Introduction
Approximately 80–90% of primary liver cancers are
hepatocellular carcinomas (HCCs). HCC is one of the three most
common types of tumors worldwide apart from gastric and esophageal
cancer. More than one million individuals are newly diagnosed with
liver cancer each year. China is the country with the highest
incidence of liver cancer. Therapeutic approaches for the treatment
of HCC can be classified into three categories: potentially
curative, palliative and symptomatic. Although liver cancer can be
surgically removed, the 2-year recurrence rate is as high as 50%.
In the past 20 years, the relative 5-year survival rate of patients
with liver cancer has increased by 3% from 4 to 7% (1). Moreover, due to underlying cirrhosis,
systemic therapy with classical cytotoxic drugs is poorly tolerated
and ineffective. Sorafenib was approved by the FDA for the
treatment of unresectable HCC and was recommended as the first-line
therapy for HCC patients who cannot benefit from therapies that are
potentially more effective, such as TACE or local ablative therapy
(2). A subanalysis was conducted to
assess the multiple risk factors involved in HCC oncogenesis, and
this revealed that sorafenib can also be beneficial for patients
with alcohol-related HCC or hepatitis B or hepatitis C infection
(3). Despite improvements in
survival after sorafenib administration, the prognosis for patients
with this stage of HCC is still poor, with a median overall
survival rate of 6.5–10.7 months (4). Accordingly, there is an urgent need to
identify new therapeutic agents for the treatment of hepatoma in
clinical practice.
Chinese medicine has achieved satisfactory results
in the treatment of liver cancer, by improving symptoms and quality
of life, and preventing recurrence and metastasis. Chemotherapy
agents commonly used in many clinical anticancer compounds are
extracted from herbal or animal substances (5,6). The
root of Cynanchum auriculatum Royle ex Wight, a traditional
Chinese medicine, has been used to nourish the blood and enhance
immunity in China and other Asian countries for a long time. C-21
steroidal glycosides is one species of important biological active
compounds widely found in the plants of the Asclepiadaceae
family, which has been shown to effectively remove hydroxyl
radicals and oxygen free radicals, regulating immunity, and
protecting liver and nerve cells (7,8). To
date, many types of C-21 steroidal glycosides have been purified
from Cynanchum bungei Decne. For example, caudatin, one species of
C-21 steroidal, has been mainly isolated from the root of Cynanchum
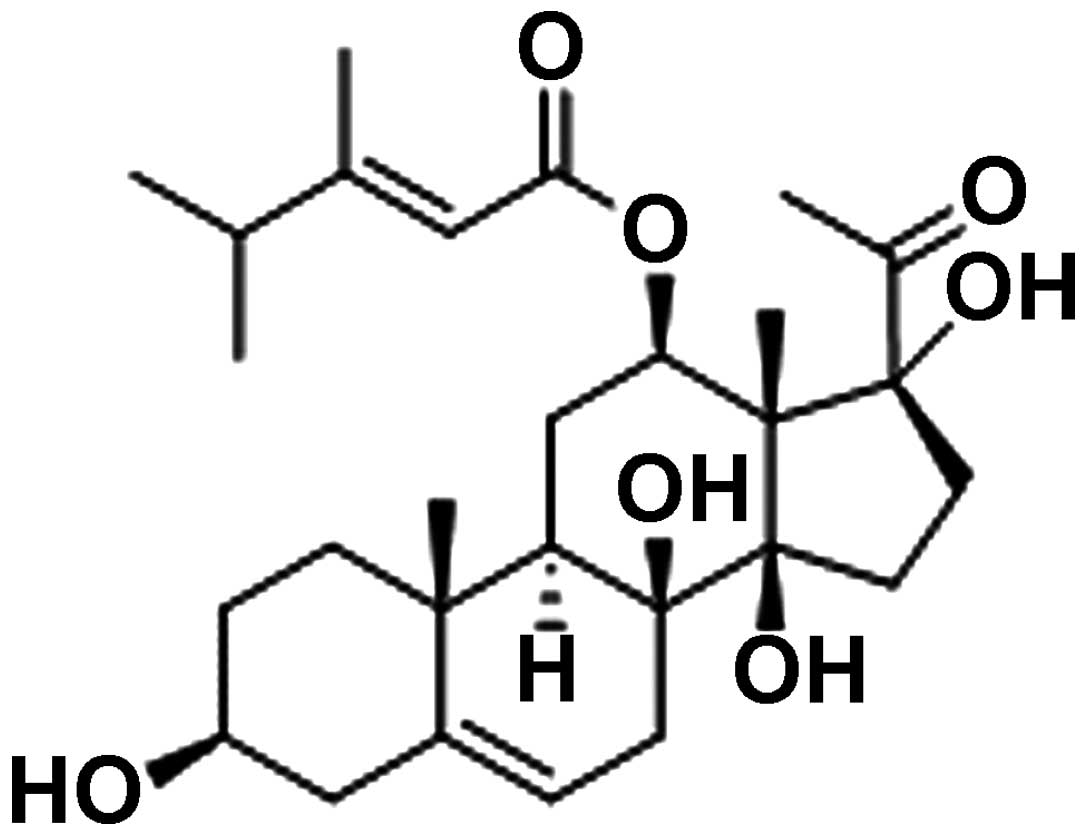
bungei Decne in China. The chemical structure of caudatin is shown
in Fig. 1. Although studies have
shown that caudatin induces apoptosis in SMMC-7721 cells (9), the underlying mechanisms of its action
are not completely understood. The purpose of the present study was
to determine the effects of caudatin on SMMC-7721 cell growth and
metastasis and to investigate the possible molecular mechanisms.
Our results showed that caudatin modulated Wnt/β-catenin signaling
and inhibited cell proliferation and induced cell apoptosis in
SMMC-7721 cells. In addition, a non-toxic dose of caudatin
suppressed the migration of SMMC-7721 cells through the inhibition
of MMP-2, MMP-9 and VEGF secretion.
Materials and methods
Materials
Caudatin (95.6% purity) was isolated from the root
tuber of Cynanchum auriculatum by our research group
(10). Caudatin was dissolved in
dimethyl sulfoxide (DMSO) and was used in all experiments. MTT
[3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide] was
obtained from Sigma (St. Louis, MO, USA). Lysis buffer was
purchased from Beyotime Institute of Biotechnology (Haimen, China).
