Introduction
α-Tomatine is a natural steroidal glycoalkaloid,
occurring mainly in stems, leaves and roots of tomatoes (Solanum
lycopersicon L.) and other Solanum species. Its content
is high in immature green tomatoes and decreases during ripening.
α-Tomatine is composed of the 6-ring steroidal aglycone tomatidine
by which a tetrasaccharide moiety (containing xylose, galactose and
2 glucose units) is bound to the 3-OH group of the aglycone. Its
partial hydrolysis leads to the loss of different sugar parts of
α-tomatine and to the formation of β1-(β2-)
tomatine (containing a trisaccharide moiety), γ-tomatine (with a
disaccharide) and δ-tomatine (with a monosaccharide) (1,2).
α-Tomatine is a biologically active compound that
possesses numerous health-related properties. It exhibits
antiviral, antibiotic and anti-inflammatory activity (3–5),
stimulates antigen-specific humoral and cellular immune response
(6), inhibits acetylcholinesterase
activity (7) and has cardiotonic
effects (8). Its other effects are
connected to its ability to form an insoluble complex with
cholesterol in a 1:1 molar ratio (2,9). Since
such complex with dietary cholesterol is not able to pass through
the intestinal wall, the p.o. administration of α-tomatine
decreases the plasma cholesterol level (10). Moreover, α-tomatine disrupts
cholesterol containing mammalian biomembranes which could lead to
various symptoms of intoxication (gastrointestinal disturbances,
haemolysis) (11).
In recent years, the anticancer effect of α-tomatine
and its mechanism of action have been studied. In tumor-bearing
mice, α-tomatine inhibited tumor growth at doses of approximately 1
mg/kg (12–14). In vitro, α-tomatine inhibits
the growth of human colon HT29 and liver HepG2 cancer cell lines
more than classical anticancer agents (doxorubicin, camptothecin)
(15). Furthermore, it is cytotoxic
to different human lung cancer cells (A549, NCI-H460) (16,17),
human prostatic adenocarcinoma PC3 cells (18), mouse EL4 lymphoma cells (19), several leukemia cell lines (14,20)
and breast adenocarcinoma MCF-7 cells (21). Previous studies demonstrated the
caspase-independent cell death in α-tomatine-treated human leukemic
cell lines (14,20). In the PC-3 cell line, α-tomatine
induced apoptosis (18).
Furthermore, it inhibited the migration and invasion of cancer
cells by inactivating FAK/PI3K/Akt and Erk signaling pathways with
a decrease in the binding activity of NF-κB (16,17,21).
In the present study, we examined the dose- and
time-dependent inhibition of the growth of the human breast
adenocarcinoma MCF-7 cell line after α-tomatine treatment. Due to
conflicting reports in the literature regarding the induction of
apoptosis by α-tomatine, we also focused on the mechanism of action
and DNA damage in MCF-7 cells. The stability of α-tomatine in
culture medium and its biotransformation were also studied.
Materials and methods
Cell culture and cytostatic
treatment
Human breast adenocarcinoma cell line MCF-7 was
obtained from the American Type Culture Collection (ATCC; Manassas,
VA, USA) and maintained in Dulbecco’s Modified Eagle’s Medium
(Sigma-Aldrich, St. Louis, MO, USA) supplemented with a 10% fetal
calf serum, L-glutamine, penicillin, streptomycin and non-essential
amino acids. Cell cultures were maintained at 37°C in a humidified
atmosphere of 5% CO2 and the culture medium was renewed
every 2–3 days. Cells were detached by incubation with trypsin.
Cell lines in a maximal range of up to 20 passages were used for
this study. Appropriate amounts of stock solution of α-tomatine
(Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA) dissolved in
DMSO were added to the culture medium to achieve the final
concentration of 1–9 μM. The control group was treated with DMSO at
the concentration of 0.6% in every assay. For irradiation
treatment, the cells were irradiated in culture flasks at room
temperature using a 60Co γ-ray source (Chisotron,
Chirana, Ostrava, Czech Republic) at a distance of 1 m from the
source, at the photon dose rate of 1 Gy/min.
WST-1
For assessing the cytotoxic effect of α-tomatine on
cell viability, WST-1 (Roche, Basel, Switzerland) reagent was used.
MCF-7 cells were seeded in 96-well microtiter plates at a
concentration of 5×103 cells/well in 200 μl of culture
medium. The cells were allowed to settle overnight at 37°C and in
5% CO2. Next, cultures were exposed to different
concentrations (1, 3, 6, 9 μM) of α-tomatine for differing time
intervals. At the end of each interval, cells were rinsed with PBS
and 100 μl of WST-1 was added. Absorbance was measured after 2 h of
incubation with WST-1 at 450 nm and a reference wavelength of 650
nm in a Tecan SpectraFluor Plus spectrometer (Tecan Austria GmbH,
Grödig, Austria).
Time lapse videomicroscopy
MCF-7 cells were seeded into plastic tissue-culture
dishes with glass bottoms and left for 24 h in an incubator with 5%
CO2 at 37°C. The next day, the growth medium was
replaced with a medium containing different concentrations of
α-tomatine (3 and 9 μM). The tissue-culture dishes were transferred
into a time-lapse imaging system BioStation IM (Nikon, Prague,
Czech Republic) combining an incubator, a motorized microscope and
a cooled CCD camera. Recording was carried out in a multipoint and
multichannel manner employing various time-lapse modes and upon
small as well as high magnifications to allow global as well as
detailed view of changes in the behavior of the treated cell
populations. Recorded sequences were subsequently
semi-automatically analyzed with the software NIS-Elements AR 3.20
(Nikon).
xCELLigence
Growth of MCF-7 cells was continuously monitored for
at least 90 h using the xCELLigence RTCA MP instrument (Roche). The
background impedance signal was measured with 50 μl of cell culture
medium/well. The final volume in a single well was adjusted to 100
μl of cell culture medium by adding an additional 50 μl of medium
containing MCF-7 cells. The impedance was recorded in 15 min
intervals. Twenty-four hours after seeding, α-tomatine was added to
the culture. All incubations were performed at a volume of 150 μl.
α-Tomatine was analyzed at 4 different concentrations (1, 3, 6 and
9 μM), each one with 4 replicates. The impedance signal was
analyzed by normalizing data of each single well to the first
measurement after starting the treatment: CI(normalized)
= CItime x/CInorm time (termed here
‘normalized cell index’). This normalized cell index was used for
representing the results graphically and exported for further
processing using Microsoft Office Excel (Microsoft Inc., Redmond,
WA, USA).
ATP
ATP content in MCF-7 cells exposed to α-tomatine at
4 different concentrations (1, 3, 6 and 9 μM) was measured by an
ATP bioluminescent assay kit (Sigma-Aldrich). ATP content was
calculated from a standard curve derived from known concentrations
of ATP and was expressed as a percentage of the control.
LDH
LDH activity was determined at individual time
points in the medium of α-tomatine exposed MCF-7 cells. An aliquot
(0.1 ml) of each sample was added to a cuvette with 0.2 ml of 2.5
mg/ml NADH solution and 0.2 ml of sodium pyruvate solution (1
mg/ml). The total LDH activity was determined after sonication of
cells. The enzyme activity was determined spectrophotometrically
and the rate of enzyme leakage/min was expressed as percentage of
the total LDH activity compared to untreated control cells.
Analysis of DNA damage (comet assay)
This method is used to assess the single-strand
breaks (SSBs) and double-strand breaks (DSBs) present in the DNA.
