Introduction
The identification and characterization of the p53
protein has relied extensively on immunological methods. Antibodies
directed against p53 protein have been valuable tools for
investigating the structure-function relationship of wild-type and
mutant p53 as well as other p53-related proteins such as p63 and
p73. Monoclonal antibodies raised against Xenopus p53 have
been demonstrated to cross-react with human p73 (1). The p53 proteins of both mouse and
human origin are detectable with PAb240, reflecting a high sequence
homology of the p53 protein between these two species. The epitope
for the monoclonal PAb421 antibody (2) is also highly conserved between the
human and the mouse (3).
The CM5 polyclonal antibody (CM5 pAb) from rabbits
immunized with mouse wild-type p53 was originally developed for
immunohistochemical analysis of mouse p53 expression (4). It has high affinity for mouse p53 and
low affinity for human p53 and is also very useful in
immunoblotting of mouse p53. CM5 pAb recognizes several antibody
epitopes of the mouse p53 protein, including the epitope for PAb240
(3). Using an anti-mouse p53 CM5
pAb to perform immunoblotting of human lung tumor tissues, we
unexpectedly found that the CM5 pAb cross-reacted with an unknown
human protein with an apparent molecular weight of 90 kDa. The
CM5-reactive protein was found to be overexpressed in human lung
tumor tissues with only minimal expression in normal adjacent lung
tissues. Therefore an investigation was carried out to identify and
characterize this protein, which may play an important role in
promoting or preventing lung cancer and could potentially be a
novel lung tumor biomarker.
Materials and methods
Patients and samples
Tissue samples were collected from patients who
underwent curative surgery for lung cancer at Maharaj Nakorn Chiang
Mai Hospital, Thailand. In each case, an adjacent normal tissue was
also collected. These specimens were immediately placed in vials,
frozen in embedded medium to preserve cell integrity, and stored at
−70°C until analyzed. The study was approved by the Ethics
Committee of the faculty of Medicine, Chiang Mai University in
compliance with the Helsinki Declaration (document no.
260/2005).
Western blot analysis
Frozen tissues were thawed, cut into small pieces
and homogenized in an SDS lysis buffer [0.5 M Tris-HCl pH 6.8, 2%
SDS (w/v) and 10% glycerol (v/v)] containing a protease inhibitor
cocktail (Complete, Mini, EDTA-free; Roche Applied Science,
Indianapolis, IN, USA). The tissue homogenate was then centrifuged
at 10,000 × g for 15 min at 4°C, after which the supernatant was
removed and the protein concentration of the supernatant was
determined using a BCA protein assay kit (Pierce Biotechnology,
Inc., Rockford, IL, USA). Protein (30 μg) from the tumor tissue and
adjacent normal tissue of each patient was resolved on 10% SDS
polyacrylamide gels under reducing conditions and
electrotransferred onto a PVDF membrane (Pierce Biotechnology,
Inc.). The membrane was blocked with 5% non-fat milk in TBS
containing 0.05% Tween-20 (TBS-Tween) for 1 h before being
incubated with polyclonal antibodies specific for mouse p53 (CM5
pAb, cat. no. NCL-p53-CM5p; Novocastra, Newcastle, UK) or
monoclonal antibodies specific for glucosidase II (cat. no.
sc-10774) or GRP-78 (cat. no. sc-13539), or GRP-94 (cat. no.
sc-53929) (all from Santa Cruz Biotechnology, Inc., Santa Cruz, CA,
USA) or human p53 (DO7; Novacastra) for 1 h at room temperature
(RT). Bound antibodies were then detected with horseradish
peroxidase (HRP)-conjugated goat anti-mouse IgG (cat. no. P 0448)
or goat anti-rabbit IgG (cat. no. P 0447) (both from
DakoCytomation, Carpinteria, CA, USA) for 1 h at RT, respectively.
After extensive washing with TBS-Tween, immunoreactive protein was
visualized with a chemiluminescence-based procedure using the ECL
Plus detection kit according to the manufacturer's protocol
(Amersham Pharmacia Biotech, Piscataway, NJ, USA).
Immunoprecipitation of the CM5
pAb-reactive protein from tumor cell lysate
Cell lysates from tumor tissues homogenized in SDS
lysis buffer containing 2% SDS were diluted with water in order to
obtain 1 mg/ml of protein and <0.05% (w/v) final concentration
of SDS. The diluted cell lysate was subjected to a preclearing step
by incubating with suspended protein G coated-agarose (cat. no.
20398; Pierce Protein Research Product) at 4°C for 45 min.
Subsequently, the precleared cell lysate was incubated with
anti-mouse p53 CM5 pAb at 4°C for 4 h before adding precleared
beads into the reaction tube and continued incubation at 4°C
overnight. After extensive washing, the immunoprecipitated proteins
were eluted from the bead particles by adding SDS lysis buffer
containing 2-mercaptoethanol and heating at 95°C for 5 min. The
eluted protein was then resolved through SDS-PAGE (10% gel) and
stained with Coomassie Blue.
Protein identification by mass
spectrophotometry
After SDS-PAGE analysis of the immunoprecipitated
CM5 pAb-reactive protein, the protein band that migrated at 90 kDa
was excised and subjected to protein identification by mass
spectrometry at the Genome Institute of Thailand (BIOTEC). The
excised protein was analyzed by liquid chromatography-tandem mass
spectrometry (LC-MS/MS) using machine model Finningan LTQ linear
ion trap mass spectrometer using a Finnigan Surveyor™ MS pump with
a flow splitter HPLC system (Thermo Scientific) according to a
previously described protocol by Mitprasat et al(5).
Characterization of the CM5 pAb-reactive
human protein in response to UV irradiation and tunicamycin-induced
endoplasmic reticulum (ER) stress
Response of CM5 pAb-reactive protein to UV
irradiation and ER stress was investigated using the A549 human
lung adenocarcinoma epithelial cell line cultured in Dulbecco's
modified Eagle's medium (DMEM) supplemented with 10% fetal bovine
serum (FBS), 100 U/ml penicillin and 100 μg/ml streptomycin. A549
(5×105) cells were seeded into a 100-mm dish containing
10 ml of DMEM and cultured at 37°C in a humidified incubator with
5% CO2 overnight. The following morning, the culture
medium was removed, and cells were irradiated with UV (15
J/m2) in order to induce DNA damage. After adding fresh
medium, culture was continued at 37°C in 5% CO2, and the
cell lysate was prepared from UV-irradiated cells at different time
points (3, 6, 24 and 36 h). In order to induce ER stress, A549
cells were cultured in DMEM containing 3 μg/ml tunicamycin (cat.
T7765; Sigma-Aldrich, St. Louis, MO, USA). Cell lysates were also
prepared from tunicamycin-treated cells at different time points
(3, 6, 24 and 36 h) and subjected to western blot analysis. Blots
were probed with specific antibodies against human p53, glucosidase
II, GRP (glucose-regulated protein)-78 and GRP-94 to characterize
changes in the level of these proteins. Blots were also probed for
GAPDH to confirm equal loading of protein.
