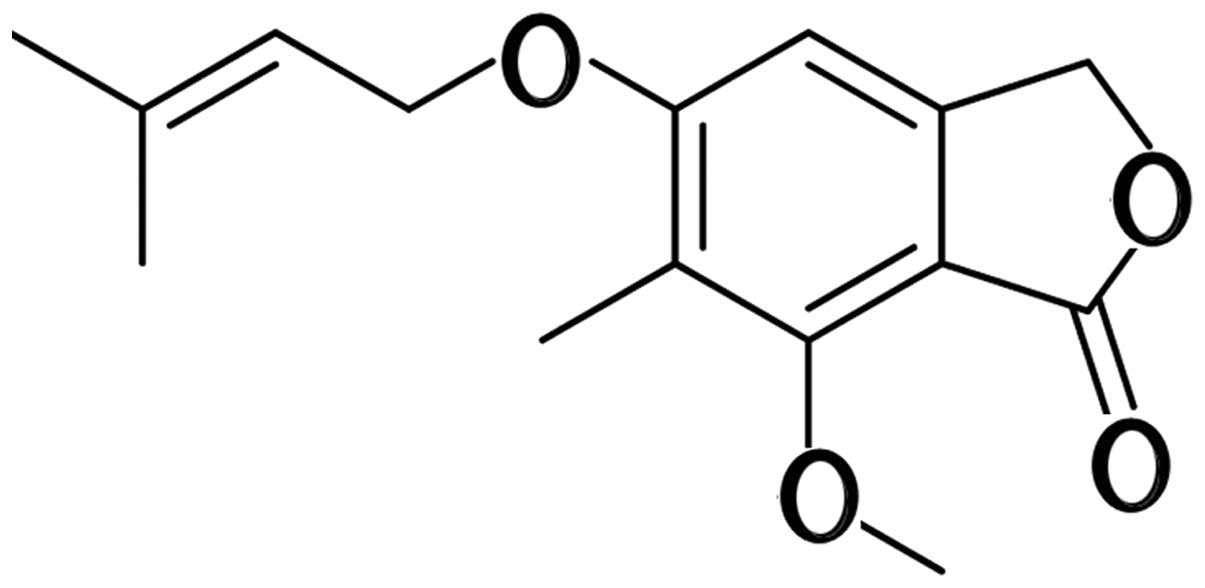
4-(3′,3′-Dimethylallyloxy)-5-methyl-6-methoxyphthalide (DMMP) was
initially found in the liquid culture of the fungus Alternaria
porri (Fig. 1) (16,17).
DMMP was reported to have antifungal activity and cytotoxic
activity in cancer cell lines. In our research, DMMP was isolated
from the plant endophytic fungus Pestalotiopsis
photiniae(18). Although the
cytotoxic activities of DMMP have been reported, little is known
concerning the molecular mechanism of its cytotoxic effect. Here,
for the first time, we discovered that DMMP inhibits the growth of
several cancer cell lines and we investigated the mechanism of its
antiproliferative effect. These results were significant in that
they provide a mechanistic framework for further exploration of the
use of DMMP as a novel antitumor agent.
Materials and methods
Materials
Caspase inhibitor Z-VAD-FMK, caspase-3 specific
inhibitor Z-DEVD-FMK, PD9805 were from Sigma-Aldrich (St. Louis,
MO, USA; MTT, acridine orange and ethidium bromide were from
Amresco LLC (Solon, OH, USA). JC-1 was from Beyotime Institute of
Biotechnology (Jiangsu, China). Rabbit polyclonal antibodies
against Fas-L, Fas, p73, p53, Bax, Bcl-2, survivin, CDK2, Akt,
p-AKT (Thr308), ERK, p-ERK1/2 (Thr202/Tyr204), p-RB (Ser807/811),
GAPDH, Bid, caspase-8, caspase-9, cyclin E, p27KIPI,
Bcl-xL and E2F1 were obtained from Santa Cruz Biotechnology, Inc.
(Santa Cruz, CA, USA). Mouse monoclonal p53 and mouse monoclonal
CDK6 were obtained from Santa Cruz Biotechnology, Inc., and rabbit
polyclonal activated caspase-3 was from Cell Signaling Technology,
Inc. (Danvers, MA, USA). MEM/NEAA medium, DMEM, L15 medium were
from Invitrogen Life Technologies (Carlsbad, CA, USA). Propidium
iodide (PI)/RNase staining buffer and the Annexin V-FITC Apoptosis
Detection kit were from BD Pharmingen (San Diego, CA, USA). PVDF
membranes were from Millipore (Billerica, MA, USA) and the
PrimeScript™ High Fidelity RT-PCR kit and SYBR® Premix
Dimer Eraser were from Takara Bio, Inc. (Shiga, Japan). DMMP
(Fig. 1) was provided by our
research group at Hebei University (purity >99%, HPLC analysis)
(18).
Cell culture
All cell lines were purchased from Cell Resource
Center, IBMS, CAMS/PUMC. MCF7 human breast adenocarcinoma and HeLa
human cervical cancer cells were grown in Dulbecco’s modified
Eagle’s medium (DMEM) supplemented with 10% heat-inactivated fetal
bovine serum (FBS) (Invitrogen Life Technologies). MRC5 normal lung
cells were grown in MEM/NEAA supplemented with 10% heat-inactivated
FBS. MDA-MB-231 human breast adenocarcinoma cells were grown in L15
medium supplemented with 10% heat-inactivated FBS and 1% glutamine.
All cell lines were cultured at 37°C in a humidified incubator
containing 5% CO2.
MTT assay
Cell viability was determined using the colorimetric
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT)
assay. Exponentially growing cells (8,000 cells/well) were seeded
in 96-well plates and cultured at 37°C, under 5% CO2 and
95% air for 24 h. Cells were treated with DMMP at different
concentrations (5, 10, 20, 40 and 80 μg/ml dissolved in DMSO) for
different time periods. The DMSO concentration was maintained below
0.5% which was found to have no antiproliferative effect on the
cell lines. Then 20 μl MTT (5 mg/ml) was added to each well. After
incubation at 37°C for 4 h, 100 μl 10% SDS-HCl was added and
incubated at 37°C overnight. The OD value of the system was
measured at a wavelength of 570 nm. Five wells were counted in 3
different experiments. The 50% inhibitory concentration
(IC50) of DMMP on cells was calculated by the MTT
assay.
Inhibitor treatment
The cell culture was preincubated for 2 h with one
of the following inhibitors: the cell-permeable pan-caspase
inhibitor Z-VAD-FMK (10 μM), the caspase-3-specific inhibitor
Z-DEVD-FMK (10 μM) or ERK inhibitor PD98059 (20 μM) before the
addition of DMMP. Cell viability was measured by the MTT assay.
Caspase activity assay
Caspase activation was measured using a caspase
colorimetric assay kit (Beyotime Institute of Biotechnology).
Briefly, following cell treatment with 40 μg/ml DMMP for the
indicated periods of time, cells were harvested and 25 μl of cold
lysis buffer/106 cells was added. The cell lysates were
incubated on ice for 10 min and then centrifuged at 16,000 × g for
15 min. The protein concentration was determined by Bradford assay.
To each reaction well, 10 μl of caspase-3 fluorogenic substrate
(Ac-DEVD-pNA) was added to 10 μl sample (1–3 mg/ml) and 80 μl
detection buffer. The plate was incubated at 37°C for 2 h. The
plate was read on a fluorescence microplate reader.
Acridine orange/ethidium bromide (AO/EB)
staining
Exponentially growing cells were seeded on
polylysine-coated glass coverslip in a 24-well-plate and cultured
at 37°C, under 5% CO2 for 24 h. After incubation with 40
μg/ml DMMP, cells were washed with PBS for 3 times and then stained
with 100 μg/ml AO/EB for 5 min. Coverslips were mounted and the
fluorescence was visualized using fluorescence microscopy
(Olympus).
JC-1 analysis
Cells (104) were seeded on
polylysine-coated glass coverslips in a 24-well plate, and incubate
at 37°C under 5% CO2 for 24 h. After being exposed to
DMMP for 12 and 24 h, cells were washed with PBS for 3 times, and
then incubated with a final concentration of 2 μM JC-1 dye at 37°C
under 5% CO2 for 20 min. Cells were treated with a final
concentration of 50 μM CCCP at 37°C for 20 min as a positive
control. After 3 washes with PBS, the coverslips were mounted and
the fluorescence was visualized using fluorescence microscopy
(Olympus).