Antibodies (GSK3β, COX-2, MMP-2, MMP-9, GAPDH and goat anti-rabbit
IgG-HRP) were obtained from BioWorld (Dublin, OH, USA). β-catenin
was obtained from Proteintech (Chicago, IL, USA). VEGF was
purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
Cell culture
Human hepatoma cell line SMMC-7721 was purchased
from the Cell Bank of Xiangya Central Experimental Laboratory.
Cells were cultured in RPMI-1640 medium supplemented with 10% fetal
bovine serum (FBS), 100 U/ml penicillin and 100 μg/ml streptomycin
(all available from Invitrogen, Grand Island, NY, USA).
Cytotoxicity assay
The cytotoxic effect of caudatin on SMMC-7721 cells
was analyzed by MTT assay. SMMC-7721 cells at mid-log phase were
seeded in a 96-well plate at a density of 5×103
cells/well in 100 μl medium. After a 24-h incubation, cells were
exposed to 0.1% DMSO (used as control in all experiments) or 15,
30, 60, 90 and 120 μM caudatin for 24, 48 and 72 h. After
treatment, 20 μl of 5 mg/ml MTT was added, and the cells were
incubated for 4 h at 37°C. The supernatant was discarded, and 150
μl of DMSO was added to each well. The mixture was shaken on a mini
shaker at room temperature for 10 min, and the spectrophotometric
absorbance was measured using the Multiskan Spectrum microplate
reader (Thermo Fisher Scientific, Waltham, MA, USA) at 490 and 630
nm (absorbance 490 nm, reference 630 nm). Triplicate experiments
were performed in a parallel manner for each concentration, and the
results are presented as means ± SD. The net OD490nm −
OD630nm was taken as the index of cell viability. The
net absorbance from the wells of cells cultured with DMSO was taken
as the 0% inhibitory rate. The percent inhibitory rate (IR %) of
the treated cells was calculated by the formula: IR % = 1 −
(OD490nm − OD630nm)
treated/(OD490nm − OD630nm)control ×
100%.
Cell cycle analysis
Cell cycle distribution was determined using a cell
cycle staining kit (Multisciences, USA). Cells treated with 0.1%
DMSO or increasing concentrations of caudatin (12.5, 25 and 50 μM)
for 48 h were trypsinized and washed twice with PBS, and fixed in
75% ethanol overnight at −20°C. The fixed cells were washed with
PBS twice before incubation with 1 ml Reagent A for 30 min at 37°C.
DNA content and the cell cycle were determined using a FACScan
laser flow cytometer (FACSVerse; Becton-Dickinson, Franklin Lakes,
NJ, USA). Data were analyzed using FlowJo software.
Apoptosis assay
SMMC-7721 cells were treated with 0.1% DMSO or
increasing concentrations of caudatin (12.5, 25 and 50 μM) for 48
h. The cells were then harvested, washed and resuspended with PBS.
Apoptotic cells were determined with an Alexa Fluor 488 Annexin
V/Dead Cell Apoptosis kit (Invitrogen, Carlsbad, CA, USA) according
to the manufacturer’s protocol. Briefly, the cells were washed and
subsequently incubated for 15 min at room temperature in the dark
in 100 μl of 1X Annexin binding buffer containing 5 μl of Annexin
V-FITC and 2 μl of propidium iodide (PI). Afterward, apoptosis was
analyzed using a FACScan laser flow cytometer (FACSVerse). Data
were analyzed using FlowJo software.
Transwell assay
The migratory ability of SMMC-7721 cells was
examined by Transwell assay. The SMMC-7721 cells with 80%
confluence were washed once with PBS and serum-starved in the basal
media (without serum and growth supplements) for 12 h. The
harvested cells were counted and reached a volume of
1×106 cells/ml. The cell migration Transwell chamber
(8.0 μm/6.5 mm; Corning Incorporated, New York, NY, USA) was
inserted at an angle of 45°, gently pressed down to avoid
generating bubbles, and 100 μl of the cell suspension was added to
each chamber. DMSO (control) or caudatin (12.5, 25 and 50 μM) was
added to the cell suspension with 1 ml of fresh medium (10% FBS) in
the lower wells (24-well plate) (Corning Incorporated). After 16 h
of incubation, the cells were fixed and stained. Five randomly
chosen fields were counted and photographed using a fluorescence
microscope (Leica DM IL).
Western blot analysis
Cells were cultured until mid-log phase and then
incubated with different concentrations of caudatin for 24 h.
Proteins were isolated by lysis buffer and measured with a BCA
protein assay. Protein samples were separated on 10%
SDS-polyacrylamide gels (SDS-PAGE) and transferred onto PVDF
membranes (Millipore, Billerica, MA, USA). After being blocked with
1% BSA in TBST (Tris-buffered saline with Tween-20) for 2 h,
membranes were incubated with the primary antibodies overnight at
4°C. Blots were washed and incubated with the secondary antibodies
for 2 h at room temperature. Membranes were again washed three
times with TBST and developed using enhanced chemiluminescence
(Beyotime). Membranes were then exposed to film.
Real-time quantitative PCR (qPCR)
Total RNA was extracted using TRIzol reagent
(Generay, Shanghai, China) according to the manufacturer’s
instructions. Total RNA (2 μg) was used for cDNA synthesis with
random hexamer primers. qPCR was carried out using the CFX Connect
Real-Time PCR system. Reactions were performed according to IQ™
SYBR Green Supermix instructions (Bio-Rad Laboratories, Hercules,
CA, USA) in triplicate in three independent experiments. The primer
sequences are provided in Table I.
The ΔΔCT method was used for qPCR determination. GAPDH
was used as a housekeeping gene to normalize the variability in
expression levels.
 | Table IPrimer sequences used for real-time
quantitative PCR (5′-3′). |
Table I
Primer sequences used for real-time
quantitative PCR (5′-3′).
| Gene | Forward primer | Reverse primer |
|---|
| hGAPDH |
AGAAGGCTGGGGCTCATTTG |
AGGGGCCATCCACAGTCTTC |
| hCOX-2 |
AAGACAGATCATAAGCGAGGGC |
AAACCGTAGATGCTCAGGGACT |
| hMMP-2 |
CTTCAAGGACCGGTTCATTTGG |
ATGAGCTTGGGGAAGCCAGGA |
| hMMP-9 |
GCGCTGGGCTTAGATCATTC |
TCAGGGCGAGGACCATAGAG |
Statistical analysis
All the data are expressed as the means ± standard
deviation (SD). Statistical analysis was performed using the
Student’s t test, and P<0.05 was considered to indicate a
statistically significant result.