Results
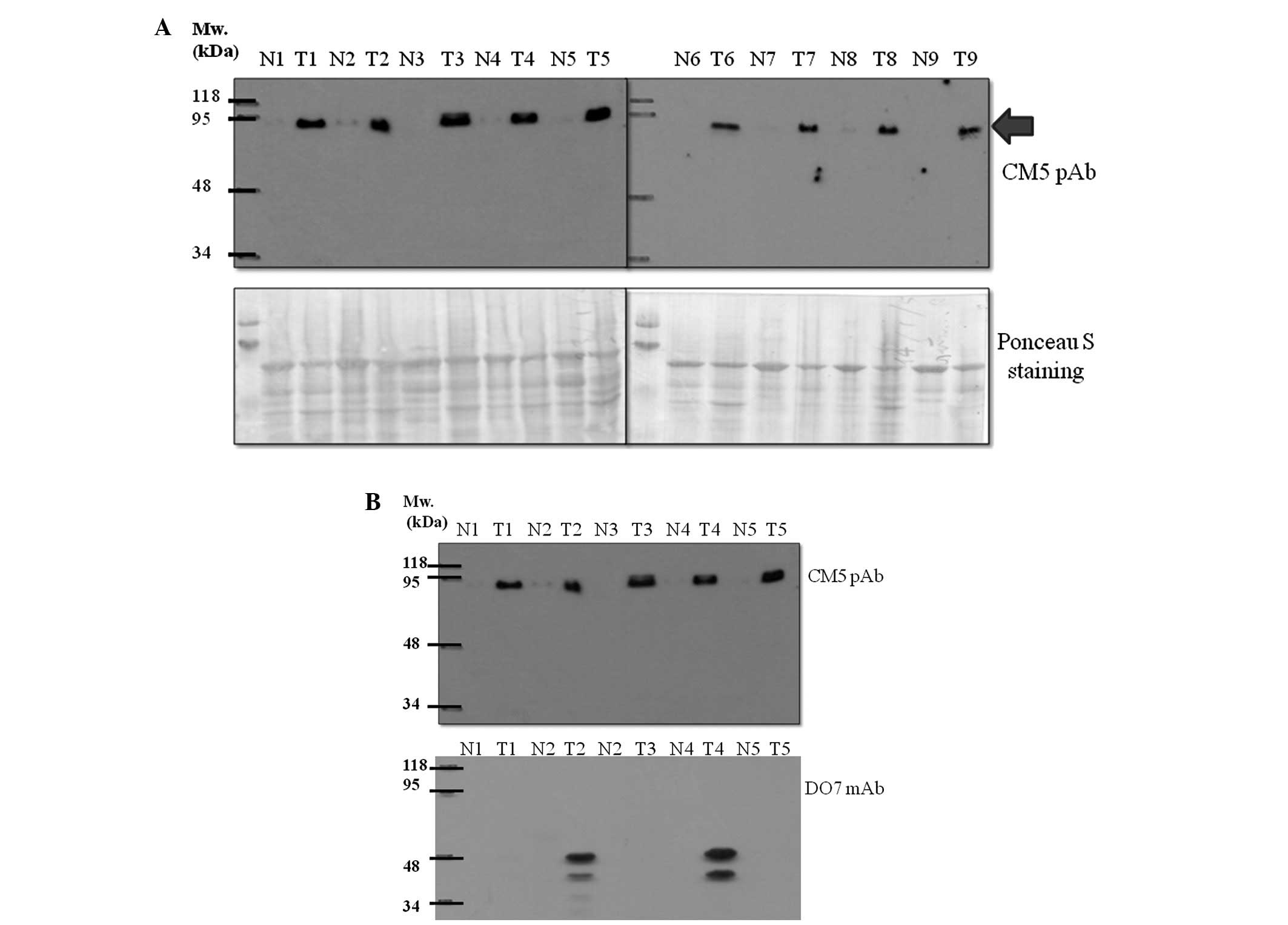
Overexpression of CM5-reactive protein in
human lung tumor tissues
Anti-mouse p53 CM5 polyclonal antibody (CM5 pAb) was
found to react with a human protein with an apparent molecular
weight of 90 kDa. Of the 37 human lung tissue samples investigated,
35 (94.6%) were found to have an increased level of this 90-kDa
protein in tumor tissues when compared to levels in the normal
tissues. The overexpression of this unknown protein in tumor vs.
normal adjacent lung tissues is shown in Fig. 1. It appeared that, in our detection
system, human p53 was not recognizable by the CM5 pAb as no band at
53 kDa was detected, although a number of tumor tissues showed a
positive band with the DO7 anti-human p53 monoclonal antibody
(Fig. 1B).
Purification and identification of the
CM5 pAb-reactive protein
To identify the unknown protein recognized by CM5
pAb, immunoprecipitation was performed in order to isolate and
purify this protein from the tumor cell lysate. The
immunoprecipitated protein was resolved through 10% polyacrylamide
gel. The protein band that migrated at ~90 kDa was excised from the
gel and subsequently subjected to protein identification using
liquid chromatography-tandem mass spectrometry (LC-MS/MS). Using MS
data searched against a mammarian protein database, the resulting
mass spectra were identified as belonging to several candidate
proteins (Table I). The most likely
candidate proteins having the highest delta Cn score were mouse
glucose-regulated protein, 78 kDa; GRP-78 (gi1304157, delta Cn
score 184.21) and human ER glucosidase II (gi2274968, delta Cn
score 174.26).
 | Table ISummary of the comparison results of
the obtained mass spectrum data from the CM5 pAb immunoprecipitated
protein against the mammalian protein database. |
Table I
Summary of the comparison results of
the obtained mass spectrum data from the CM5 pAb immunoprecipitated
protein against the mammalian protein database.
| Reference
scan(s) | Sequence | MH+ | Charge | XC | Score delta Cn | Accession Sp | | Peptides (hits)
Ions | Count |
|---|
| 1.