Flow cytometric assay
HeLa cells were treated with DMMP at a concentration
of 40 μg/ml for 12, 24 and 36 h. Cells (106)were
collected by centrifuging at 1,000 rpm for 5 min, and then cells
were washed twice with ice-cold PBS. For cell cycle analysis, cells
were fixed in ice-cold ethanol (70% v/v) and stained with 0.5 ml
PI/RNase staining buffer (BD Pharmingen) for 15 min at room
temperature and analyzed by flow cytometry (Becton-Dickinson,
Franklin Lakes, NJ, USA). Apoptotic/necrotic cells were detected
using the Annexin V-FITC Apoptosis Detection kit (BD Pharmingen).
Briefly, cells were incubated with binding buffer (10 mM
HEPES/NaOH, pH 7.5, 140 mM NaCl and 2.5 mM CaCl2) and
stained with PI and FITC-labeled Annexin V for 15 min at room
temperature in the dark. Cell fluorescence was evaluated by flow
cytometry using a FACSCalibur instrument and analyzed by Cell Quest
software.
Western blotting
Western blotting was used to determine the
expression of various key proteins. Cells were seeded in a culture
bottle at 37°C under 5% CO2 and 95% air. The cells were
treated with DMMP (40 μg/ml for 12, 24 and 36 h). Approximately
2×106 cells were collected and washed with ice-cold PBS.
Subsequently, cells were incubated in lysis buffer (50 mM
HEPES-NaOH, 100 mM NaCl, 0.5% NP-40, 2.5 mM EDTA, 10% glycerol, 1
mM DTT, 1 mM Na3VO4, 5 mM NaF, 1 mM PMSF, 2
μg/ml pepstatin, 5 μg/ml leupetin, 2 μg/ml aprotintin) for 10 min
on ice, collected by scraping and kept on ice for 30 min. Finally,
after freezing and thawing, cells were centrifuged at 14,000 rpm
for 30 min at 4°C. Cell lysates were centrifuged at 12,000 rpm for
15 min at 4°C. Protein concentrations in lysates were determined by
Bradford assay. Fifty micrograms of protein from the protein
lysates per sample was denatured in 2X sample buffer and was loaded
on sodium dodecyl sulfate-polyacrylamide (SDS) gel electrophoresis
on 8–12% Tris-glycine gel. After electrophoresis, proteins were
transferred onto PVDF membranes (Millipore) followed by blocking
with 5% non-fat milk powder (w/v) in Tris-buffered saline [10 mM
Tris-HCl (pH 7.5), 100 mM NaCl, 0.1% Tween-20] for 1 h at room
temperature. The membranes were then incubated with specific
primary antibodies for 1 h. After washing, the membranes were
incubated with appropriate secondary HRP-conjugated antibodies and
visualized by ECL. Each membrane was stripped and reprobed with
anti-GADPH antibody to ensure equal protein loading.
Real-time RT-PCR
Approximately 106 cells were harvested at
the indicated time points and total RNA was isolated using TRIzol
reagent (Invitrogen Life Technologies) as described by the
manufacturer. The integration of RNA was detected by agarose gel
analysis and spectrophotometry. Reverse transcription of total RNA
was performed by PrimeScript™ High Fidelity RT-PCR kit (Takara Bio,
Inc). Real-time RT-PCR was performed using a Bio-Rad (Hercules, CA,
USA) iCycler PCR machine. Each PCR mixture contained 100 ng of cDNA
template and primers at a concentration of 10 μM in a final volume
of 25 μl of SYBR Premix Dimer Eraser. Specificity of each PCR was
examined by the melting temperature profiles of the final products.
Standard curves were calculated using cDNA to determine the linear
range and efficiency of each primer pair (primer sequences are
provided upon request). Reactions were conducted in triplicate, and
relative amounts of gene were normalized to GAPDH. The relative
gene expression data were analyzed by the comparative CT method
(ΔΔCT method).
Statistical analysis
The statistical significance of the differences
between the control and treated groups was determined using the
2-tailed Student’s t-test. P-values <0.05 were considered to
indicate a statistically significant result.
Results
Effect of DMMP on cell proliferation
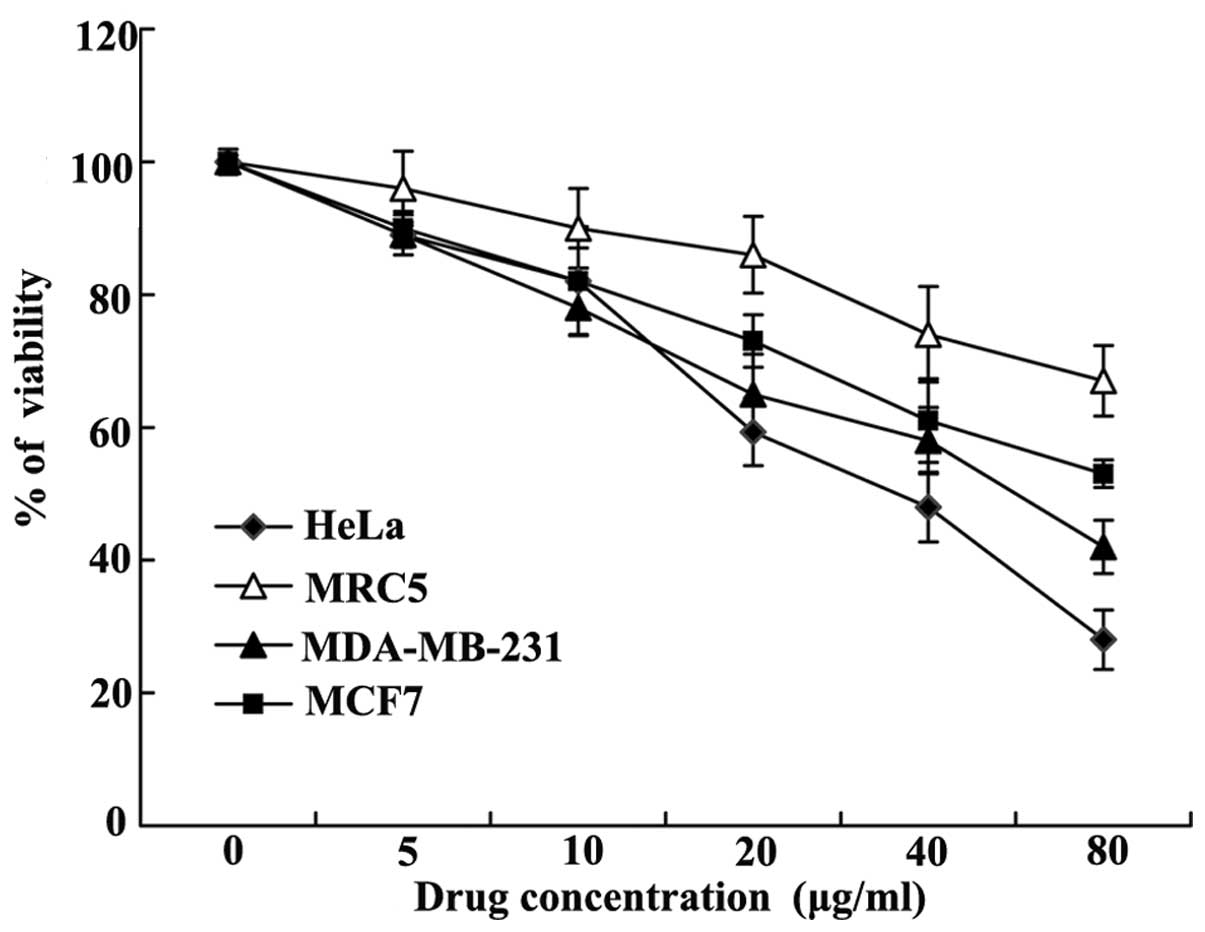
To assess the effect of DMMP on cell proliferation,
human cancer cell lines, HeLa, MCF7 and MDA-MB-231, and the normal
lung fibroblast cell line MRC5 were treated with increasing
concentrations of DMMP at 24 h, and cell viability was assessed
using the MTT assay (Fig. 2).
