Introduction
Lung cancer is a malignant tumor with some of the
highest incidence and mortality rates worldwide; moreover, for most
patients, radical cure operation is not an option when the symptoms
appear and the disease is found, therefore, their prognosis is very
poor (1). The efficiency of
chemotherapy and radiotherapy is low and the considerable
side-effects are too strong for some patients to endure (2). With the advancements in the research,
a significant number of drugs against lung cancer are being
developed and although considerable achievements have been made, a
lot remains to be studied in order to control lung cancer.
Telomeres, the end of eukaryote linear chromosomes
consisting of tandem arrays of telomeric repeats, protect the
genome from degradation. In somatic cells, telomeres shorten with
each cell division due to the ‘end-replication-problem’. Critically
short telomeres induce an irreversible exit from the cell cycle,
known as senescence (3). Thus,
telomere loss is considered as a ‘mitotic clock’ that limits the
proliferation capacity of somatic cells. However, in immortal
cells, including tumor cells, telomerase provides a means to
replace telomere repeats which are lost during replication as a
result of the inability of DNA polymerase to replicate to the end
of linear chromosomes. Telomerase activity not only maintains the
telomeres of proliferating cells but is implicated in the process
of cellular immortalization and oncogenesis (4). In particular, tumor cells with
positive expression of telomerase have undergone senescence and
have obtained a considerable ability to proliferate.
Inhibition of telomerase has been discussed as a
promising approach for treating a variety of malignant tumors, as
the prerequisites agreed that telomerase was the main mechanism of
telomere maintenance (5). However,
it was independent of (uncoupled from) the proliferation of tumor
cells even if telomerase activity is diminished. In other words,
there is a lag phase between the times telomerase is inhibited and
when telomeres shorten sufficiently to cause cellular crisis with
proliferation strangled, and no host can suffer from so much
neoplastic burden so many times (6).
In this case, another factors of regulator for
telomere maintenance, tankyrase was advert, which poly
(ADP-ribosyl) ate telomere-repeat-binding factor 1 (TRF1) and
releases it from telomeres, allowing access of telomerase to
telomeres and enhancing telomere elongation. Conversely, it was
possible that the combination of both enzyme inhibitors can
increase the risk of critically shortened telomere and promote the
following crisis and mortality of lung cancer cells (7).
Materials and methods
Mice and cell lines
BALB/c nude mice, 6–8 weeks old, were purchased from
the Center of Medical Experimental Animals of Hubei Province
(Wuhan, China). The animals were housed and used for studies
approved by the Animal Care and Use Committee of the central
hospital of Wuhan. A549 human lung adenocarcinoma cell lines were
obtained from the China Center for Type Culture Collection (CCTCC)
in Wuhan University, China. After thawing, cells were transferred
into culture flasks containing RPMI-1640 (Gibco) supplemented with
1% penicillin-streptomycin and 10% fetal bovine serum (FBS), and
propagated in a humidified 5% CO2 incubator at 37°C. The
exponentially growing cells were harvested when reaching 80–90%
confluence.
Oligonucleotide synthesis and preparation
of reagents
An optimal target is a 25-base sequence that lies
within the region from the 5′-cap through the first 25 bases of
coding sequence, has ~50% GC content and has little or no secondary
structure (7). All oligonucleotides
in these experiments were designed by a computational neural
network. BLAST confirmed they are specific for their mRNA
(http://www.ncbi.nlm.nih.gov/BLAST/).
The oligonucleotides were synthesized by Sangon Biotechnology
Engineering Company of Shanghai, China, including antisense
oligonucleotides for human telomerase reverse transcriptase
(ashTERT) and tankyrase (asTANKS), and corresponding sense
oligonucleotides for human telomerase reverse transcriptase
(shTERT) and tankyrase (sTANKS) used as control as well. The
sequences of oligonucleotides were: ashTERT,
5′-GGAGCGCGCGGCATCGCGGG-3′; asTANKS,
5′-CATCTTCGGACTCCCCTAGCACTG-3′; shTERT, 5′-CCCGCGATGCCGCGCGCTCC-3′;
sTANKS, 5′-CAGTGCTAGGGGAGTCCGAAGATG-3′. The synthesized
oligonucleotides were modified by phosphorothiolation, purified by
HPLC, stored at −20°C.
Oligofectamin Reagent (Invitrogen) was applied in
order to improve oligonucleotide merging with cell membrane. RNA
TRIzol Reagent was obtained from Invitrogen, First Strand cDNA
Synthesis Reagent was purchased from Gibco, TRAPeze ELISA
Telomerase Detection kit was obtained from Millipore, Telomere PNA
kit was provided by Dako and Quantum premixed fluorescein
isothiocyanate (FITC) MESF beads were purchased by Flow Cytometry
Standards Co.
Transfection of oligonucleotides
Owing to different oligonucleotides, A549 cells were
divided into 6 groups: shTERT, ashTERT, ashTERT plus asTANKS,
asTANKS, sTANKS and blank control, and the concentration applied
was 2 μM. Oligofectamin concentration depended on oligonucleotide
dose according to the manufacturer’s protocol. Briefly, cells were
plated into 96-well plates and incubated until the cells reached
30–50% confluence. Before transfection, oligonucleotides drug were
diluted with serum-free medium. Then, the desired amount of
oligonucleotide was incubated for 15–20 min with diluted
Oligofectamin. The oligonucleotide/Oligofectamin mixture (20 μl)
was added drop-wise in 80 μl RPMI-1640. After co-incubating for 48
h at 37°C, 50 μl RPMI-1640 containing 5% FBS was added into each
well instead of drugs and cells were analyzed respectively.
