Introduction
Gastric cancer is one of the most frequently
occurring types of cancer in China (1). Metastatic disease is one of the major
causes of mortality in cancer patients, and exploring the
mechanisms that are the acquisition of a metastatic phenotype may
offer new therapeutic strategies for metastatic gastric cancer
patients.
The ETS family of transcription factors is involved
in several physiological and pathological processes including tumor
progression (2). The ETS
transcription factors are divided into subfamilies based on the
sequence and location of the ETS domain. ETV1 (Ets variant gene 1;
also known as ER81), is a member of the PEA3 subfamily, which has
been found to promote metastatic progression in several types of
human cancer. In prostate cancer, ETV1 performs its oncogenic
effects through upregulating the matrix metalloproteinase-7 (MMP-7)
gene expression (3); ETV1 has also
been shown to cooperate with the androgen receptor (AR) to bind to
the prostate-specific antigen enhancer and enhance gene
transcription (4,5). In gastrointestinal stromal tumor
(GIST), ETV1 was shown to be highly expressed and to be activated
by KIT mutations (6), but this was
disputed by another study recently (7). Although previous studies demonstrated
that ETV1 expression was upregulated in gastric adenocarcinomas
(8,9), the underlying mechanisms of
ETV1-induced metastatic progression in gastric cancer remain
elusive.
Epithelial-mesenchymal transition (EMT) is a process
characterized by loss of cell-cell adhesion, repression of
E-cadherin expression and gain of cell motility (10). In cancer cells, the EMT process may
promote their metastatic potential. Transcription factors,
including Snail, Slug and TWIST, have been demonstrated to be
master regulators of EMT (11).
These proteins are transcriptional repressors of E-cadherin and
their expression induces EMT, thereby increasing motility and
invasiveness of cancer cells.
The role of Snail in EMT and metastatic progression
of cancer cells has been extensively studied (12). In the present study, we present data
showing that ETV1 promotes Snail expression to induce EMT-like
metastatic progression in breast cancer.
Materials and methods
Cell culture and transfection
Human normal gastric epithelial cell line GES-1,
gastric cancer cell lines MKN-45, MGC-803 and SGC-7901, HEK 293T
cells were maintained and cultured in Dulbecco’s Modified Eagle’s
Medium (DMEM; Invitrogen) supplemented with 10% fetal bovine serum
(FBS; Invitrogen). All cell lines were incubated at 37°C in
humidified air with 5% CO2.
Cells were transfected with Flag-tagged ETV1 using
Lipofectamine™ 2000 reagent (Invitrogen), according to the
manufacturer’s protocol. siRNA oligonucleotides against human ETV1
(ON-TARGETplus SMARTpool) and non-targeting siRNAs control
oligonucleotides were obtained from Dharmacon using DharmaFECT1
siRNA transfection reagent (Dharmacon).
Western blot analysis
Whole cell extracts were prepared using Laemmli
Buffer. Samples were run on a 10% sodium dodecyl sulfate
(SDS)-polyacrylamide gel and transferred to a nitrocellulose
membrane. Membranes were blocked in 5% milk solution [Tris buffered
saline (TBS)-0.1% Tween]for 1 h at room temperature and incubated
with indicated primary antibody in 5% milk solution overnight at
4°C. The membranes were washed three times for 10 min in TBS-0.1%
Tween at room temperature and incubated for 1 h with the
corresponding horseradish peroxidase (HRP)-conjugated secondary
antibody. Proteins were detected by the enhanced chemiluminescence
system (Amersham Pharmacia Biotech) as described by the
manufacturer’s instructions. The primary antibodies used for
western blot analysis were: anti-ETV1 (ab81086; Abcam), anti-Snail
(sc-10432), anti-GAPDH (sc-25778; Santa Cruz Biotechnology),
anti-Vimentin (550513), anti-N-cadherin (610921), anti-E-cadherin
(610182), anti-β-catenin (610153; BD Transduction
Laboratories).
mRNA isolation and quantitative PCR
(qPCR)
mRNA was isolated using TRIzol (Invitrogen) and cDNA
was prepared using transcriptor first-strand cDNA synthesis kit
(Roche) according to the manufacturer’s protocol. qRT-PCR was
carried out using SYBR-Green (Roche) on a MasterCycler RealPlex4
instrument. Gene expression was normalized to Gapdh. The
upstream and downstream primers used for ETV1 gene were:
5′-TACCCCATGGACCACAGATT-3′ and 5′-CACTGGGTCGTGGTACTCCT-3′.