The alkaline and neutral version of the comet assay was used as
previously described (22,23). Cells were embedded in 1% agarose
(Sigma-Aldrich) on microscope slides and lysed in 10 mM of
Tris-buffered 2.5 M NaCl (pH 10.0; Penta, Prague, Czech Republic)
with 1% Triton X-100 (Merck KGaA, Darmstadt, Germany), and 100 mM
EDTA (Penta) at 4°C for 1 h. The alkaline comet assay (NaOH, EDTA)
was carried out at 40 V, 300 mA, for 30 min at 4°C after 40 min of
alkaline unwinding. The neutral comet assay (90 mM Tris, 90 mM
boric acid, 2 mM EDTA, pH 8.0) was performed at 29 V, 6 mA, for 40
min, at 4°C.
Comets were scored by the Lucia 6.20 image analysis
comet module (Laboratory Imaging, Prague, Czech Republic) after
staining of cells with ethidium bromide (Sigma-Aldrich). The
percentage of DNA in the comet tail was assessed in at least 50
cells/slide and the median DNA damage score for each gel was
calculated. Generally, 4–6 gels for each treatment group were
scored, and the means ± SD (standard deviation) of medians of these
gels are presented.
Western blot assay
The cells treated with α-tomatine were harvested for
the preparation of whole-cell lysates (Cell Lysis Buffer; Cell
Signaling Technology, Inc., Boston, MA, USA) and the protein
content was quantified using BCA assay (Sigma-Aldrich). Lysates
with an equal amount of protein (10 μg) were loaded on 12% SDS-PAGE
gels. After electrophoresis, the separated proteins were
transferred to a PVDF membrane (Bio-Rad, Hercules, CA, USA). The
membranes were blocked in Tris-buffered saline containing 0.05%
Tween-20 and 10% non-fat dry milk and incubated with primary
antibody (p53; Exbio, Prague, Czech Republic; p53_serine15;
Calbiochem-Merck Co., USA; Chk1, Chk1_serine345; Chk2,
Chk2_threonine68; Cell Signaling Technology; β-actin,
p21WAF1/Cip1; Sigma-Aldrich) at 4°C overnight. Then, the
membranes were incubated with polyclonal anti-mouse or anti-rabbit
secondary antibody (DakoCytomation, Glostrup, Denmark). For band
detection, a chemiluminescence detection kit (Roche) was used.
Caspase activity
In order to detect caspase-8 and -9 activity,
Caspase-Glo® Assays (Promega Corporations, Madison, WI,
USA) were used according to the manufacturer’s instructions.
Briefly, the cells (1×104) treated with the different
concentrations of α-tomatine were seeded per well using a 96-well
plate format (Sigma-Aldrich) and incubated with 50 μl of
Caspase-Glo® Assays reagent for 30 min before the
luminescence was measured using a Tecan Infinite M200 spectrometer
(Tecan Group, Männedorf, Switzerland).
Flow cytometry
After 24 and 72 h of incubation with α-tomatine,
cells were harvested, washed twice with ice-cold PBS buffer, fixed
and permeabilized with 70% ethanol. For detecting
low-molecular-weight fragments of DNA, the cells were incubated for
5 min at room temperature in a buffer (192 ml 0.2 M
Na2HPO4 + 8 ml 0.1 M citric acid, pH 7.8) and
then stained with propidium iodide in Vindelov’s solution for 60
min at 37°C (all reagents from Sigma-Aldrich). The DNA content was
determined by flow cytometer Dako CyAn (Beckman Coulter, Brea, CA,
USA) with an excitation wave length of 488 nm; the total emission
above 560 nm was recorded. List mode data were analyzed using
Multicycle AV software (Phoenix Flow Systems, San Diego, CA, USA)
and the percentage of cycling cells was determined.
Transmission electron microscopy
MCF-7 cells were treated with 6 μM of α-tomatine for
4 h. The cells were fixed in 2% paraformaldehyde and 1%
glutaraldehyde in phosphate buffer for 4 h at 4°C. The cells were
then post-fixed in 2% OsO4 in phosphate buffer for 2 h
at room temperature. The dehydration was carried out in graded
ethanol and propylene glycol and embedding in Epon-Durcupan
(Sigma-Aldrich). Ultra-thin sections (60 nm) were prepared and
placed on grids, stained with 2% uranyl acetate solution and 0.2%
lead citrate in 0.1 M NaOH, and examined in a Tesla BS-500
transmission electron microscope (Tesla, Brno, Czech Republic). The
calibration of magnification was carried out using the calibrator
02902-AB (SPI, West Chester, PA, USA).
Liquid chromatography, mass
spectrometry
Analysis of α-tomatine
Separation was performed on a 1200 series liquid
chromatography system (Agilent Technologies, Palo Alto, CA, USA)
consisting of degasser, quaternary pump, autosampler and
thermostated column compartment. The HPLC column used was Accucore
C18 (100×2.1 mm, 2.6 μm) (Thermo Scientific, San Jose, CA, USA)
held at 40°C. The mobile phase flowing at a rate of 0.6 ml/min
consisted of ammonium formate (5 mM, pH=3.8) (solvent A) and
acetonitrile (solvent B). The gradient elution program was: from 20
to 27% (v/v) of solvent B in 4.5 min, for 1 min at 50% solvent B
and equilibration at 20% (v/v) of solvent B for 2 min.
Column output was coupled with a 3D ion trap LCQ
Fleet mass spectrometer with electrospray interface (Thermo
Scientific). All mass scans were operated in positive SIM mode at
m/z 1033.5–1035.5 for α-tomatine and 883.5–885.5 for solasonine
(IS) (Research Plus Inc., Bayonne, NJ, USA) with the following
optimized settings for the ion source: spray voltage 4 kV, sheath
gas 40 AU, auxiliary gas 10 AU, capillary voltage 20 V, capillary
temperature 260°C, tube lens voltage 92 V.
Metabolic screening
For metabolite screening, the same chromatographic
system and mass spectrometer was used as in the case of α-tomatine
analysis. Chromatography was carried out on HPLC column Kinetex C18
(150×3 mm, 2.6 μm) (Phenomenex, Torrance, CA, USA) held at 45°C.
The mobile phase flowing at a rate of 0.6 ml/min consisted of
ammonium formate (5 mM, pH=3.8) (solvent A) and acetonitrile
(solvent B). The gradient elution program was: from 20 to 60% (v/v)
of solvent B in 30 min, equilibration at 20% (v/v) of solvent B for
5 min. Mass spectrometric conditions were the same as in the
analysis of α-tomatine, data were scanned in full scan mode
(150–2,000 m/z).
Sample preparation for analysis and
metabolic screening
Acid hydrolysis of α-tomatine
Putative α-tomatine metabolites arising from
deglycosidation were prepared by acid hydrolysis of α-tomatine
standard. The hydrolysis proceeded in a solution of 0.25 M
hydrochloric acid prepared in methanol. α-Tomatine was dissolved at
a final concentration of 10 μM in the above-mentioned solution and
left 1 h at 60°C. After hydrolysis, 5 μl was injected directly onto
the column.
Analysis of α-tomatine
Ten microliters of the internal standard
(solasonine, 50 μM) and 100 μl of acetonitrile were added to a 100
μl sample aliquot. The sample was vortex mixed, centrifuged (12,700
× g, 10 min, 4°C) and the supernatant transferred to a vial with a
glass insert. Five microliters of the sample were injected onto the
column.
Metabolic screening
Frozen samples (−80°C) of incubation media and MCF-7
cell pellet reconstituted in 100 μl of water were thawed at room
temperature. Two hundred microliters of 10% (v/v) formic acid in
N,N-dimethylformamide (DMF) were added to a 100 μl sample aliquot.
The sample was vortex mixed, centrifuged (12,700 × g, 10 min, 4°C)
and transferred to a vial with a glass insert. Five microliters of
the sample were injected onto the column. The 10% (v/v) formic acid
in DMF was able to solubilize serum proteins, including albumin and
to dissociate the cholesterol/α-tomatine complex. In contrast to
the precipitation with acetonitrile, the putative metabolites could
neither be retained by protein precipitate nor in complexes with
cholesterol when 10% (v/v) formic acid in DMF was used for the
sample treatment.
