Introduction
Radiotherapy remains the standard radiation modality
used for the treatment of breast cancer, which is a common
malignancy worldwide and carries a high mortality rate (1). Previous studies have shown that
radiation improves overall survival from breast cancer in women
with early stage and advanced disease (2,3).
Despite the fact that improvements in the management of breast
cancer have been made, the clinical application of radiotherapy has
improved only marginally, due to the tumor resistance to ionizing
radiation (IR). The emerging reasonable explanation for this
phenomenon is the existence of a rare subpopulation of cells which
are purported cancer stem cells (CSCs) or cancer-initiating cells
(CICs) that may be contribute to some cases of resistance to cancer
therapy (4,5). Pre-clinical data suggest that breast
CSCs/CICs can be enriched after radiation and that breast cancer
stem/initiating clonogens are particularly resistant to radiation
(6,7). However, the molecular mechanisms that
mediate radiation resistance of breast CSCs remain unidentified.
Therefore, uncovering key genes which are responsible for
maintaining the radiation resistance of breast CSCs is a critical
approach for improving the effects of radiotherapy.
Glucose-regulated protein 78KD (GRP78), one of the
best-characterized endoplasmic reticulum (ER) chaperones, serves
multiple functions in maintaining cellular homeostasis. GRP78 has
been implicated as a mediator of tumor proliferation and
metastasis, therapeutic resistance and recurrence (8,9).
Moreover, recent data indicate that GRP78 plays a crucial role in
stem cell biology (10). For
instance, GRP78 is required for the survival of embryonic stem cell
precursors and is also highly expressed in hematopoietic stem cells
(11). Additionally, GRP78 has been
reported to be highly elevated in breast disseminated tumor cells,
which shared similar biological properties of CICs (12). In agreement, differential systemic
analysis revealed elevated GRP78 expression in head and neck CICs
(13). However, the role of GRP78
in breast CICs has yet to be determined. Based on these findings,
it is worth investigating the role of GRP78 in breast CICs if GRP78
is preferentially overexpressed in CICs.
In order to test this hypothesis, we first
identified breast CICs from MCF-7 cell lines by utilizing flow
cytometry based cell-sorting base on ABCG2 efflux pump-mediated
Hoechst 33342 dye exclusion, which enables the isolation of a rare
stem-like side population (SP) cells (14–17).
This method allows estimating the GRP78 expression and determining
whether isolated SP fraction which harbors the cancer stem
cell-like properties is involved in resistance to radiation. This,
in turn, could provide an explanation as to why tumors have some
degree of intrinsic resistance to radiotherapy.
Materials and methods
Cell culture and reagents
The MCF-7 breast cancer cell line was purchased from
the American Type Culture Collection (Manassas, VA, USA) and
cultured in Roswell Park Memorial Institute (RPMI)-1640 culture
medium (Macgene, Beijing, China) supplemented with 10% (v/v) fetal
bovine serum (FBS; Hyclone, Utah, USA) at 37°C in a humidified
atmosphere containing 5% CO2. Hoechst 33342, epidermal
growth factor (EGF), human recombinant basic fibroblast growth
factor-basic (bFGF), B27 supplement and Lipofectamine®
2000 were purchased from Invitrogen (Carlsbad, CA, USA). Verapamil
was obtained from Sigma (St. Louis, MO, USA) and dissolved in
distilled deionized water (ddH2O). Serum-free
Opti-MEM® I and DMEM/F12 medium were purchased from
Gibco (Grand Island, NY, USA).
Analysis and isolation of SP and non-SP
cell fractions from MCF-7
The SP analysis was performed based on the method
reported by Goodell et al(14) with slight modifications. MCF-7 cells
were digested with 0.25% trypsin (Sigma), washed twice with PBS,
resuspended in pre-warmed RPMI-1640 culture (supplemented with 2%
FBS) at a density of 1×106 cells/ml. Then, the cells
were pre-incubated with or without 50 μM verapamil for 30 min at
37°C before adding the Hoechst 33342 dye at a final concentration
of 5 μg/ml. The mixture was incubated in the dark at 37°C for 90
min with interval mixing. Following incubation, the cells were
washed twice with ice-cold PBS, and the cells were then filtered
through a 40 μm nylon mesh to obtain single cell suspension and
kept at 4°C in the dark. Cell analysis and purification were
performed using FACS (FACSAria II; Becton-Dickinson, CA, USA).
Hoechst 33342 was excited with UV light at 355 nm and fluorescence
emission was measured with 450/BP50 (Hoechst blue) and a 660/BP50
(Hoechst red) optical filters. At the end of sorting, both
collected SP and non-SP cells were reanalyzed to evaluate sorting
purity and to conduct further experiments. To minimize the
non-specific effects of the Hoechst dye on the sorted cells, we
cultured both SP and non-SP cells for 24 h to remove dead cells and
then performed all experiments described below.
Long-term differentiation of SP and
non-SP cells
Sorted SP and non-SP cells were cultured in
RPMI-1640 (supplemented with 10% FBS) for 28 days. Then the
cultured SP and non-SP cells were stained with Hoechst 33342 and
analyzed by FACS to determine the differentiation ability of the
two subpopulations.
Sphere formation and clone formation
assay
Sorted SP and non-SP cells were cultured in tumor
sphere medium consisting of serum-free DMEM/F12 medium, B27
supplement, 20 ng/ml EGF and 10 ng/ml bFGF. Cells were plated at a
density of 5×103 cells/well in ultra-low-attachment
6-well plate triplicates and the medium was changed every other
day. After 10 days in culture, colonies that contained >20 cells
were counted.
For clone formation assay, sorted SP and non-SP
cells were plated in triplicate at 600 cells on each
25-cm2 flasks and cultured with RPMI-1640 (supplemented
with 10% FBS) for 10 days. Then, cells were fixed and stained with
0.5% crystal violet. Colonies containing >50 cells were manually
counted. The clone formation efficiency was the ratio of the clone
number to the planted cell number.