IC50 for HeLa, MDA-MB-231, MCF7 and MRC5 cells were 36,
51, 81 and 147 μg/ml, respectively. DMMP inhibited the
proliferation of the cancer cells in a concentration-dependent
manner. DMMP showed the most antiproliferative effect on the HeLa
cell line. While the normal lung fibroblast cell line MRC5
exhibited less sensitivity. DMMP exhibited slight selective
cytotoxic effect.
Effect of DMMP on cell cycle
progression
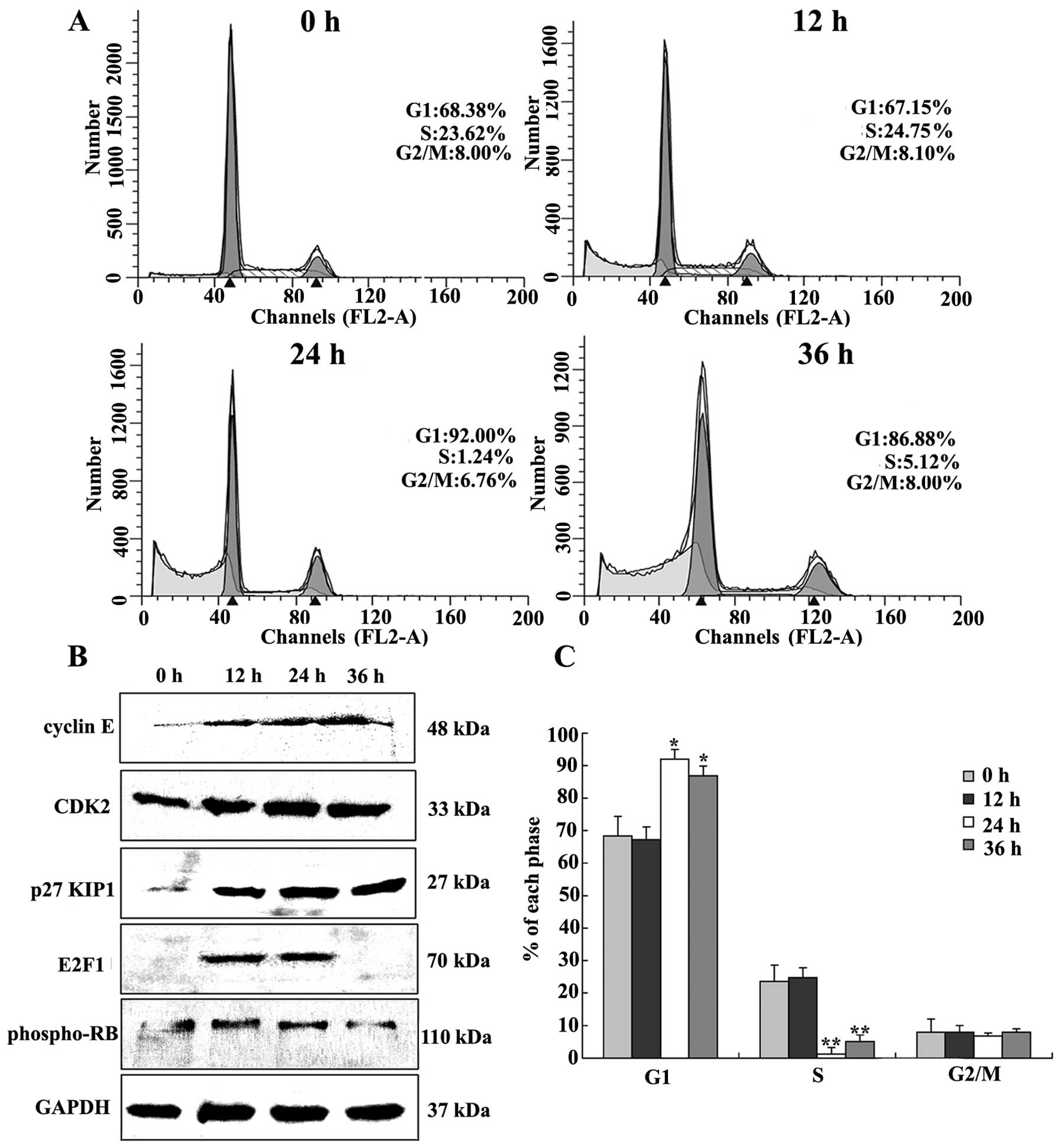
The effect of DMMP treatment on cell cycle arrest
was examined. Compared to DMSO treatment, cells treated with DMMP
at the indicated times notably accumulated in the G1 phase of the
cell cycle with a reduction in the percentage of cells in S phase
(Fig. 3A). DMMP treatment for 24 h
increased the percentage of cells in the G1 phase from 68.38 to
92.0%. These results suggest that DMMP inhibited cellular
proliferation of HeLa cells via the G1 phase arrest of the cell
cycle.
The expression of cell cycle-regulating proteins was
examined. Expression levels of cyclin E were gradually increased
after DMMP treatment, with slight upregulation of CDK2 (Fig. 3B). p27KIP1 was able to
directly bind several different classes of cyclins and CDKs leading
to cell cycle arrest in the G1 phase (19). As shown in Fig. 3B, p27KIP1 was
dramatically upregulated at 12 h and this effect increased over
time. pRb is an important regulator of G1/S phase transition.
Phosphorylated pRb was slightly increased (Fig. 3B). E2F1, the binding partner of pRb,
was increased at 12 and 24 h.
DMMP induces apoptosis in HeLa cancer
cells
To further investigate the mechanism of the
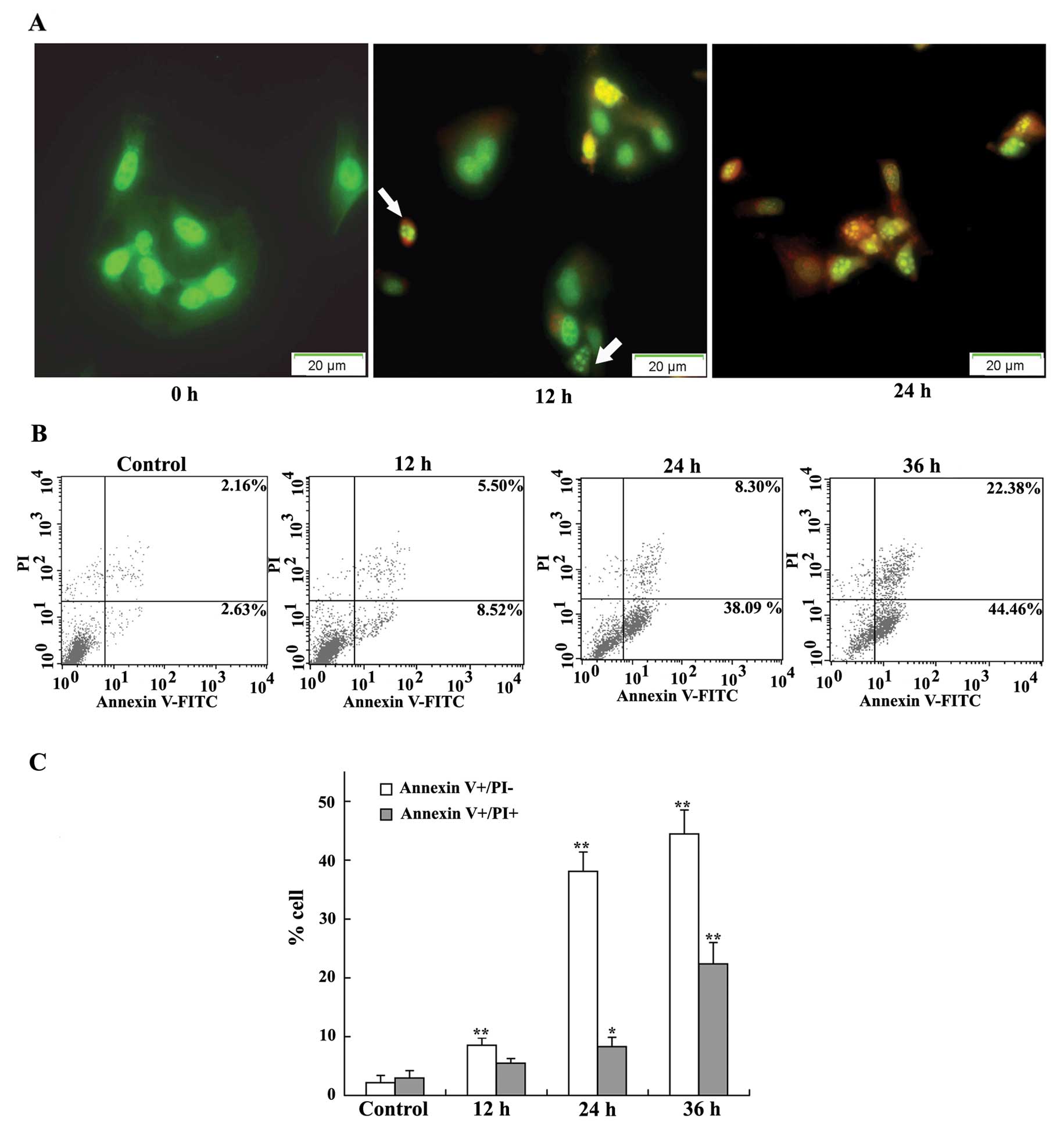
inhibitory effect of DMMP, acridine orange/ethidium bromide (AO/EB)
staining was used to visualize nuclear changes and apoptotic body
formation which are characteristic of apoptosis. AO is a
membrane-permeable dye that can pass through the membrane of intact
cells and embed in the nuclear DNA emitting bright green
fluorescence. EB can pass through the membrane of a dead or dying
cell with orange fluorescence. After HeLa cells were exposed to
DMMP for 12 h, early apoptotic cells with green pyknotic-like
nuclear chromatin were noted. At 24 h, late apoptotic cells with
pyknotic-like deep orange nuclear chromatin were detected. Control
cells with normal morphology showed green fluorescence (Fig. 4A). The results suggest that DMMP was
able to induce marked apoptotic morphology in HeLa cells in a
time-dependent manner.