Statistical analysis
The statistical data for caspase-8 and -9 assays,
flow cytometry and comet assay were evaluated with descriptive
statistics using Microsoft Office Excel (Microsoft Inc.). Data are
expressed as an arithmetic means ± SD of at least 3 independent
experiments and statistical analysis was performed using a
Student’s t-test. Significant differences were established at
p≤0.001.
The statistical data for WST-1, xCELLigence, ATP and
LDH assays were evaluated with GraphPad Prism (GraphPad Software
version 4.0, Inc., San Diego, CA, USA) software. Data are expressed
as an arithmetic means ± SD of at least 3 independent experiments
and statistical analysis was performed using one-way ANOVA test and
Dunnett’s post-hoc test for multiple comparisons. Significant
differences were established at p≤0.05.
Results
The cytotoxic effect of α-tomatine in
MCF-7 cells
We studied the effect of α-tomatine at various
concentrations (0, 1, 3, 6 and 9 μM) on cell viability and
proliferation of the MCF-7 cell line at different times of exposure
(24, 48 and 72 h). A WST-1 assay was used for viability measurement
and an xCELLigence system was used to measure proliferation. As
shown in Fig. 1A, α-tomatine
exerted a dose-dependent inhibitory effect on the viability of
MCF-7 cells. Treatment with α-tomatine at concentrations of 1 and 3
μM did not significantly alter the viability of the MCF-7 cell line
(compared with the untreated control) after 24, 48 and 72 h.
Treatment with 6 and 9 μM caused a significant decrease in the cell
viability after 24, 48 and 72 h. Similarly, the proliferation of
MCF-7 cells after α-tomatine treatment decreased in a
dose-dependent manner and the EC50 value after 72 h was
7.17 μM as determined by the xCELLigence system (Fig. 1B). The inhibition of proliferation
and viability of MCF-7 cells was the most pronounced after 24 h of
α-tomatine action.
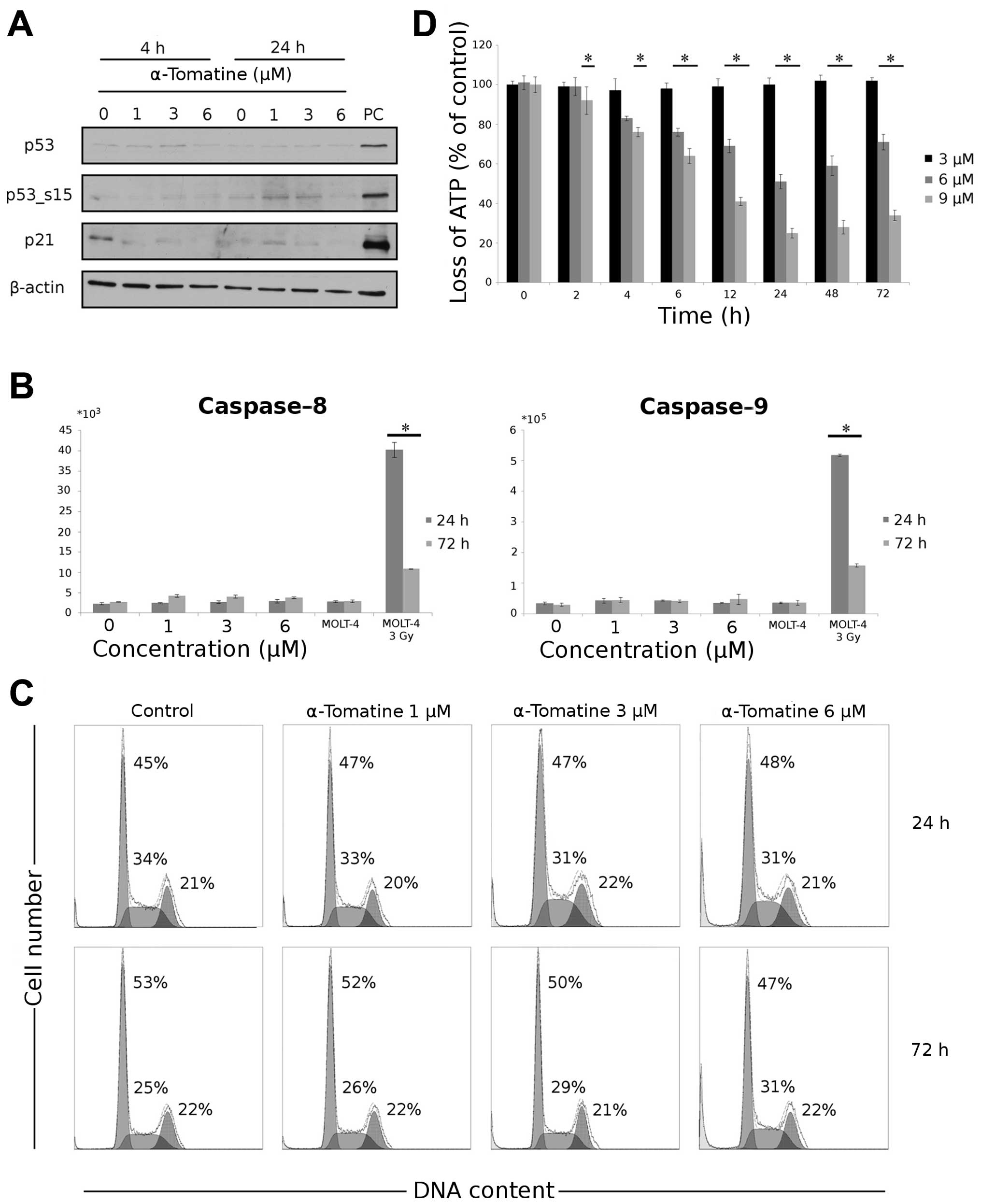
 | Figure 1Cytotoxic effect of α-tomatine in
human breast adenocarcinoma cell line MCF-7. Cultured cells were
treated with α-tomatine at various concentrations for 24, 48 and 72
h, then examined using (A) WST-1 test or (B) xCELLigence assay over
72 h. (C) Cultured cells were treated with α-tomatine (3, 6 and 9
μM) for 2, 4, 6, 12, 24, 48 and 72 h, then LDH release was
measured. (D) Induction and activation of Chk1 in MCF-7 cells
exposed to 1, 3 and 6 μM α-tomatine 4 h after the application of
the drug detected by western blotting. To confirm equal protein
loading, membranes were reincubated with β-actin. Chk1_s345 - Chk1
phosphorylated on serine 345. Chk1, check-point kinase 1. (E)
Time-lapse videomicroscopy was performed in MCF-7 cells after
α-tomatine treatment at various concentrations for 24, 48 and 72 h.
Values represent means ± SD of 3 independent experiments
(*p≤0.05 compared with the untreated control group with
one-way ANOVA test and Dunnett’s post-hoc test for multiple
comparisons). |
Furthermore, we studied the cytotoxic effect on
MCF-7 cells using the measurement of LDH release from damaged
cells. Again, the concentrations of 6 μM (after 4–72 h) and 9 μM
(after 2–72 h) of α-tomatine caused significant damage (Fig. 1C) to treated cells, with the highest
effect occurring after 24 h. The release of LDH is a result of
membrane destabilization.
The inhibition of proliferation corresponds with the
finding of check-point kinase 1 (Chk1) activation. In MCF-7 cells
treated with α-tomatine at concentrations of 1, 3 and 6 μM after 4
h, the amount of Chk1 increased. This increase was accompanied by
phosphorylation of Chk1 on serine 345 after the same incubation
time (Fig. 1D). After 24 h, the
changes in Chk1 and its phosphorylated form disappeared (data not
shown).