Tumorigenicity assay
Numbers of sorted SP and non-SP cells
(1×105) suspended in 200 μl PBS were injected
subcutaneously in the flank region of 5-week-old female BALB/c nude
mice obtained from the Institute of Laboratory Animal Science of
Peking University Health Science Center. The mice were monitored
weekly and euthanized 4 weeks after transplantation to assess tumor
formation. Tumors were measured using a vernier caliper, weighed
and photographed. Tumor volume (TV) was calculated using the
following formula: TV (mm3) = (length ×
width2)/2. A portion of the subcutaneous tumor tissue
was collected, fixed in 10% formaldehyde and embedded in paraffin
for hematoxylin and eosin (H&E) staining to assess tumor
pathology. All animal practices were in accordance with the
guidelines of Peking University Health Science Center for the use
of laboratory animals.
Radiation and clonogenic assay
Subsequently, 600–8,000 cells were plated on
25-cm2 flasks in triplicate for each experiment. Twelve
hours later, the cells irradiated at room temperature with a
60Co laboratory irradiator (Beijing Normal University,
Beijing) at a dose rate of 1 Gy/min for the time required to
generate a dose curve of 0, 0.5, 1, 2, 4, 6 and 8 Gy. The control
was sham irradiated. Following the irradiation, the cells were
incubated for an additional 9 days, and the cells were fixed with
100% carbinol and stained with 0.5% crystal violet. Only colonies
containing >50 cells were manually counted. The surviving
fraction was calculated as follows: plating efficiency (PE) =
(colony number/inoculating cell number) × 100%. SF = PE (tested
group)/PE (0 Gy group) × 100%. The cell-survival was calculated
according to the single-hit multi-target formula: SF = 1 − (1 −
e−D/D0)N(18). The radiobiological parameters of
cellular radiosensitivity (D0, mean lethal dose), the
capacity for sublethal damage repair (Dq,
quasi-threshold dose) and the extrapolation number (N) were
calculated. Then, those values were used to calculate the SF after
irradiation at a dose of 2 Gy (SF2) and the sensitization
enhancement ratio (SER).
RNA extraction and quantitative real-time
PCR
Total RNA was extracted from newly sorted SP and
non-SP cells using Takara RNAiso Plus (Dalian, China) and reverse
transcriptions were performed according to the protocol supplied by
the manufacturer (Takara, Japan).
Real-time PCR was performed using SYBR-Green I
master mix kit (Takara) on a Bio-Rad IQ5 Real-time-PCR Reaction
System (Bio-Rad Laboratories, Inc., CA, USA). The relative amounts
of mRNA were calculated from the values of comparative threshold
cycle by using GAPDH as control (primers are depicted in Table I). The reaction was carried out with
the following cycling conditions: 95°C for 2 min followed by 45
cycles of amplification (denaturation at 95°C for 15 sec, annealing
at 58°C for 20 sec and extension at 72°C for 30 sec).
 | Table IPrimer sequences used for the
real-time PCR. |
Table I
Primer sequences used for the
real-time PCR.
| Gene | Primer | Product size
(bp) | Temperature
(°C) |
|---|
| ABCG2 | S:
5′-CATGTACTGGCGAAGAATATTTGGT-3′ | 74 | 65.2 |
| (NM_004827) | A:
5′-CACGTGATTCTTCCACAAGCC-3′ | | 64.3 |
| Bmi1 | S:
5′-AAATGCTGGAGAACTGGAAAG-3′ | 124 | 60.9 |
| (NM_ 005180) | A:
5′-CTGTGGATGAGGAGACTGC-3′ | | 61.1 |
| Nanog | S:
5′-ATTCAGGACAGCCCTGATTCTTC-3′ | 76 | 65.5 |
| (NM_024865) | A:
5′-TTTTTGCGACACTCTTCTCTGC-3′ | | 64.5 |
| Sox2 | S:
5′-CGAGTGGAAACTTTTGTCGGA-3′ | 74 | 63.3 |
| (NM_003106) | A:
5′-TGTGCAGCGCTCGCAG-3′ | | 63.4 |
| Oct4 | S:
5′-GTGGAGAGCAACTCCGATG-3′ | 86 | 61.8 |
| (NM_002701) | A:
5′-TGCTCCAGCTTCTCCTTCTC-3′ | | 63.4 |
| GRP78 | S:
5′-CACGCCGTCCTATGTCGC-3′ | 238 | 60 |
| (NM_005347) | A:
5′-AAATGTCTTTGTTTGCCCACC-3′ | | 55.9 |
| GAPDH | S:
5′-AATTGAGCCCGCAGCCTCCC-3′ | 153 | 69.7 |
| (NM_002046) | A:
5′-CCAGGCGCCCAATACGACCA-3′ | | 69.3 |
Transient overexpression and silencing of
GRP78 in SP cells
To overexpress and silence the GRP78 in SP cells,
the plasmid (pcDNA3.1/hGRP78, a gift from Dr RC Austin, McMaster
University, Ontario, Canada; pSuper/GRP78 RNAi, designed by our
laboratory) which can overexpress and silence the GRP78 in
mammalian cells was introduced by transfection. For transient
transfection, SP cells were cultured in 6-well plates and
transfected at 80% confluence with Lipofectamine® 2000
according to the manufacturer’s instructions. After transfection,
the cells were left for another 36 h before they were harvested by
trypsinization and resuspended for clonogenic experiments. The
siRNA sequences for human GRP78 are: sense
5′-GATCCCCGATCACAATCACCAATGACTTCAAGAGAGTCATTGGTGATTGTGATCTTTTTA-3′
and antisense
5′-AGCTTAAAAAGATCACAATCACCAATGACTCTCTTGAAGTCATTGGTGATTGTGATCGTG-3′.
Statistical analysis
Statistical analysis was performed with Sigmaplot
10.0 software. Statistical differences between SP and non-SP cells
were analyzed using the Student’s t-test. Data are presented as the
means ± SEM. P-values <0.05 were considered to indicate
statistically significant differences.