RT-PCR assay
The expression of hTERT mRNA was detected by RT-PCR
assay. Total cellular mRNA was isolated with TRIzol Reagent. RNA
sample was quantitated by measurement of optic absorbance (A) at
260/280 nm in a spectrophotometer and the rate of A260/A280 ranged
between 1.8–2.0. Two micrograms of total RNA were reverse
transcribed into cDNA in a volume of 20 μl by Reverse Transcription
kit using M-MuLV reverse transcriptase and oligo (dT) 18 primer
(MDI Fermentas). The mixture was incubated at 42°C for 60 min and
heated to 70°C for 10 min to stop reaction. PCR was performed on a
DNA thermal cycler with primers (Sangon Biotechnology Engineering
Company of Shanghai, China) specific for hTERT, tankyrase, B-cell
CLL/lymphoma 2 (BCL-2), BAX, myeloid cell leukemia-1 (MCL-1) and
β-actin; all the sequences were blasted (http://www.ncbi.nlm.nih.gov/BLAST/). The primers were:
hTERT (263 bp) forward, 5′-TGCGTTTGGTGGATGATTTCTTGT-3′ and reverse,
5′-CCGGGCATAGCTGGAGTAGTCG-3′; tankyrase (763 bp) forward,
5′-GGGCGGAAAGACGTAGTTGA-3′ and reverse, 5′-TTAACTGTGGTGTGGGAGCC-3′;
BCL-2 (708 bp) forward, 5′-CGGAATTCTATGGCGCAAGCCGG GAG-3′ and
reverse, 5′-CGGTACCTCACTTGTGGCCCAG GTATGCACC-3′; BAX (360 bp)
forward, 5′-AAGCTGAGC GAGTGTCTCCGGCG-3′ and reverse, 5′-GCCACAAAGA
TGGTCACTGTCTGCC-3′; MCL-1 (970 bp) forward, 5′-CGG
TAATCGGACTCAACCTCT-3′ and reverse, 5′-ACATTC CTGATGCCACCTCTA-3′;
β-actin (541 bp) forward, 5′-GTG GGGCGCCCCAGGCACCA-3′ and reverse,
5′-CTCCTT AATGTCACGCACGATTTC-3′. Reverse transcription product
(cDNA) (2 μl) was added to the PCR mixture (Takara Co.) for
amplification in a volume of 50 μl. The amplification profile was:
94°C 45 sec, 57–62°C 55 sec, 72°C 60 sec, 32 cycles, finally 72°C
10 min. The products amplified were subjected to 2% agarose gel
electrophoresis and bands were visualized by staining with ethidium
bromide and images were captured under ultraviolet lamp.
Polymerase chain reaction enzyme-linked
immunosorbent assay (PCR-ELISA)
PCR-ELISA assay was measured according to the
manufacturer’s protocol of TRAPeze ELISA Telomerase Detection kit
(Invitrogen) with a minor modification. Cultured A549 cells were
harvested at a density of 1×105/well and washed with
PBS, then homogenized in 200 μl CHAPS lysis buffer and left on ice
for 20 min, and 60 μl of supernatant was collected after
centrifugation (12,000 × g, 20 min, 4°C). PCR was performed in 50
μl supernatant containing 10 μl transfer reaction mixture, 2 μl of
cell extracts was added to 23 μl of nuclease-free water. The PCR
conditions were: the telomerase reaction was carried out at 25°C
for 10 min, followed by a two-step PCR amplification (94°C 40 sec,
50°C 40 sec, 72°C 90 sec, 33 cycles). After equilibrating at 72°C
for 10 min, 5 μl amplified product and 20 μl denatured reagent were
incubated at room temperature. Hybridization buffer (225 μl) was
then added and mixed, and 100 μl of anti-DIG-POD working solution
was added and incubated for another 30 min followed by the addition
of 100 μl TMB substrate solution and 100 μl of stop reagent was
added. The absorbance in each well was read at the wavelength of
450 and 690 nm by microtiter plate reader and the value for each
sample was computed as Asample = A450nm -
A690nm. TSR8 with 8 telomeric repeats was used as
PCR-ELISA positive control (the range of A450nm -
A690nm must be >0.8), and CHAPS lysis buffer without
tissue extract and heat-treated (80°C, 20 min) as negative control
(the range of A450nm - A690nm must be <
0.25). The net increased rate of absorbance for the sample was
recorded as ΔA (%) = (Asample -
Anegative)/(Apositive - Anegative)
× 100%.
Q-FISH assay
Briefly, FACSCalibur System (BD Biosciences) was
calibrated by MESF Quantum 24 Fluorescent Beads and slope
calculated on account of the linear relationship between the MESF
beads and fluorescent channel. According to the manufacturer’s
protocol for the Telomere PNA kit for Flow Cytometry (Dako),
hybridization was performed in 300 μl buffer solution PNA probe at
0.3 μg/ml containing pellet centrifuged from 5×105
washed A549 cells. After incubating at 82°C for 20 min and mixed by
vortexing 2 h in the dark, resuspended cells were collected in 1 ml
wash buffer, mixed and incubated in a 40°C water bath for 10 min.
Centrifuged cells (500 × g, 7 min) were added to 500 μl of DNA
staining buffer, transferred to standard polystyrene flow cytometry
tubes and incubated at room temperature for 2 h. As the same
installation as that used for the MESF beads, samples were detected
and G0/G1 cells were gated from the scatter
plot forward scatter (FSC) vs. DNA dye fluorescence with linear
scaling. FITC fluorescence was displayed on histogram gated on the
population, while mean fluorescence intensity (MFI) was recorded
and buffer without cell sample blank control as well. The mean
telomere length was calculated as: TL (kb) = (MFIsample
- MFIblank) × 0.019 × 0.02604/sec.
Western blotting
A549 cells (5×105) were rinsed with
ice-cold PBS, agitated constantly at 4°C for 30 min followed by
centrifugation with RIPA buffer (12,000 × g, 20 min). The
supernatant was aspirated and the protein concentration levels were
detected by a Bradford assay, which determined whether samples were
fit for test as followed. Through denaturation by anionic detergent
sodium dodecyl sulfate (SDS), 20 μg samples were loaded to 8%
polyacrylamide gel and β-actin as positive control.
SDS-polyacrylamide gel electrophoresis (PAGE) was carried out and
protein blotting was transferred to PVDF membranes, in which
transferred efficiency was checked by ponceau staining soon after.
Membranes were blocked for 2 h at room temperature in 10% blocking
buffer and incubated overnight with first antibody at 4°C, followed
by peroxidase-conjugated secondary lgG. Finally, chemiluminescence
emanating from membranes was revealed and images were captured. The
first antibodies of β-actin, MCL-1, BCL-2 and BAX were purchased
from Santa Cruz and the first antibodies of telomerase and
tankyrase were purchased from Abcam; all the peroxidase-conjugated
second antibodies were purchased from EarthOx.