gi|1304157|dbj|BAA11462.1| 78 kDa glucose-regulated protein (Mus
musculus) | 184.21 | 1304157.0 | | 20 (14 5 0 1 0) | |
| 2498–2500 | -.ETAEAYLGK.- | 982.07 | 1 | 1.60 | 0.18 | 185.0 | 113 | 10/16 | 99 |
| 2506–2508 | -.M#KETAEAYLGK.- | 1257.44 | 2 | 2.87 | 0.21 | 1382.7 | 1 | 18/20 | 99 |
| 2798–2800 | -.TWNDPSVQQDIK.- | 1431.53 | 2 | 3.03 | 0.13 | 1199.5 | 2 | 18/22 | 32 |
| 2938 |
-.NQLTSNPENTVFDAK.- | 1678.78 | 2 | 4.06 | 0.24 | 1346.5 | 1 | 21/28 | 38 |
| 2970 |
-.TKPYIQVDIGGGQTK.- | 1605.82 | 2 | 3.85 | 0.32 | 1461.3 | 1 | 24/28 | 24 |
| 2972–2974 |
-.SQIFSTASDNQPTVTIK.- | 1838.01 | 2 | 4.24 | 0.28 | 1953.0 | 1 | 24/32 | 35 |
| 2986 |
-.TKPYIQVDIGGGQTK.- | 1605.82 | 2 | 3.81 | 0.26 | 1137.4 | 1 | 22/28 | 24 |
| 3032 | -.TWNDPSVQQDIK.- | 1431.53 | 2 | 2.69 | 0.04 | 490.3 | 87 | 15/22 | 32 |
| 3096 |
-.ITPSYVAFTPEGER.- | 1567.72 | 2 | 2.72 | 0.12 | 643.3 | 190 | 16/26 | 31 |
| 3104 |
-.NQLTSNPENTVFDAK.- | 1678.78 | 2 | 4.25 | 0.24 | 1623.3 | 1 | 23/28 | 38 |
| 3108 |
-.ITPSYVAFTPEGER.- | 1567.72 | 2 | 2.52 | 0.07 | 745.4 | 9 | 17/26 | 31 |
| 3118 |
-.NQLTSNPENTVFDAK.- | 1678.78 | 2 | 3.86 | 0.23 | 1605.2 | 1 | 23/28 | 38 |
| 3180 | -.NELESYAYSLK.- | 1317.43 | 2 | 3.03 | 0.11 | 1177.2 | 17 | 16/20 | 44 |
| 3192 | -.NELESYAYSLK.- | 1317.43 | 2 | 3.38 | 0.00 | 1020.5 | 54 | 16/20 | 44 |
| 3322 | -.ELEEIVQPIISK.- | 1398.63 | 2 | 2.26 | 0.12 | 915.4 | 2 | 17/22 | 25 |
| 3388 |
-.TFAPEEISAM#VLTK.- | 1553.80 | 2 | 2.15 | 0.13 | 860.0 | 7 | 17/26 | 33 |
| 3608–3610 |
-.TFAPEEISAM#VLTK.- | 1553.80 | 2 | 3.33 | 0.01 | 1210.7 | 1 | 20/26 | 33 |
| 3800 |
-.TFAPEEISAMVLTK.- | 1537.80 | 2 | 2.56 | 0.09 | 1026.5 | 3 | 20/26 | 33 |
| 3806 |
-.TFAPEEISAMVLTK.- | 1537.80 | 2 | 3.22 | 0.24 | 1334.8 | 2 | 21/26 | 33 |
| 3874 |
-.TFAPEEISAM#VLTK.- | 1553.80 | 2 | 2.29 | 0.08 | 651.7 | 1 | 20/26 | 33 |
| 2.
gi|2274968|emb|CAA04006.1| Glucosidase II (Homo
sapiens) | 174.26 | 2274968.0 | | 18 (15 3 0 0
0) | |
| 2010–2012 |
-.DPAEGDGAQPEETPR.- | 1569.57 | 2 | 4.23 | 0.40 | 1491.0 | 1 | 22/28 | 7 |
| 2654 | -.PAAVVLQTK.- | 927.12 | 2 | 2.45 | 0.17 | 604.9 | 39 | 13/16 | 15 |
| 2660 | -.PAAVVLQTK.- | 927.12 | 2 | 2.21 | 0.26 | 904.1 | 1 | 14/16 | 15 |
| 2692 | -.SIRPGLSPYR.- | 1146.32 | 2 | 2.21 | 0.11 | 698.2 | 45 | 14/18 | 12 |
| 2720 |
-.M#M#DYLQGSGETPQTDVR.- | 1961.12 | 2 | 5.13 | 0.45 | 2861.2 | 1 | 26/32 | 9 |
| 2734 |
-.M#M#DYLQGSGETPQTDVR.- | 1961.12 | 2 | 5.04 | 0.42 | 2968.0 | 1 | 26/32 | 9 |
| 2876 |
-.MM#DYLQGSGETPQTDVR.- | 1945.12 | 2 | 5.12 | 0.45 | 1984.8 | 1 | 25/32 | 9 |
| 2876 |
-.M#MDYLQGSGETPQTDVR.- | 1945.12 | 2 | 5.19 | 0.01 | 1984.8 | 1 | 25/32 | 9 |
| 2894 |
-.MM#DYLQGSGETPQTDVR.- | 1945.12 | 2 | 4.91 | 0.00 | 1566.9 | 1 | 23/32 | 9 |
| 2894 |
-.M#MDYLQGSGETPQTDVR.- | 1945.12 | 2 | 4.91 | 0.45 | 1566.9 | 1 | 23/32 | 9 |
| 3012 |
-.MMDYLQGSGETPQTDVR.- | 1929.12 | 2 | 4.54 | 0.16 | 2176.9 | 1 | 24/32 | 9 |
| 3018 | -.LVAIVDPHIK.- | 1105.36 | 2 | 2.53 | 0.17 | 1689.6 | 1 | 16/18 | 18 |
| 3022 | -.LVAIVDPHIK.- | 1105.36 | 2 | 2.50 | 0.12 | 1473.5 | 1 | 15/18 | 18 |
| 3028 |
-.MMDYLQGSGETPQTDVR.- | 1929.12 | 2 | 5.10 | 0.41 | 1951.7 | 1 | 24/32 | 9 |
| 3146 |
-.DENSVELTMAEGPYK.- | 1683.82 | 2 | 4.76 | 0.48 | 2096.1 | 1 | 22/28 | 7 |
| 3168 |
-.DENSVELTMAEGPYK.- | 1683.82 | 2 | 4.83 | 0.35 | 1994.7 | 1 | 22/28 | 7 |
| 3292 | -.SLLLSVNAR.- | 973.15 | 2 | 2.51 | 0.20 | 1262.5 | 13 | 14/16 | 16 |
| 3314 | -.SLLLSVNAR.- | 973.15 | 2 | 2.47 | 0.08 | 986.5 | 52 | 13/16 | 16 |
| 3.
gi|224970|prf||1205208A heat shock protein hsp70 | 74.23 | 224970.0 | | 8 (5 3 0 0 0) | |
| 2096–2098 |
-.VEIIANDQGNR.- | 1229.33 | 2 | 3.63 | 0.00 | 1872.9 | 1 | 19/20 | 99 |
| 2326 |
-.VEIIANDQGNR.- | 1229.33 | 2 | 2.81 | 0.00 | 1083.6 | 2 | 16/20 | 99 |
| 2330 |
-.VEIIANDQGNR.- | 1229.33 | 2 | 3.14 | 0.11 | 1103.4 | 23 | 16/20 | 99 |
| 3372 |
-.IINEPTAAAIAYGLDK.- | 1660.89 | 2 | 3.66 | 0.00 | 1810.4 | 1 | 24/30 | 99 |
| 3538 |
-.FEELNM#DLFR.- | 1330.49 | 2 | 3.09 | 0.28 | 1014.2 | 1 | 15/18 | 99 |
| 3550 |
-.FEELNM#DLFR.- | 1330.49 | 2 | 3.48 | 0.20 | 986.9 | 25 | 14/18 | 99 |
| 3580 |
-.IINEPTAAAIAYGLDK.- | 1660.89 | 2 | 4.50 | 0.00 | 1673.4 | 1 | 25/30 | 99 |
| 3590 |
-.IINEPTAAAIAYGLDK.- | 1660.89 | 2 | 3.97 | 0.00 | 1549.3 | 1 | 24/30 | 99 |
| 4.