Results
Isolation of SP and non-SP cell fractions
from MCF-7 cell line
We first attempted to isolate a side-population
within MCF-7 breast cancer cell lines. Following trypsinization and
staining with fluorescent dye Hoechst 33342, the cells were
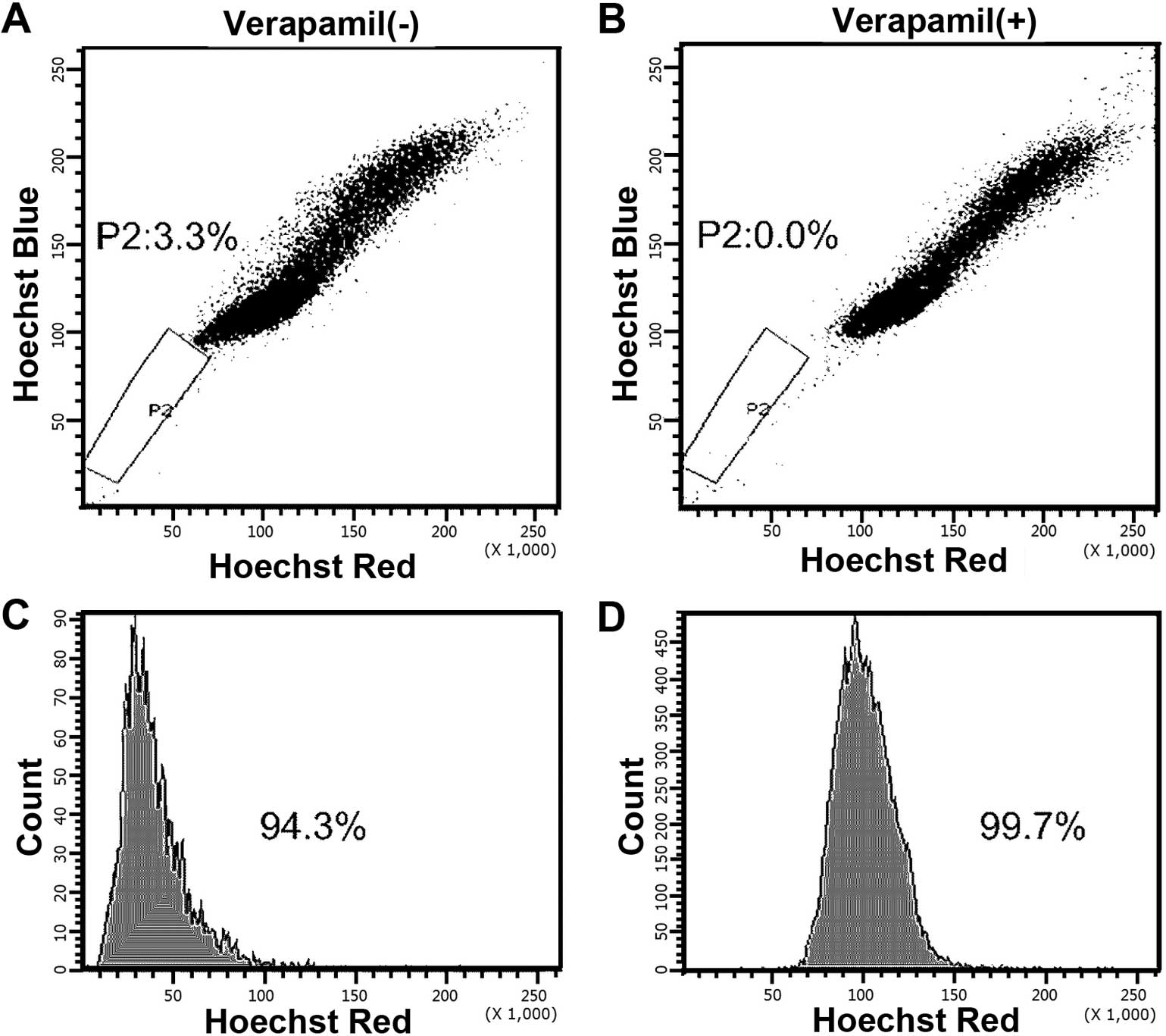
analyzed by flow cytometry; the P2 gate showed that the SP cells
were Hoechst 33342 negative/low and the P3 gate indicated the
non-SP cells that were Hoechst 33342 positive (Fig. 1A). MCF-7 cells presented a distinct
SP, accounting for 3.3% of the whole population. The percentage of
SP cells diminished to ~0.0% of the total cells when pretreated
with verapamil (Fig. 1B),
confirming that SP cells extrude Hoechst 33342 dye actively via a
verapamil-sensitive ABC transporter. Then the SP (P2) and non-SP
(P3) cells were sorted separately and applied for further
experiments; the purity of SP and non-SP cells was 94.3 and 99.7%,
respectively (Fig. 1C and D).
SP cells show high tumorigenicity and
clonogenic capacity
High tumorigenic potential is considered a hallmark
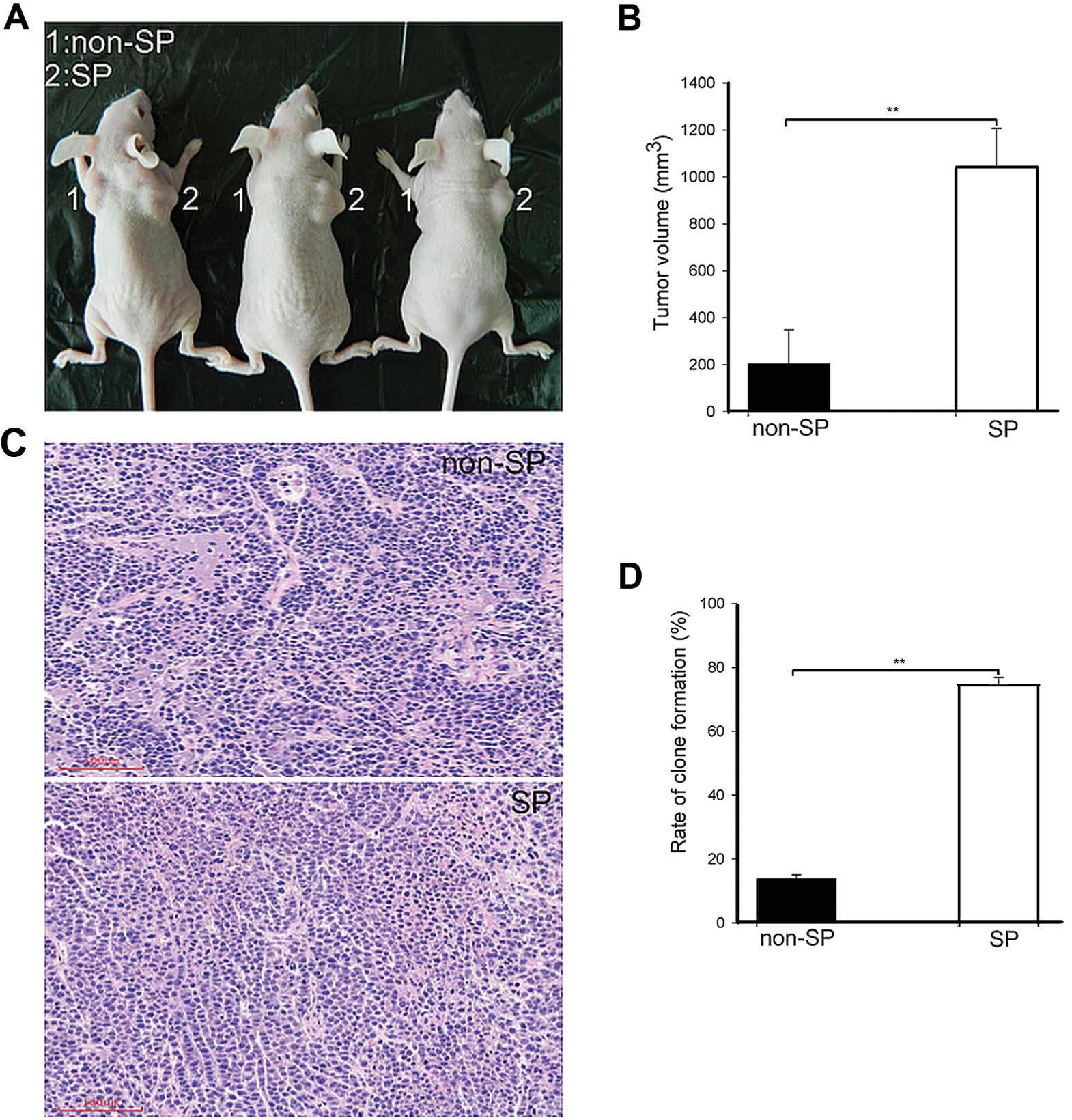
of CICs (19). We first performed
tumorigenicity assay to compare the tumorigenic potential of SP and
non-SP cells, respectively. Both 1×105 numbers of SP and
non-SP cells were injected into the flank region of nude mice
subcutaneously (n=3). Four weeks after inoculation, both SP and
non-SP cells were able to produce tumors as shown in Fig. 2A. Then, the mice were euthanized and
the tumors were measured with a vernier caliper. The results showed
