Introduction
Prostate cancer is known to be a chronic disease and
better detection and prognostication allowing long-term management
is needed to improve quality of life for patients. Different
prostate cancer cell lines express different levels of cluster
designation (CD) antigens on their cell surface. Cell lines such as
LNCaP, PC3 and DU145 are derived from metastatic lesions and, thus,
have metastatic patterns which can be predicted by their CD
phenotypes (1). The pattern of
expression of CD antigens, thus, potentially provides prognostic
information (2,3). Some of these CD molecules are
transmembrane-4-superfamily (TM4SF) members or tetraspanins, a
family of 33 members in mammals (4–6). The
exact biochemical functions of tetraspanins are still unknown,
however, they have been linked to various processes including
signal transduction pathways, cellular activation, proliferation,
motility, adhesion, tissue differentiation, angiogenesis, tumour
progression and metastasis. Most tetraspanins become downregulated
in metastatic tumours; however, the CD151 glycoprotein was
identified by Testa et al(7)
to be the first tetraspanin member to play a role as a promoter of
metastasis.
Binding of CD151 to the laminin binding integrins
α3β1, α6β1, α6β4 and α7β1 promotes extracellular matrix (ECM)
remodelling and there is evidence of localisation of CD151 and α3
integrins at the intercellular junction between endothelial
cell-tumour cells, leading to tumour cell transendothelial
migration across secondary sites (8). CD151 is a point of motility regulation
on both cell-cell and cell-stroma levels which involves control of
integrin activation and distribution (9). The findings from our laboratory show
that CD151 has a better prognostication value than traditional
Gleason grading and can distinguish aggressive forms of prostate
cancer (10), and that in
vitro manipulation of CD151 leads to change in prostate cancer
migration (11). It is predicted
that heightened CD151-positivity in prostate cancer cells will lead
to more progressive disease; we have explored this in a mouse model
of prostate cancer in the present study.
Materials and methods
Cell lines and reagents
The human prostate cancer cell lines PC-3, LNCaP and
DU145 were obtained from the American Type Culture Collection
(ATCC, Manassas, VA, USA). Cell lines were maintained and
propagated as monolayer cultures in RPMI-1640 medium (Invitrogen
Life Technologies, Carlsbad, CA, USA) respectively, with 10% foetal
bovine serum (FBS) (Invitrogen Life Technologies) and 100 U/ml
penicillin and 100 μg/ml streptomycin (Invitrogen Life
Technologies). Antibodies used are monoclonal mouse anti-human
CD151 antibody, 11B1.G4 (IgG2a) a kind gift from Professor L.
Ashman’s laboratory (Newcastle University, NSW, Australia)
(12,13), monoclonal mouse anti-human β-tubulin
antibody (Invitrogen Life Technologies), monoclonal rat anti-mouse
CD31 antibody (MEC 13.3) (BD Pharmigen™; BD Biosciences, San Jose,
CA, USA) and monoclonal rat anti-mouse LYVE-1 antibody (XB-13)
(Santa Cruz Biotechnology, Dallas, TX, USA).
Total cellular protein isolation and
western blotting
Modified radioimmunoprecipitation (RIPA) buffer was
used to extract proteins from the cells. Medium was removed from
cells and cells were washed twice with ice-cold PBS before addition
of ice-cold RIPA buffer containing 1× Complete Mini EDTA-free
protease inhibitor tablet (Roche Diagnostics, Castle Hill, NSW,
Australia). Protein concentrations of the whole cell lysates were
determined using the Bradford assay (14). Western blot analysis was used to
determine CD151 expression. Proteins were separated by SDS-PAGE.
Protein bands were then transferred to nitrocellulose paper and
incubated with CD151 antibody (Ab) and peroxidase conjugated
anti-mouse Ab, respectively. Peroxidase linked anti-mouse antibody
was purchased from Amersham™ (GE Healthcare Biosciences, Rydalmere,
NSW, Australia). β-tubulin levels were used as a loading control.