Apoptosis assay
Apoptosis was detected by flow cytometry using
Annexin V Apoptosis Detection kit FITC (eBioscience). A549 cells
transfected with or without oligonucleotide were harvested 48 h
later. After washing cells once with binding buffer, cells were
stained by Apoptosis Detection kit according to the manufacturer’s
instructions. The samples were tested by FACSCalibur System
(Becton-Dickinson) and the data were analyzed with FlowJo (v7.6.5)
software.
Hoechst 33342 staining
A549 cells were plated in 6-well plates and were
fixed with 4% paraformaldehyde for 10 min at 25°C and cells were
washed twice in PBS. Hoechst 33342 staining (1 mM; Sigma-Aldrich)
solution was added to the plate and incubated for 15 min in the
dark at 37°C; the staining solution was discarded and washed twice
in PBS. Stained cells were observed by fluorescence microscopy
(Olympus).
MTT assay
A549 cells were seeded in 96-well plates at a
density of 2,000 cells/well containing 200 μl RPMI-1640 medium and
transfected with oligonucleotide. Forty-eight hours later, 20 μl of
MTT (5 mg/ml; Sigma-Aldrich) was added to each well and incubated
for 4 h at 37°C avoiding light. The medium was removed and the
formazan was dissolved in 150 μl of DMSO. Absorbance was measured
at 490 nm. Cell inhibition rate was calculated as % IR = (1 -
ODexperiment/ODcontrol) × 100%.
Cell proliferation assay
A549 cells (1×106) were suspended in 1 ml
RPMI-1640 without FBS, 1 μl of 5 mM carboxyfluorescein diacetate
succinimidyl ester (CFSE; Molecular Probes) was added in the tube.
The cells were place at 37°C for 10 min away from light; then, 5
volumes FBS was added and mixed, centrifuged at 300 × g for 5 min
at 20°C and then washed with 10 ml RPMI-1640 complete medium twice.
The labeled cells were the parent generation and the cultured cells
were the daughter generation. Data are described as proliferation
index (PI) analyzed by FlowJo (v7.6.5) software.
Tumor growth experiment in vivo
Female BALB/c nude mice were injected subcutaneously
with 5×106 A549 tumor cells transfected with or without
oligonucleotide to the left flank. The length (L) and width (W) of
tumors were measured regularly, and the volumes of tumor (V) were
calculated by the following formula: V = (L × W2)/2.
Statistical analysis
Data are expressed as mean ± standard error of the
mean and determined by ANOVA test. Differences were considered to
be statistically significant when P<0.05.
Results
The corresponding antisense
oligonucleotide decreases the expression of telomerase reverse
transcriptase (TERT) and tankyrase
TERT, which is the telomerase catalytic subunit, is
a ribonucleoprotein that synthesizes telomeric DNA repeats and
keeps the length of telomere. In the present study, we transfected
the ashTERT and asTANKS into A549 cells respectively or
synchronously, at the same time corresponding shTERT and sTANKS
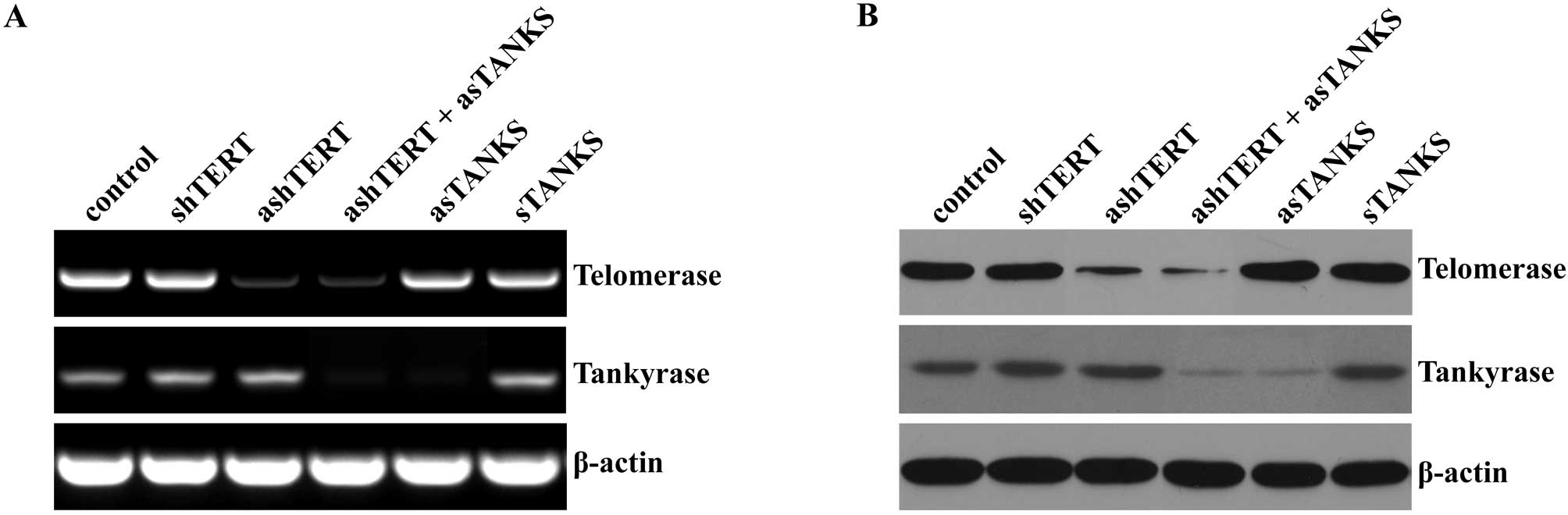
were used as control. Our results showed that the expression of
TERT and tankyrase decreased significantly after transfecting
ashTERT or asTANKS 48 h later in both mRNA and protein level, but
the corresponding sense control oligonucleotide showed no apparent
change (Fig. 1). Of note, neither
ashTERT nor asTANKS affected the expression to each other, while
the combination of ashTERT and asTANKS downregulated TERT and
tankyrase more efficiently than transfecting one of them alone
(Fig. 1).