Results
Cytotoxic effect of caudatin on SMMC-7721
cells
The inhibitory effect of caudatin on cell growth was
assessed by the commonly used MTT assay at different intervals (24,
48 and 72 h) of treatment. Caudatin treatment significantly
inhibited the growth of SMMC-7721 cells in both a concentration-
and time-dependent manner at 24 and 48 h (Fig. 2). The inhibitory effect decreased at
72 h, possibly due to drug degradation and cell resistance. The
IC50 values at 24, 48 and 72 h were 89.49, 54.43 and
118.07 μM. We choose the 48 h time-point to carry out the follow-up
experiments.
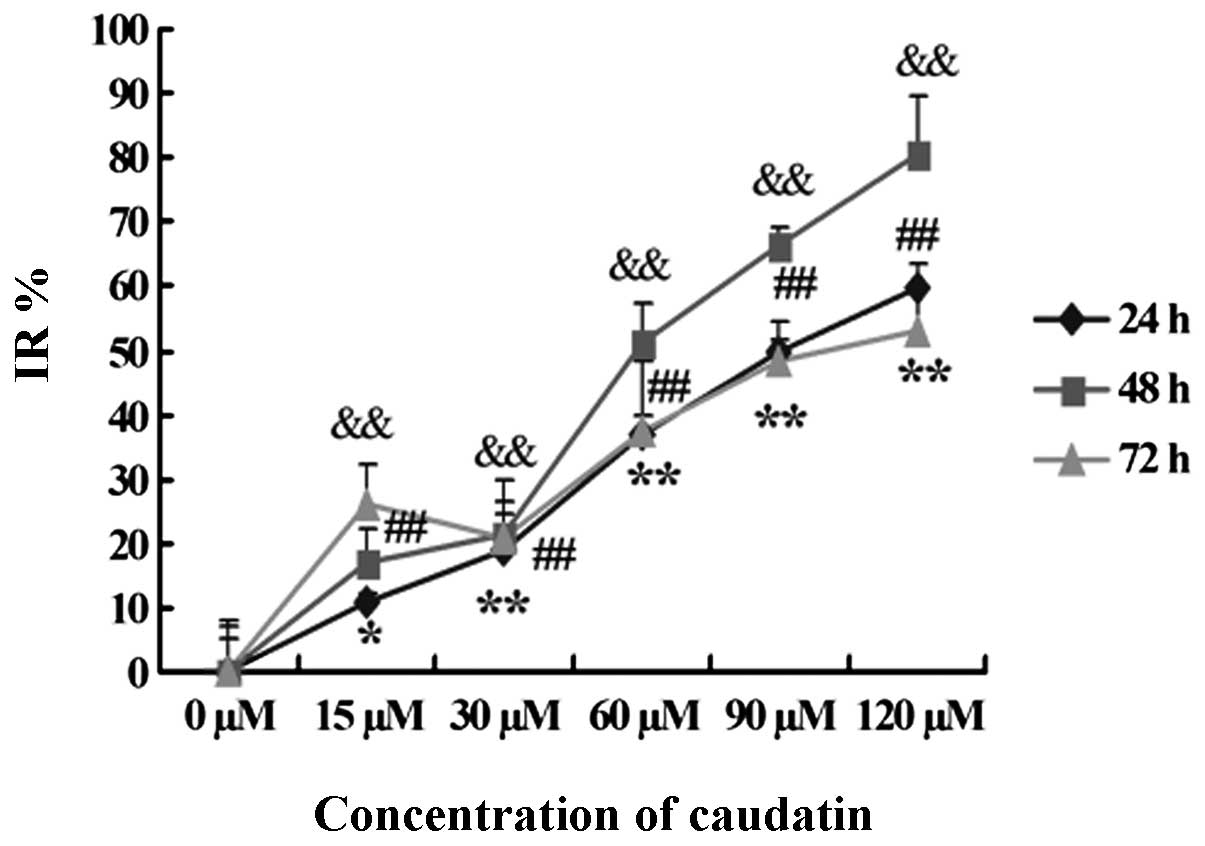 | Figure 2Cytotoxic effect of caudatin on
SMMC-7721 cells. The results shown are the mean of three parallel
experiments (triplicate wells) for each concentration (15, 30, 60,
90 and 120 μM) at 24, 48 and 72 h. *Indicates 24 h,
#indicates 48 h, &indicates 72 h.