The morphology and behavior of MCF-7 cells after
treatment with 3 and 9 μM of α-tomatine were recorded over a 72-h
period using time-lapse videomicroscopy. Control cultures showed a
time-dependent increase in the number of cells culminating at 72 h,
when full confluence was reached. Similar timing and dynamics were
also observed upon 3 μM of α-tomatine treatment with no
morphological or behavioral alterations. Conversely, 9 μM of
α-tomatine significantly reduced proliferation of MCF-7 cells over
a 24-h period. Cells did not divide, their membranes demonstrated
reduced undulations and many detached from the substratum and
rapidly collapsed, releasing their content into the medium. On the
other hand, no specific apoptotic morphologies (i.e. blebbing) were
observed. In subsequent time intervals (24–72 h of treatment), a
number of thus affected cells decreased and the remaining cells
started to behave actively including mitoses. At the end of the
experiment (72 h), the treated culture clearly recovered and
continued in proliferation (Fig.
1E).
These results indicated the significant cytotoxic
effects of α-tomatine at concentrations of 6 and 9 μM after 4, 6,
12 and 24 h of treatment. After 48 and 72 h of treatment, the
cytotoxic effects persisted, but the cells started to proliferate
again and their viability increased.
Assessment of DNA damage in MCF-7 cells
after α-tomatine treatment
Since DNA is the main target of most cytotoxic
anticancer drugs, we focused on the induction of DNA damage in
MCF-7 cells after α-tomatine treatment. To assess DNA damage in
MCF-7 cells, we used the alkaline and neutral version of the comet
assay. The alkaline version is employed for the quantification of
SSBs of DNA. The concentrations of α-tomatine used (1–9 μM) did not
cause any significant SSBs after 4 h compared with
H2O2-treated MCF-7 cells (Fig. 2A–C and G). The neutral version
specifically determines DSBs of DNA. The same concentrations of
α-tomatine, as they were used in the alkaline version of the comet
assay, did not cause any significant DSBs after 4 h of treatment in
comparison with irradiated cells (20 Gy after 0.5 h). These results
indicated that α-tomatine caused no DNA damage in MCF-7 cells
(Fig. 2D–F and H).
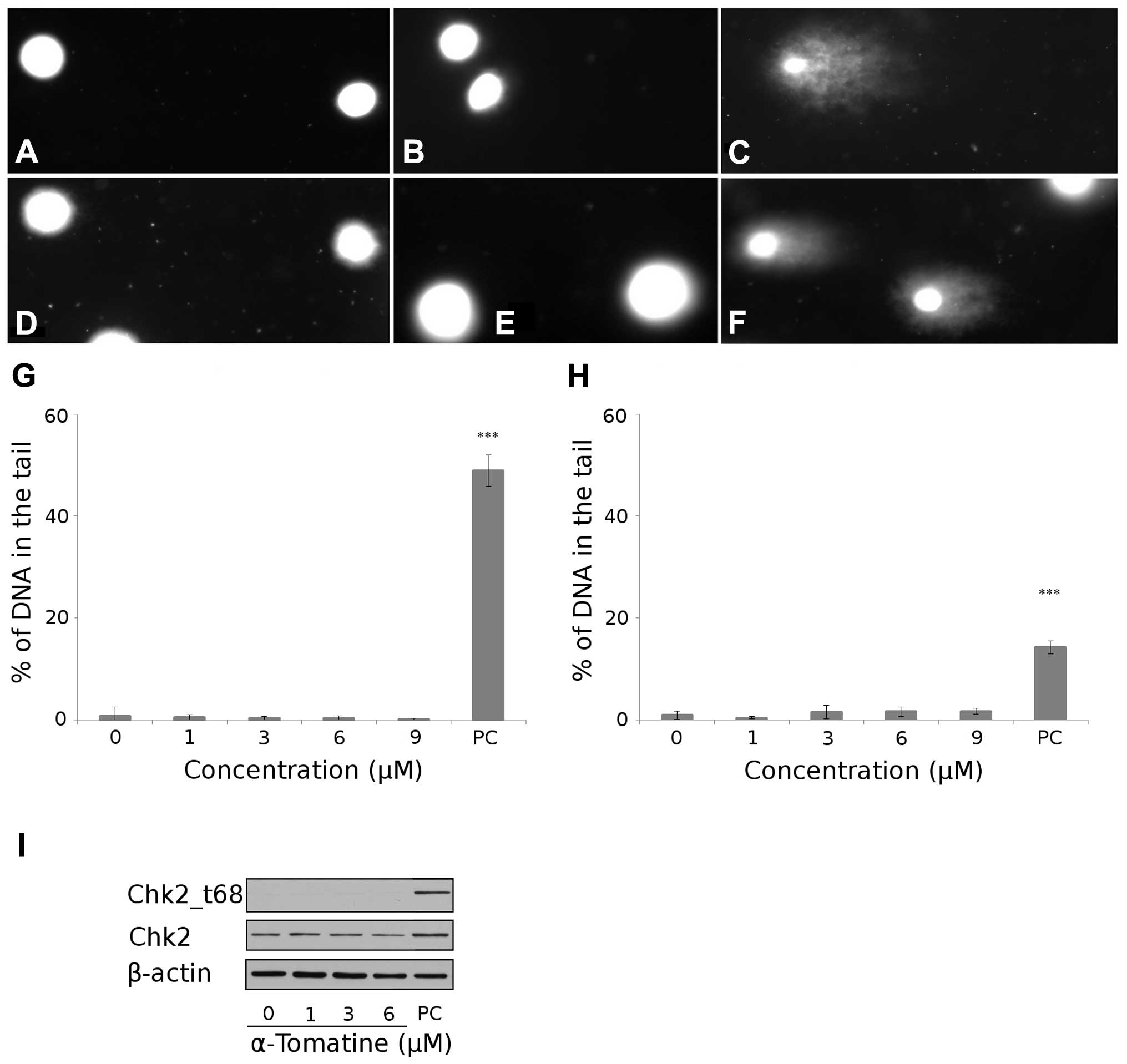 | Figure 2Detection of DNA damage. Digital
camera image of the comet assay; the alkaline version (top row),
(A) untreated MCF-7 cells, (B) 6 μM of α-tomatine-treated cells and
(C) 1.5% H2O2-treated cells (positive
control). Digital camera image of comet assay: the neutral version
(bottom row), (D) untreated MCF-7 cells, (E) 6 μM of
α-tomatine-treated cells and (F) cells after exposure of 20 Gy of
irradiation (positive control). Microscope magnification, ×20. (G)
The dependence of DNA single-strand breaks on the concentration of
α-tomatine after 4 h of exposure in MCF-7 cells. Asterisks indicate
values significantly (p<0.001) different from the control
(distinguished using Student’s t-test). PC, positive control (1.5%
H2O2). (H) The dependence of DNA
double-strand breaks on the concentration of α-tomatine after 4 h
of exposure in MCF-7 cells. Asterisk indicate values significantly
(p<0.001) different from the control (Student’s t-test). PC,
positive control (20 Gy γ radiation). (I) Changes in Chk2 and Chk2
phosphorylated on threonine 68 (Chk2_t68) detected by western
blotting 4 h after exposure to 1, 3 and 6 μM of α-tomatine. To
confirm equal protein loading, membranes were reincubated with
β-actin. PC, positive control (10 Gy γ radiation after 24 h). Chk2,
check-point kinase 2. |
In the next step, we confirmed no DSBs of DNA by
determining check-point kinase 2 (Chk2) non-activation. After 4 h
of α-tomatine treatment (1, 3 and 6 μM), no phosphorylation on
threonine 68 of Chk2 occurred in comparison with the positive
control group (10 Gy irradiation after 24 h) (Fig. 2I).