gi|229552|prf||754920A albumin | 60.19 | 229552.0 | | 6 (6 0 0 0 0) | |
| 2902–2904 |
-.HLVDEPQNLIK.- | 1306.49 | 2 | 2.70 | 0.09 | 710.5 | 3 | 15/20 | 5 |
| 2966 |
-.KVPQVSTPTLVEVSR.- | 1640.91 | 2 | 3.25 | 0.27 | 657.3 | 1 | 18/28 | 34 |
| 2992 |
-.KVPQVSTPTLVEVSR.- | 1640.91 | 2 | 3.77 | 0.16 | 976.9 | 1 | 20/28 | 34 |
| 3030 | -.YLYEIAR.- | 928.07 | 2 | 2.28 | 0.00 | 593.3 | 21 | 11/12 | 30 |
| 3128 |
-.VPQVSTPTLVEVSR.- | 1512.73 | 2 | 3.03 | 0.21 | 1104.1 | 2 | 18/26 | 34 |
| 5.
gi|2392283|pdb|1DKG|D chain D, crystal structure of the nucleotide
exchange factor grpe bound to Th | 28.23 | 2392283.0 | | 3 (2 1 0 0 0) | |
| 3372 |
-.IINEPTAAALAYGLDK.- | 1660.89 | 2 | 3.66 | 0.00 | 1810.4 | 1 | 24/30 | 99 |
| 3580 |
-.IINEPTAAALAYGLDK.- | 1660.89 | 2 | 4.50 | 0.00 | 1673.4 | 1 | 25/30 | 99 |
| 3590 |
-.IINEPTAAALAYGLDK.- | 1660.89 | 2 | 3.97 | 0.00 | 1549.3 | 1 | 24/30 | 99 |
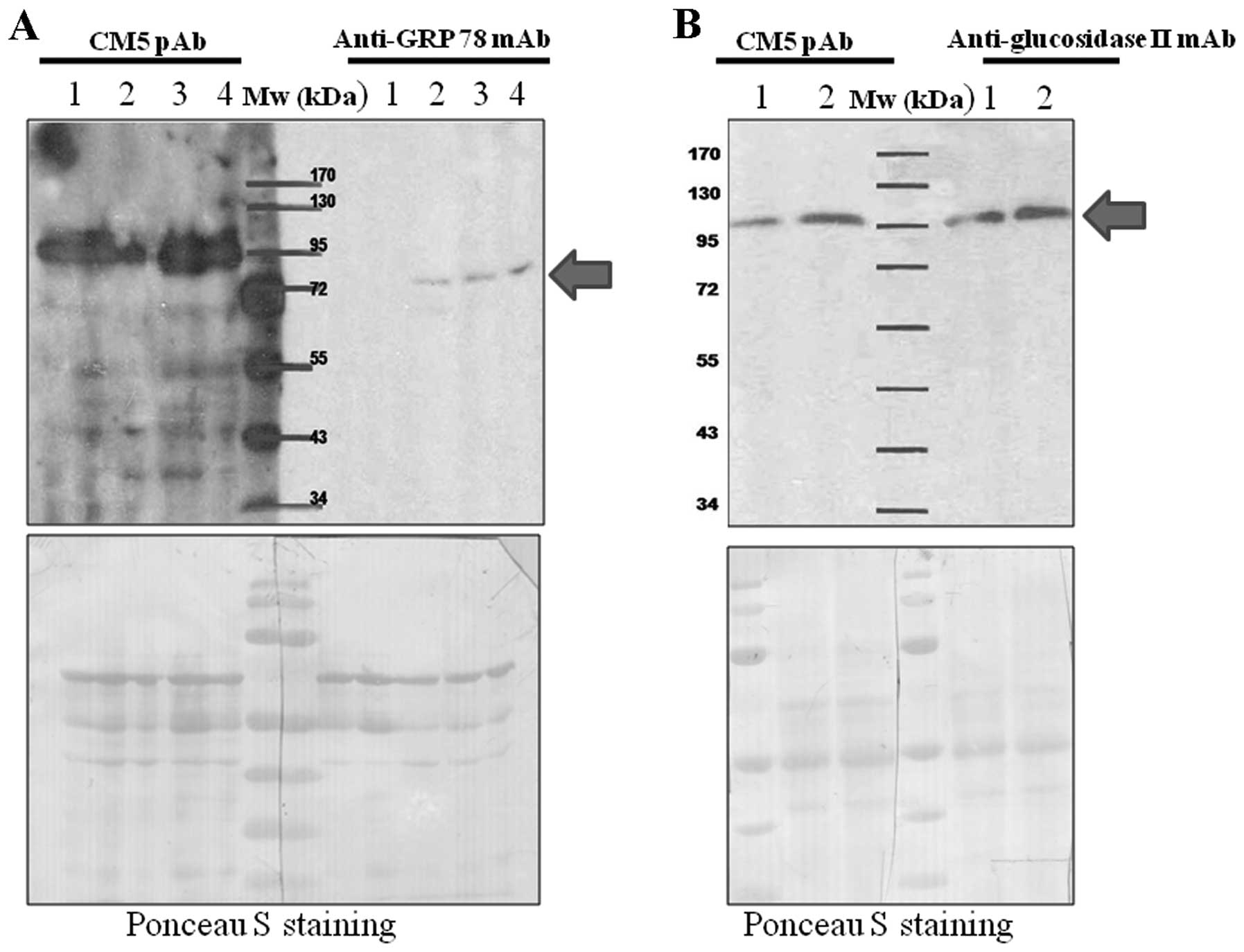
Verification of the protein identified by
mass spectrophotometry
In order to verify the identification results of
LC-MS/MS, protein molecular weight comparison and
immunoprecipitation were performed. Whole cell lysate from various
lung tumor tissues known to overexpress the CM5 pAb-reactive
protein was resolved through 10% polyacrylamide gel, and separated
proteins were transfered onto a PVDF membrane. The blots were cut
in half and immunodetected with either CM5 pAb and anti-GRP-78 or
anti-glucosidase II. As shown in Fig.
2, the unknown protein recognized by CM5 pAb had an apparent
molecular weight similar to glucosidase II but not GRP-78.