To investigate the kinetics of the apoptotic cells
induced by DMMP, HeLa cells were treated with DMMP for various
periods of time, and the apoptotic cell death was analyzed by flow
cytometry using Annexin V-PI staining. The proportion of early
apoptotic cells (Annexin V-positive and PI-negative) was
significantly increased from 2.63% in the control cells to 8.52% at
12 h, 38.09% at 24 h, and 44.46% at 36 h following DMMP treatment
(Fig. 4B and C). These results
indicate that cell death induced by DMMP was mainly caused by
induction of apoptosis in a time-dependent-manner.
Caspase activity is induced by DMMP
We investigated whether DMMP induces HeLa cell
apoptosis through the caspase pathway. Caspase-8, -9 and -3 were
detected by western blotting as shown in Fig. 5A. Following DMMP treatment
full-length caspase-9 was decreased in a time-dependent manner.
Cleaved caspase-9 was detected indicating activation of caspase-9
and cleaved caspase-3 was also detected, while activated caspase-8
was not detected (Fig. 5A).
Meanwhile, expression of the death receptor, Fas, was unchanged and
its ligand, FasL was slightly increased (Fig. 5A). Survivin expression was gradually
increased and peaked at 24 h and AIF was also slightly increased in
DMMP-induced apoptotic cell death (Fig.
5A).
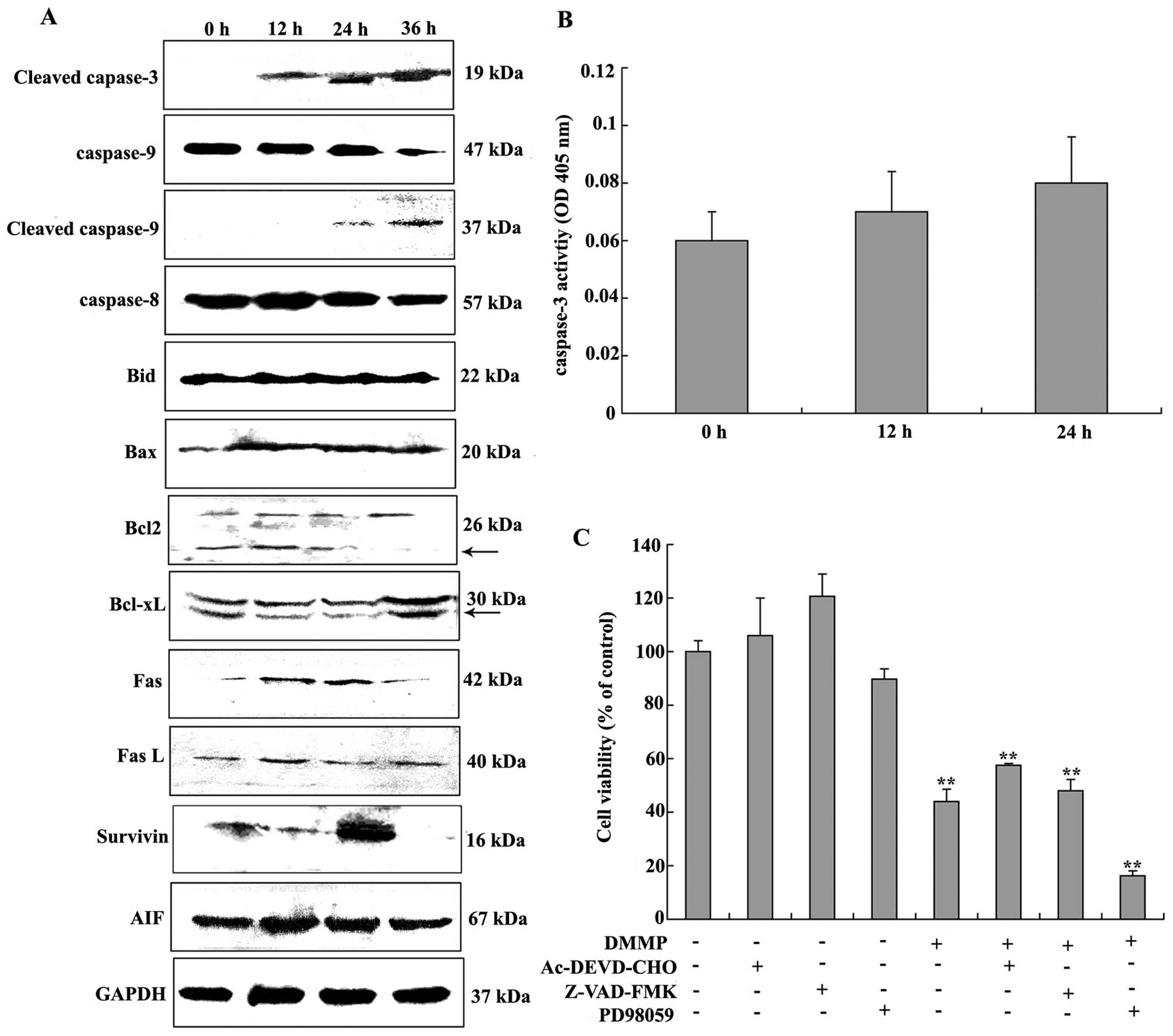 | Figure 5Activation of caspases induced by
DMMP. (A) HeLa cells were treated with or without 40 μg/ml DMMP for
different time periods. The cells were harvested and proteins were
separated by SDS-PAGE and transferred onto PVDF membranes. The
expression of apoptosis-related proteins was detected by western
blotting probed with cleaved caspase-3, -9 and -8, Bid, Bax, Bcl-2,
Bcl-xL, Fas, FasL, survivin, AIF and GAPDH antibodies. GAPDH was
used as a loading control for western blotting. Arrowheads indicate
the non-specific bands. (B) Activation of caspase-3 by DMMP. HeLa
cells were incubated with 40 μg/ml DMMP for 12 and 24 h before the
caspase-3 substrate Ac-DEVD-pNA was added. Incubation of the plate
was carried out at 37°C for 2 h, and the optical density (OD) was
read on a fluorescence microplate reader. (C) Effects of caspase
inhibitors and an ERK inhibitor on apoptosis induced by DMMP. HeLa
cells were pretreated with the broad-spectrum caspase inhibitor
(Z-VAD-FMK) and caspase-3 inhibitor (Ac-DEVD-CHO) at 10 μM and ERK
inhibitor (PD98059) at 20 μM for 2 h followed by incubation with 40
μg/ml DMMP for 24 h. The cell viability was evaluated using the MTT
assay. Error bars represent ± SD from triplicate independent
experiments at each time point compared to the control group;
*P<0.05, **P<0.01. |
Caspase-3 can be activated either in the intrinsic
or the extrinsic apoptotic pathway. To further identify whether
caspase-3 is involved in DMMP-induced apoptosis, the catalytic
activity of caspase-3 was measured using a caspase colorimetric
assay kit. Caspase activity was not significantly increased when
compared to the control after DMMP treatment (Fig. 5B). To further evaluate the role of
caspases in DMMP-induced apoptosis pathway, HeLa cells were
pretreated in the presence of the broad-spectrum caspase inhibitor
(Z-VAD-FMK) and caspase-3 inhibitor (Ac-DEVD-CHO) for 2 h before
drug treatment. Pretreatment of caspase-3 and broad-spectrum
caspase inhibitors failed to attenuate cell death following DMMP
treatment (Fig. 5C). These results
showed that DMMP-induced apoptosis may be through a
caspase-independent mechanism.