Mechanism of cell death and cell cycle
analysis
Although our results indicate no α-tomatine-induced
DNA damage in MCF-7 cells, we elucidated whether apoptosis is
induced in the cells exposed to α-tomatine by another triggering
mechanism.
Initially, we evaluated changes in the levels of
proteins p53, p53 phosphorylated on serine 15 and
p21WAF1/CIP1 in α-tomatine-treated (1, 3 and 6 μM) MCF-7
cells after 4 and 24 h of incubation. No changes in the levels of
these proteins were observed (Fig.
3A).
Then, we measured the caspase-8 and -9 activity
after 24 and 72 h of incubation with α-tomatine (1, 3 and 6 μM). No
significant increase in the activity of these caspases was observed
after α-tomatine treatment (Fig.
3B).
We used cell cycle analysis by flow cytometry to
assess possible apoptosis after α-tomatine treatment (1, 3 and 6
μM) for 24 and 72 h. As shown in Fig.
3C, the concentrations of α-tomatine used caused no specific
apoptotic sub-G1 peak after 24 and 72 h of treatment.
To confirm our theory that α-tomatine did not cause
apoptosis in MCF-7 cells, we measured the loss of ATP in
α-tomatine-treated cells (3, 6 and 9 μM) after 2, 4, 6, 12, 24, 48
and 72 h. Fig. 3D demonstrates a
significant decrease, in comparison to untreated cells, in the
content of ATP in cells treated with 9 μM of α-tomatine at all
treatment intervals between 4 and 72 h and with 6 μM of α-tomatine
at all treatment intervals between 6 and 72 h. Moreover, to gain
further insight into the mechanism of cell death caused by
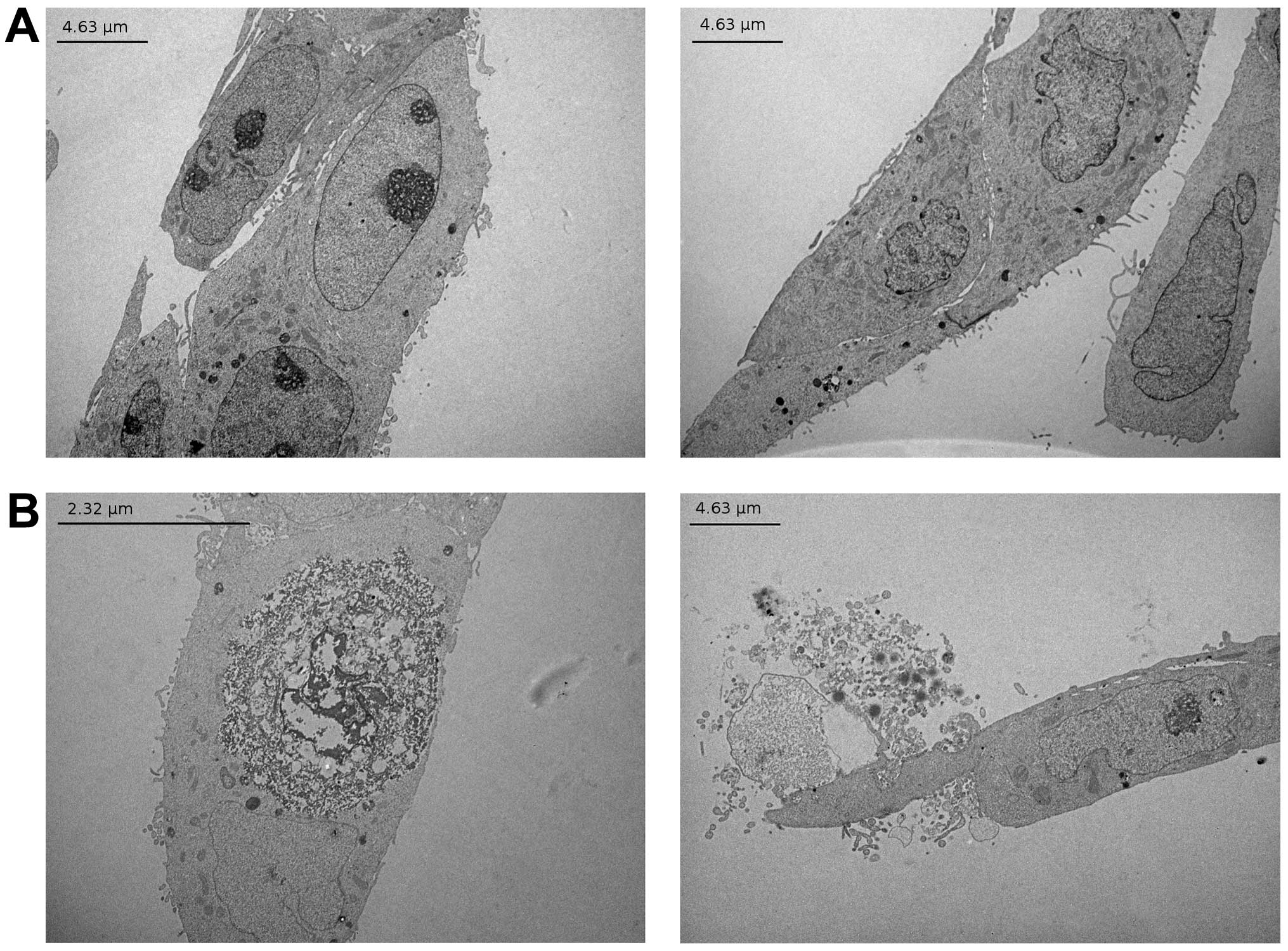
α-tomatine, transmission electron microscopy of MCF-7 cells treated
with 6 μM of α-tomatine was carried out (Fig. 4). The treated cells showed swelling
and disintegration of both nuclear and plasma membranes,
characteristic of a rapid necrosis, but no typical morphologic
signs of apoptosis, such as cell shrinkage, nuclear fragmentation,
chromatin condensation, nuclear and cytoplasmic blebbing and the
formation of apoptotic bodies.