Protein bands were visualized after chemiluminescent reaction.
RNA isolation
RNA was isolated using the Rneasy Mini kit (Qiagen)
according to the manufacturer’s instructions. RNA quantity was
assessed using a UV spectrophotometer at A260 nm and RNA quality
was determined using the A260 nm/A280 nm ratio.
Reverse-transcript PCR and quantitative
PCR
RNA was extracted from prostate cancer cell lines,
LNCaP, PC-3 and DU-145 as previously described. cDNA synthesis were
performed using M-MuLV kit (Invitrogen Life Technologies). cDNA
samples were then analysed using CD151 (Assay ID Hs00388381_m1) and
18S (Assay ID Hs99999901_s1) (as control) TaqMan Gene Expression
assays (Applied Biosystems/Life Technologies). PCR amplification
was performed in a 25 μl final volume (total 54 ng cDNA per
reaction) using Applied Biosystems 7500 real-time PCR system. mRNA
expression of CD151 was normalized in relation to the control 18S
expression. Data are expressed as fold difference to LNCaP cell
line.
In vivo mouse model of prostate
cancer
A severe combined immunodeficiency (SCID) mouse
model orthotopically implanted with the human prostate cancer cell
line, PC-3 constitutively expressing a fluorescent marker dsRED was
used (15). In total, 12 SCID mice
were implanted intraprostatically with human PC-3-DsRed cells, as a
model of prostate cancer metastasis. Cells were inoculated in 0.01
ml PBS into the prostate (1×106 cells/mouse). Mice were
sacrificed 7 weeks after tumour cell inoculation.
The extent of tumour spread was examined by in
vivo fluorescent imaging. For regular weekly in vivo
imaging, the mice under study were first denuded of all hair in the
region of interest. They were then anaesthetised with a 1–5%
isofluorane/95% oxygen air mixture and placed under the light for
imaging. The emission from DsRed was measured using a Fuji Film
LAS-3000 camera housed alongside an image intensifier and light
source in a custom built machine. Primary tumour volume was
calculated using a common ellipsoid estimation length ×
width2 × 0.5 as previously described (16). Upon harvesting, mice were
anaesthetised and primary tumours and regional lymph nodes were
removed, weighed and processed for histology and
immunohistochemical analyses. Tissues were fixed in 10% formalin,
followed by paraffin embedding and sectioning. Mouse organs
(spleen, liver and kidneys) were also removed and weighed for
examination of any organ damage.
Immunohistochemistry
For histological analysis of the xenograft mouse
model, tissue sections (5 μm) were cut from formalin-fixed,
paraffin-embedded tumour specimens of the implanted prostate cancer
specimens and neighbouring lymph nodes (if tumour was found).
Routine haematoxylin and eosin (H&E) staining was performed for
each specimen for histological analysis. CD151, CD31 (angiogenesis
marker) and LYVE-1 (lymphatic marker) protein expression in tissue
sections were measured using Dako LSAB+ kit (Dako).
CD151, CD31 and LYVE-1 stainings were performed as previously
described (10,15,17,18).
Ab used was anti-human CD151 Ab (4 μg/ml working concentration),
CD31 Ab (diluted 1:10) and LYVE-1 Ab (diluted 1:50). Negative
control immunostaining using isotype control antibody substituted
for each primary antibody was also performed. Immunostainings were
visualized with 3,3′-diaminobenzidine (DAB; Dako Australia Pty,
Ltd., Campbellfield, VIC, Australia) and sections were then
counterstained with haematoxylin.
Quantification of endothelial and
lymphatic vasculature
Staining quantification was performed using light
microscopy. CD31 and LYVE-1 stainings were analysed on digitised
colour images and evaluated by counting of positive staining
vessels. An intratumoural microvessel and lymphatic vessel were
defined as any brown-stained (DAB immunoperoxidase stain with
anti-CD31 antigen and anti-LYVE-1 antigen, respectively) cell or
cell cluster clearly separated from adjacent vessels (19). Tumour vascularity was measured as
microvessel density (MVD) and lymphatic vessel density (LVD) in
areas containing intense neovascularisation (neovascular ‘hot
spot’), as previously described (19,20).