DMMP induces loss of mitochondrial
transmembrane potential (ΔΨm)
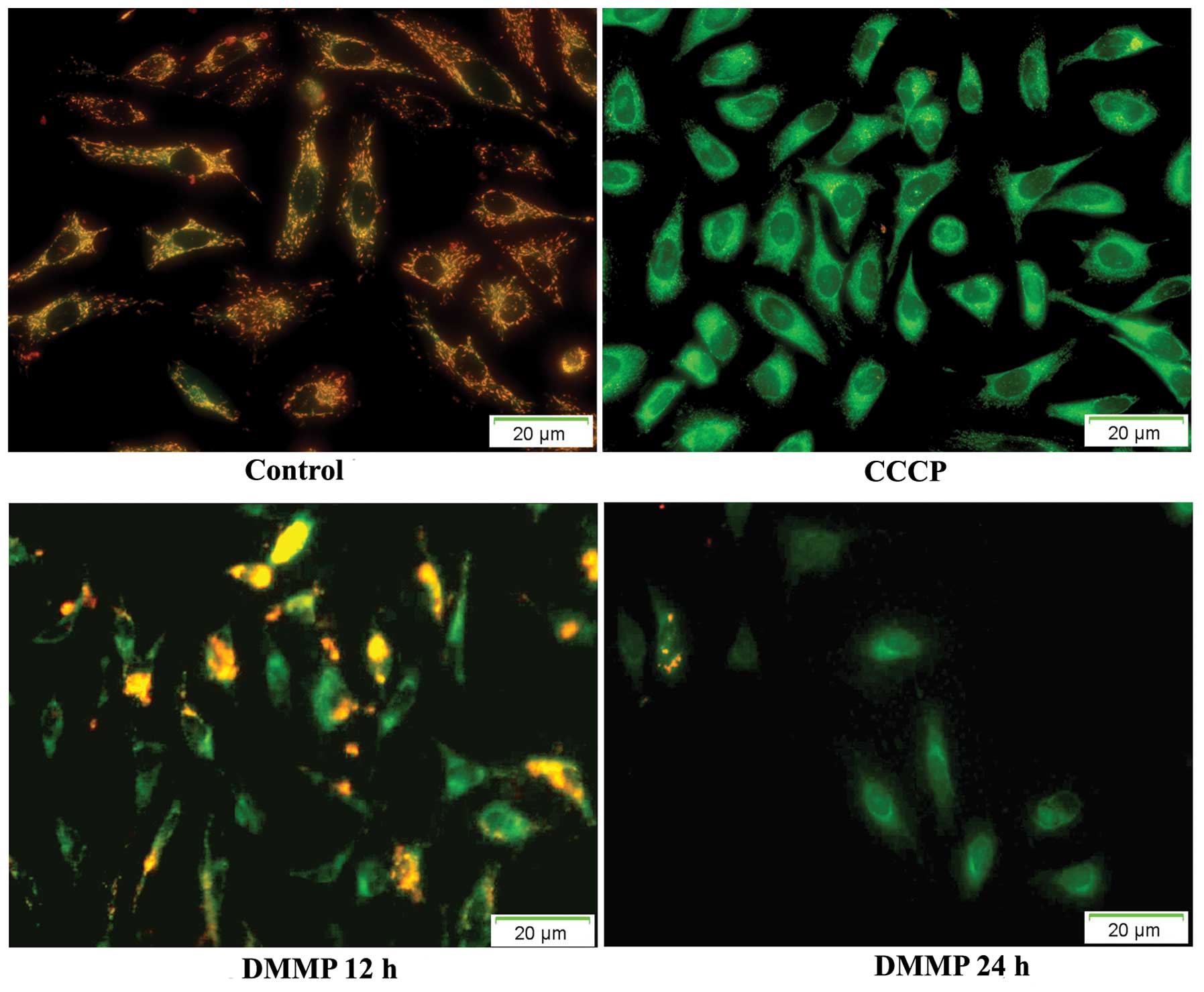
Loss of mitochondrial transmembrane potential is one
of several key events that occur in the mitochondria during
apoptosis. JC-1 is the most commonly used fluorescent probe to
detect the change in mitochondrial membrane potential (ΔΨm). In
healthy cells with high ΔΨm, JC-1 spontaneously shows intense red
fluorescence and in contrast in apoptotic or unhealthy cells with
low ΔΨm, JC-1 shows only green fluorescence. In untreated HeLa
cells, JC-1 showed intense red fluorescence. At 12 h after drug
treatment, green fluorescence was noted in some cells indicating
the dissipation of ΔΨm. At 24 h, most cells exhibited green
fluorescence (Fig. 6). The time
course analysis revealed that significant numbers of cells
following treatment with DMMP lost ΔΨm.
Effect of DMMP on the Bcl-2 family
Bcl-2 family proteins are key regulators of
mitochondrial permeability. Therefore, we investigated whether
apoptosis in HeLa cells induced by DMMP is modulated by Bcl-2
family members. Western blotting results revealed that pro-survival
Bcl-2 family proteins, Bcl-2, was decreased at 36 h, and Bcl-xL was
decreased at 12 and 24 h (Fig. 5A).