*,#,&P<0.05,
**,##,&&P<0.01. IR, inhibitory rate |
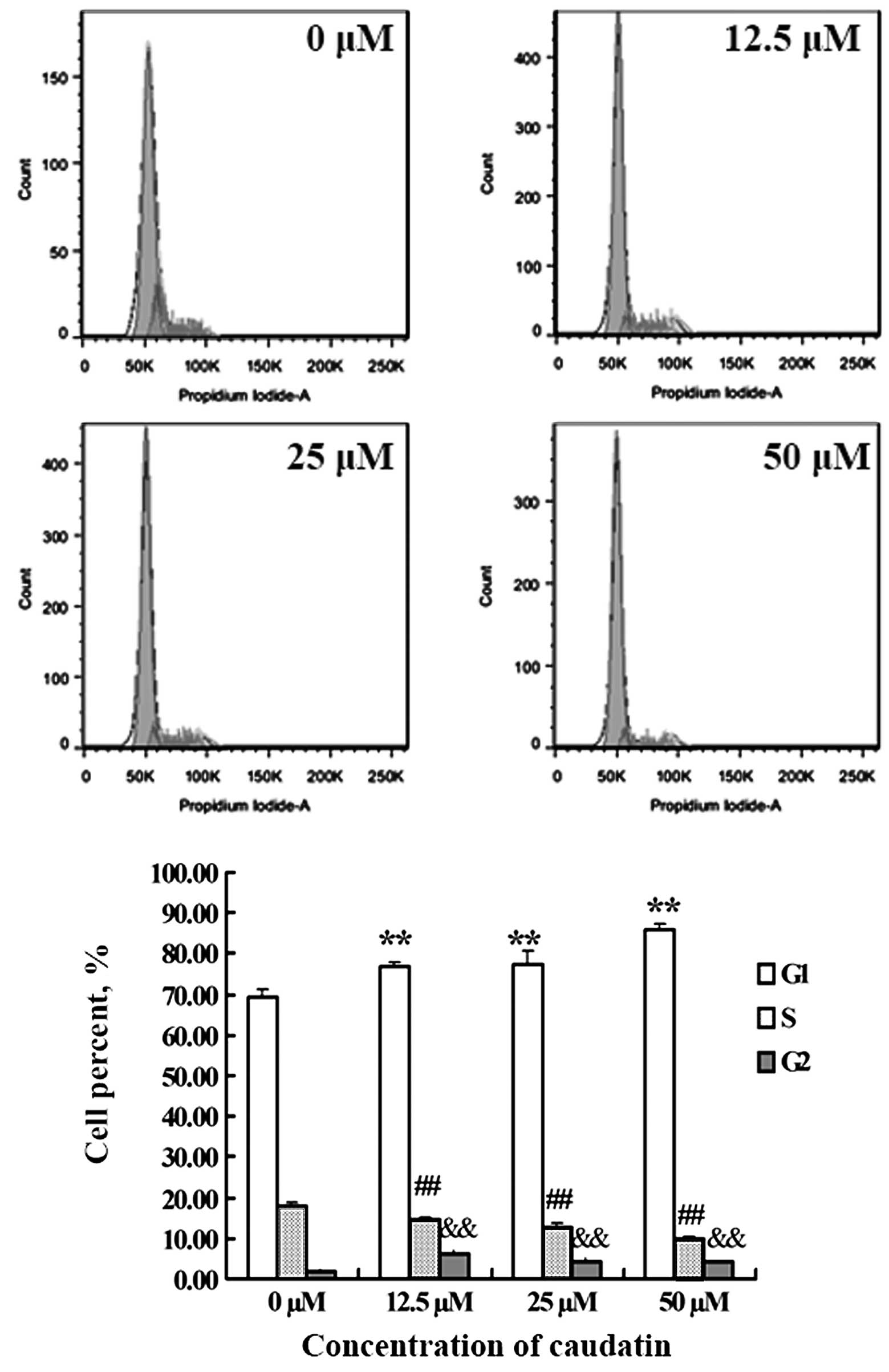
Caudatin treatment causes G2 phase cell
cycle arrest
To test whether caudatin affects the cell cycle of
SMMC-7721 cells, cells treated with DMSO or different
concentrations of caudatin for 48 h were subjected to flow
cytometric analysis after DNA staining. As shown in Fig. 3, exposure of SMMC-7721 cells to
growth suppressive concentrations of caudatin resulted in a
statistically significant increase in the G2 phase cell population
which was accompanied by a decrease in the S phase population. For
example, the percentage of cells in the G2 phase increased by
~2-fold following the treatment of SMMC-7721 cells with 12.5 μM
caudatin when compared with the control. This indicated that
caudatin suppressed SMMC-7721 cell proliferation associated with
cell cycle arrest at the G2 phase.
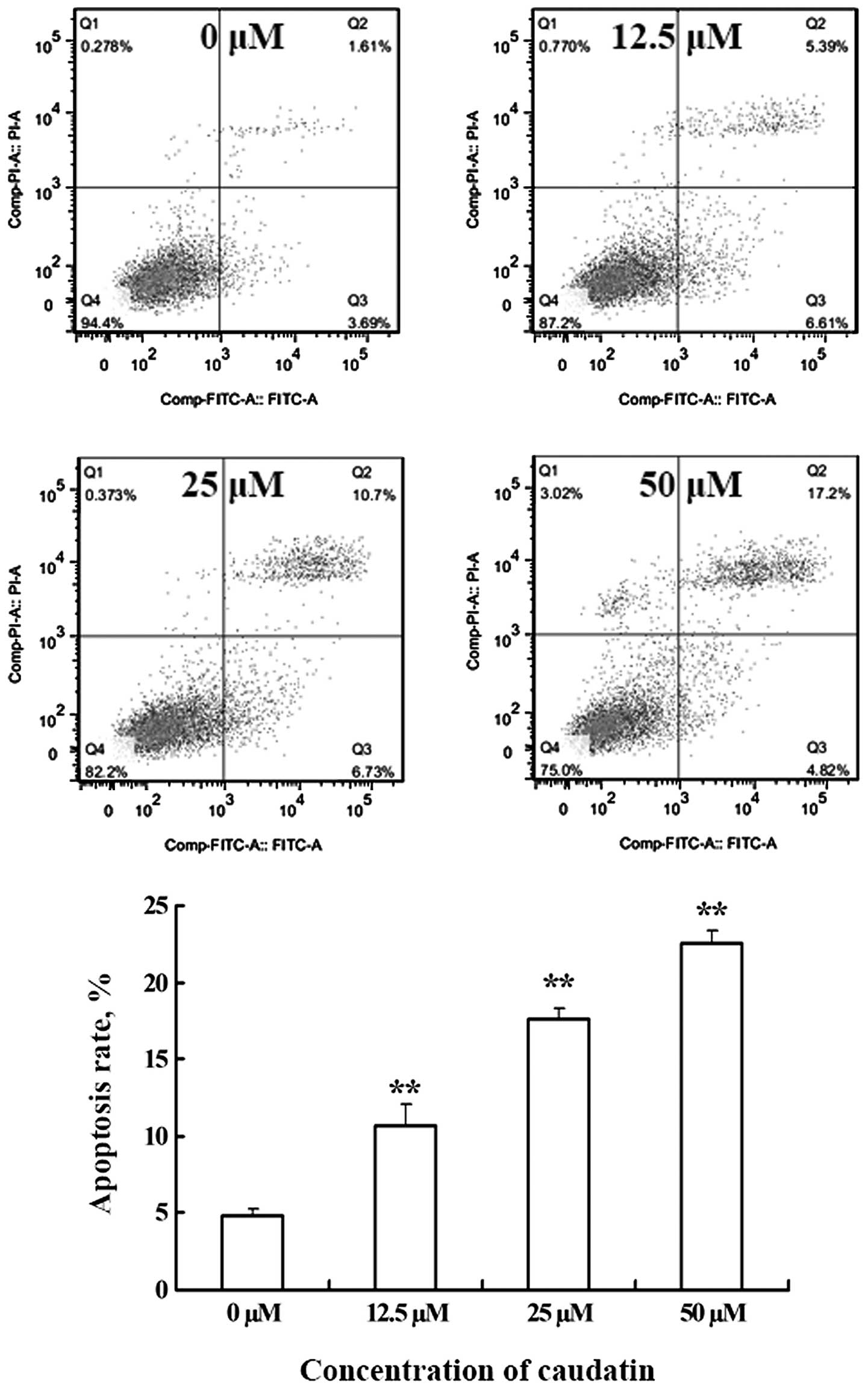
Pro-apoptotic effect of caudatin on
SMMC-7721 cells
The occurrence of apoptosis was obtained by double
staining of the cultures with PI and Annexin V-FITC. Annexin V is a
protein that binds with high affinity to phosphatidylserine, which
is translocated from the inner to the outer membrane leaflet early
in the apoptotic process. As shown in Fig. 4, living cells stained negative for
both PI and Annexin V-FITC (Q4). Caudatin-induced cells, on the
other hand, showed many Annexin V-positive, PI-negative cells (Q3),
indicating that they were at an early stage of apoptosis. The
double positive staining of particular cells revealed that these
cells were in a late apoptotic stage or were necrotic (Q2). These
findings provided strong evidence that caudatin has a pro-apoptosis
effect on SMMC-7721 cells.