The step-by-step procedures to identify these ‘hot spots’ were
previously described (21).
Primary and secondary lesions were examined for MVD
and LVD. Up to 10 hot spots (depending on tumour size) were
selected at low magnification (×100) and vessels were counted in a
representative high magnification (×200). The MVD or LVD ratios
were calculated as vascular density/(0.74 mm2 for each
×200 field). The vasculature ratios of primary tumour and secondary
lymph node metastasis were compared and analysed. In addition, mice
were also assigned into two groups of those that formed metastasis
and those without metastasis. MVD and LVD of primary tumours of
these two groups were also compared and analysed.
Statistical analysis
Statistical analysis was carried out using
independent samples Student’s t-tests. All P-values of <0.05
were considered to indicate a statistically significant result.
Results are expressed as mean ± standard error of the mean (SEM).
The statistical tests performed for MVD and LVD were two-tailed,
equal (F-value >0.05) or unequal variance (F-value ≤0.05),
Student’s t-test. Pearson correlation was also performed to examine
the relationship between CD151 staining, MVD, LVD and tumour
weights.
Results
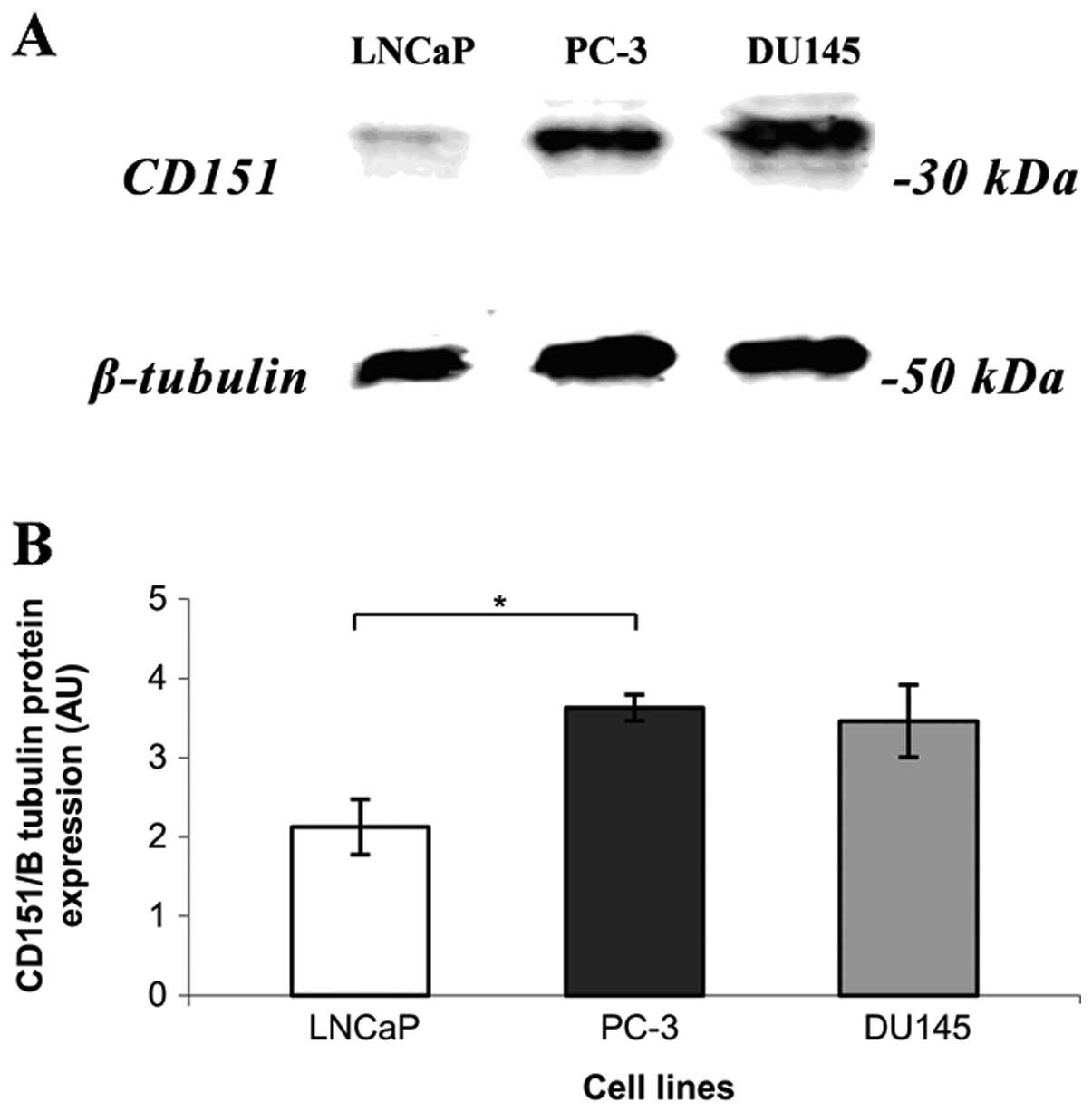
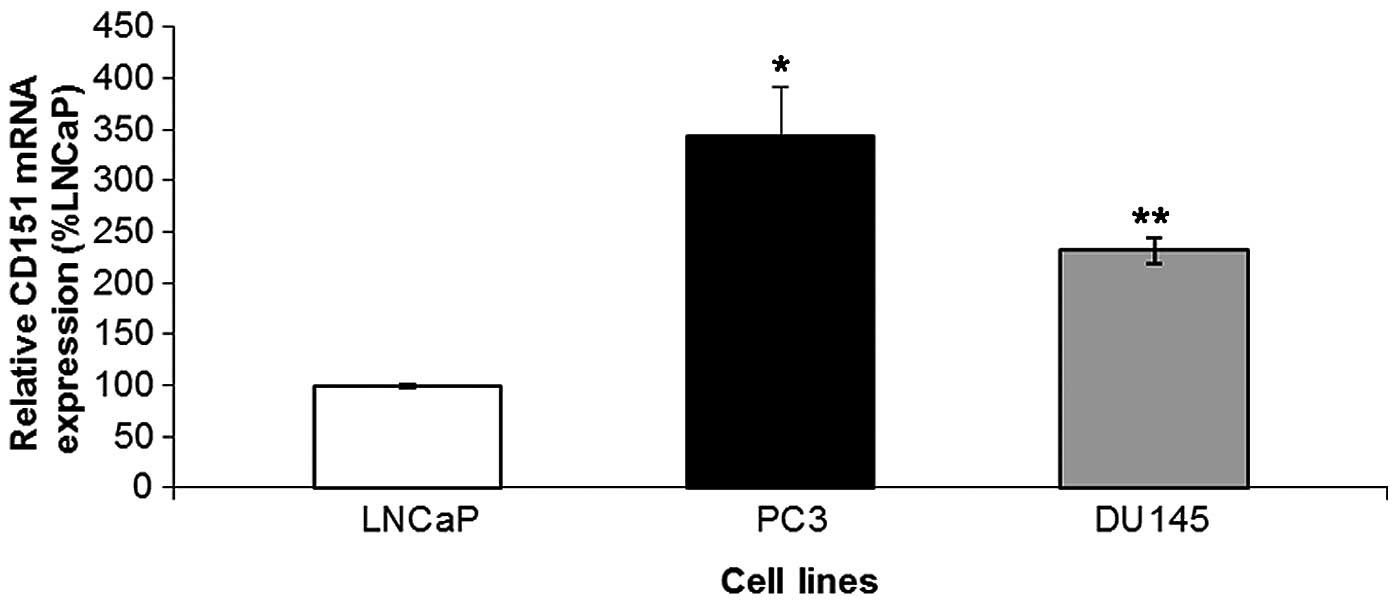
CD151 is expressed in commonly used
prostate cancer cell lines
CD151 is heterogeneously expressed across all
prostate cancer cell lines investigated. LNCaP, an
androgen-responsive cell line, expressed low levels of CD151
protein and mRNA compared to two androgen-insensitive cell lines,
PC-3 and DU-145 (Figs. 1 and
2, respectively).