Bcl-2 mRNA expression significantly increased at 12 h (2.5-fold )
and 24 h (2.2-fold), and then decreased at 36 h after treatment
consistent with the results of the western blotting. Meanwhile, a
pro-apoptotic Bcl-2 family member, Bax, was significantly increased
(Fig. 5A). The mRNA expression
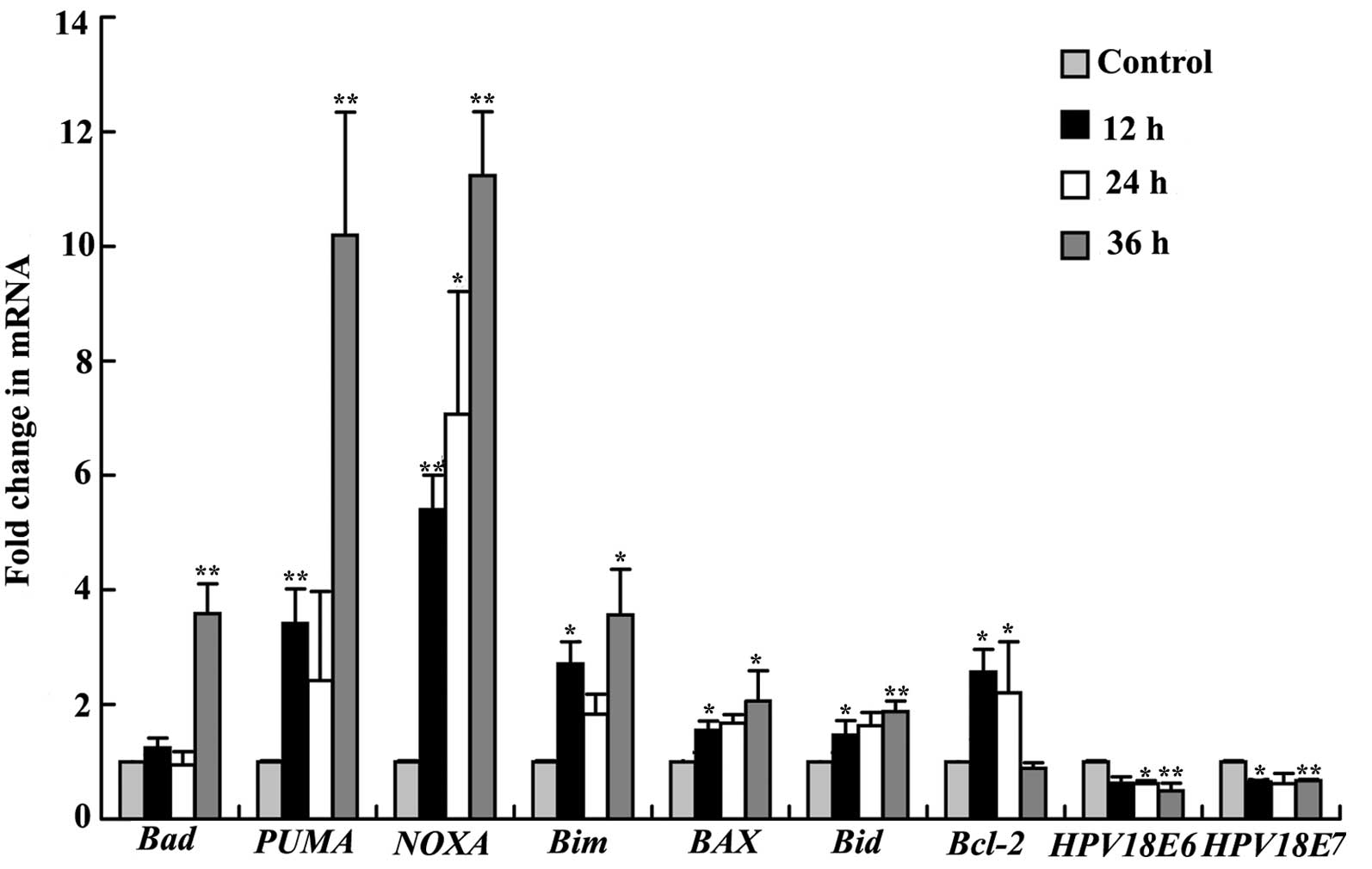
levels of PUMA, NOXA, Bax, Bad and Bid were
significantly upregulated after treatment (Fig. 7). The mRNA expression levels of
PUMA and NOXA were significantly increased (3.4- and
5.4-fold, respectively) compared with the control at 12 h and
achieved a maximum increase at 36 h after treatment (10.1- and
11.2-fold). Bax and Bid mRNA expression was
significantly increased at 24 h after treatment (2.0- and 1.8-fold,
respectively). Bim and Bad mRNA expression was
significantly increased only at 36 h (3.5- and 3.6-fold,
respectively).
Effect of DMMP on p53 and p73
expression
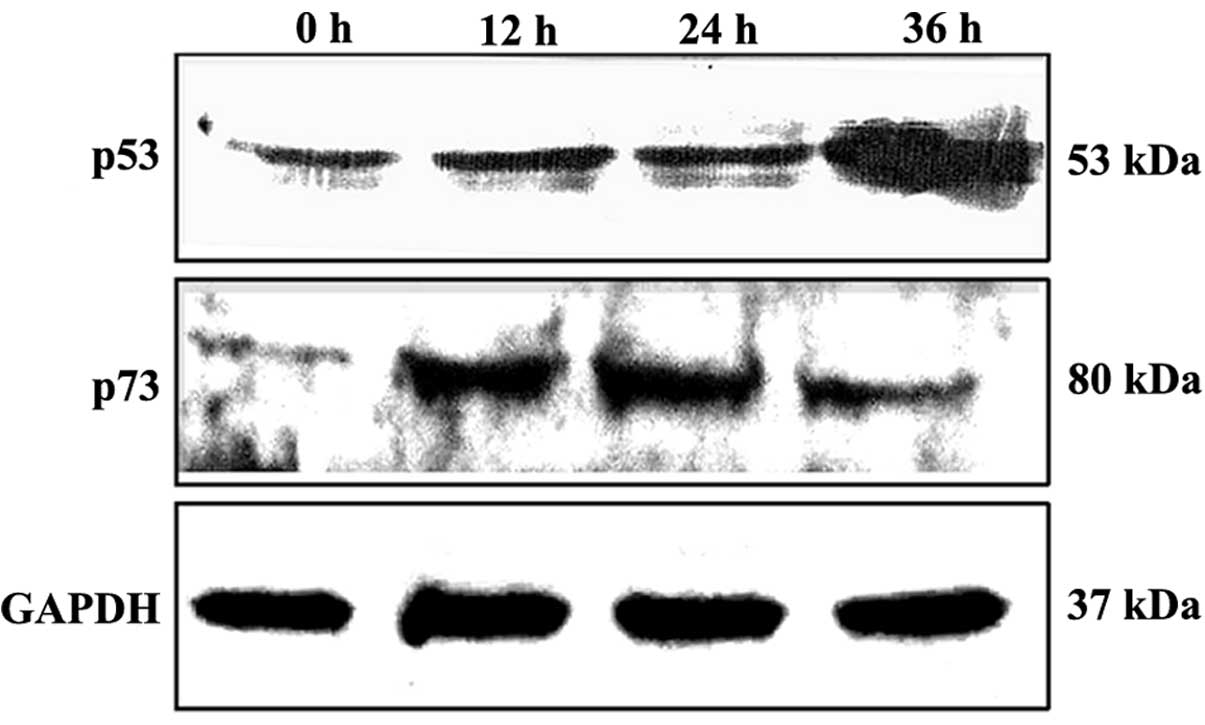
To investigate whether DMMP has an effect on p53
protein expression, we determined the p53 levels in HeLa cells
treated with DMMP at the indicated time periods. Western blot
analysis showed that DMMP treatment resulted in a time-dependent
accumulation of p53 in HeLa cells (Fig.
8). Another p53 family member, p73, was also increased.
Moreover, HeLa cells contain HPV E6–E7 genes which promote the
degradation of p53 and the retinoblastoma protein. HPV E6–E7 mRNA
levels were decreased by 50% following the drug treatment (Fig. 7).
Effect of DMMP on ERK and Akt
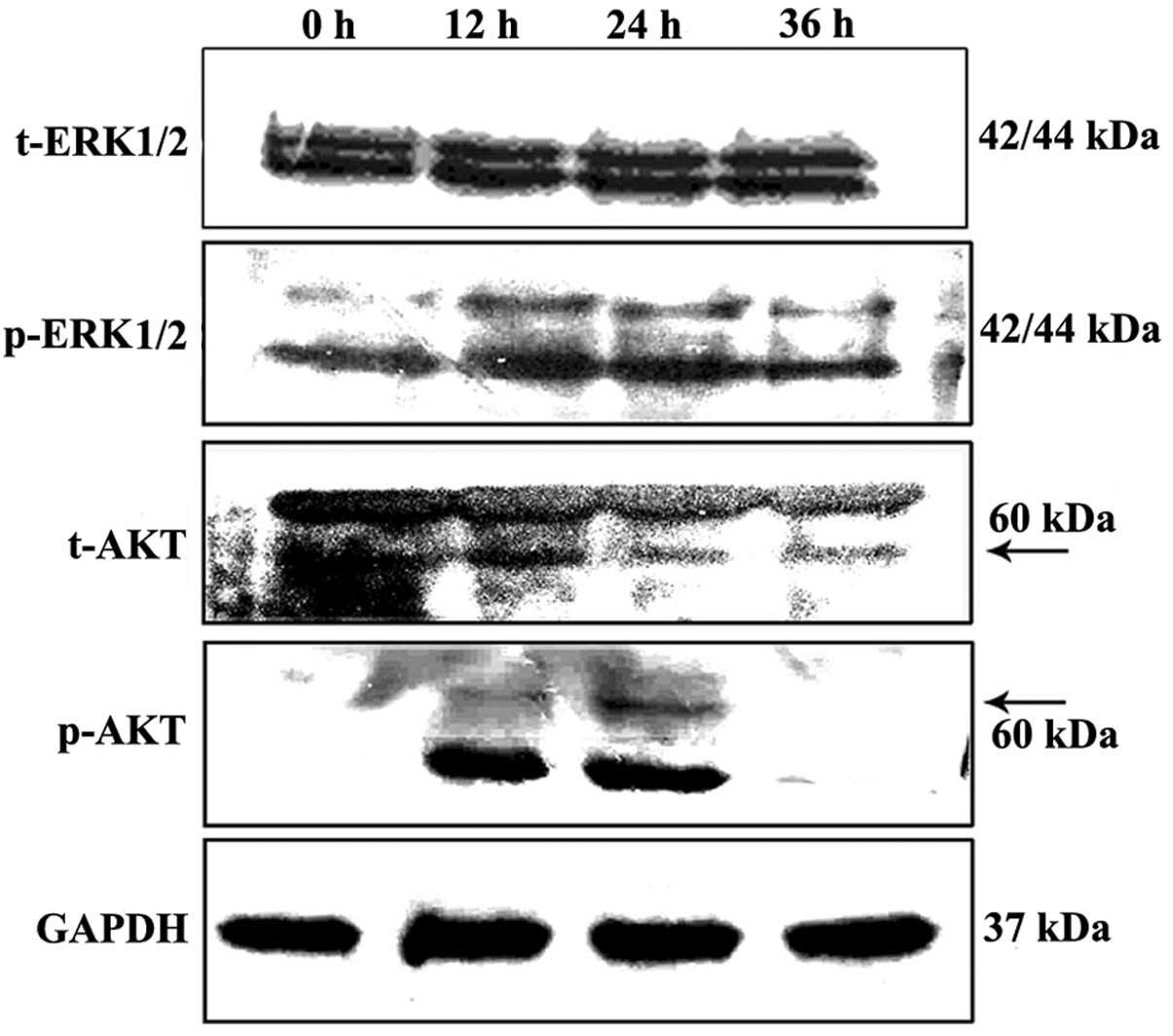
Western blot analysis showed that treatment with
DMMP led to a slight upregulation of p-ERK in the HeLa cells
following treatment while levels of total extracellular
signal-regulated protein kinases 1 and 2 (ERK1/2) were not altered
(Fig. 9). To further demonstrate
the functions of ERK in inducing apoptosis, the effects of the
highly specific ERK inhibitor PD98059 were examined. As shown in
Fig. 5C, PD98059 significantly
increased cell death. This indicates that sustained phosphorylation
of ERK inhibits DMMP-induced cell death.