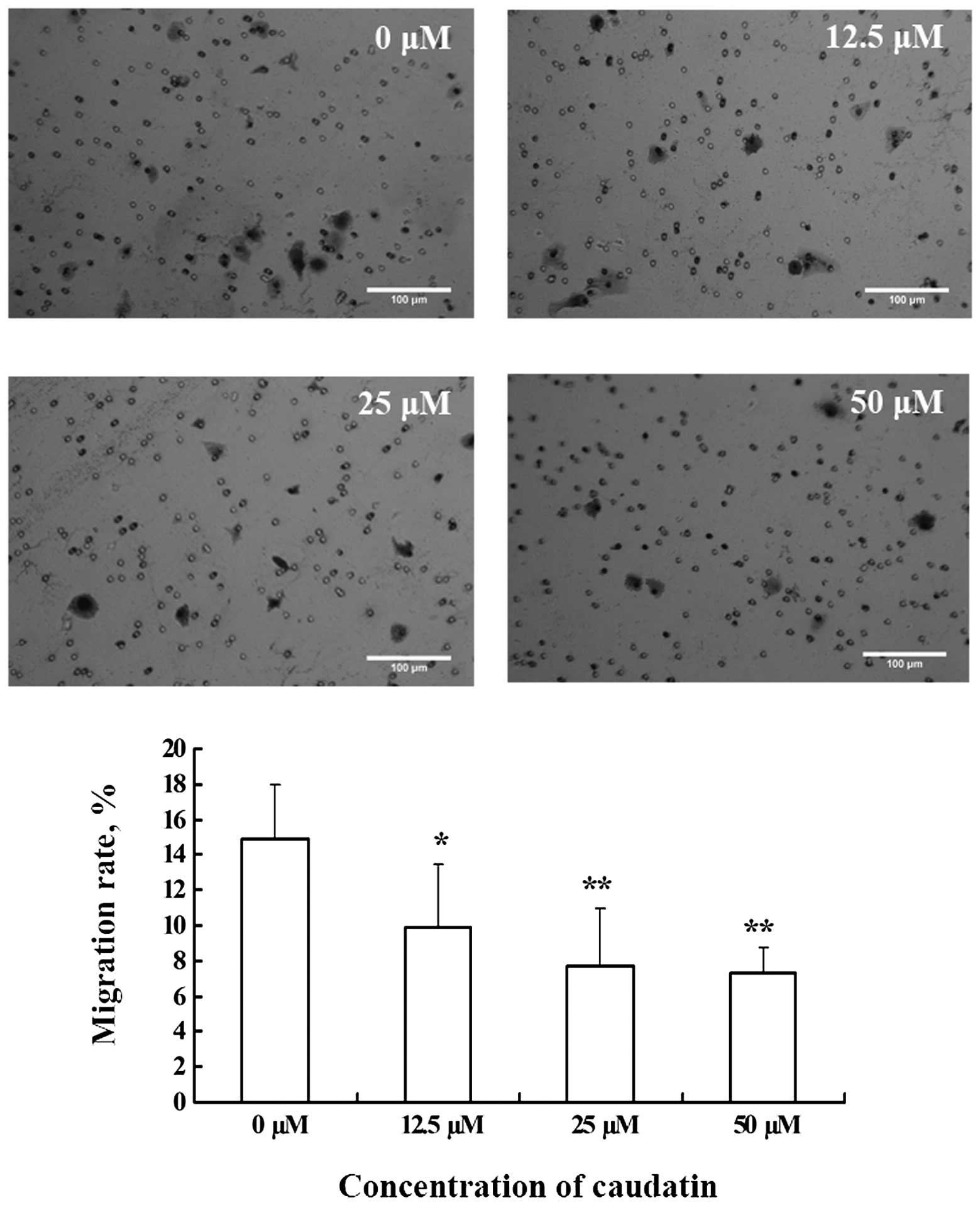
Caudatin inhibits SMMC-7721 cell
migration
The migration of SMMC-7721 cells is a prerequisite
for tumor metastasis. We determined the effect of caudatin on
SMMC-7721 cell migration stimulated with FBS using the Boyden
Chamber assay. After stimulation for 16 h, a high number of cells
migrated to the lower side of the Transwell filter in the control
group. However, addition of caudatin to the top chamber
significantly reduced the number of migratory cells (Fig. 5).