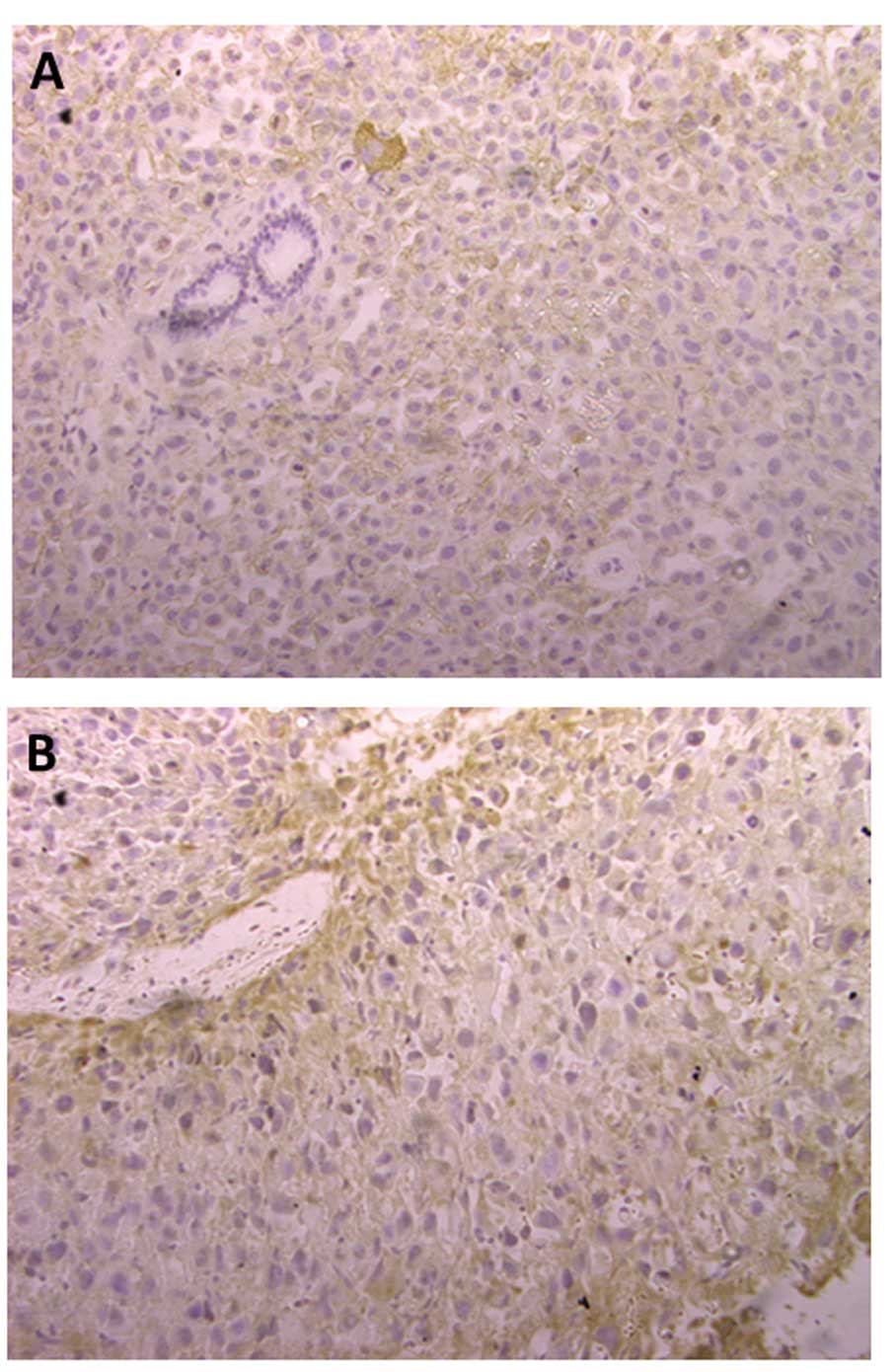
CD151 is a prognostic marker in mouse
model of prostate cancer
All mice developed primary tumours at the site of
inoculation (100% tumour take rate) and 8 of 12 (67%) mice
developed pelvic lymph node metastases. CD151 expression was
investigated via immunohistochemistry (Fig. 3). CD151 expression is predominantly
localised in the cellular membrane and cytosol in prostate cancer