ashTERT and asTANKS reduce telomerase
activity and shorten telomere length of A549 cells
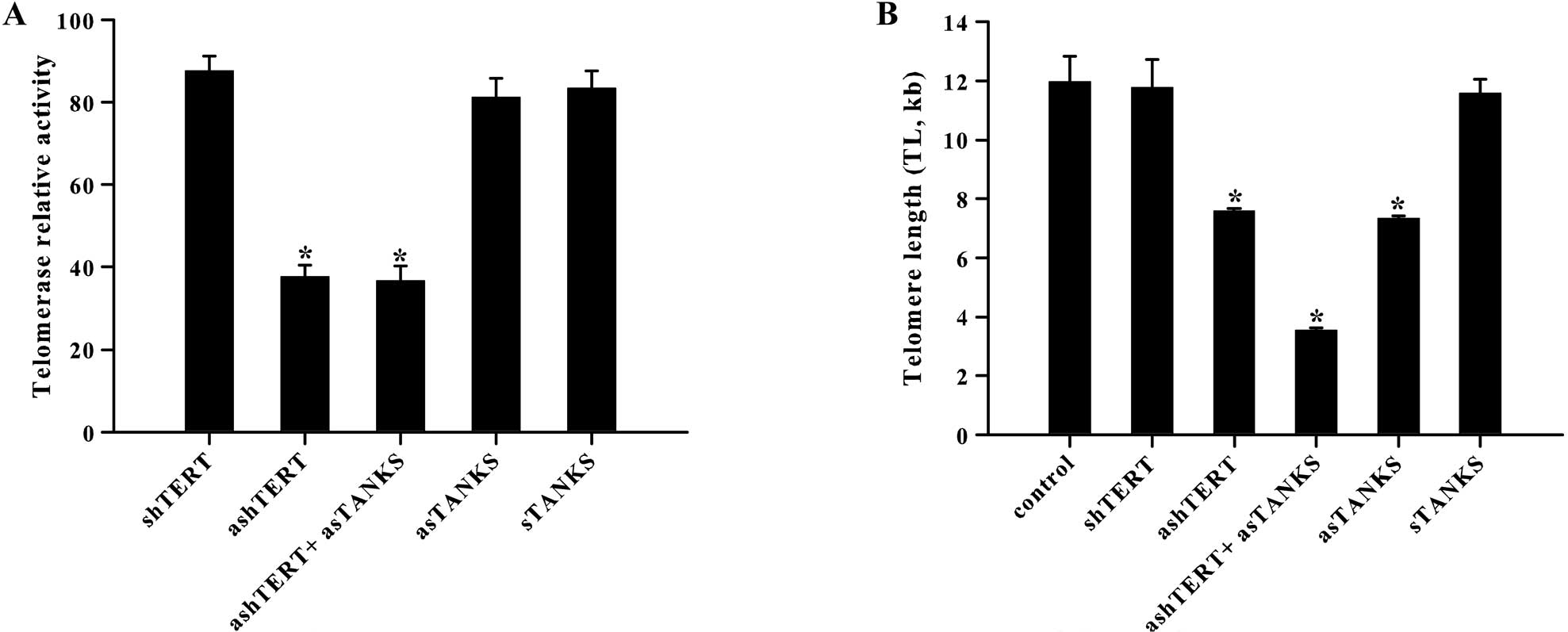
Telomerase activity was decreased markedly by
ashTERT and was uncorrelated with asTANKS (Fig. 2A). The related activity of
telomerase in shTERT, ashTERT, ashTERT plus asTANKS, asTANKS and
sTANKS group was 87.55±3.69, 37.75±2.79, 36.70±3.51, 81.14±4.65 and
83.44±4.13, respectively. There was a significant difference in
telomerase activity between groups with and without ashTERT
(P<0.001). However, regardless of asTANKS or sTANKS, telomerase
activity was independent of them (P>0.05). The data supplied
other evidence that the effect of tankyrase was unrelated to
telomerase. Owing to different handling, there was statistical
significance in mean telomere length, which corresponded to channel
fluorescence intensity (FITC-labeled PNA probe) (Fig. 2B). With either asTANKS or ashTERT
for 48 h, mean telomere length was 7.33±0.09, 7.59±0.07 kb,
respectively, which was markedly lower than controls (P<0.001).
However, regarding asTANKS or ashTERT, the combinated effect of
both sides was more marked in shortening telomere length, in which
mean telomere length was 3.55±0.08 kb (P<0.001).
ashTERT and asTANKS promote A549 cell
apoptosis
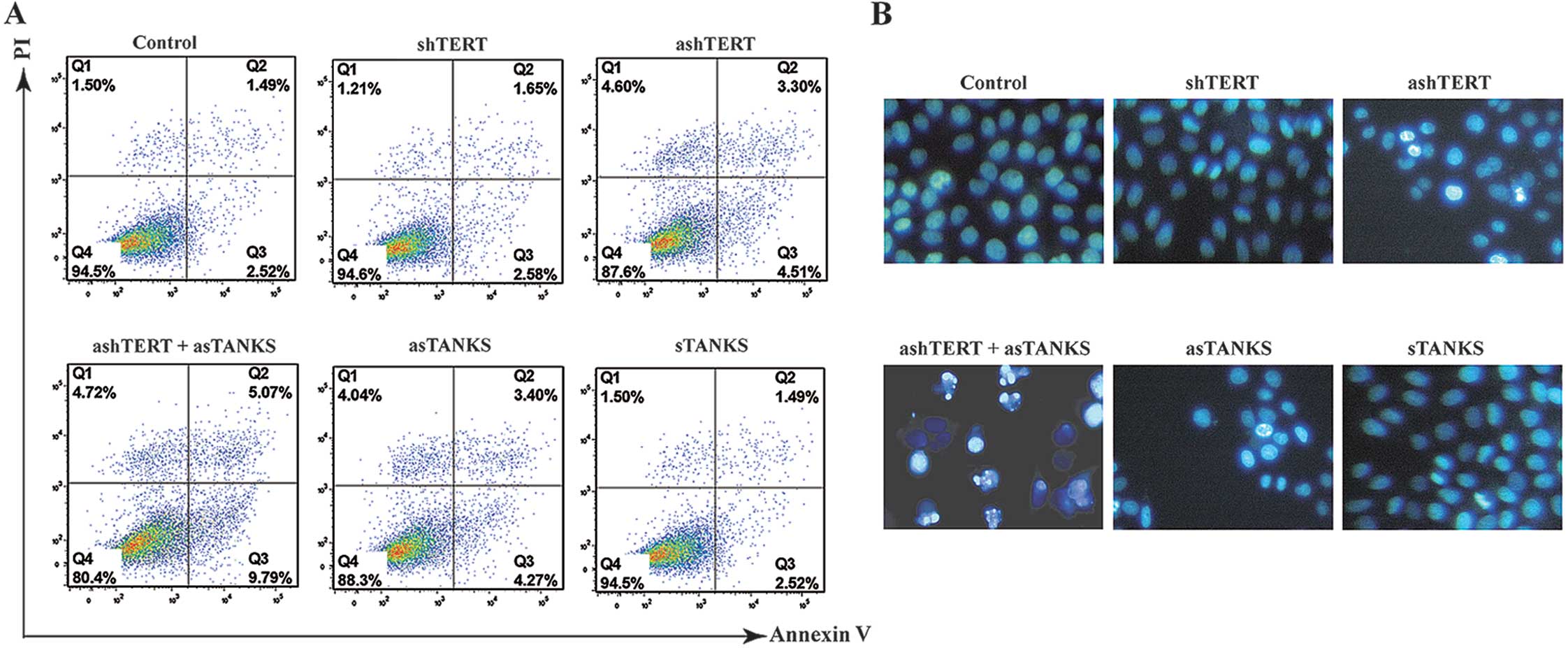
In accordance with our hypothesis, ashTERT and
asTANKS promoted A549 cell apoptosis. Furthermore, the ashTERT plus
asTANKS group had the higher apoptosis percentage of all (Fig. 3A), the apoptosis rate (Q2 + Q3) in
control, shTERT, ashTERT, ashTERT plus asTANKS, asTANKS and sTANKS
group was 4.11±0.25, 4.23±0.21, 7.78±0.35, 14.87±0.42, 7.56±0.34