that SP cells formed a tumor with a mean volume of 1043.12±163.65
mm3 while non-SP cells formed tumor with 204.04±144.86
mm3 mean volume (Fig.
2B). H&E staining results confirmed that the tumors formed
by SP and non-SP cells were typical adenomatous carcinoma (Fig. 2C). H&E staining of tumors grown
in mice after injection of SP cells showed the presence of
malignant cells, with large nuclei and prominent nucleoli; some
cells showed a dark basophilic cytoplasm. Moreover, cells were in a
chaotic arrangement with necrosis (Fig.
2C).
Additionally, the clone formation assay showed that
the mean clone formation efficiency was 77.56±3.67 and 26.39±3.25%
in SP and non-SP cells, respectively (Fig. 2D). In vitro clonogenic
potential indicated the efficiency of SP and non-SP cells to form a
tumor, which is consistent with the in vivo tumor transplant
results.
SP cells harbor long-differentiation
ability
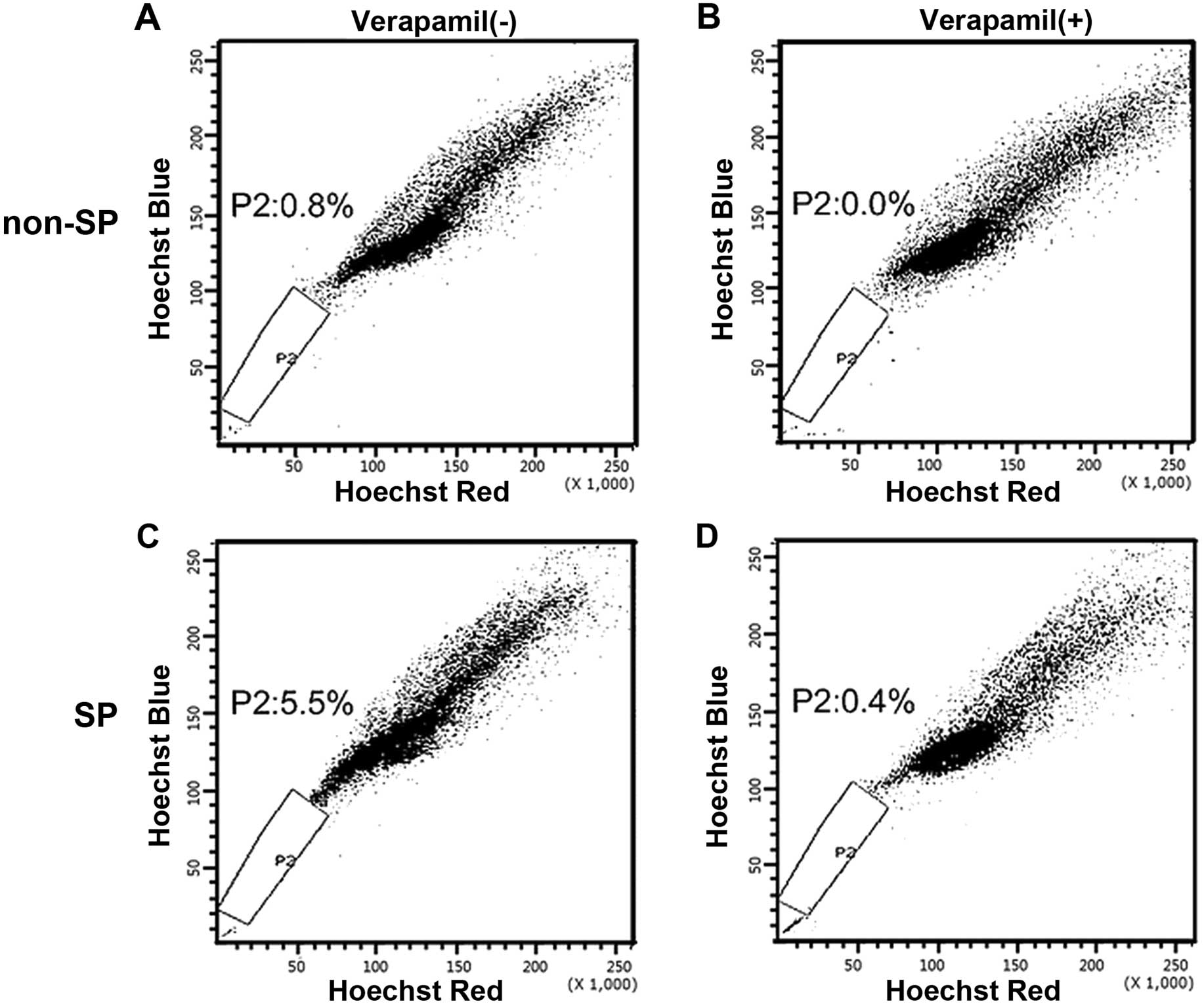
SP cells and non-SP cells were cultured for 4 weeks
in normal RPMI-1640 medium, stained again with Hoechst 33342 and
analyzed using the cell sorter. As shown in Fig. 3, SP and non-SP fractions were
obtained again from the former SP subpopulation after being
cultured for 4 weeks (Fig. 3C). By
contrast, SP fraction could not be obtained from the former non-SP
population (Fig. 3D). This
indicated that SP cells may undergo asymmetrical division to
generate heterogeneous phenotypes of low-tumorigenic cells, such as
non-SP cells that form the bulk of the tumor.