cells. Weaker nuclear staining can also be seen. CD151 expression
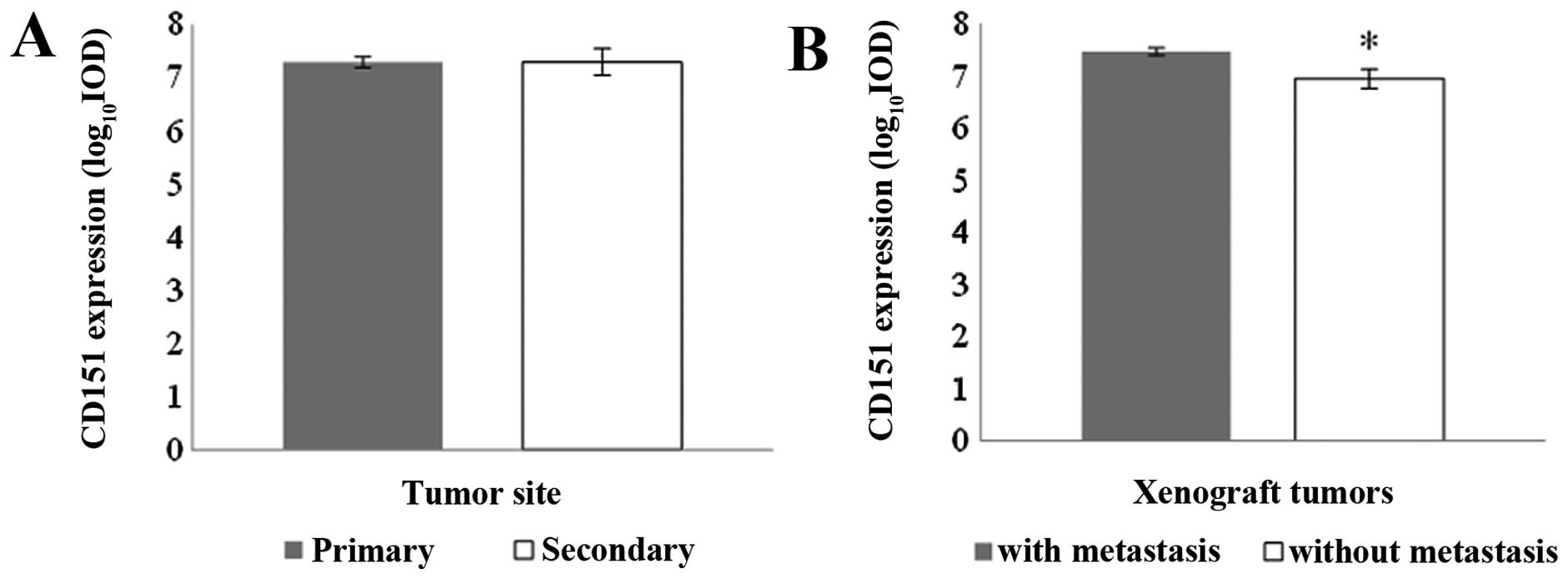
was examined in the primary and secondary lesions and no difference
was seen between the two tumour sites (Fig. 4A). However, the expression of CD151
in primary tumour specimens from the group of mice forming
metastasis was found to be significantly higher than in those mice
without metastasis formation (Fig.