Akt is a serine/threonine kinase involved in
cellular survival pathways by inhibiting apoptotic processes. To
determine whether Akt is involved in DMMP-induced apoptosis, the
phosphorylation status of Akt was examined by western blot
analysis. Akt phosphorylation was significantly increased at 12 h
and declined at 36 h, while the level of total Akt protein was not
altered (Fig. 9).
Discussion
During a search for novel anticancer agents in the
culture broth of various endophytic fungi, DMMP was isolated from
the endophytic fungus Pestalotiopsis photiniae. In the
present study, DMMP inhibited the proliferation of various cancer
cell lines and had little effect on normal lung MRC5 cells. As
compared with the tumor cell lines analyzed, we found that DMMP had
selective cytotoxic effect. Thus, the molecular mechanism of the
cytotoxic effect of DMMP was investigated in HeLa cells.
DMMP caused cell cycle arrest in the G1 phase of the
cell cycle. DMMP significantly elevated p27KIP1
expression (Fig. 3).
p27KIP1 causes cell cycle arrest in G1 phase by binding
cyclin E-CDK2 complexes (20–22).
After DMMP treatment, phospho-Rb was slightly elevated, and E2F1
expression was significantly increased at 12 and 24 h. Meanwhile,
CDK2 and cyclin E proteins were also increased (Fig. 3B). The retinoblastoma protein (Rb)
exhibits growth inhibitory activity by binding with E2F1. The
Rb-E2F1 pathway plays an important role in the regulation of cell
cycle progression. Cyclin E-CDK2 can phosphorylate Rb resulting in
the activation of E2F-responsive genes (23). Whether the increased levels of CDK2
and cyclin E proteins facilitated E2F function requires further
investigation.
DMMP efficiently induced HeLa cell apoptosis
characterized by compaction and fragmentation of nuclear chromatin.
To further confirm the effects on apoptosis, FITC Annexin V and PI
double staining and flow cytometry were performed. Results showed
that DMMP induced apoptosis in the HeLa cells. Depletion of ΔΨm in
HeLa cells was also detected after treatment. The mitochondrial
apoptotic pathway is mainly regulated by Bcl-2 family proteins. Any
imbalance in the expression level of pro-apoptotic and
anti-apoptotic Bcl-2 members leads to the disruption of the outer
mitochondrial membrane (24,25).
DMMP significantly upregulated the mRNA levels of pro-apoptotic
genes such as Bax, PUMA, NOXA, Bim, Bid and Bad (3-
to 11-fold) (Fig. 7). Meanwhile,
Bcl-2 and Bcl-xL levels were decreased. Bax protein was
significantly increased (Fig. 5A),
and Bcl-2 mRNA expression was increased by 2-fold and declined at
36 h. As a result of these changes, the ratios of anti-apoptotic
proteins and pro-apoptotic proteins of the Bcl-2 family were
significantly reduced during apoptosis. The imbalance led to the
loss of ΔΨm after DMMP treatment.
The extrinsic apoptosis pathway is indicated by the
ligation of cell surface death receptors to their specific ligands
such as Fas/FasL (26). The
formation of active caspase-8 is involved in the extrinsic
apoptosis pathway. Extrinsic apoptosis in some cells is dependent
on the cleavage of Bid. In our study, although FasL was slightly
increased, we did not detect the activation of caspase-8 and the
truncated form of Bid (Fig. 5A).
These results showed that DMMP-induced apoptosis did not occur
through the Fas/FasL extrinsic apoptosis pathway.
Sequential activation of caspases plays a central
role in the execution-phase of cell apoptosis. As an effector
caspase, caspase-3 plays a central role in the extrinsic apoptotic
pathway and intrinsic apoptotic pathway. In our study, although
cleaved caspase-3 and cleaved caspase-9 were detected by western
blotting, no significant activation of caspase-3 proteolytic
activity was detected after drug treatment. Survivin, which
functions to inhibit caspase activation, was elevated after drug
treatment. Elevated survivin may inhibit caspase-3 proteolytic
activity. Further experiments showed that pretreatment with caspase
inhibitors (Ac-DEVD-CHO and Z-VAD-FMK) failed to attenuate
DMMP-induced cell death (Fig. 5C).
DMMP induced a slightly higher level of AIF after drug treatment
compared with the control (Fig.
5A). Although caspase activation is considered a hallmark of
apoptotic cell death, caspase-independent apoptosis has been
reported. Apoptosis-induced factors (AIF) and endonuclease G
(EndoG) residing in mitochondria are involved in the
caspase-independent cell death pathway (27). DMMP may induce apoptosis in a
caspase-independent manner. Further experimental investigation is
needed for confirmation.
The tumor suppressor transcription factor p53 plays
a vital role in cell cycle arrest and apoptosis in response to
cellular stress. Western blot analysis showed that DMMP treatment
resulted in a significant accumulation of p53 and p73 proteins in
HeLa cells (Fig. 8). Normally, p53
protein is rapidly degraded by the ubiquitin-proteome system. Mdm2
is an important negative regulator of the p53 tumor suppressor
(28,29). But in HPV-infected cervical cancer
cells, HPV E6 is the protein responsible for repressing p53
replacing Mdm2 function (30). In
the present study, real-time RT-PCR showed that DMMP caused a ~50%
reduction of E6 and E7 mRNA expression when compared to the control
(Fig. 7). The dramatic upregulation
of p53 protein may be due to the low expression of HPV E6.
In large part, the potent anticancer activity of p53
has been linked to its ability to induce apoptosis through the
intrinsic mitochondrial-mediated apoptotic pathway (31). p53 enhances the expression of Bcl-2
family members including Bax, Bid, PUMA and Noxa (32). BH3-only proteins, NOXA and PUMA, are
important mediators of p53-induced apoptosis (33). E2F1 is the first member of the E2F
transcription factor family. Evidence shows that E2F1 also
possesses tumor suppressor functions which can induce apoptosis via
p53-dependent and p53-independent pathways (34). Oncogenic signaling by E2F1 has
recently been linked to stabilization and activation of the tumor
suppressor p53 (35). Puma and Noxa
can be regulated by both p53 and E2F1. In our study, E2F1 and p53
were both increased after DMMP treatment. The mRNA levels of
Bax, Puma, Noxa and Bid genes were significantly
increased (Fig. 7). Whether p53 and
E2F1 were involved in DMMP-induced apoptosis requires further
investigation using p53 or E2F1 knockdown cells.