and 4.15±0.28%, respectively. Furthermore, we stained A549 cells
with Hoechst 33342, a fluorescent dye that accumulated in apoptosis
cells and showed that the dye increased in the ashTERT, asTANKS and
ashTERT plus asTANKS groups, and, in particular, it clearly
accumulated in the ashTERT plus asTANKS group (Fig. 3B).
ashTERT and asTANKS inhibit A549 cell
proliferation
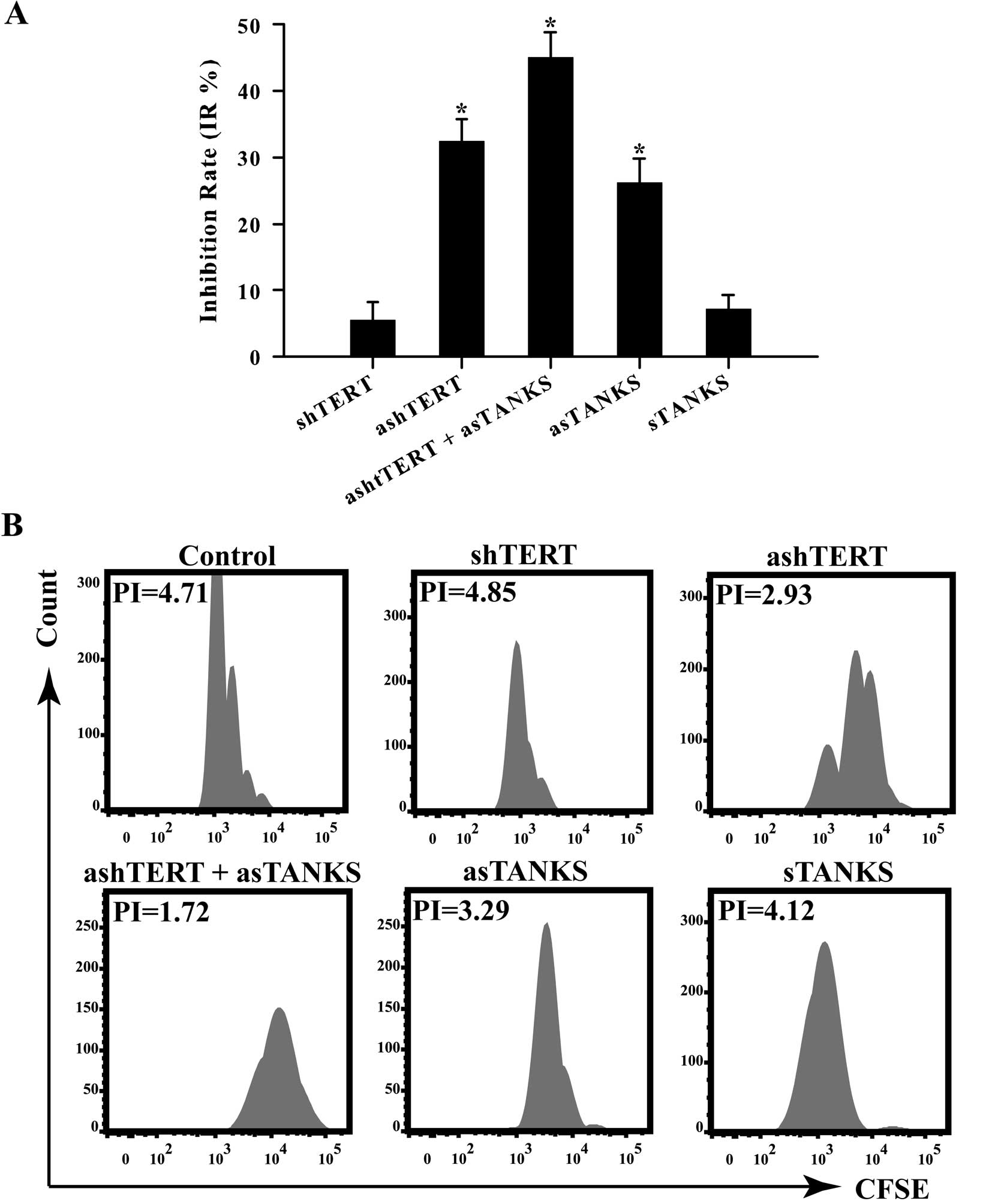
MTT proliferation assays were employed to further
confirm the inhibitory effect on A549 cells caused by ashTERT and
asTANKS. We measured absorbance at 490 nm and calculated inhibition
rate as IR = (1 - A490testing group/A490control
group) × 100%. Forty-eight hours after transfection, the
absorbance at 490 nm was detected and the corresponding IR in
shTERT, ashTERT, ashTERT plus asTANKS, asTANKS and sTANKS group was
5.51±2.76, 32.39±3.31, 45.06±3.73, 26.21±3.54, 7.07±2.23%; our
findings showed that the ashTERT plus asTANKS group inhibited A549
cells proliferation the most efficiently (Fig. 4A). We labeled A549 cells with
5(6)-Carboxyfluorescein diacetate
N-succinimidyl ester (CFSE), a fluorescent dye that can stain live
cells and the fluorescence intensity detected by flow cytometry
weakens followed by mitosis which was used to calculate PI by
software FlowJo (v7.6.5). The PI in control, shTERT, ashTERT,
ashTERT plus asTANKS, asTANKS and sTANKS group was 4.68±0.17,
4.82±0.20, 2.95±0.15, 1.70±0.12, 3.28±0.16, 4.21±0.23 (Fig. 4B). The results were consistent with
the MTT assay that the asTANKS plus sTANKS group had the strongest
ability to inhibit A549 cell proliferation.