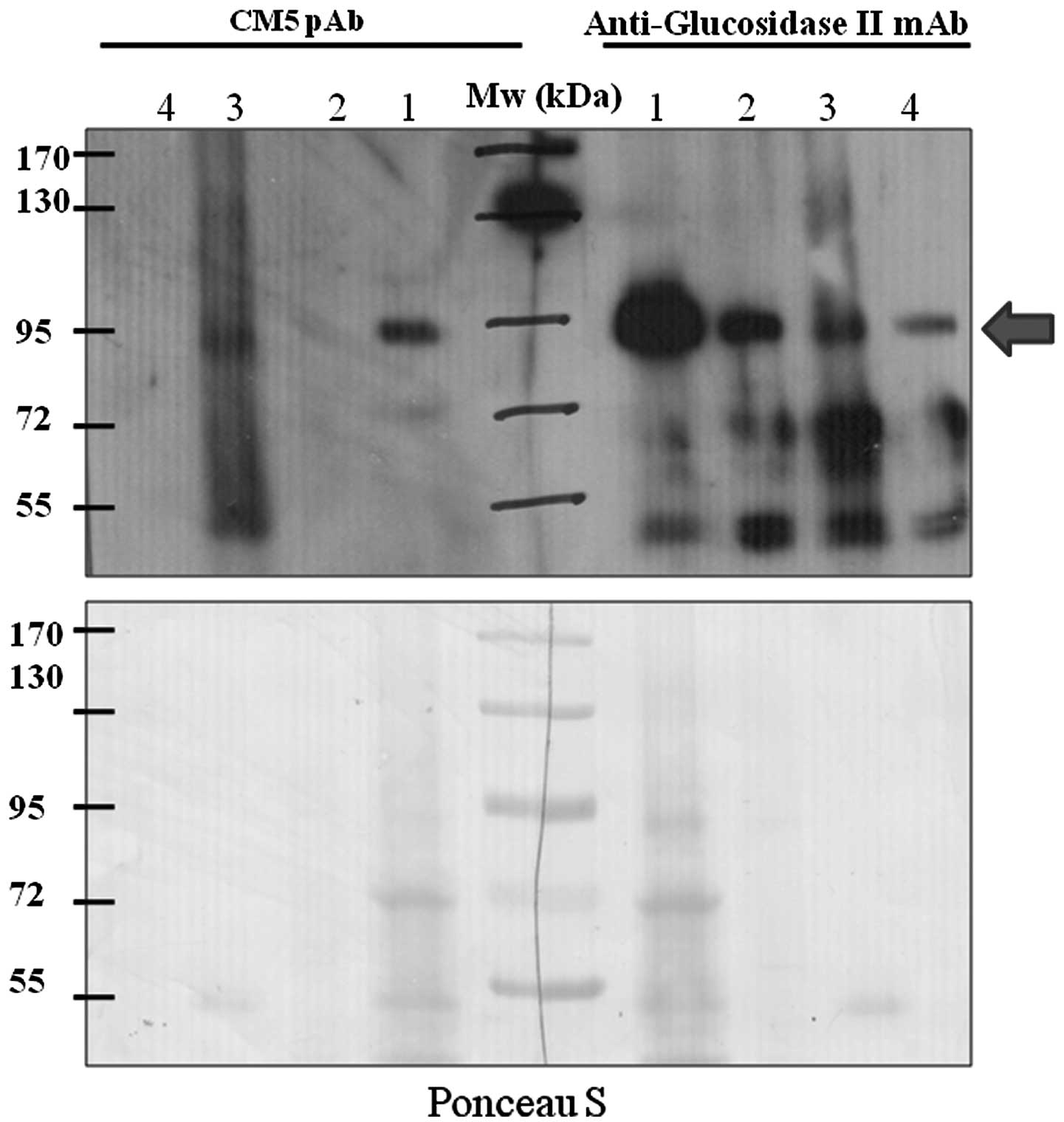
Therefore, immunoprecipitation was performed using anti-glucosidase
II mAb. The immunoprecipitated protein was resolved through
SDS-PAGE and subjected to immunodetection with CM5 pAb. The
immunoprecipitated protein was recognizable by anti-glucosidase II
mAb thus indicating the successful immunoprecipitation process.
Notably, the immunoprecipitated protein was also recognized by CM5
pAb (Fig. 3). In addition, the
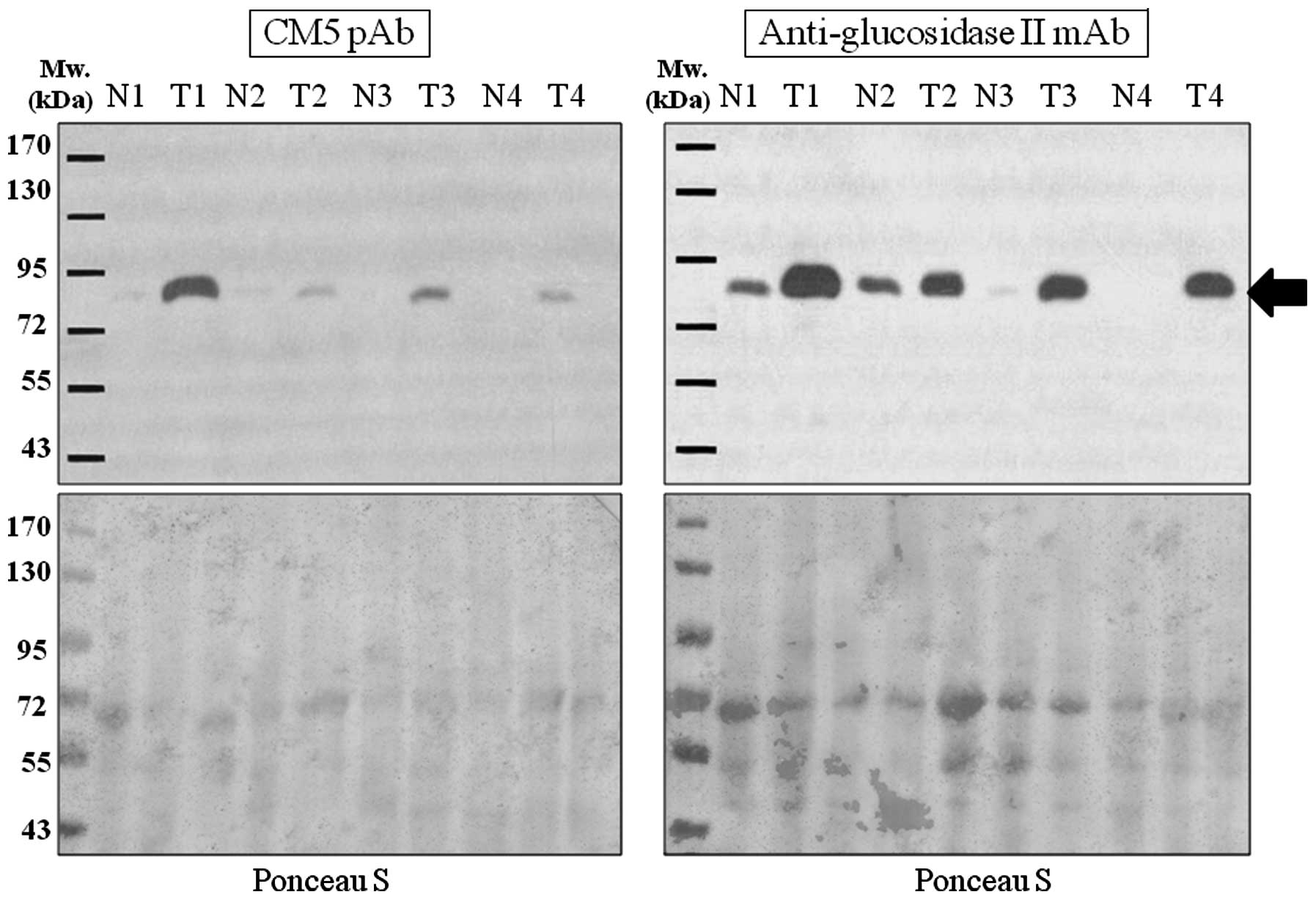
results of the western blot analysis demonstrated an identical
expression pattern between the proteins recognized by CM5 pAb and
anti-glucosidase II mAb in each lung tumor tissue tested
(representative western blot results are shown in Fig. 4).