Extracellular signal-regulated protein kinases 1 and
2 (ERK1/2) are members of the mitogen-activated protein kinase
superfamily that can mediate cell proliferation and apoptosis
(36). PhosphoERK1/2 was slightly
increased after DMMP treatment (Fig.
8). Treatment with the ERK inhibitor PD98059 and DMMP
efficiently induced HeLa cell death. This implied that DMMP
combined with PD98059 may represent a novel anticancer strategy.
The Akt serine/threonine kinases are critical regulators of cell
survival. The activation of the PI3K-Akt pathways promotes
tumorigenesis by inhibiting apoptotic processes (37). Following DMMP treatment,
phospho-Akt, increased at 12 and 24 h, was slightly detected at 36
h while total Akt was unaltered (Fig.
9). The reason for the elevated phosphorylation of Akt at an
early stage requires further investigation.
In conclusion, for the first time we demonstrated
that DMMP significantly induced HeLa cell apoptosis and caused cell
cycle arrest in G1 phase. We investigated the possible mechanisms
involved in DMMP-induced apoptosis. We demonstrated that
DMMP-induced apoptosis was through the mitochondrial intrinsic
pathway in HeLa cells. DMMP upregulated p27 protein, and caused
cell cycle arrest in the G1 phase. The p53 family members, p53 and
p73, were increased after drug treatment. Our results indicate that
DMMP is a promising cancer therapeutic agent.
Acknowledgements
This study was supported by programs for New Century
Excellent Talents in University (NCET-09-0112), the National
Natural Science Foundation of China (30901755 and 31171885), the
Scientific Research Program of Hebei Provincial Education Bureau
(2011107), the Hebei Province Science Fund for Distinguished Young
Scholars (C2011201113), and the Program for Changjiang Scholars and
Innovative Research Team in the University (IRT1124).
References
|
1
|
Koehn FE and Carter GT: The evolving role
of natural products in drug discovery. Nat Rev Drug Discov.
4:206–220. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Miller KI, Qing C, Sze DM and Neilan BA:
Investigation of the biosynthetic potential of endophytes in
traditional Chinese anticancer herbs. PLoS One. 7:e359532012.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tan RX and Zou WX: Endophytes: a rich
source of functional metabolites. Nat Prod Rep. 18:448–459.
2001.PubMed/NCBI
|
|
4
|
Reed JC: Drug insight: cancer therapy
strategies based on restoration of endogenous cell death
mechanisms. Nat Clin Pract Oncol. 3:388–398. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Frankfurt OS and Krishan A:
Apoptosis-based drug screening and detection of selective toxicity
to cancer cells. Anticancer Drugs. 14:555–561. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen F, Wang W and El-Deiry WS: Current
strategies to target p53 in cancer. Biochem Pharmacol. 80:724–730.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Collavin L, Lunardi A and Del Sal G:
p53-family proteins and their regulators: hubs and spokes in tumor
suppression. Cell Death Differ. 17:901–911. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ferreira CG, Tolis C and Giaccone G: p53
and chemosensitivity. Ann Oncol. 10:1011–1021. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lunghi P, Costanzo A, Mazzera L, Rizzoli
V, Levrero M and Bonati A: The p53 family protein p73 provides new
insights into cancer chemosensitivity and targeting. Clin Cancer
Res. 15:6495–6502. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bisso A, Collavin L and Del Sal G: p73 as
a pharmaceutical target for cancer therapy. Curr Pharm Des.
17:578–590. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Slade N and Horvat A: Targeting p73 - a
potential approach in cancer treatment. Curr Pharm Des. 17:591–602.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lain S, Hollick JJ, Campbell J, et al:
Discovery, in vivo activity, and mechanism of action of a
small-molecule p53 activator. Cancer Cell. 13:454–463. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Vassilev LT, Vu BT, Graves B, et al: In
vivo activation of the p53 pathway by small-molecule antagonists of
MDM2. Science. 303:844–848. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Peirce SK and Findley HW: The MDM2
antagonist nutlin-3 sensitizes p53-null neuroblastoma cells to
doxorubicin via E2F1 and TAp73. Int J Oncol. 34:1395–1402.
2009.PubMed/NCBI
|
|
15
|
Sampath D, Calin GA, Puduvalli VK, et al:
Specific activation of microRNA106b enables the p73 apoptotic
response in chronic lymphocytic leukemia by targeting the ubiquitin
ligase Itch for degradation. Blood. 113:3744–3753. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Phuwapraisirisan P, Rangsan J, Siripong P
and Tip-Pyang S: New antitumour fungal metabolites from
Alternaria porri. Nat Prod Res. 23:1063–1071. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Suemitsu R, Ohnishi K, Morikawa Y and
Nagatomo S: Zinnimidine and
5-(3′,3′-dimethylallyloxy)-7-methoxy-6-methylphthalide from
Alternaria porri. Phytochemistry. 38:495–497.
1995.PubMed/NCBI
|
|
18
|
Yang XL, Zhang S, Hu QB, Luo DQ and Zhang
Y: Phthalide derivatives with antifungal activities against the
plant pathogens isolated from the liquid culture of
Pestalotiopsis photiniae. J Antibiot (Tokyo). 64:723–727.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Malumbres M and Barbacid M: To cycle or
not to cycle: a critical decision in cancer. Nat Rev Cancer.
1:222–231. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hengst L and Reed SI: Translational
control of p27KIP1 accumulation during the cell cycle.
Science. 271:1861–1864. 1996. View Article : Google Scholar
|
|
21
|
Pagano M, Tam SW, Theodoras AM, et al:
Role of the ubiquitin-proteasome pathway in regulating abundance of
the cyclin-dependent kinase inhibitor p27. Science. 269:682–685.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Vervoorts J and Lüscher B:
Post-translational regulation of the tumor suppressor p27(KIP1).
Cell Mol Life Sci. 65:3255–3264. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Harbour JW and Dean DC: The Rb/E2F
pathway: expanding roles and emerging paradigms. Genes Dev.
14:2393–2409. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Danial NN and Korsmeyer SJ: Cell death:
critical control points. Cell. 116:205–219. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chipuk JE and Green DR: How do BCL-2
proteins induce mitochondrial outer membrane permeabilization?
Trends Cell Biol. 18:157–164. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ashkenazi A and Dixit VM: Death receptors:
signaling and modulation. Science. 281:1305–1308. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chipuk JE and Green DR: Do inducers of
apoptosis trigger caspase-independent cell death? Nat Rev Mol Cell
Biol. 6:268–275. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Brooks CL and Gu W: p53 ubiquitination:
Mdm2 and beyond. Mol Cell. 21:307–315. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sullivan KD, Gallant-Behm CL, Henry RE,
Fraikin JL and Espinosa JM: The p53 circuit board. Biochim Biophys
Acta. 1825:229–244. 2012.PubMed/NCBI
|
|
30
|
Diaz D, Santander MA and Chavez JA: HPV-16
E6 and E7 oncogene expression is downregulated as a
result of Mdm2 knockdown. Int J Oncol. 41:141–146. 2012.
|
|
31
|
Chipuk JE and Green DR: Dissecting
p53-dependent apoptosis. Cell Death Differ. 13:994–1002. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hemann MT and Lowe SW: The p53-Bcl-2
connection. Cell Death Differ. 13:1256–1259. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Villunger A, Michalak EM, Coultas L, et
al: p53- and drug-induced apoptotic responses mediated by BH3-only
proteins puma and noxa. Science. 302:1036–1038. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wu Z, Zheng S and Yu Q: The E2F family and
the role of E2F1 in apoptosis. Int J Biochem Cell Biol.
41:2389–2397. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Stiewe T and Pützer BM: Role of the
p53-homologue p73 in E2F1-induced apoptosis. Nat Genet. 26:464–469.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mebratu Y and Tesfaigzi Y: How ERK1/2
activation controls cell proliferation and cell death: is
subcellular localization the answer? Cell Cycle. 8:1168–1175. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Amin AR, Paul RK, Thakur VS and Agarwal
ML: A novel role for p73 in the regulation of Akt-Foxo1a-Bim
signaling and apoptosis induced by the plant lectin, Concanavalin
A. Cancer Res. 67:5617–5621. 2007. View Article : Google Scholar : PubMed/NCBI
|