Transwell invasion assay
Transwell invasion experiments were performed with
24-well Matrigel-coated chambers (8 μm pore size) from BD
Biosciences. Briefly, cells were allowed to grow to subconfluency
(~80%) and were serum-starved for 24 h. After detachment with
trypsin, cells were washed with phosphate-buffered saline,
resuspended in serum-free medium and 5×104 cells were
added to the upper chamber. Complete medium was added to the bottom
wells of the chambers. After 12 h, the cells that had not migrated
were removed from the upper face of the filters using cotton swabs,
and the cells that had migrated were fixed and stained by crystal
violet solution.
Luciferase assay
HEK 293T cells were co-transfected with the
indicated plasmids. TK-Renilla expression plasmid was used
as an internal control. Luciferase activity was measured after 24 h
using a dual luciferase assay kit according to the manufacturer’s
protocol (Promega). Statistical analysis was performed using
GraphPad Prism 4.0.
Chromatin immunoprecipitation (ChIP)
The ChIP kit was obtained from Millipore and ChIP
was performed according to the manufacturer’s instructions.
Immunoprecipitation was carried out using 2 mg of ETV1 antibody, 2
mg normal rabbit IgG (Sigma-Aldrich). PCR was carried out using the
following primers: forward, 5′-CCAGTGATGTGCGTTTCCCT-3′ and reverse,
5′-AAGCGAGGCCTCTGCGAGGT-3′.
Immunohistochemistry
Expression of ETV1 and Snail was analyzed using