Expression of glucosidase II in response
to ER stress and UV irradiation
The A549 human lung adenocarcinoma epithelial cell
line was used as a cell model to investigate the expression pattern
of glucosidase II in response to UV-irradiation and
tunicamycin-induced ER stress in comparison to p53. The whole cell
lysate was prepared at different time points (0, 3, 6, 24 and 36 h)
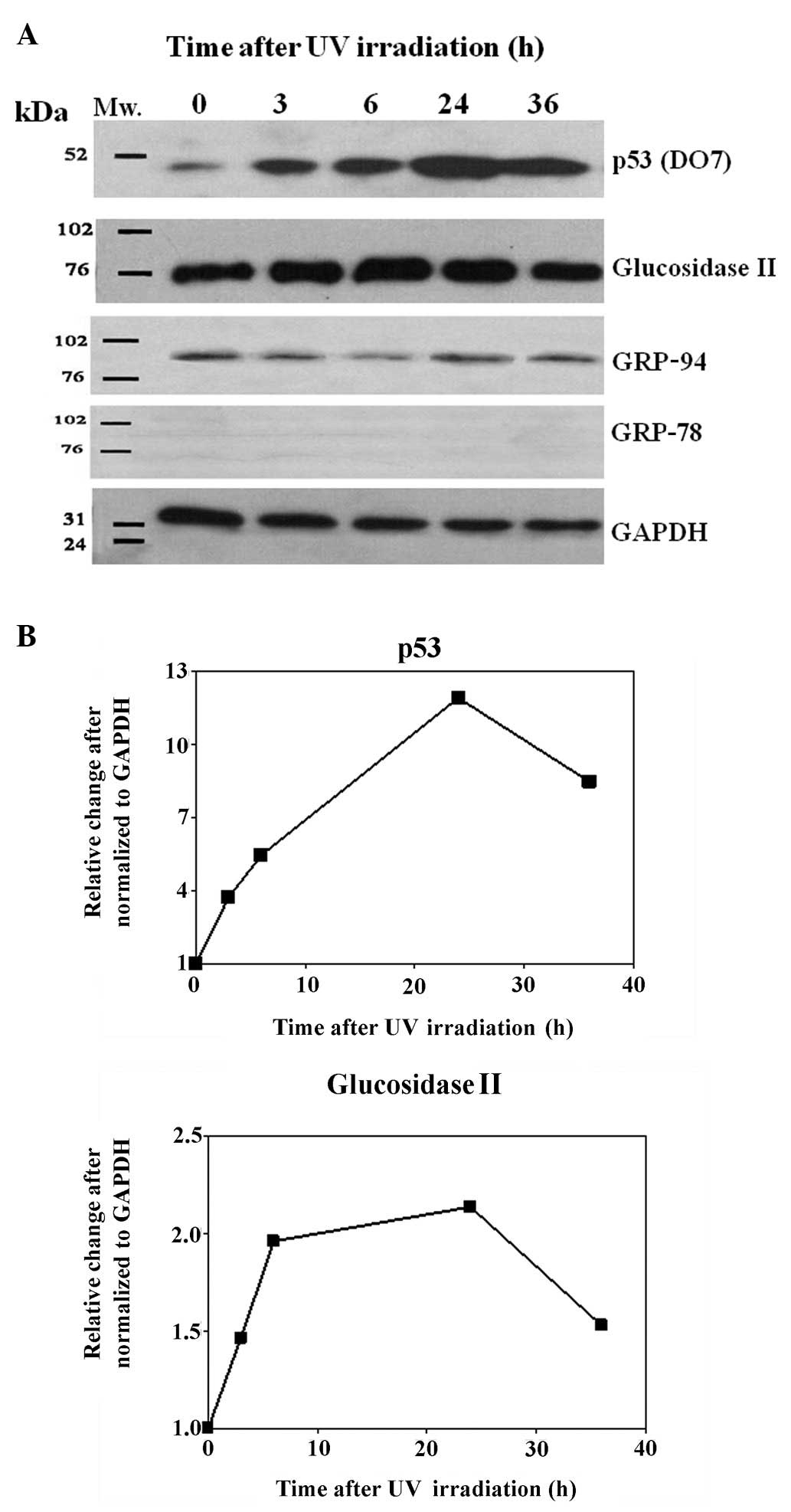
following treatment. As shown in Fig.