SP cells display higher stemness gene
expression, self-renewal ability and resistance to IR
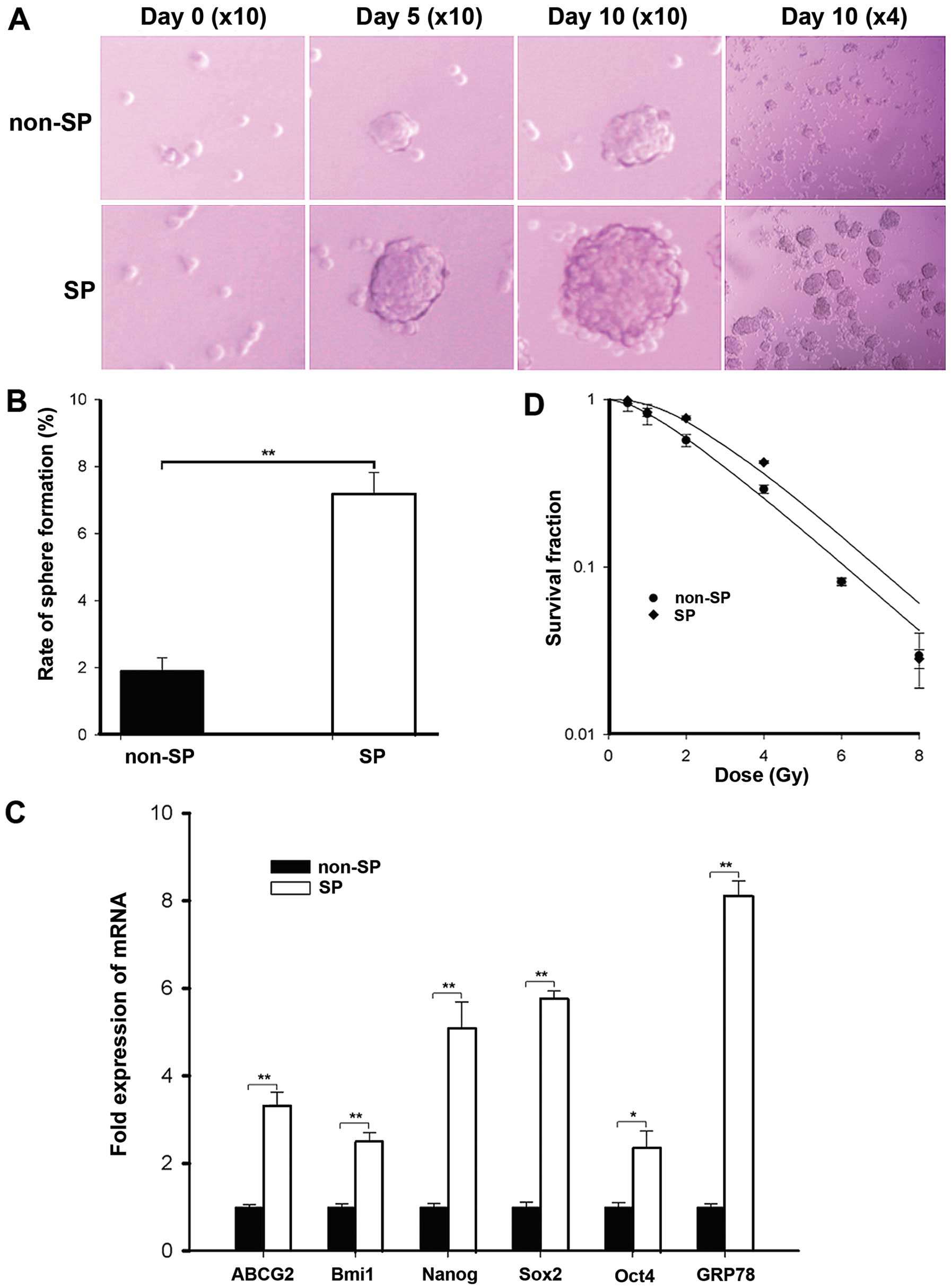
Sphere formation has been well described as a
typical characteristic of CICs that reflects the potential for
self-renewal (20). We evaluated
the ability of SP and non-SP cells to generate spherical colonies
in an ultra-low attachment and serum-starved culture system. After
10 days of culture, spherical colonies were counted. As shown in
Fig. 4A, SP cells displayed much
higher tumor sphere-forming ability than non-SP cells; the sphere
formation efficiency was 10.17±2.33% for the SP cells vs.
2.33±0.25% for the non-SP cells (Fig.
4B).