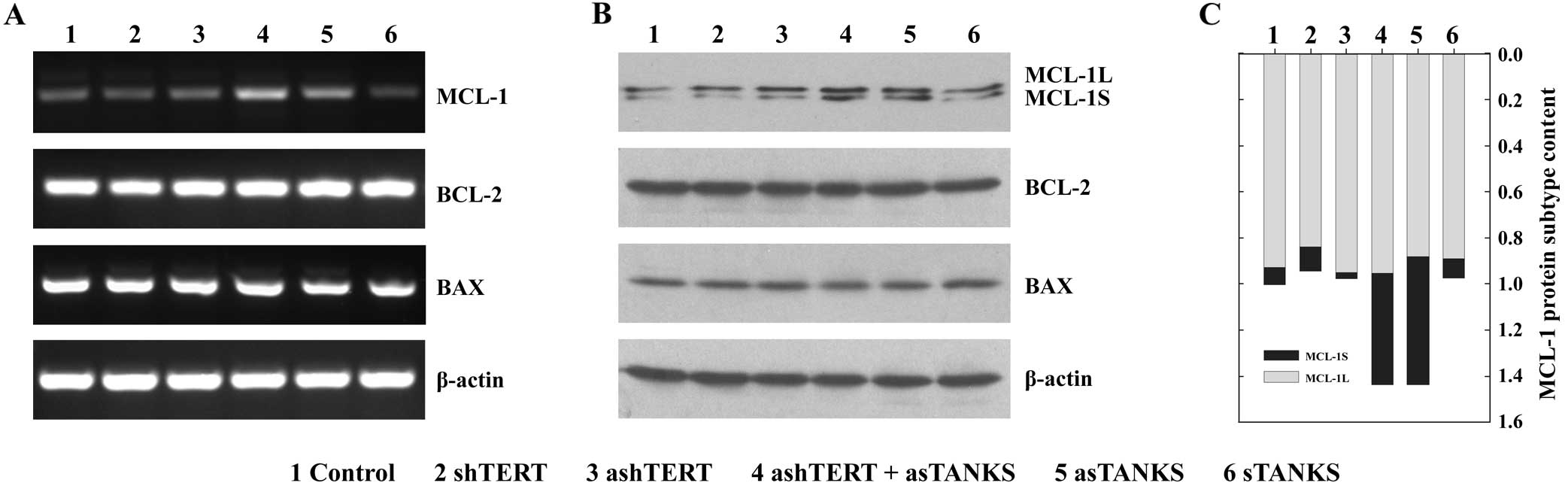
asTANKS increases the expression of MCL-1
in A549 cells
ashTERT and asTANKS enhanced A549 cell apoptosis in
the above study, and, thus, it is likely that some
apoptosis-associated gene may have changed after transfecting by
them. In order to confirm this hypothesis, the anti-apoptotic gene
BCL-2, the pro-apoptotic gene BCL-2-associated X protein (BAX) and
the bcl-2-like protein (MCL-1) were measured by RT-PCR (Fig. 5A) and western blotting (Fig. 5B). The results showed that neither
ashTERT nor shTERT affected the expression of BCL-2, BAX and MCL-1
in mRNA and protein level in A549 cells, but asTANKS upregulated
the MCL-1 mRNA markedly. The combination of asTANKS and ashTERT
also led to a clear MCL-1 overexpression but it did not influence
BCL-2 and BAX expression. Of note, MCL-1Short (MCL-1S), a
pro-apoptotic short isoform, increased more than the alternatively
spliced longer gene product MCL-1Long (MCL-1L) (Fig. 5B and C), which is an anti-apoptotic
isoform. This may explain the molecular mechanism for asTANKS
accelerating A549 cells apoptosis.