human gastric cancer tissues from 20 patients in grade I and 20
patients in grade III. Use of the tissue samples was approved by
the Harbin Medical University Institutional Review Board. Standard
immunohistochemical procedures were carried out using an anti-ETV1
or anti-Snail antibody. The staining results were scored by two
investigators blinded to the clinical data. As negative controls,
the primary antibodies were omitted and replaced with a related
strain of IgG used as a negative control.
Statistical analysis
The Student’s t-test was used for two-group
comparisons. Comparisons between three or more groups were analyzed
by one-way ANOVA followed by the Duncan’s test in SPSS 15.0 (SPSS
Inc.). The probability of P<0.05 was considered to indicate a
statistically significant result.
Results
Expression of ETV1 in normal gastric
epithelial and gastric cancer cell lines
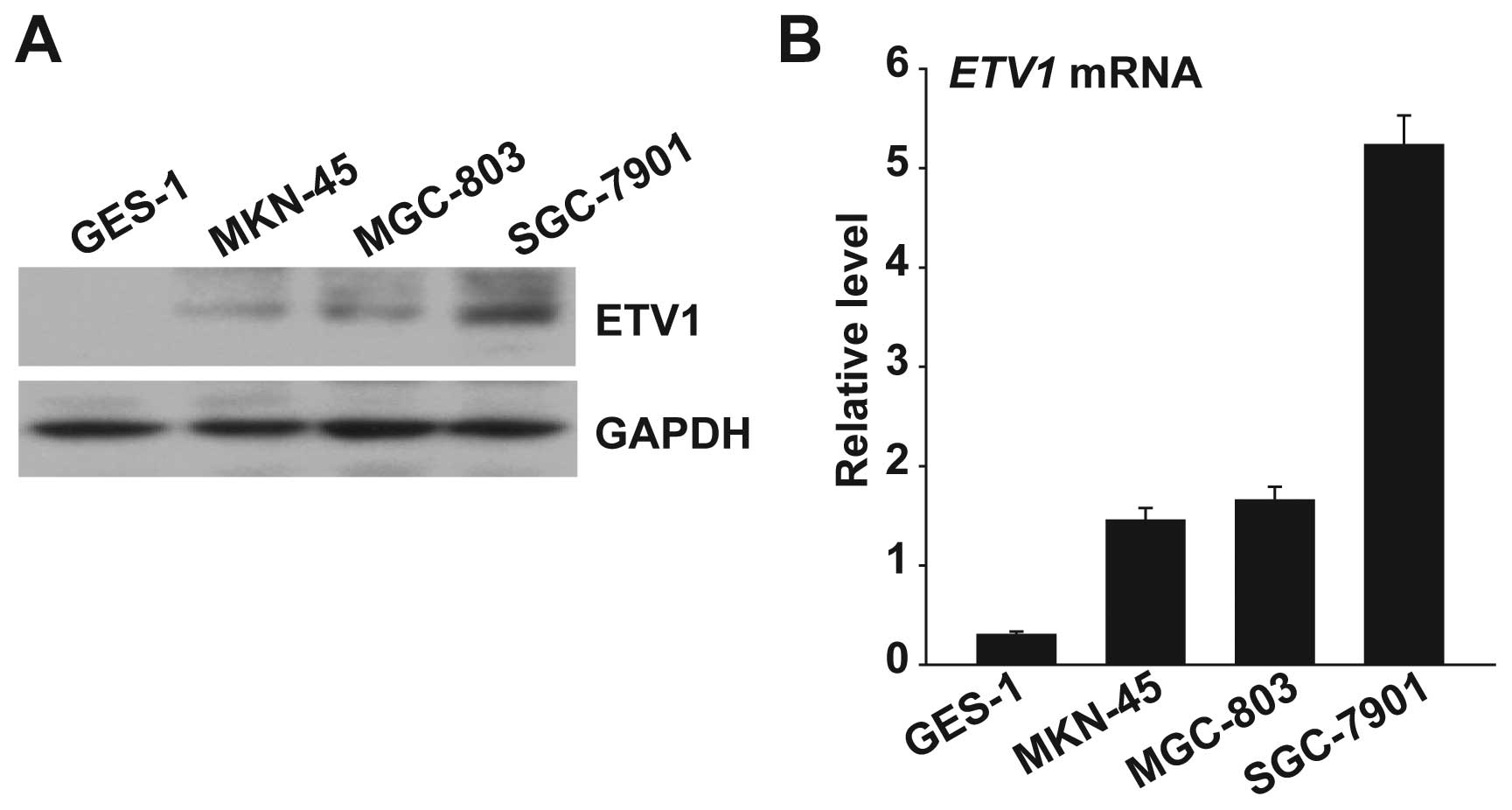
First, we determined the ETV1 expression level using
qPCR and western blot analysis in human normal gastric epithelial
cell line GES-1 and the gastric cancer cell lines (MKN-45, MGC-803
and SGC-7901). The level of ETV1 expression in the three gastric
cancer cell lines was significantly higher than in the normal
gastric epithelial cell line GES-1 at both the protein and the mRNA
level (Fig. 1). The highest levels
of ETV1 expression were observed in the SGC-7901 gastric cancer