Stability of α-tomatine in culture medium
and its biotransformation
Previous results indicated that α-tomatine at
concentrations of 6 and 9 μM caused a cytotoxic effect in MCF-7
cells during the first 24 h, but after 48 and 72 h of incubation
cell proliferation and viability began to increase. Thus, we
monitored the concentration of α-tomatine in the medium during the
incubation with cells in order to determine whether it changes with
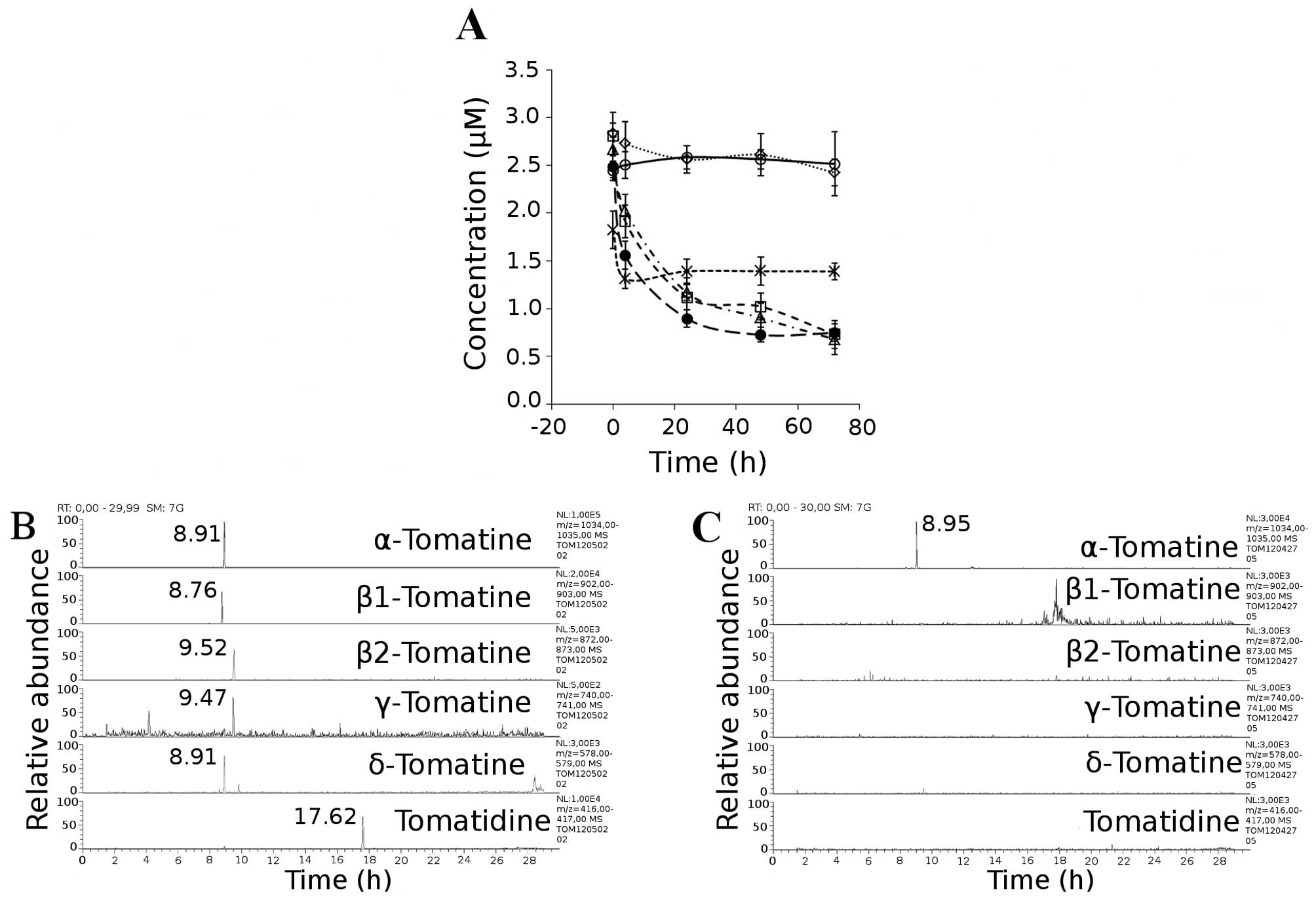
time. We detected a decrease in the level of α-tomatine from the
initial value of 2.49 to 1.56 μM after 4 h, to 0.90 μM after 24 h,
to 0.73 μM after 48 h, and to 0.75 μM after 72 h.
As cholesterol is known to form an insoluble
precipitate with α-tomatine, its effect in the incubation media was
evaluated. Initially, the stability of 3 μM of α-tomatine in PBS
buffer (pH 7.4) during the 72 h of incubation was confirmed
(Fig. 5A). Subsequently, the effect
of cholesterol was tested. Since cholesterol is insoluble in
aqueous solutions, albumin was used as a carrier. When fetal calf
serum was replaced by pure albumin in the standard incubation
medium, a decrease in α-tomatine in the cell-free medium was not
observed during the incubation time. By contrast, >50% of
α-tomatine was lost during the first 4 h of incubation when albumin
containing the same concentration of cholesterol as the fetal calf
serum (100 μM) was used. Therefore, interaction of α-tomatine with
cholesterol appears to be the main reason for the decrease in
α-tomatine in the medium during incubation, at least in the absence
of cells.
To support this hypothesis, we also carried out
tests to exclude the possible metabolism of α-tomatine caused by
MCF-7 cells or enzymes present in the fetal calf serum added to an
incubation medium. The incubation of 3 μM of α-tomatine in the
standard incubation medium with and without MCF-7 cells led to a
60–70% loss of α-tomatine during the first 24 h. A comparable
decrease in α-tomatine was observed when inactivated serum was
added to the medium. Thus, the decrease is not due to
biotransformation by plasma enzymes.
To screen the potential biotransformation by MCF-7
cells, a standard incubation medium with native fetal calf serum
was used. Based on the α-tomatine structure, we anticipated that
its biotransformation can proceed by deglycosidation or by
oxidation (hydroxylation or N-oxidation) (24) of the aglycone. As these
deglycosidated derivatives, with the exception of tomatidine, are
not available on the market, acid hydrolysis of α-tomatine was
carried out in order to get a reaction mixture containing compounds
derived from α-tomatine by consecutive loss of sugar units.
Hydrolysis products found in the reaction mixture are presented in
Fig. 5B. Representative
chromatograms for each compound were extracted from full scan data
(Fig. 5B) and the retention times
and masses (m/z) are presented in Table
I.
 | Table IRetention times and m/z of
pseudomolecular ions [M+H]+ of α-tomatine hydrolysis
products. |
Table I
Retention times and m/z of
pseudomolecular ions [M+H]+ of α-tomatine hydrolysis
products.
| Compound | tr
(min) | m/z |
|---|
| Tomatine | 8.91 | 1034.5 |
|
GLC-GLC-GAL-TOD | 8.76 | 902.5 |
|
XYL-GLC-GAL-TOD | 9.52 | 872.5 |
| GLC-GAL-TOD | 9.47 | 740.5 |
| GAL-TOD | 8.91 | 578.5 |
| Tomatidine
(TOD) | 17.62 | 416.4 |
Subsequently, samples of the incubation media and
MCF-7 cell pellets after 4, 24, 48 and 72 h of incubation with 3 μM
of α-tomatine were obtained and targeted metabolic screening for
deglycosidated metabolites was carried out. Representative
chromatograms (Fig. 5C) for
metabolites possibly arising from deglycosidation were extracted
from mass spectrometric data and compared with chromatograms of
derivatives obtained by hydrolysis. Target metabolites were not
found either in the incubation media or in the cell pellet. With
this experiment, we proved that during incubation with an MCF-7
cell line, α-tomatine is not subjected to deglycosidation by
biotransformation or by chemical decomposition.
Samples were also tested for the presence of
oxidative biotransformation products (hydroxylation, N-oxidation)
of α-tomatine and tomatidine. Since chemical standards of these
metabolites are not available and chemical synthesis is
problematic, the mass spectrometric data obtained was only
inspected for masses of suspected mono-hydroxy (Δ m/z 16),
di-hydroxy (Δ m/z 32) or N-oxide derivatives (Δ m/z 16). None of
these suspected metabolites were found in the samples.