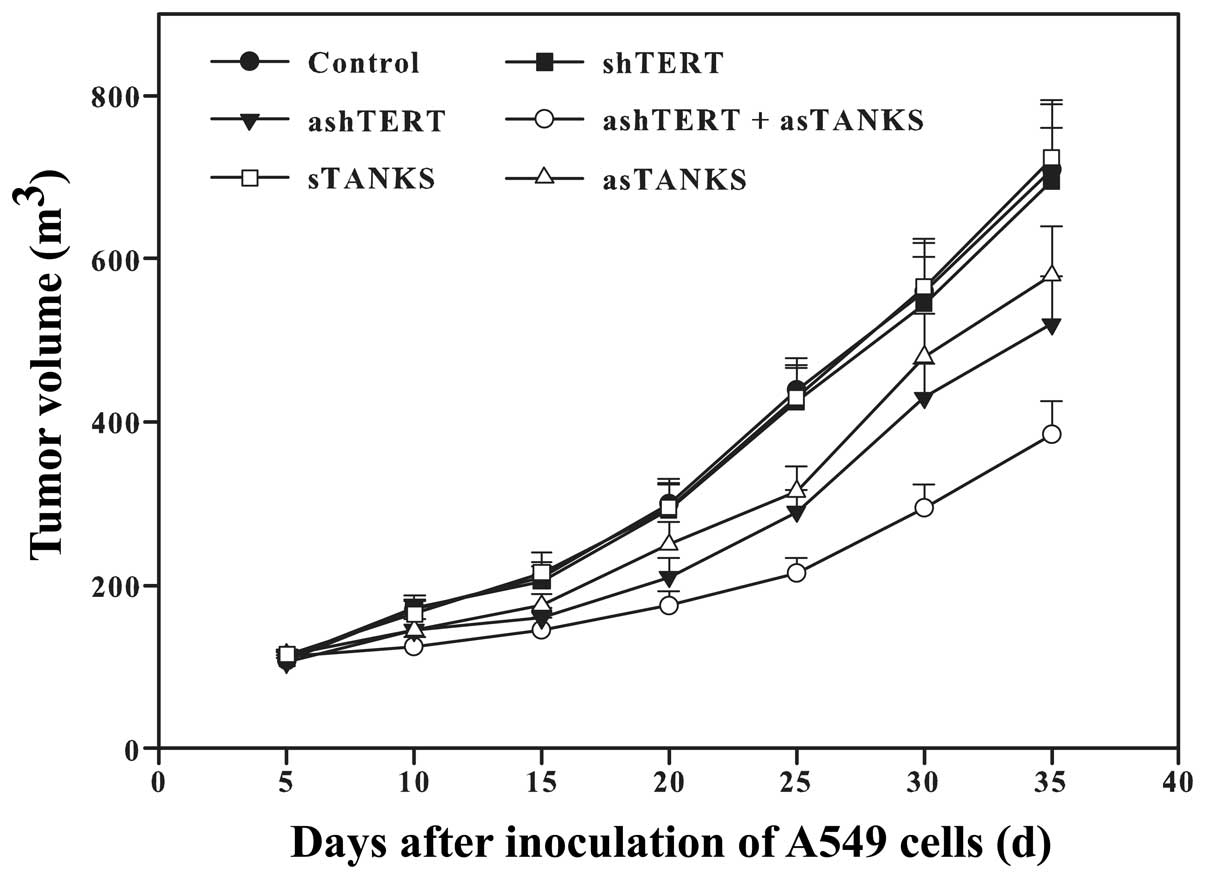
ashTERT and asTANKS inhibit A549 growth
in vivo
The above results revealed that ashTERT and asTANKS
promoted A549 cell apoptosis and inhibited their proliferation
in vitro. In order to observe their effect in vivo,
we inoculated A549 cells subcutaneously in the nude mice after
transfection for 48 h. It showed that both ashTERT and asTANKS
inhibited A549 cell proliferation and the tumor volume in the
ashTERT plus asTANKS group was the least of all (Fig. 6).
Discussion
Telomerase and tankyrase play an important role in
maintaining cell telomere length and cell division capacity. They
are always at high levels in several types of malignant cancer,
such as lung cancer (8), gastric
cancer (9) and prostate cancer
(10). Additionally, the higher
their expression in cancer the poorer the prognosis in patients,
therefore telomerase and tankyrase are gradually becoming targets
of anticancer drugs.
Several telomerase and tankyrase inhibitors have
been developed to treat cancer and present favorable antitumor
effects, including azidothymidine (AZT), a telomerase inhibitor
that can suppress reverse transcriptase activity of telomerase,
promote apoptosis of human liver cancer cells HepG2 and SMMC-7721
and inhibit their proliferation (11). The tankyrase inhibitor JW55 which
suppresses poly(ADP-ribose) polymerase (PARP) domain of tankyrase
also suppressed SW480 human colon cancer cell proliferation by
blocking the Wnt signaling pathway (12).
In the present study, ashTERT and asTANKS were used
as the special inhibitors to silence the expression of target genes
in human lung adenocarcinoma cells by transfection. At the same
time, the corresponding sense oligonucleotides were used as
control. Similar to the previously published reports, both ashTERT
and asTANKS led A549 cells to apoptosis and suppressed
proliferation. Moreover, combined ashTERT and asTANKS enhanced
these effects; thus, the results indicated that asTANKS could
improve the antitumor effect of ashTERT. In this report, it was
also observed that ashTERT and asTANKS shortened the telomere
length and, similarly, the combined group presented the more
evident effect. Only ashTERT suppressed the telomerase reverse
transcriptase activity, but asTANKS had almost no effect on it.
Furthermore, asTANKS did not yet enhance this effect of ashTERT.
Consistent with a previous study, the antisense oligonucleotide of
tankyrase also shortened the length of telomere in SGC-7901 human
gastric cancer cells and did not affect telomerase activity
(13). Thus, it should be noted
that asTANKS did not affect the telomerase activity directly, but
it reduced the length of telomere in A549 cells by inhibiting the
expression of tankyrase, leading to the telomere structure becoming
tight and inhibiting telomerase approaching the telomere to
elongate.
The telomere shortening can cause apoptosis of tumor
cells (14). In the present study,
we detected the cardinal anti-apoptotic gene BCL-2 and the
pro-apoptotic gene BAX, but neither ashTERT nor asTANKS altered the
expression of these genes. MCL-1, which can interact with tankyrase
directly and regulate apoptosis, has two alternative splicing
transcript variants which have distinct functions (15). MCL-1L is a long transcript variant
that has been shown to form heterodimers with pro-apoptotic
proteins BIM and BAK, and which suppresses the release of
cytochrome c to inhibit apoptosis (16). MCL-1S combines with MCL-1L directly
to promote apoptosis (17), thus
the rate of these two isoforms determines the cell apoptosis. In
this study, asTANKS upregulated the expression of MCL-1L and
MCL-1S, and MCL-1S increased more significantly, ashTERT
strengthened asTANKS effect but ashTERT alone had no such effect.
Similarly, in vivo ashTERT and asTANKS suppressed the growth
of A549 in nude mice and the A549 tumor grew the slowest in the
combined group.
Telomere maintenance by telomerase is the key point
of infinite growth for most cancer cells. Continuous treatment of
cancer cells with telomerase inhibitors shortens telomeres and
eventually induces cellular apoptosis. Moreover, there were 19
phase I, II or III clinical trials regarding telomerase inhibitor
running in 2012 worldwide, and it may become the target of cancer
therapy (18).