cell line, which is a high metastatic potential cell.
Overexpression of ETV1 in normal gastric
epithelial cells results in EMT and increased invasiveness
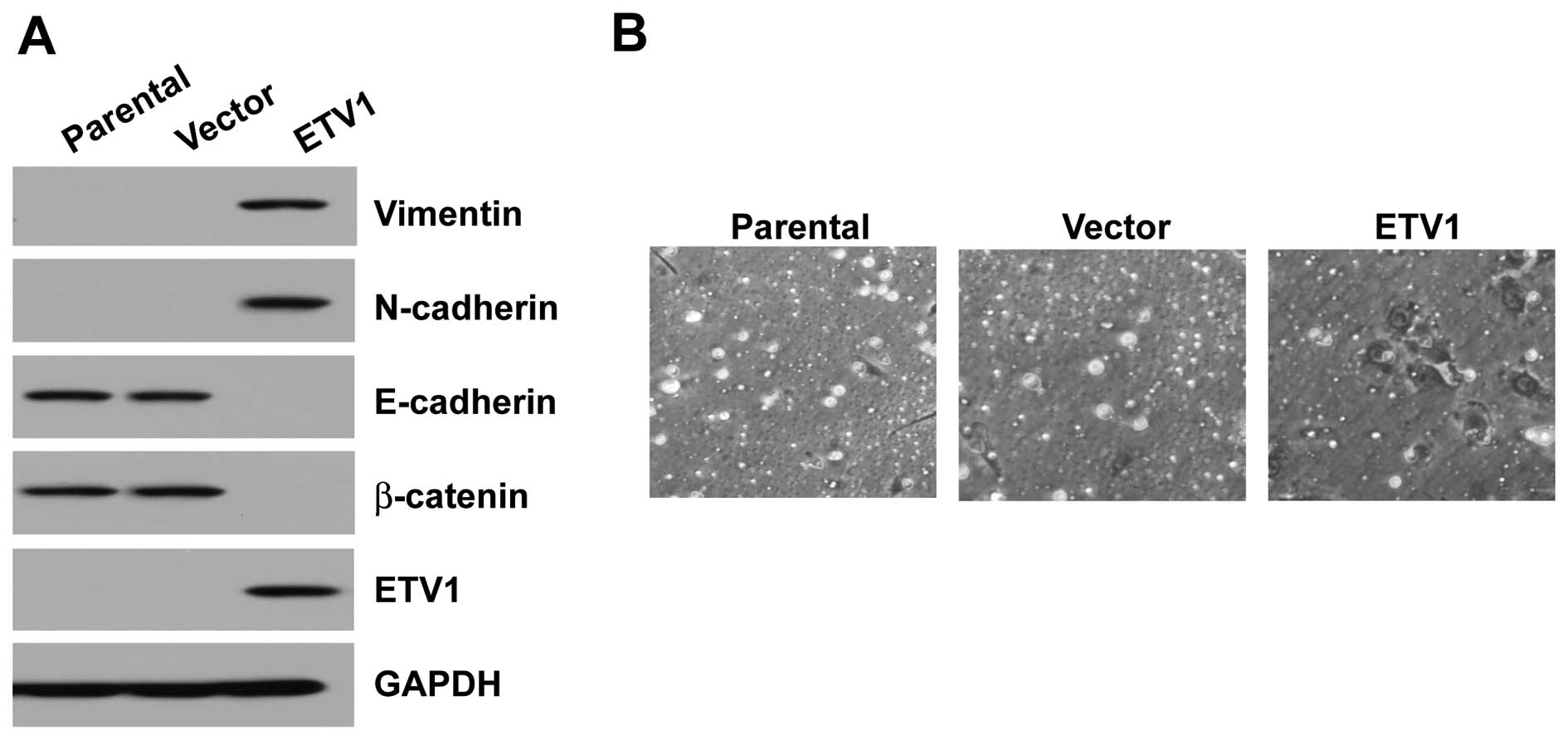
GES-1 cells overexpressing ETV1 expressed higher
levels of mesenchymal markers, such as vimentin and N-cadherin, and
lower levels of epithelial markers, such as E-cadherin and
β-catenin, compared with their vector control (Fig. 2A). To investigate whether the
ETV1-induced EMT-like phenotype could be translated into enhanced
metastatic ability of the GES-1 cells, the invasion of ETV1 cells
was tested. ETV1-overexpressing GES-1 cells displayed an increase
in motility through the Matrigel in Transwell invasion assays,
compared with the vector control cells (Fig. 2B).
Knockdown of ETV1 results in decreased
aggressiveness of the invasive gastric cancer cells
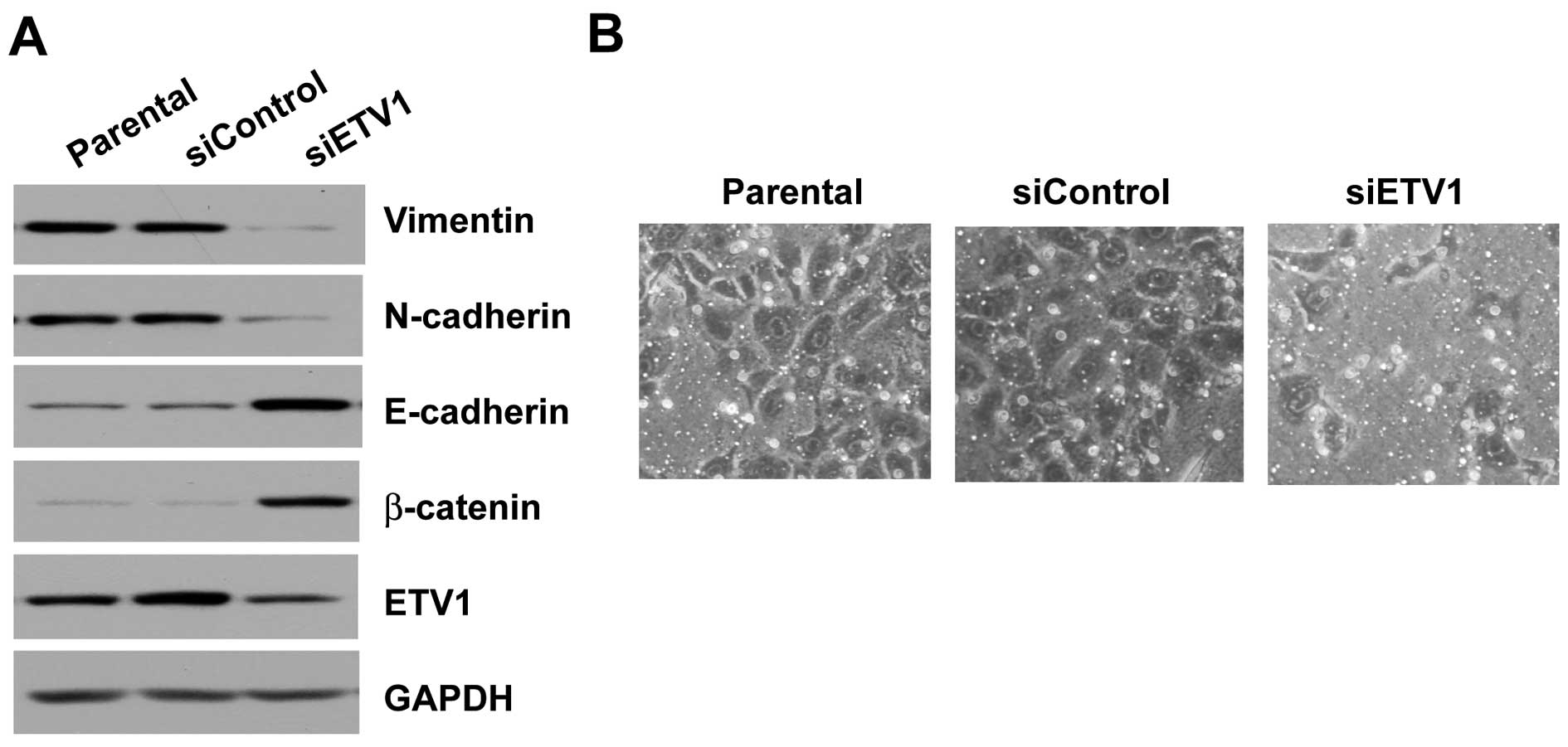
ETV1 was knocked down by siRNA in the high
metastatic potential cell line SGC-7901, which expresses high
levels of ETV1. Knockdown of ETV1 in the cell line resulted in a
significant decrease at the protein level. Protein expression of
the mesenchymal marker vimentin and N-cadherin was also decreased.
The level of E-cadherin and β-catenin, both epithelial markers, was
induced (Fig. 3A). Furthermore,
knockdown of ETV1 in SGC-7901 cells resulted in decreased ability
of the cells to invade through the Matrigel in the invasion assay
(Fig. 3B).