Caudatin treatment decreases expression
of β-catenin
The prominent role of Wnt/β-catenin signaling in
tumorigenesis has attracted considerable interest in the drug
discovery research community, and identification of inhibitors for
this signaling pathway has been a goal of researchers. To assess
whether caudatin affects the expression of β-catenin, SMMC-7721
cells were exposed to various concentrations of caudatin for 24 h.
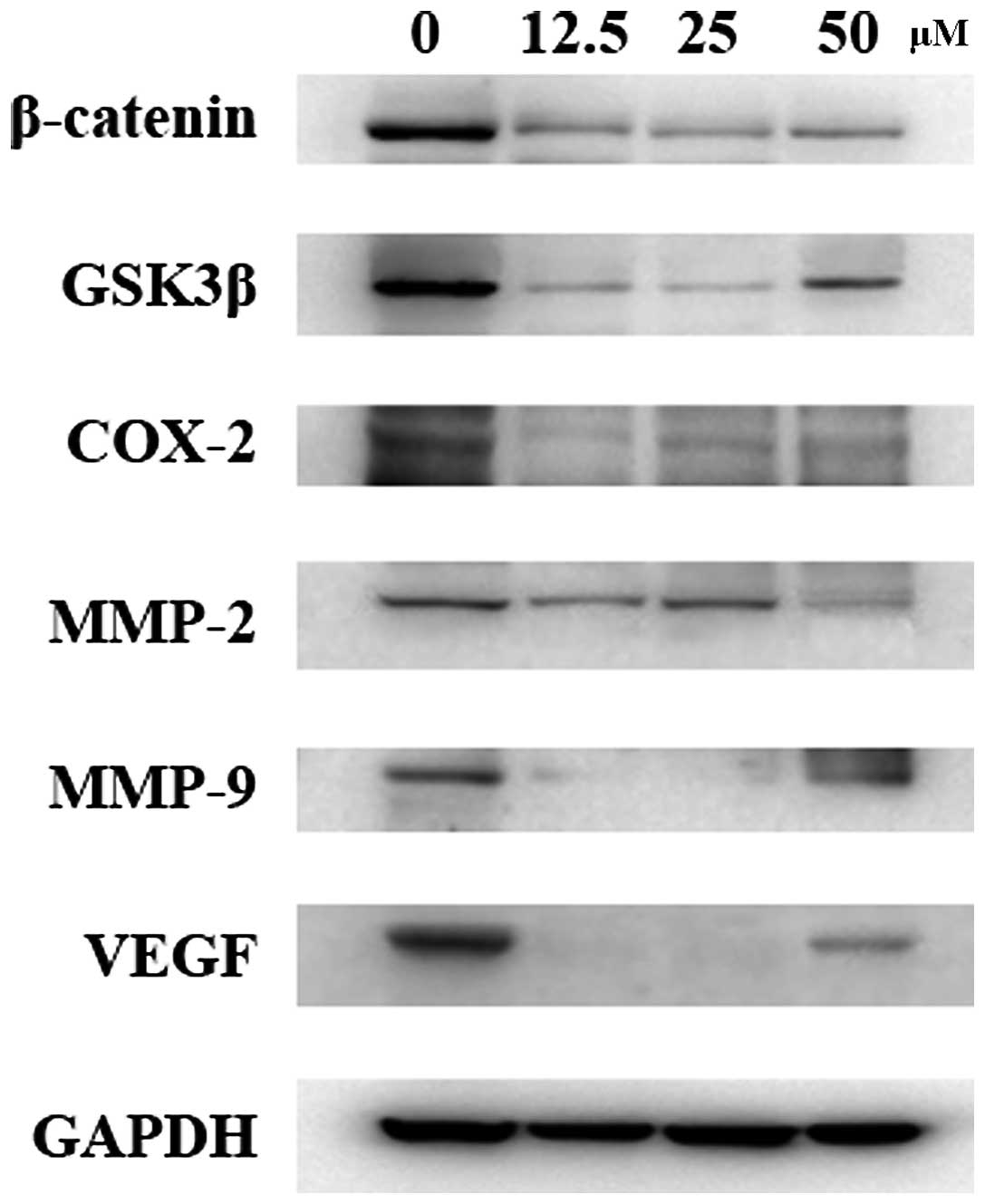
Western blot results indicated that caudatin significantly
downregulated the expression of β-catenin which was also associated
with a significant decrease in GSK3β levels in SMMC-7721 cells
(Fig. 6), suggesting that caudatin
regulates the β-catenin pathway by inducing β-catenin degradation.
In addition, treatment of SMMC-7721 cells with caudatin caused a
significant reduction in the level of COX-2, MMP-2, MMP-9 and VEGF,
the downstream targets of β-catenin.
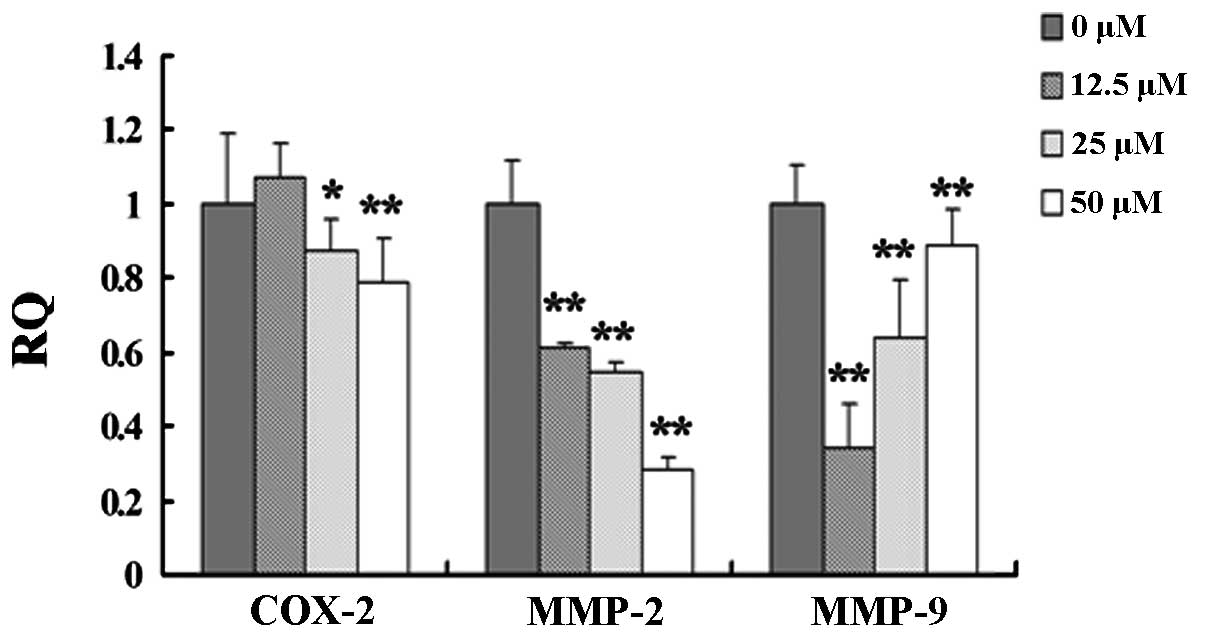
Caudatin suppresses the expression of
β-catenin pathway downstream target genes
Since cox-2, mmp-2 and mmp-9
are the downstream target genes of the Wnt/β-catenin pathway
(11), we examined whether caudatin
downregulates the expression of these genes in SMMC-7721 cells. At
the dose range of 12.5–50 μM, caudatin was able to inhibit
cox-2, mmp-2 and mmp-9 mRNA expression in a
dose-dependent manner. The inhibitory effect on cox-2 and
mmp-2 mRNA expression was positively correlated with the
dose of caudatin while mmp-9 exhibited a negative
correlation (Fig. 7).