A potential disadvantage is that telomere shortening
depends on the repetitive occurrence of the DNA end replication
problem resulting from cell division (19). For this reason, it is essential that
telomerase inhibitors are not cytotoxic. Furthermore, as telomere
loss is a gradual process there is a lag between the time
telomerase is inhibited and the time telomeres shorten sufficiently
to disrupt the capping function. This would necessitate long
treatment schedules that may lead to acquired drug resistance both
in the cell and throughout the body. In general, longer telomeres
provide more binding sites for TRF1, which blocks telomere access
to telomerase. Accordingly, telomere shortening compromises the
effect of telomerase inhibitors since shorter telomeres have fewer
TRF1 molecules and therefore allow easier access to residual
telomerase activity. Thus, the rate of telomere shortening per cell
division decreases with telomere shortening itself. This phenomenon
results from the incomplete shutdown of telomerase activity by
telomerase inhibitors. A better therapeutic outcome may result from
increasing the efficiency of telomere shortening to hasten the
telomere crisis.
Telomere accessibility is also a potential target
for telomerase inhibition. Inhibition of tankyrases, that enhance
telomerase access to telomeres, may indirectly induce cancer cell
senescence by abrogating telomerase activity (20). Tankyrase inhibitors enhance the rate
of telomere shortening by means of a telomerase inhibitor, and
induce earlier cell crisis. Tankyrase inhibitors do not directly
inhibit telomerase activity but lead to telomere shortening, to a
small extent, presumably by reducing telomere access to telomerase.
Thus, it is expected that the effect of such tankyrase inhibitors
on telomere length is selective to telomerase-positive cells and
these provide support for tankyrase 1 as a suitable target for
telomere-directed cancer therapy.
These observations suggest that the pharmacological
targeting of tankyrase oligos is a potentially significant
anticancer strategy if used in conjunction with telomerase
inhibitors. This trend would further promote development not only
of telomerase but also of tankyrase inhibitors.
Acknowledgements
The present study was supported by Grant 81101550
from the National Natural Science Foundation of China and the
Natural Science Foundation of Hubei Province, China (no.
2012FFB05904).
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar
|
|
2
|
Gadgeel SM, Ramalingam SS and Kalemkerian
GP: Treatment of lung cancer. Radiol Clin North Am. 50:961–974.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tümpel S and Rudolph KL: The role of
telomere shortening in somatic stem cells and tissue aging: lessons
from telomerase model systems. Ann NY Acad Sci. 1266:28–39.
2012.PubMed/NCBI
|
|
4
|
Gomez DE, Armando RG, Farina HG, Menna PL,
Cerrudo CS, Ghiringhelli PD and Alonso DF: Telomere structure and
telomerase in health and disease (Review). Int J Oncol.
41:1561–1569. 2012.PubMed/NCBI
|
|
5
|
Shay JW and Wright WE: Role of telomeres
and telomerase in cancer. Semin Cancer Biol. 21:349–353. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ruden M and Puri N: Novel anticancer
therapeutics targeting telomerase. Cancer Treat Rev. 39:444–456.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Riffell JL, Lord CJ and Ashworth A:
Tankyrase-targeted therapeutics: expanding opportunities in the
PARP family. Nat Rev Drug Discov. 11:923–936. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cha N, Li XY, Zhao YJ, Wang EH and Wu GP:
hTERT gene amplification and clinical significance in pleural
effusions of patients with lung cancer. Clin Lung Cancer.
13:494–499. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gigek CO, Leal MF, Silva PN, Lisboa LC,
Lima EM, Calcagno DQ, Assumpção PP, Burbano RR and de Smith MA:
hTERT methylation and expression in gastric cancer. Biomarkers.
14:630–636. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bantis A, Patsouris E, Gonidi M, Kavantzas
N, Tsipis A, Athanassiadou AM, Aggelonidou E and Athanassiadou P:
Telomerase RNA expression and DNA ploidy as prognostic markers of
prostate carcinomas. Tumori. 95:744–752. 2009.PubMed/NCBI
|
|
11
|
Chen C, Zhang Y, Wang Y, Huang D, Xi Y and
Qi Y: Synergic effect of 3′-azido-3′-deoxythymidine and arsenic
trioxide in suppressing hepatoma cells. Anticancer Drugs.
22:435–443. 2011.
|
|
12
|
Waaler J, Machon O, Tumova L, Dinh H,
Korinek V, Wilson SR, Paulsen JE, Pedersen NM, Eide TJ, Machonova
O, Gradl D, Voronkov A, von Kries JP and Krauss S: A novel
tankyrase inhibitor decreases canonical Wnt signaling in colon
carcinoma cells and reduces tumor growth in conditional APC mutant
mice. Cancer Res. 72:2822–2832. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang H, Yang MH, Zhao JJ, Chen L, Yu ST,
Tang XD, Fang DC and Yang SM: Inhibition of tankyrase 1 in human
gastric cancer cells enhances telomere shortening by telomerase
inhibitors. Oncol Rep. 24:1059–1065. 2010.PubMed/NCBI
|
|
14
|
Zhang X, Mar V, Zhou W, Harrington L and
Robinson MO: Telomere shortening and apoptosis in
telomerase-inhibited human tumor cells. Genes Dev. 13:2388–2399.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bae J, Donigian JR and Hsueh AJ: Tankyrase
1 interacts with Mcl-1 proteins and inhibits their regulation of
apoptosis. J Biol Chem. 278:5195–5204. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Adams KW and Cooper GM: Rapid turnover of
mcl-1 couples translation to cell survival and apoptosis. J Biol
Chem. 282:6192–6200. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bae J, Leo CP, Hsu SY and Hsueh AJ:
MCL-1S, a splicing variant of the antiapoptotic BCL-2 family member
MCL-1, encodes a proapoptotic protein possessing only the BH3
domain. J Biol Chem. 275:25255–25261. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Buseman CM, Wright WE and Shay JW: Is
telomerase a viable target in cancer? Mutat Res. 730:90–97. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tian X, Chen B and Liu X: Telomere and
telomerase as targets for cancer therapy. Appl Biochem Biotechnol.
160:1460–1472. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Seimiya H: The telomeric PARP, tankyrases,
as targets for cancer therapy. Br J Cancer. 94:341–345. 2006.
View Article : Google Scholar : PubMed/NCBI
|