5A, the protein levels of both p53 and glucosidase II were
increased in the UV-irradiated cells. The induction appeared as
rapidly as 3 h after treatment and continued to increase even after
24 h. The maximum induction was observed at 24 h for both p53 and
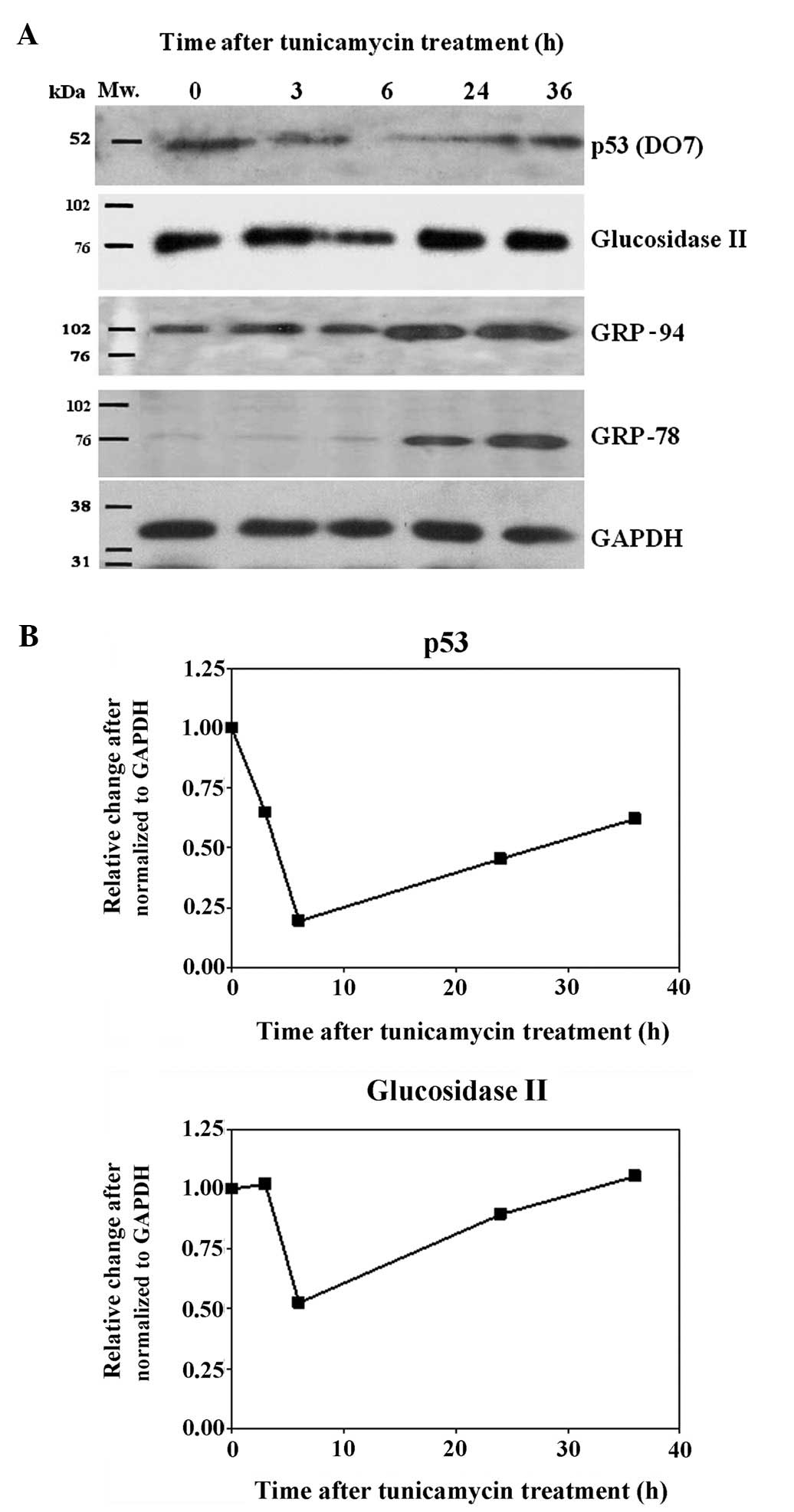
glucosidase II (Fig. 5B). In
contrast, levels of both p53 and glucosidase II were found to be
suppressed in the tunicamycin-treated cells and the maximum
reduction appeared at 6 h after treatment for both p53 and
glucosidase II (Fig. 6A and B).
Discussion
Using the CM5 anti-mouse p53 polyclonal antibody to
performed western blot analysis of human lung tumor tissues, we
fortuitously discovered that the protein level of glucosidase II
was greatly increased in a high proportion of lung tumor tissues,
while the corresponding normal adjacent tissues showed barely
detectable expression levels. Glucosidase II plays a key role in
the processing of N-linked oligosaccharide chains of glycoprotein
and is involved in the quality control mechanism of glycoprotein
folding (6). However, to the best
of our knowledge, overexpression of glucosidase II in tumor tissues
has not been previously described. ER is the site of folding for
protein destined for different compartments of the cell. As the
newly synthesized proteins enter the ER lumen, they are covalently
modified by the addition of a pre-assembled oligosacharide composed
of 2 N-acetylglucosamines, 9 mannoses and 3 glucoses. Glycosylation
of the newly synthesized protein is believed to aid in the
increasing hydrophilicity of the un-structured proteins.
Subsequently, the glycosylated protein chains enter a
glycoprotein-dedicated chaperone system comprising calnexin and
calreticulin (7). Access to the
calnexin/calreticulin system requires removal of the two outermost
glucose residues by the sequential action of glucosidase I and II,
respectively (8). Removal of the
third glucose is also mediated through glucosidase II action, which
is required to dissociate folding substrates from calnexin and the
release of the native proteins from the ER and transport to their
final destination. However, if the protein is improperly folded,
the folding sensor UDP-glucose:glycoprotein glucosyltransferase 1
(UGT1) will add back a terminal glucose to promote re-association
of the non-native protein and calnexin, thus prolonging their time
in the ER folding environment. Cycles of de-/re-glycosylation may
be extended until the protein released from calnexin fufills the
quality requirement (reviewed in ref. 6). Therefore, balance of glucosidase II
and UGT1 activity is crucial in order to maintain protein quality
control of the ER.
Disturbance in the folding capacity of the ER caused
by a variety of endogenous and exogenous stimulants could initiate
a cellular stress condition known as ER stress. ER stress is
initially induced to re-establish ER homeostasis through the
activation of an integrated intracellular signal transduction
termed the unfolded protein response (UPR). However, when ER stress
is too severe or prolonged, the pro-survival function of the UPR
turns into a toxic signal, which is predominantly executed by
mitochondrial apoptosis (9). Since
glucosidase II plays a major role in the release of proteins from
the ER folding system, cells with overexpression of glucosidase II
may be impervious to ER stress. Even overloaded with misfolded
proteins, UPR and apoptosis may not be triggered in glucosidase
II-overexpressed cells. In agreement with this hypothesis,
inhibition of glucosidase II has been reported to reduce
proliferation and induce apoptosis (10,11) of
tumor cells.
There is increasing evidence that the UPR is
compromised in a large variety of human tumors (reviewed in ref.
12). More importantly, several
recent studies have demonstrated that interfering with the
activation of different arms of the UPR (i.e., PERK-eIF2α-ATF4
axis) or altering the levels of the ER molecular chaperone
GRP-78/BIP (a master regulator of ER function and the UPR) can
inhibit tumor growth (13–15). This evidence indicates that not only
the UPR is compromised in tumors but that it contributes to
survival or growth of the cancer cells. Therefore, inhibitors of
glucosidase II could also be regarded as potential anticancer
agents (16,17).