Discussion
Considering the high recurrence and high transfer
characteristics of liver cancer, treatment after tumor recurrence
and metastasis is a key factor for prolonging survival time.
Metastasis and invasion are also important factors affecting the
effective treatment of liver cancer. At present, basic research on
the metastasis and invasion of liver cancer is ongoing, but no
important breakthrough has been reported.
Wnt signaling is an evolutionarily conserved
signaling cascade with imperative roles in regulating developmental
decisions as well as adult tissue homeostasis. The protein
β-catenin is the central player in one major arm of the Wnt pathway
called the canonical Wnt pathway (12,13).
Characterization of this pathway has shown that the Wnt/β-catenin
signaling pathway is indispensible in processes as diverse as cell
fate, proliferation, differentiation, growth and cell survival
(14–16). Hyperactivation of β-catenin
signaling has been implicated as a driver of various types of
cancers, including liver cancer. Invasion and metastasis of
malignant tumors depend on angiogenesis. Furthermore, research has
found that the Wnt/β-catenin signaling pathway plays an important
role in angiogenesis. In vivo experiments have demonstrated
that a large number of vascular cells express multiple Wnt
proteins, such as Wnt-2, Wnt-5a and Wnt receptor FZD. In many
developmental and pathological conditions, stable intravascular
expression of β-catenin was found, which further confirmed the
regulatory effect of the Wnt/β-catenin signaling pathway on
angiogenesis (17). Recent studies
have shown that MMPs as tumor invasion and metastasis-promoting
factors are closely related to the Wnt/β-catenin signaling pathway
(18). It has been found that HCC
tumor angiogenesis, tumor progression and liver metastasis are
closely related to the high expression of MMP-2 in patients with a
poor prognosis. MMP-2 overexpression causes the degradation of type
IV collagen in the extracellular matrix (ECM) and basement membrane
(BM) and this may be the main cause of of cell invasion and
metastasis of HCC. Arii et al(19) found that MMP-9 gene expression
levels can be used as indicators of primary liver cancer
recurrence, invasion and metastasis. Vascular endothelial growth
factor (VEGF) is a highly specific endothelial mitogen which
promotes endothelial cell division, proliferation and induces the
occurrence of blood vessels during tumor occurrence and
development. Kamel et al(20) found that VEGF expression promotes
HCC recurrence, invasion and metastasis. Zhang et
al(21) reported on 805 bp
upstream of VEGF promoter locus TCF4 binding components, indicating
that the Wnt signaling pathway significantly upregulated the
expression of VEGF. COX-2 in addition to regulating the expression
of MMP, is also involved in tumor angiogenesis. APC mutations can
greatly enhance the activity of COX-2, indicating that
Wnt/β-catenin signaling also regulates the expression of COX-2 and
promotes tumor angiogenesis.
Caudatin is mainly isolated from the root of
Cynanchum bungei Decne, a traditional Chinese medicine and
health food, which has been used to nourish the blood and enhance
immunity in China and other Asian countries for a long time
(10). Recently, other researchers
have observed that caudatin induces cell growth arrest and
apoptosis in human hepatoma cells. However, the molecular
mechanisms are still unclear. In the present study, we used human
hepatoma cells as experimental material to confirm the antitumor
effect of caudatin, and to illustrate the underlying mechanisms of
its anticancer activity. We first examined the cell cycle and cell
apoptosis by flow cytometric assay, and found that caudatin induced
SMMC-7721 cell apoptosis and arrested the cell cycle in the G2
phase. To further investigate the mechanism of caudatin involved in
the regulation of cell proliferation and apoptosis, the protein
level of β-catenin was determined by western blot assay. We found
that caudatin treatment inhibited the expression of β-catenin and
GSK3β. Activation of Wnt signaling by binding of Wnt ligands to a
Frizzled receptor inhibits GSK3β-mediated phosphorylation of
β-catenin, resulting in an accumulation of hypophosphorylated
β-catenin in the cytosol (22).
Stabilized hypophosphorylated or dephosphorylated β-catenin
eventually translocates to the nucleus, leading to modulated
expression of a broad range of genes, such as cyclin D1 and Myc
(23,24). In the present study, we also
investigated expression levels of the downstream target genes
associated with the Wnt/β-catenin pathway. Cox-2,
mmp-2 and mmp-9 mRNA was inhibited by caudatin as a
result of its inhibitory effect on β-catenin and GSK3β.