Overexpression of ETV1 promotes Snail
expression but knockdown of ETV1 inhibits it
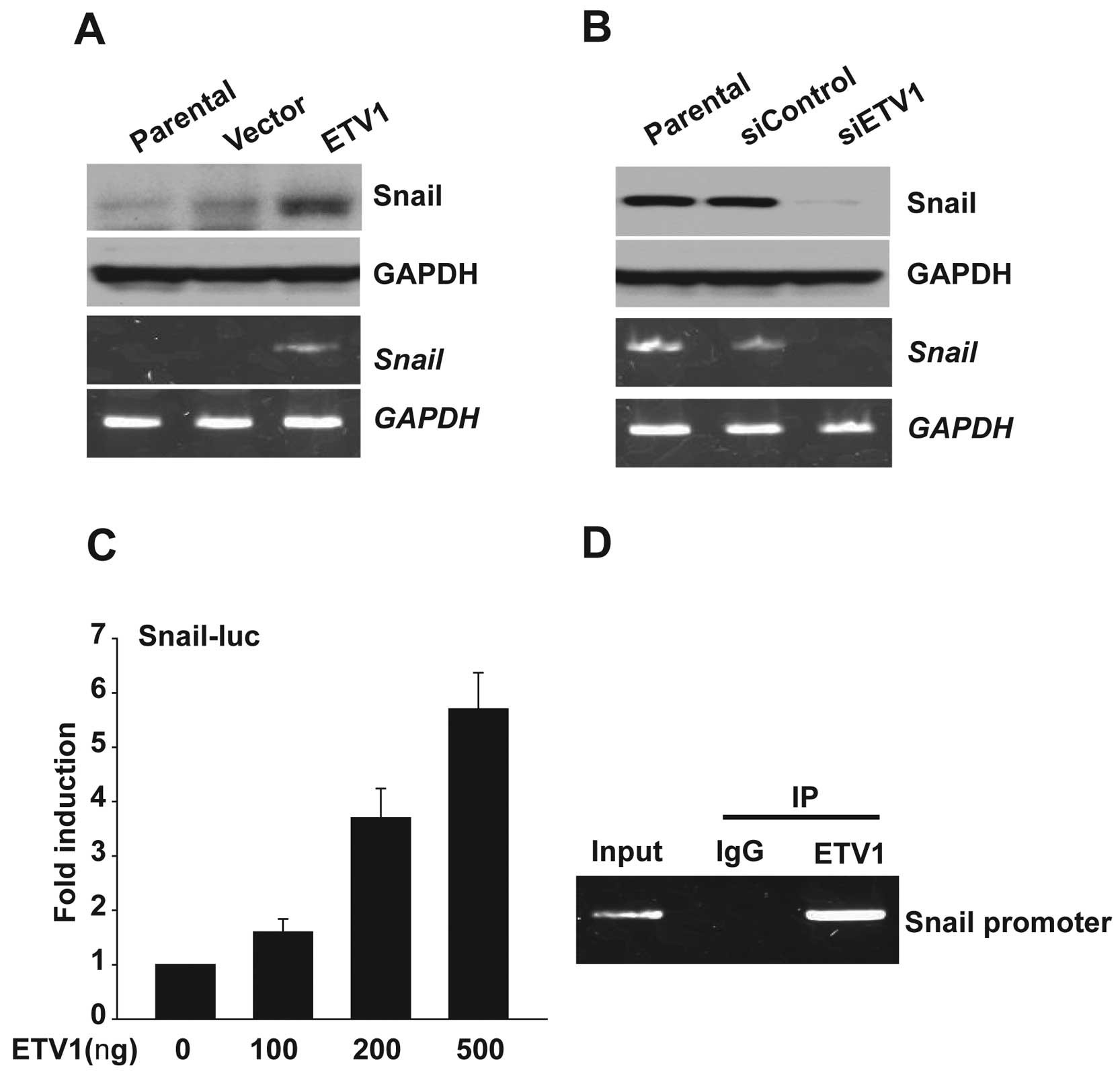
To elucidate the mechanism of ETV1 that leads to EMT
and invasiveness, we hypothesized that ETV1 regulates the
expression of Snail. Indeed, ectopic ETV1 expression greatly
increased Snail at both the mRNA and the protein levels.
Conversely, knockdown of ETV1 reduced the Snail expression
(Fig. 4A and B). Next, we assessed
whether ETV1 promotes the activity of the Snail gene
promoter. To test this possibility, the Snail luciferase reporter
construct (Snail-Luc) transiently transfected into HEK 293T
cells with and without ETV1 expression plasmids. Luciferase
reporter activity increased in a dose-dependent manner in cells
with ectopic ETV1 expression compared with empty vector (Fig. 4C). This result suggested that
ectopic ETV1 expression promotes the promoter activity of
Snail. To determine if ETV1 interacted with the endogenous
Snail promoter, we performed ChIP assays. Primers flanking
the predicted ETV1 binding site of the Snail promoter were
used to amplify chromatin fragments enriched by ETV1 binding to
this region. The PCR products were amplified from DNA fragments
immunoprecipitated with an anti-ETV1 antibody but not from DNA
fragments precipitated with an IgG control antibody. As shown as
Fig. 4D, ETV1 bound to the
Snail promoter.
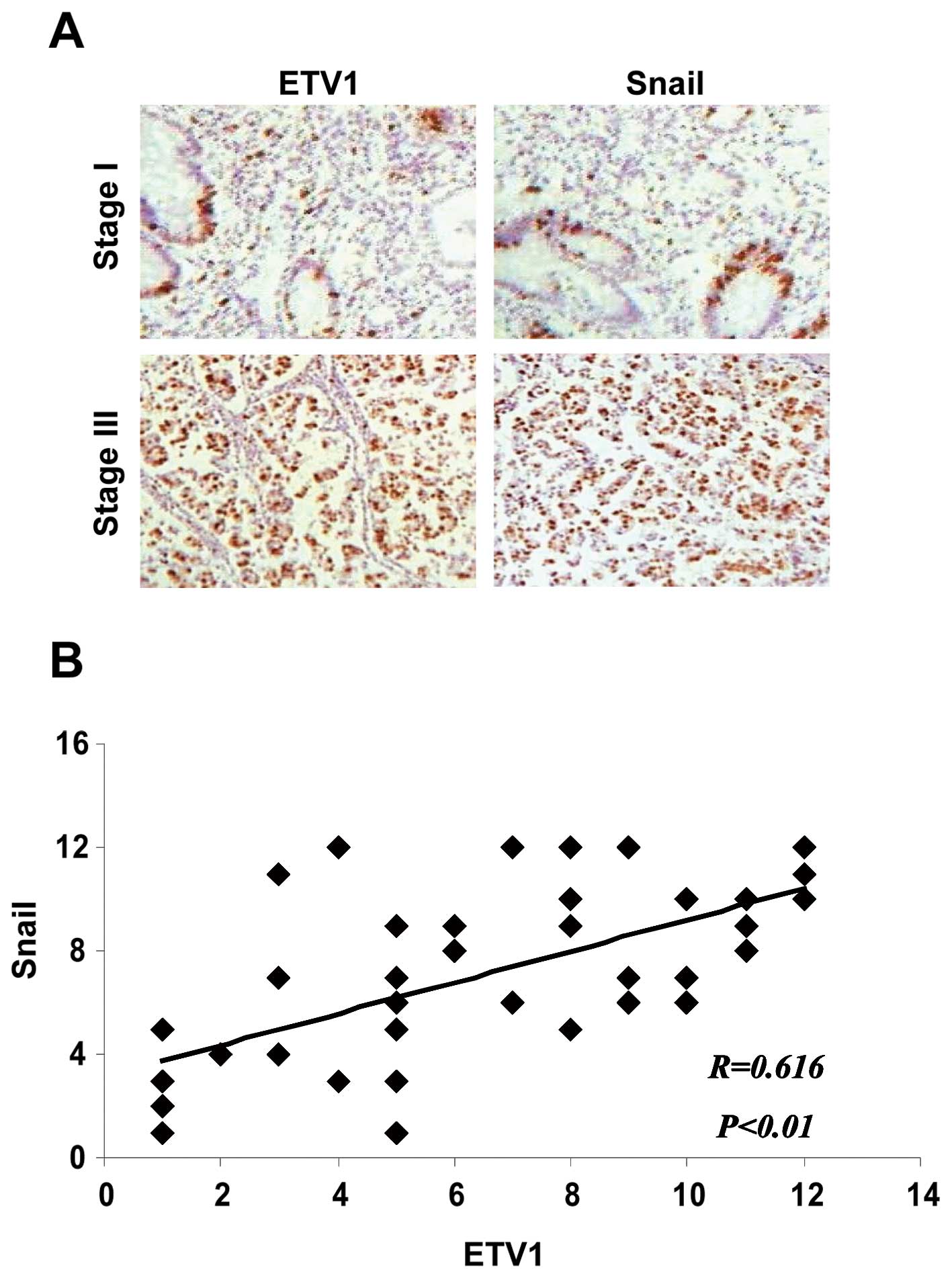
Correlative expression of ETV1 with high