To further confirm the stem phenotype of the SP
cells, the expression of embryonic stem cell (ES) marker genes were
also investigated, as CICs are considered to share similar
characteristics with normal stem cells (21). As shown in Fig. 4C, the q-PCR analysis indicated that
the mRNA expression of stemness genes such as Oct4, Nanog, Sox2 and
Bmi1 in SP cells was significantly higher than non-SP cells.
Furthermore, the results also showed that the mRNA levels of ABCG2
in SP cells were more highly expressed as compared to non-SP cells,
according to the sorting phenotype of Hoechst 33342 exclusion.
Current data demonstrate that breast CICs can be
enriched after radiation and CSCs/CICs are particularly more
resistant to radiotherapy (6,7,22). To
compare the SP and non-SP cell response to radiation-induced
cytotoxicity, clonogenic assays were performed. The dose-dependent
survival curves of SP and non-SP cells are presented in Fig. 4D, the SP cells exhibited more
resistance than the non-SP cells. Accordingly, SP cells had larger
survival fraction values at 2 Gy irradiation than non-SP cells
(Table II). By application of the
single-hit multi-target model, the values of D0, N and
Dq of SP and non-SP cells were analyzed. As shown in
Table III, SP cells display
significantly larger values of Dq than non-SP cells,
indicating that enhanced repair of sublethal damage may contribute
to higher surviving fraction after irradiation in SP cells.
| Table IIThe survival fraction of 2
Gy-irradiated cells in different treatment. |
Table II
The survival fraction of 2
Gy-irradiated cells in different treatment.
| Treatment | SF 2 Gy (mean ±
SEM)a | P-valueb |
|---|
| non-SP | 0.570±0.046 | 0.002 |
| SP | 0.774±0.022 | |
| pcDNA3.1(+) | 0.675±0.033 | 0.004 |
|
pcDNA3.1(+)/hGRP78 | 0.791±0.012 | |
| pSuper | 0.800±0.059 | 0.010 |
| pSuper/GRP78
RNAi | 0.596±0.049 | |
| Table IIIRadiobiological parameters from
different treatment. |
Table III
Radiobiological parameters from
different treatment.
| Treatment | D0 | N | Dq |
SERDq |
|---|
| non-SP | 2.210±0.564 | 1.914±0.895 | 1.082±0.437 | 0.777±0.408 |
| SP | 2.111±0.177 | 2.811±0.607 | 2.127±0.124 | |
| pcDNA3.1(+) | 2.330±0.046 | 1.960±0.181 | 1.560±0.063 | 0.823±0.058 |
|
pcDNA3.1(+)/hGRP78 | 2.178±0.153 | 3.310±0.735 | 2.552±0.108 | |
| pSuper | 2.494±0.131 | 2.645±0.414 | 2.390±0.088 | 1.348±0.094 |
| pSuper/GRP78
RNAi | 2.349±0.139 | 1.737±0.204 | 1.277±0.110 | |
GRP78 has been hypothesized to be a key regulator of
the therapeutic resistance properties of cancer stem-like cells. We
then examined the GRP78 expression in SP cells. q-PCR analysis
demonstrated that the expression level of GRP78, which is required
for survival of embryonic stem cell precursors, was highly
expressed in SP cells (Fig.
4C).
GRP78 is necessary for maintaining the
radiation resistance properties of SP cells
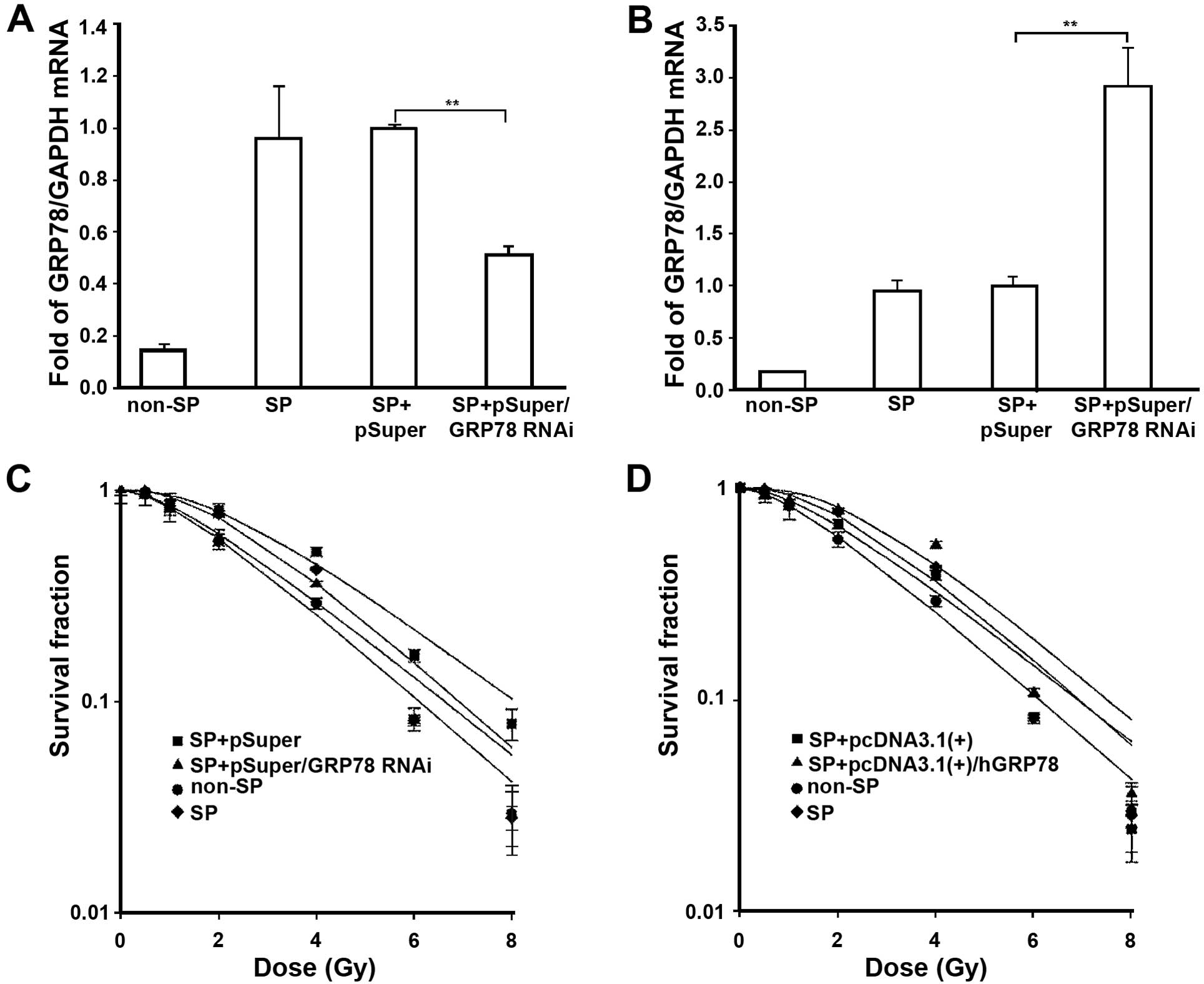
To evaluate the correlation between GRP78 expression
profile and radiation resistance, the GRP78 gene was silenced using
a small interfering RNA expressed in a pSuper vector. The SP cells
were transfected with pSuper and pSuper/GRP78 RNAi vector. After 36
h transfection, q-PCR analysis confirmed that the expression of
GRP78 was markedly suppressed in pSuper/GRP78 RNAi
vector-transfected SP cells (Fig.