The discovery of the induction of glucosidase II
protein in lung tumor tissues through the use of the antibody
against p53 indicates their structure similarity, and prompted us
to elucidate the connection between these two proteins. Although
our findings remain to be confirmed in different cell lines and
primary cultures from patients, we showed that the pattern of
changes in the protein levels of p53 and glucosidase II in response
to ER stress and genotoxic stress was similar in A549 cells. The
p53 tumor-suppressor protein becomes stabilized and activated in
response to a number of stressful stimuli, i.e., hypoxia,
nucleotide depletion or oncogene activation (18). The main form of stress that
activates p53 is genotoxic stress (19), which if left unchecked can lead to
loss of genomic integrity and tumorigenesis. Activation of p53
allows it to carry out its function as a tumor suppressor protein
through a number of controlling endpoints i.e., cell cycle arrest
or apoptosis (20). However, it was
not recognized until recently that ER stress also plays a pivotal
role in modulating p53 activity. The protein level (21) and function (22) of p53 have been reported to be
reduced or suppressed in response to ER stress. This inhibition is
believed to help ensure that p53 activation is restricted to agents
that induce genotoxic stress (23).
However, if the ER stress is not properly responded or if the UPR
is compromised by an altered level of regulator proteins, i.e.
glucosidase II, this may lead to the overproduction of misfolded
proteins which may be oncogenic and together with the suppression
of p53 function, the cell may finally become cancerous. The fact
that both p53 and glucosidase II respond to ER stress and
genotoxicity in a similar fashion indicates the possible connection
between the two proteins; our findings warrant further
investigation.
In conclusion, we demonstrated that human
glucosidase II protein was frequently overexpressed in human lung
tumor tissues. The high frequency of glucosidase II overexpression,
which to the best of our knowledge has not been previously
described, indicates the crucial role of glucosidase II in lung
tumorigenesis and its potential value as a biomarker for aiding the
screening and/or diagnosis of lung cancer. For example, a highly
sensitive glucosidase assay system could be developed to measure
glucosidase enzymatic activity from non-invasive samples i.e.,
exhaled breath condensate (EBC) or pleural effusion, in order to
screen for lung cancer. Further investigation concerning the
underlying mechanism of the protein induction and its relationship
with p53, genotoxic stress and ER stress are warranted.
Acknowledgements
This study was financially sponsored by the Research
Chair Grant, National Sciences and Technology Development Agency
(Thailand), the National Research Council of Thailand (NRCT) and
the Faculty of Associated Medical Sciences, Chiang Mai University.
We thank Dr Tim R. Cressey, Harvard School of Public Health and
Chiang Mai University for his review and editing of the
manuscript.
References
|
1
|
Le Bras M, Delattre V, Bensaad K, Blandino
G and Soussi T: Monoclonal antibodies raised against Xenopus p53
interact with human p73. Oncogene. 21:1304–1308. 2002.PubMed/NCBI
|
|
2
|
Harlow E, Crawford L, Pim D and Williamson
N: Monoclonal antibodies specific for simian virus 40 tumor
antigens. J Virol. 39:861–869. 1981.PubMed/NCBI
|
|
3
|
Lane DP, Stephen CW, Midgley CA, et al:
Epitope analysis of the murine p53 tumour suppressor protein.
Oncogene. 12:2461–2466. 1996.PubMed/NCBI
|
|
4
|
Midgley CA, Owens B, Briscoe CV, Thomas
DB, Lane DP and Hall PA: Coupling between gamma irradiation, p53
induction and the apoptotic response depends upon cell type in
vivo. J Cell Sci. 108:1843–1848. 1995.PubMed/NCBI
|
|
5
|
Mitprasat M, Roytrakul S, Jiemsup S,
Boonseng O and Yokthongwattana K: Leaf proteomic analysis in
cassava (Manihot esculenta, Crantz) during plant
development, from planting of stem cutting to storage root
formation. Planta. 233:1209–1221. 2011.PubMed/NCBI
|
|
6
|
Ruddock LW and Molinari M: N-glycan
processing in ER quality control. J Cell Sci. 119:4373–4380. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ellgaard L, Molinari M and Helenius A:
Setting the standards: quality control in the secretory pathway.
Science. 286:1882–1888. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang N, Daniels R and Hebert DN: The
cotranslational maturation of the type I membrane glycoprotein
tyrosinase: the heat shock protein 70 system hands off to the
lectin-based chaperone system. Mol Biol Cell. 16:3740–3752. 2005.
View Article : Google Scholar
|
|
9
|
Lee AS and Hendershot LM: ER stress and
cancer. Cancer Biol Ther. 5:721–722. 2006. View Article : Google Scholar
|
|
10
|
Magyar JE, Gamberucci A, Konta L, et al:
Endoplasmic reticulum stress underlying the pro-apoptotic effect of
epigallocatechin gallate in mouse hepatoma cells. Int J Biochem
Cell Biol. 41:694–700. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gamberucci A, Konta L, Colucci A, et al:
Green tea flavonols inhibit glucosidase II. Biochem Pharmacol.
72:640–646. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ranganathan AC, Adam AP, Zhang L and
Aguirre-Ghiso JA: Tumor cell dormancy induced by p38SAPK and
ER-stress signaling: an adaptive advantage for metastatic cells?
Cancer Biol Ther. 5:729–735. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cho HY, Thomas S, Golden EB, et al:
Enhanced killing of chemo-resistant breast cancer cells via
controlled aggravation of ER stress. Cancer Lett. 282:87–97. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jakobsen CH, Størvold GL, Bremseth H, et
al: DHA induces ER stress and growth arrest in human colon cancer
cells: associations with cholesterol and calcium homeostasis. J
Lipid Res. 49:2089–2100. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang LJ, Chen S, Wu P, et al: Inhibition
of MEK blocks GRP78 up-regulation and enhances apoptosis induced by
ER stress in gastric cancer cells. Cancer Lett. 274:40–46. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Elbein AD: Glycosidase inhibitors as
antiviral and/or antitumor agents. Semin Cell Biol. 2:309–317.
1991.PubMed/NCBI
|
|
17
|
Elbein AD: Glycosidase inhibitors:
inhibitors of N-linked oligosaccharide processing. FASEB J.
5:3055–3063. 1991.PubMed/NCBI
|
|
18
|
Horn HF and Vousden KH: Coping with
stress: multiple ways to activate p53. Oncogene. 26:1306–1316.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Colman MS, Afshari CA and Barrett JC:
Regulation of p53 stability and activity in response to genotoxic
stress. Mutat Res. 462:179–188. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Amundson SA, Myers TG and Fornace AJ Jr:
Roles for p53 in growth arrest and apoptosis: putting on the brakes
after genotoxic stress. Oncogene. 17:3287–3299. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Pluquet O, Qu LK, Baltzis D and Koromilas
AE: Endoplasmic reticulum stress accelerates p53 degradation by the
cooperative actions of Hdm2 and glycogen synthase kinase 3β. Mol
Cell Biol. 25:9392–9405. 2005.PubMed/NCBI
|
|
22
|
Qu L and Koromilas AE: Control of tumor
suppressor p53 function by endoplasmic reticulum stress. Cell
Cycle. 3:567–570. 2004.PubMed/NCBI
|
|
23
|
Stavridi ES and Halazonetis TD: p53 and
stress in the ER. Genes Dev. 18:241–244. 2004. View Article : Google Scholar : PubMed/NCBI
|