levels of Snail in gastric adenocarcinoma specimens
To examine whether an increase in ETV1 expression
was associated with an increase in Snail level in human gastric
tumor samples, we performed immunohistochemistry to assess the
expression of ETV1 and Snail at the protein levels from 40 gastric
adenocarcinomas. We found that the protein levels of both molecules
were significantly correlated (Fig.
5).
Discussion
ETV1 belongs to the PEA3 subfamily, which is
associated with a variety of cancers including colon, breast,
prostate and gastric cancer (6,13–15).
In the presents study, we showed that ETV1 transcriptionally
activates Snail, thereby promoting EMT and invasive progression
in vitro. In human specimens, we further confirmed that the
expression levels and nuclear localization of ETV1, as well as that
of Snail, are potential prognostic markers for gastric cancer
patients.
In the present study, we first examined the
expression of ETV1 in normal gastric epithelial and gastric cancer
cell lines at the mRNA level by real-time PCR and at the protein
level by western blotting. EMT is a process that enables cancer
cells to lose their cell-cell and cell-matrix contacts to gain
migratory properties through transcriptional reprogramming
(16,17). To demonstrate the role of ETV1 in
EMT, we silenced the expression of ETV1 using siRNA in SGC-7901
cells with highly invasive potential and ETV1 high expression.
Moreover, we also overexpressed ETV1 in normal gastric epithelial
cell line GES-1 without invasion and ETV1 expression. We found that
the variation of ETV1 expression significantly correlates with
several putative EMT marker expressions. In addition, assessment of
the invasive potential, following transfection with ETV1-siRNA,
indicated that the rate of cell invasion was markedly reduced
compared to those in siControl and mock control, suggesting that
ETV1 contributes to the invasive potential of gastric cancer
cells.
Although ETV1 overexpression is thought to
contribute to gastric carcinogenesis, it has remained unknown by
which molecular mechanism the transcription factor achieves its
deleterious effects. It is well-known that Snail is required for
EMT-mediated metastatic development and its upregulation is
associated with recurrence (18).
The present study provides a mechanistic explanation in the case of
ETV1, i.e., through inducing transcription as exemplified for the
Snail gene.
Immunohistochemical expression of ETV1 was
associated with an increase in the tumor grade and tumor stage
suggesting that increasing levels of ETV1 in primary tumors are
associated with a more differentiated phenotype and initial stages
of carcinogenesis. ETV1 expression was significantly correlated
with Snail expression, further supporting the hypothesis that ETV1
plays an important role in the induction of Snail expression.