5A). Then, the transfected cells were harvested by
trypsinization and resuspended for clonogenic experiments. As shown
in dose-dependent survival curves (Fig.
5C), the IR effect on SP cells treated with pSuper/GRP78 RNAi
vector was significantly stronger as compared to SP cells treated
with a scrambled control. These data demonstrated that the
silencing of GRP78 significantly enhanced the sensitivity of SP
cells to IR.
In order to further verify the role of GRP78 in the
resistance of SP cells to irradiation, we performed the
gain-of-function approach by transfection with pcDNA3.1(+)/hGRP78
plasmids transient overexpressing GRP78 into SP cells. Total mRNA
from SP cells with transfection of GRP78-expressing plasmids
displayed elevated expression of GRP78 (Fig. 5B). Then, exposed to IR, the results
indicated that GRP78 overexpression resulted in increased radiation
resistance in SP cells (Fig. 5D).
As shown in Tables II and III, the survival fraction of 2 Gy and
radiobiological parameters demonstrated that knockdown of the GRP78
gene increases the effects of IR, while overexpression of GRP78
decreases the radiation-induced cytotoxicity to SP cells.
Collectively, our data provide evidence that GRP78 upregulation in
SP cells mediates the resistance to IR.
Discussion
Despite advances in the detection and treatment of
breast cancer, mortality from this disease remains high as current
therapies are limited by the emergence of therapy resistance
(23,24). Several reports support the existence
of a subset of cells bearing stem cell characteristics within
breast tumors (20,25), giving rise to the possibility that
tumor therapy resistance is mainly due to the CSC-resistance to
antitumor treatment (26,27). Phillips et al(6) and Lagadec et al(7) reported that a population of
CD24−/low/CD44+ cells, which regards the
breast progenitor cells were resistant to radiation and the
population of CICs increased during the course of fractionated
radiation. This provided strong support for the hypothesis that
CSCs are responsible for the resistance to cancer treatment.
In order to investigate the role of CICs and the
molecular characteristics in radiation resistance, the first key
step is to identify and isolate CICs. As previously described,
reports clearly support that a functionally distinct subpopulation
of CICs can be isolated from breast cancer using either prospective
surface marker-based FACS analysis or SP cell sorting (15–17,20).
The isolation of SP cells is based on the technique initially
described by Goodell et al(14). SP assay has emerged as a promising
method for identifying cancer stem-like cell and progenitor
populations in different types of cancer (16,17,28,29).
In the present study, we isolated SP cells from human MCF-7 cell
lines, helping to further characterize the biological properties of
this cell type.
Compared with the bulk of non-SP cancer cells, SP
cells have been shown to display increased ability to form tumor
spheres and had a high clonogenic efficiency and tumorigenicity
when transplanted into immunocompromised mice (Figs. 2 and 4). The SP cells also displayed higher ES
cell marker expression (e.g., Bmi1, Nanog, Oct4 and Sox2) (Fig. 4C) and self-renewal capacity. SP
cells can differentiate into the bulk of non-SP cells (Fig. 3). Our result shows that the SP cells
are more resistant to radiation than non-SP cells, which is
consistent with findings of other reports that CICs are more
resistant to IR. These results provide direct evidence that the SP
cells we sorted bear some of the phenotypic characteristics of
CICs.
Although the putative breast CICs are considered to
be mediators of resistance to current therapies, the underlying
molecular mechanism determining the radiation resistance remains
elusive. Currently, emerging evidence indicates that the stress
response and molecular chaperones play an important role in stem
cell oncogenesis (10,30). In the present study, we found GRP78,
a major stress-inducible ER chaperone which has been reported to
play a crucial role in tumor therapeutic resistance (9,31,32),
was significantly increased in isolated SP cells. Based on the
facts, both CSCs and GRP78 are closely associated with the
resistance to cancer treatment. We therefore hypothesized that
GRP78 may be involved in the radiation resistance of SP cells.
We thus directly knocked down the expression of
GRP78 by transfecting with pSuper/GRP78 RNAi vector. The results
showed that silencing GRP78 increased the effects of radiation,
while increase of the expression of GRP78 elevated the resistance
of SP to radiation. The present study demonstrated that the
overexpression of GRP78 in SP cells mediates the resistance to
radiation.
Collectively, the present study indicated that GRP78
plays an important role in maintaining the radiation resistance of
SP cells. Based on our finding, targeting GRP78 may be a potential
therapeutic target for eliminating breast CICs. Furthermore,
combined with anti-GRP78 strategy, some cases of resistance to
radiotherapy may be overcome.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (no. 30470846). The authors
thank Professor Tianyan Zhou for the valuable input and
suggestions.
References
|
1
|
Veronesi U, Boyle P, Goldhirsch A,
Orecchia R and Viale G: Breast cancer. Lancet. 365:1727–1741. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Clarke M, Collins R, Darby S, et al:
Effects of radiotherapy and of differences in the extent of surgery
for early breast cancer on local recurrence and 15-year survival:
an overview of the randomised trials. Lancet. 366:2087–2106.