Discussion
The mechanism of the cytotoxic action of α-tomatine
has been studied intensively; nevertheless, the existing results
differ significantly. In mouse EL4 lymphoma cells, α-tomatine at
the concentration of 8 μM induces apoptosis with changes in the
cell cycle distribution of these cells (53.8% sub-G1 peak)
(19). In human prostatic
adenocarcinoma PC-3 cells, a marked increase of apoptotic cells in
sub-G1 phase (95%) was observed after treatment with 2 μM of
α-tomatine for 24 h (18). The
induction of apoptosis is closely related to the activation of
cysteine proteases called caspases (25). In PC-3 cells, the activation of
caspase-3, -8 and -9 in 2 μM of α-tomatine-treated cells was
demonstrated (18). Contrary to
these results, the induction of programmed cell death with no
changes in caspase-3, -6, -7, -8 and -9 activity and with no
changes in cell cycle distribution was demonstrated in
α-tomatine-treated human leukemic cells HL60 and K562 (14) as well as MOLT-4 (20). Similarly, we demonstrated that
caspase-8 and -9 are not activated. Our results indicate no changes
in cell cycle distribution, no apoptotic sub-G1 peak in MCF-7 cells
and no morphological signs of apoptosis after treatment with
α-tomatine. α-Tomatine at the concentrations of 1–9 μM after 4 h
induced neither SSBs nor DSBs detectable by comet assay in MCF-7
cells. Moreover, the activation of Chk2 related to DNA damage
(26) was not detected. The
tumor-suppressor phosphoprotein p53 plays a significant role in
preventing inappropriate cell proliferation and maintaining genome
integrity in response to genotoxic stress, resulting in cell-cycle
arrest, DNA repair, apoptosis or senescence (27). Our experimental data show no changes
in the level of p53 and p53 phosphorylated on serine 15 in MCF-7
cells after 4 and 24 h of α-tomatine treatment (1–6 μM). The levels
of the cyclin-dependent kinase inhibitor p21WAF1/CIP1
also remained unchanged. Apoptosis is a process highly dependent on
the content of intracellular ATP; there is an evident physiological
difference in cells undergoing apoptosis or necrosis (28). We demonstrated a loss of ATP in
α-tomatine-treated cells (6 and 9 μM) after 72 h, which supports
the results of this study discussed above. A possible explanation
of the cytotoxic mechanism could be the fact that cholesterol in
biological membranes serves as a target for α-tomatine (see
below).
A number of studies have indicated that α-tomatine
inhibits the growth of various human cancer cell lines at low μM
concentrations within 1–48 h of treatment (14–18,20,21).
Our results showed a cytotoxic effect of α-tomatine at slightly
higher concentrations than in the previous studies. We demonstrated
that 6 and 9 μM of α-tomatine caused an inhibition of proliferation
and a decrease in viability after 24 h in MCF-7 cells. However,
after 48 and 72 h of α-tomatine treatment, a recovery of these
cells occurred. Our study also revealed a transient increase and
activation of check-point kinase 1 (Chk1) 4 h after α-tomatine
application, which corresponds with the inhibition of
proliferation. These changes in the levels of proteins disappeared
after 24 h of incubation - an effect that corresponds with cell
recovery. Chk1 is essential for cell viability and proliferation
and for maintaining DNA integrity. Although it is active even in
unperturbed cell cycles, it is further activated in response to DNA
damage or stalled replication fork (26,29,30).
We investigated why the cells recovered after 24 h
of incubation. In the course of that, we observed a decrease in the
levels of α-tomatine solved in the media throughout the incubation.
As this phenomenon occurred only when cholesterol was present in
the media, the formation of an insoluble α-tomatine cholesterol
complex (2) should be the main
reason. It was reported (31,32)
that α-tomatine is, in contrast to the related saponins, not
interface seeking. The accumulation of α-tomatine at the water-air
interface should therefore not be the cause of its potential
disappearing from the solution.
Regarding biotransformation, fungal pathogens of
tomato produce tomatinase enzymes which hydrolyze α-tomatine by
removing the complete sugar moiety (β-lycotetraose) or partial
sugars (9,24,33).
Only aglycone (tomatidine) released is susceptible to hydroxylation
by cytochrome P-450. Since the removal of even a single
monosaccharide from the sugar moiety reduces the membranolytic
activity of α-tomatine substantially (34), deglycosylation serves as a
resistance mechanism of fungal pathogens. Our findings of no
putative metabolites of α-tomatine, neither in the medium nor in
cell pellets, are consistent with the current knowledge which
suggests that mammalian cells do not metabolize α-tomatine.
Cholesterol in the membranes of MCF-7 cells probably
served as a target for α-tomatine as well. The release of the
cytosolic enzyme LDH which occurred especially at concentrations of
α-tomatine 6 and 9 μM, is a marker of membrane integrity loss
typical for early necrosis and only the late stage of apoptosis
(35). The absence of any
characteristic features of apoptosis, the morphological signs as
well as the ATP depletion observed, rather indicate necrotic cell
death. Although the rupture of membranes occurs in necrosis
mediated by different mechanisms (36), the binding of α-tomatine to membrane
cholesterol and the consequent membrane disruption may have had a
triggering effect in the present study (37,38).
Membrane disintegration by several other agents was shown to induce
necrosis (35,39). Steroidal glycoalkaloids form
complexes with free 3β-hydroxy sterols, such as cholesterol,
present in the outer leaflet of the plasma and outer mitochondrial
membranes of animal cells. A network of these complexes undergoes a
lateral aggregation through sugar-sugar interactions between the
sugar moieties, causing a rearrangement and subsequent loss of the
barrier function. α-Tomatine has the strongest membrane-disruptive
effect of all Solanum glycoalkaloids studied thus far
(34,38).
In conclusion, our results show that a single
application of α-tomatine has an antiproliferative effect on cancer
cells within 24 h of incubation, but then the cells recover. Since
α-tomatine is not biotransformed in the MCF-7 cell line via
deglycosidation or oxidation during 72 h but the concentration of
α-tomatine significantly decreases in solutions and media
containing cholesterol, this effect is most probably related to the
ability of α-tomatine to bind with cholesterol present in the
culture medium. Our study is the first to describe this phenomenon,
which should be taken into consideration when interpreting results
from an in vitro assay with α-tomatine. α-Tomatine does not
induce DNA damage or the activation of caspases; it does not change
the levels of proteins p53 and p21 in the MCF-7 cell line but
rather causes a decrease in the cellular ATP. These facts, along
with the morphological changes observed, suggest that the decrease
in viability of MCF-7 cells by α-tomatine is not due to apoptosis
induction. A probable mechanism involved in the cytotoxicity is the
membrane-disruptive effect due to its binding with membrane
cholesterol.
Acknowledgements
The authors thank Ms. Nadezda Mazankova and Ms. Zora
Komarkova for their technical assistance. This study was supported
by the grant GACR P303/12/P536 and the programs PRVOUK P37/01 and
SVV-2013-266901 of the Charles University in Prague. The
publication was co-financed by the European Social Fund and the
state budget of the Czech Republic. Project no.
CZ.1.07/2.3.00/30.0022.
References
|
1
|
Blankemeyer JT, White JB, Stringer BK and
Friedman M: Effect of α-tomatine and tomatidine on membrane
potential of frog embryos and active transport of ions in frog
skin. Food Chem Toxicol. 35:639–646. 1997.
|
|
2
|
Roddick JG: The steroidal glycoalkaloid
α-tomatine. Phytochemistry. 13:9–25. 1974.
|
|
3
|
Simons V, Morrissey JP, Latijnhouwers M,
Csukai M, Cleaver A, Yarrow C and Osbourn A: Dual effects of plant
steroidal alkaloids on Saccharomyces cerevisiae. Antimicrob
Agents Chemother. 50:2732–2740. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Thorne HV, Clarke GF and Skuce R: The
inactivation of herpes simplex virus by some Solanaceae
glycoalkaloids. Antiviral Res. 5:335–343. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Filderman RB and Kovacs BA:
Anti-inflammatory activity of the steroid alkaloid glycoside,
tomatine. Br J Pharmacol. 37:748–755. 1969. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Morrow WJ, Yang YW and Sheikh NA:
Immunobiology of the Tomatine adjuvant. Vaccine. 22:2380–2384.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Roddick JG: The
acetylcholinesterase-inhibitory activity of steroidal
glycoalkaloids and their aglycones. Phytochemistry. 28:2631–2634.