In conclusion, the present study is the first to
show that ETV1 may induce EMT to acquire invasion in gastric cancer
cells. The function of ETV1 as an oncogene may be associated with
several important molecules involved in the invasion of cancer
cells. These results further suggest that ETV1 may serve as a
potential target for the development of therapies for gastric
cancer.
Acknowledgements
The present study was supported by the Foundation of
Educational Commission of Heilongjiang Province, China (11541224)
to Z.L.
References
|
1
|
Zhu X and Li J: Gastric carcinoma in
China: Current status and future perspectives (Review). Oncol Lett.
1:407–412. 2010.PubMed/NCBI
|
|
2
|
Llauradó M, Abal M, Castellví J, et al:
ETV5 transcription factor is overexpressed in ovarian cancer and
regulates cell adhesion in ovarian cancer cells. Int J Cancer.
130:1532–1543. 2012.PubMed/NCBI
|
|
3
|
Shin S, Oh S, An S and Janknecht R: ETS
variant 1 regulates matrix metalloproteinase-7 transcription in
LNCaP prostate cancer cells. Oncol Rep. 29:306–314. 2013.PubMed/NCBI
|
|
4
|
Shin S, Kim TD, Jin F, et al: Induction of
prostatic intraepithelial neoplasia and modulation of androgen
receptor by ETS variant 1/ETS-related protein 81. Cancer Res.
69:8102–8110. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cai C, Hsieh CL, Omwancha J, et al: ETV1
is a novel androgen receptor-regulated gene that mediates prostate
cancer cell invasion. Mol Endocrinol. 21:1835–1846. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chi P, Chen Y, Zhang L, et al: ETV1 is a
lineage survival factor that cooperates with KIT in
gastrointestinal stromal tumours. Nature. 467:849–853. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Birner P, Beer A, Vinatzer U, et al:
MAPKAP kinase 2 overexpression influences prognosis in
gastrointestinal stromal tumors and associates with copy number
variations on chromosome 1 and expression of p38 MAP kinase and
ETV1. Clin Cancer Res. 18:1879–1887. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Keld R, Guo B, Downey P, et al:
PEA3/ETV4-related transcription factors coupled with active ERK
signalling are associated with poor prognosis in gastric
adenocarcinoma. Br J Cancer. 105:124–130. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yamamoto H, Horiuchi S, Adachi Y, et al:
Expression of ets-related transcriptional factor E1AF is associated
with tumor progression and over-expression of matrilysin in human
gastric cancer. Carcinogenesis. 25:325–332. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
De Craene B and Berx G: Regulatory
networks defining EMT during cancer initiation and progression. Nat
Rev Cancer. 13:97–110. 2013.PubMed/NCBI
|
|
11
|
Heldin CH, Vanlandewijck M and Moustakas
A: Regulation of EMT by TGFβ in cancer. FEBS Lett. 586:1959–1970.
2012.
|
|
12
|
Sánchez-Tilló E, Liu Y, de Barrios O, et
al: EMT-activating transcription factors in cancer: beyond EMT and
tumor invasiveness. Cell Mol Life Sci. 69:3429–3456.
2012.PubMed/NCBI
|
|
13
|
Horiuchi S, Yamamoto H, Min Y, Adachi Y,
Itoh F and Imai K: Association of ets-related transcriptional
factor E1AF expression with tumour progression and overexpression
of MMP-1 and matrilysin in human colorectal cancer. J Pathol.
200:568–576. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shin S, Bosc DG, Ingle JN, Spelsberg TC
and Janknecht R: Rcl is a novel ETV1/ER81 target gene upregulated
in breast tumors. J Cell Biochem. 105:866–874. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Baena E, Shao Z, Linn DE, et al: ETV1
directs androgen metabolism and confers aggressive prostate cancer
in targeted mice and patients. Genes Dev. 27:683–698. 2013.
View Article : Google Scholar
|
|
16
|
Thiery JP: Epithelial-mesenchymal
transitions in tumour progression. Nat Rev Cancer. 2:442–454. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Evdokimova V, Tognon C, Ng T and Sorensen
PH: Reduced proliferation and enhanced migration: two sides of the
same coin? molecular mechanisms of metastatic progression by YB-1.
Cell Cycle. 8:2901–2906. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Naber HP, Drabsch Y, Snaar-Jagalska BE,
Ten Dijke P and van Laar T: Snail and Slug, key regulators of
TGF-β-induced EMT, are sufficient for the induction of single-cell
invasion. Biochem Biophys Res Commun. 435:58–63. 2013.
|