2005.PubMed/NCBI
|
|
3
|
Gebski V, Lagleva M, Keech A, Simes J and
Langlands AO: Survival effects of postmastectomy adjuvant radiation
therapy using biologically equivalent doses: a clinical
perspective. J Natl Cancer Inst. 98:26–38. 2006. View Article : Google Scholar
|
|
4
|
Rosen JM and Jordan CT: The increasing
complexity of the cancer stem cell paradigm. Science.
324:1670–1673. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gupta PB, Chaffer CL and Weinberg RA:
Cancer stem cells: mirage or reality? Nat Med. 15:1010–1012. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Phillips TM, McBride WH and Pajonk F: The
response of CD24(−/low)/CD44+ breast cancer-initiating
cells to radiation. J Natl Cancer Inst. 98:1777–1785. 2006.
|
|
7
|
Lagadec C, Vlashi E, Della Donna L, et al:
Survival and self-renewing capacity of breast cancer initiating
cells during fractionated radiation treatment. Breast Cancer Res.
12:R132010. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Luo B and Lee AS: The critical roles of
endoplasmic reticulum chaperones and unfolded protein response in
tumorigenesis and anticancer therapies. Oncogene. 32:805–818. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li J and Lee AS: Stress induction of
GRP78/BiP and its role in cancer. Curr Mol Med. 6:45–54. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mousa SA, Sudha T, Dyskin E, et al: Stress
resistant human embryonic stem cells as a potential source for the
identification of novel cancer stem cell markers. Cancer Lett.
289:208–216. 2010.PubMed/NCBI
|
|
11
|
Luo S, Mao C, Lee B and Lee AS: GRP78/BiP
is required for cell proliferation and protecting the inner cell
mass from apoptosis during early mouse embryonic development. Mol
Cell Biol. 26:5688–5697. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bartkowiak K, Effenberger KE, Harder S, et
al: Discovery of a novel unfolded protein response phenotype of
cancer stem/progenitor cells from the bone marrow of breast cancer
patients. J Proteome Res. 9:3158–3168. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wu MJ, Jan CI, Tsay YG, et al: Elimination
of head and neck cancer initiating cells through targeting glucose
regulated protein78 signaling. Mol Cancer. 9:2832010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Goodell MA, Brose K, Paradis G, Conner AS
and Mulligan RC: Isolation and functional properties of murine
hematopoietic stem cells that are replicating in vivo. J Exp Med.
183:1797–1806. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wright MH, Calcagno AM, Salcido CD,
Carlson MD, Ambudkar SV and Varticovski L: Brca1 breast tumors
contain distinct CD44+/CD24− and
CD133+ cells with cancer stem cell characteristics.
Breast Cancer Res. 10:R102008. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kondo T, Setoguchi T and Taga T:
Persistence of a small subpopulation of cancer stem-like cells in
the C6 glioma cell line. Proc Natl Acad Sci USA. 101:781–786. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Patrawala L, Calhoun T,
Schneider-Broussard R, Zhou J, Claypool K and Tang DG: Side
population is enriched in tumorigenic, stem-like cancer cells,
whereas ABCG2+ and ABCG2− cancer cells are
similarly tumorigenic. Cancer Res. 65:6207–6219. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xia S, Zhao Y, Yu S and Zhang M: Activated
PI3K/Akt/COX-2 pathway induces resistance to radiation in human
cervical cancer HeLa cells. Cancer Biother Radiopharm. 25:317–323.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Al-Hajj M, Wicha MS, Benito-Hernandez A,
Morrison SJ and Clarke MF: Prospective identification of
tumorigenic breast cancer cells. Proc Natl Acad Sci USA.
100:3983–3988. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Pastrana E, Silva-Vargas V and Doetsch F:
Eyes wide open: a critical review of sphere-formation as an assay
for stem cells. Cell Stem Cell. 8:486–498. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang P, Zhang Y, Mao L, Zhang Z and Chen
W: Side population in oral squamous cell carcinoma possesses tumor
stem cell phenotypes. Cancer Lett. 277:227–234. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Debeb BG, Xu W and Woodward WA: Radiation
resistance of breast cancer stem cells: understanding the clinical
framework. J Mammary Gland Biol Neoplasia. 14:11–17. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Stockler M, Wilcken NR, Ghersi D and Simes
RJ: Systematic reviews of chemotherapy and endocrine therapy in
metastatic breast cancer. Cancer Treat Rev. 26:151–168. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Al-Ejeh F, Smart CE, Morrison BJ, et al:
Breast cancer stem cells: treatment resistance and therapeutic
opportunities. Carcinogenesis. 32:650–658. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fillmore CM and Kuperwasser C: Human
breast cancer cell lines contain stem-like cells that self-renew,
give rise to phenotypically diverse progeny and survive
chemotherapy. Breast Cancer Res. 10:R252008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kakarala M and Wicha MS: Implications of
the cancer stem-cell hypothesis for breast cancer prevention and
therapy. J Clin Oncol. 26:2813–2820. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ablett MP, Singh JK and Clarke RB: Stem
cells in breast tumours: are they ready for the clinic? Eur J
Cancer. 48:2104–2116. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ho MM, Ng AV, Lam S and Hung JY: Side
population in human lung cancer cell lines and tumors is enriched
with stem-like cancer cells. Cancer Res. 67:4827–4833. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chiba T, Kita K, Zheng YW, et al: Side
population purified from hepatocellular carcinoma cells harbors
cancer stem cell-like properties. Hepatology. 44:240–251. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kang J, Shakya A and Tantin D: Stem cells,
stress, metabolism and cancer: a drama in two Octs. Trends Biochem
Sci. 34:491–499. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Dong D, Ko B, Baumeister P, et al:
Vascular targeting and antiangiogenesis agents induce drug
resistance effector GRP78 within the tumor microenvironment. Cancer
Res. 65:5785–5791. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Fu Y, Li J and Lee AS: GRP78/BiP inhibits
endoplasmic reticulum BIK and protects human breast cancer cells
against estrogen starvation-induced apoptosis. Cancer Res.
67:3734–3740. 2007. View Article : Google Scholar
|