1989. View Article : Google Scholar
|
|
8
|
Bergers WW and Alink GM: Toxic effect of
the glycoalkaloids solanine and tomatine on cultured neonatal rat
heart cells. Toxicol Lett. 6:29–32. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Friedman M: Tomato glycoalkaloids: role in
the plant and in the diet. J Agric Food Chem. 50:5751–5780. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Friedman M, Fitch TE and Yokoyama WE:
Lowering of plasma LDL cholesterol in hamsters by the tomato
glycoalkaloid tomatine. Food Chem Toxicol. 38:549–553. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Smith DB, Roddick JG and Jones JL: Potato
glycoalkaloids: some unanswered questions. Trends Food Sci Tech.
7:126–131. 1996. View Article : Google Scholar
|
|
12
|
Lee ST, Wong PF, He H, Hooper JD and
Mustafa MR: Alpha-tomatine attenuation of in vivo growth of
subcutaneous and orthotopic xenograft tumors of human prostate
carcinoma PC-3 cells is accompanied by inactivation of nuclear
factor-kappa B signaling. PLoS One. 8:e577082013. View Article : Google Scholar
|
|
13
|
Tomsik P, Micuda S, Sucha L, Cermakova E,
Suba P, Zivny P, Mazurova Y, Knizek J, Niang M and Rezacova M: The
anticancer activity of alpha-tomatine against mammary
adenocarcinoma in mice. Biomed Pap Med Fac Univ Palacky Olomouc
Czech Repub. 157:153–161. 2013.PubMed/NCBI
|
|
14
|
Chao MW, Chen CH, Chang YL, Teng CM and
Pan SL: α-Tomatine-mediated anti-cancer activity in vitro and in
vivo through cell cycle- and caspase-independent pathways. PLoS
One. 7:e440932012.
|
|
15
|
Lee KR, Kozukue N, Han JS, Park JH, Chang
EY, Baek EJ, Chang JS and Friedman M: Glycoalkaloids and
metabolites inhibit the growth of human colon (HT29) and liver
(HepG2) cancer cells. J Agric Food Chem. 52:2832–2839. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shieh JM, Cheng TH, Shi MD, Wu PF, Chen Y,
Ko SC and Shih YW: α-Tomatine suppresses invasion and migration of
human non-small cell lung cancer NCI-H460 cells through
inactivating FAK/PI3K/Akt signaling pathway and reducing binding
activity of NF-κB. Cell Biochem Biophys. 60:279–310. 2011.
|
|
17
|
Shih YW, Shieh JM, Wu PF, Lee YC, Chen YZ
and Chiang TA: α-Tomatine inactivates PI3K/Akt and ERK signaling
pathways in human lung adenocarcinoma A549 cells: effect on
metastasis. Food Chem Toxicol. 47:1985–1995. 2009.
|
|
18
|
Lee ST, Wong PF, Cheah SC and Mustafa MR:
Alpha-tomatine induces apoptosis and inhibits nuclear factor-kappa
B activation on human prostatic adenocarcinoma PC-3 cells. PLoS
One. 6:e189152011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang YW, Wu CA and Morrow WJ: The
apoptotic and necrotic effects of tomatine adjuvant. Vaccine.
22:2316–2327. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kúdelová J, Seifrtová M, Suchá L, Tomšík
P, Havelek R and Řezáčová M: A-tomatine activates cell cycle
checkpoints in the absence of DNA damage in human leukemic MOLT-4
cells. J Appl Biomed. 11:93–103. 2013.
|
|
21
|
Shi MD, Shih YW, Lee YS, Cheng YF and Tsai
LY: Suppression of
12-O-tetradecanoylphorbol-13-acetate-induced MCF-7 breast
adenocarcinoma cells invasion/migration by α-tomatine through
activation PKCα/ERK/NF-κB-dependent MMP-2/MMP-9 expressions. Cell
Biochem Biophys. 66:161–174. 2013.
|
|
22
|
Olive PL and Banáth JP: The comet assay: a
method to measure DNA damage in individual cells. Nat Protoc.
1:23–29. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Collins AR, Dušinská M, Gedik CM and
Štětina R: Oxidative damage to DNA: do we have a reliable
biomarker? Environ Health Perspect. 104(Suppl 3): S465–S469. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Weltring KM, Wessels J and Pauli GF:
Metabolism of the tomato saponin α-tomatine by Gebberella
pulicaris. Phytochemistry. 48:1321–1328. 1998.
|
|
25
|
Thornberry NA and Lazebnik Y: Caspases:
enemies within. Science. 281:1312–1316. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bartek J and Lukas J: Chk1 and Chk2
kinases in checkpoint control and cancer. Cancer Cell. 3:421–429.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bai L and Zhu WG: p53: structure, function
and therapeutic applications. J Cancer Mol. 2:141–153. 2006.
|
|
28
|
Eguchi Y, Shimizu S and Tsujimoto Y:
Intracellular ATP levels determine cell death fate by apoptosis or
necrosis. Cancer Res. 57:1835–1840. 1997.PubMed/NCBI
|
|
29
|
Cho SH, Toouli CD, Fujii GH, Crain C and
Parry D: Chk1 is essential for tumor cell viability following
activation of the replication checkpoint. Cell Cycle. 4:131–139.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu Q, Guntuku S, Cui XS, Matsuoka S,
Cortez D, Tamai K, Luo G, Carattini-Rivera S, DeMayo F, Bradley A,
Donehower LA and Elledge SJ: Chk1 is an essential kinase that is
regulated by Atr and required for the G2/M DNA damage
checkpoint. Genes Dev. 14:1448–1459. 2000.PubMed/NCBI
|
|
31
|
Keukens EA, de Vrije T, Fabrie CH, Demel
RA, Jongen WM and de Kruijff B: Dual specificity of sterol-mediated
glycoalkaloid induced membrane disruption. Biochim Biophys Acta.
1110:127–136. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Walker BW, Manhanke N and Stine KJ:
Comparison of the interaction of tomatine with mixed monolayers
containing phospholipid, egg sphingomyelin, and sterols. Biochim
Biophys Acta. 1778:2244–2257. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sandrock RW and Vanetten HD: Fungal
sensitivity to and enzymatic degradation of the phytoanticipin
α-tomatine. Phytopathology. 88:137–143. 1998.PubMed/NCBI
|
|
34
|
Keukens EA, de Vrije T, van den Boom C, de
Waard P, Plasman HH, Thiel F, Chupin V, Jongen WM and de Kruijff B:
Molecular basis of glycoalkaloid induced membrane disruption.
Biochim Biophys Acta. 1240:216–228. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Do TN, Rosal RV, Drew L, Raffo AJ, Michl
J, Pincus MR, Friedman FK, Petrylak DP, Cassai N, Szmulewicz J,
Sidhu G, Fine RL and Brandt-Rauf PW: Preferential induction of
necrosis in human breast cancer cells by a p53 peptide derived from
the MDM2 binding site. Oncogene. 22:1431–1444. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Vanlangenakker N, Vanden Berghe T, Krysko
DV, Festjens N and Vandenabeele P: Molecular mechanisms and
pathophysiology of necrotic cell death. Curr Mol Med. 8:207–220.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gee JM, Wortley GM, Johnson IT, Price KR,
Rutten AA, Houben GF and Penninks AH: Effects of saponins and
glycoalkaloids on the permeability and viability of mammalian
intestinal cells and on the integrity of tissue preparations in
vitro. Toxicol In Vitro. 10:117–128. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Keukens EA, de Vrije T, Jansen LA, de Boer
H, Janssen M, de Kroon AI, Jongen WM and de Kruijff B:
Glycoalkaloids selectively permeabilize cholesterol containing
biomembranes. Biochim Biophys Acta. 1279:243–250. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Dong Z, Saikumar P, Weinberg JM and
Venkatachalam MA: Internucleosomal DNA cleavage triggered by plasma
membrane damage during necrotic cell death. Involvement of serine
but not cysteine proteases. Am J Pathol. 151:1205–1213.
1997.PubMed/NCBI
|