Traditional Chinese medicine with its unique manner
of syndrome differentiation and ‘preventive treatment of disease’
has demonstrated efficiency in the control of tumor metastasis and
has received scientific attention and affirmation. Treatment of
liver cancer with Chinese medicine offers the advantages of
multi-component, multi-link, multi-target effects. A variety of
Chinese herbal medicinal ingredients has been proven to play roles
in liver cancer through a series of signal transduction pathways;
yet, the specific areas in which they interfere with signal
transduction remain unclear. Current research has mainly focused on
tumor cell proliferation, apoptosis and tumor angiogenesis, as the
signal transduction pathway is extremely complex. To elucidate the
specific mechanisms of the effects of Chinese medicine on liver
cancer, further in-depth study and discussion must be carried
out.
Acknowledgements
The present study was supported by the second part
of the Jiangsu Province Outstanding Young Chinese Medical Talents
Training Project, Jiangsu Province Administration of Traditional
Chinese Medicine (no. YX1214).
References
|
1
|
O’Brien K, Cokkinides V, Jemal A, et al:
Cancer statistics for Hispanics, 2003. CA Cancer J Clin.
53:208–226. 2003.
|
|
2
|
Bruix J and Sherman M: Management of
hepatocellular carcinoma: an update. Hepatology. 53:1020–1022.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bruix J, Raoul JL, Sherman M, et al:
Efficacy and safety of sorafenib in patients with advanced
hepatocellular carcinoma: subanalyses of a phase III trial. J
Hepatol. 57:821–829. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Llovet JM, Ricci S, Mazzaferro V, et al:
Sorafenib in advanced hepatocellular carcinoma. N Engl J Med.
359:378–390. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Su M, Wu X, Chung HY, Li Y and Ye W:
Antiproliferative activities of five Chinese medicinal herbs and
active compounds in Elephantopus scaber. Nat Prod Commun.
4:1025–1030. 2009.PubMed/NCBI
|
|
6
|
Chang C, Zhu Y, Tang X and Tao W: The
anti-proliferative effects of norcantharidin on human HepG2 cells
in cell culture. Mol Biol Rep. 38:163–169. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lee MK, Yeo H, Kim J, Markelonis GJ, Oh TH
and Kim YC: Cynandione A from Cynanchum wilfordii protects
cultured cortical neurons from toxicity induced by
H2O2, L-glutamate, and kainate. J Neurosci
Res. 59:259–264. 2000.
|
|
8
|
Lee MK, Yeo H, Kim J and Kim YC:
Protection of rat hepatocytes exposed to CCl4 in-vitro
by cynandione A, a biacetophenone from Cynanchum wilfordii.
J Pharm Pharmacol. 52:341–345. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Peng YR, Ding YF, Wei YJ, Shu B, Li YB and
Liu XD: Caudatin-2,6-dideoxy-3-O-methy-β-D-cymaropyranoside 1
induced apoptosis through caspase 3-dependent pathway in human
hepatoma cell line SMMC-7721. Phytother Res. 25:631–637. 2011.
|
|
10
|
Zhang JF, Li YB, Li CL and Jiang JQ:
Studies on chemical constituents in root tuber of Cynanchum
auriculatum. Zhongguo Zhong Yao Za Zhi. 31:814–816. 2006.(In
Chinese).
|
|
11
|
Pongracz JE and Stockley RA: Wnt
signalling in lung development and diseases. Respir Res. 7:152006.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
MacDonald BT, Tamai K and He X:
Wnt/β-catenin signaling: components, mechanisms, and diseases. Dev
Cell. 17:9–26. 2009.
|
|
13
|
Behari J: The Wnt/β-catenin signaling
pathway in liver biology and disease. Expert Rev Gastroenterol
Hepatol. 4:745–756. 2010.
|
|
14
|
El Wakil A and Lalli E: The
Wnt/beta-catenin pathway in adrenocortical development and cancer.
Mol Cell Endocrinol. 332:32–37. 2011.PubMed/NCBI
|
|
15
|
Monga SP: Role of Wnt/β-catenin signaling
in liver metabolism and cancer. Int J Biochem Cell Biol.
43:1021–1029. 2011.
|
|
16
|
Xiong F, Leonov S, Howard AC, et al:
Receptor for advanced glycation end products (RAGE) prevents
endothelial cell membrane resealing and regulates F-actin
remodeling in a β-catenin-dependent manner. J Biol Chem.
286:35061–35070. 2011.PubMed/NCBI
|
|
17
|
Goodwin AM and D’Amore PA: Wnt signaling
in the vasculature. Angiogenesis. 5:1–9. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mei JM, Borchert GL, Donald SP and Phang
JM: Matrix metalloproteinase(s) mediate(s) NO-induced dissociation
of β-catenin from membrane bound E-cadherin and formation of
nuclear beta-catenin/LEF-1 complex. Carcinogenesis. 23:2119–2122.
2002.PubMed/NCBI
|
|
19
|
Arii S, Mise M, Harada T, et al:
Overexpression of matrix metalloproteinase 9 gene in hepatocellular
carcinoma with invasive potential. Hepatology. 24:316–322. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kamel L, Nessim I, Abd-el-Hady A, Ghali A
and Ismail A: Assessment of the clinical significance of serum
vascular endothelial growth factor and matrix metalloproteinase-9
in patients with hepatocellular carcinoma. J Egypt Soc Parasitol.
35:875–890. 2005.
|
|
21
|
Zhang X, Gaspard JP and Chung DC:
Regulation of vascular endothelial growth factor by the Wnt and
K-ras pathways in colonic neoplasia. Cancer Res. 61:6050–6054.
2001.PubMed/NCBI
|
|
22
|
Baryawno N, Sveinbjornsson B, Eksborg S,
Chen CS, Kogner P and Johnsen JI: Small-molecule inhibitors of
phosphatidylinositol 3-kinase/Akt signaling inhibit Wnt/β-catenin
pathway cross-talk and suppress medulloblastoma growth. Cancer Res.
70:266–276. 2010.PubMed/NCBI
|
|
23
|
Tung JN, Chiang CC, Tsai YY, et al:
CyclinD1 protein expressed in pterygia is associated with β-catenin
protein localization. Mol Vis. 16:2733–2738. 2010.PubMed/NCBI
|
|
24
|
Zhang JY, Tao LY, Liang YJ, et al:
Secalonic acid D induced leukemia cell apoptosis and cell cycle
arrest of G(1) with involvement of GSK-3β/β-catenin/c-Myc pathway.
Cell Cycle. 8:2444–2450. 2009.PubMed/NCBI
|