4B). There was no evidence of non-specific staining in the
control slides (data not shown). This confirms that CD151 has a
prognostic value in this mouse model of prostate cancer.
Angiogenesis and lymphangiogenesis
markers in mouse model of prostate cancer
We next investigated angiogenesis and
lymphangiogenesis markers. The angiogenesis marker CD31 was used as
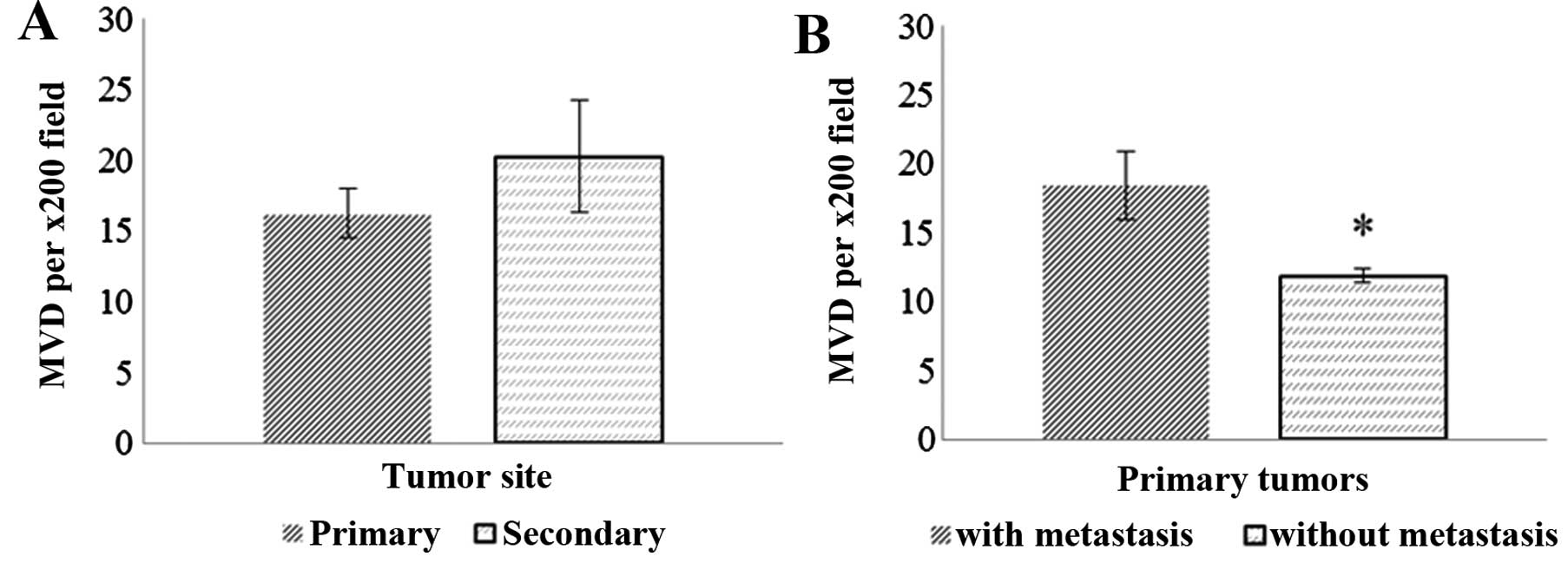
a marker and microvessel density (MVD) and was measured in primary
and secondary lesions. No difference in MVD was detected between
primary and secondary lesions. However, MVD was significantly
higher in primary prostatic xenografts forming metastases at nearby
lymph nodes compared to primary lesions that did not form
metastasis (Fig. 5). There was no
association between tumour weight and MVD of the metastasis forming
xenografts (data not shown).
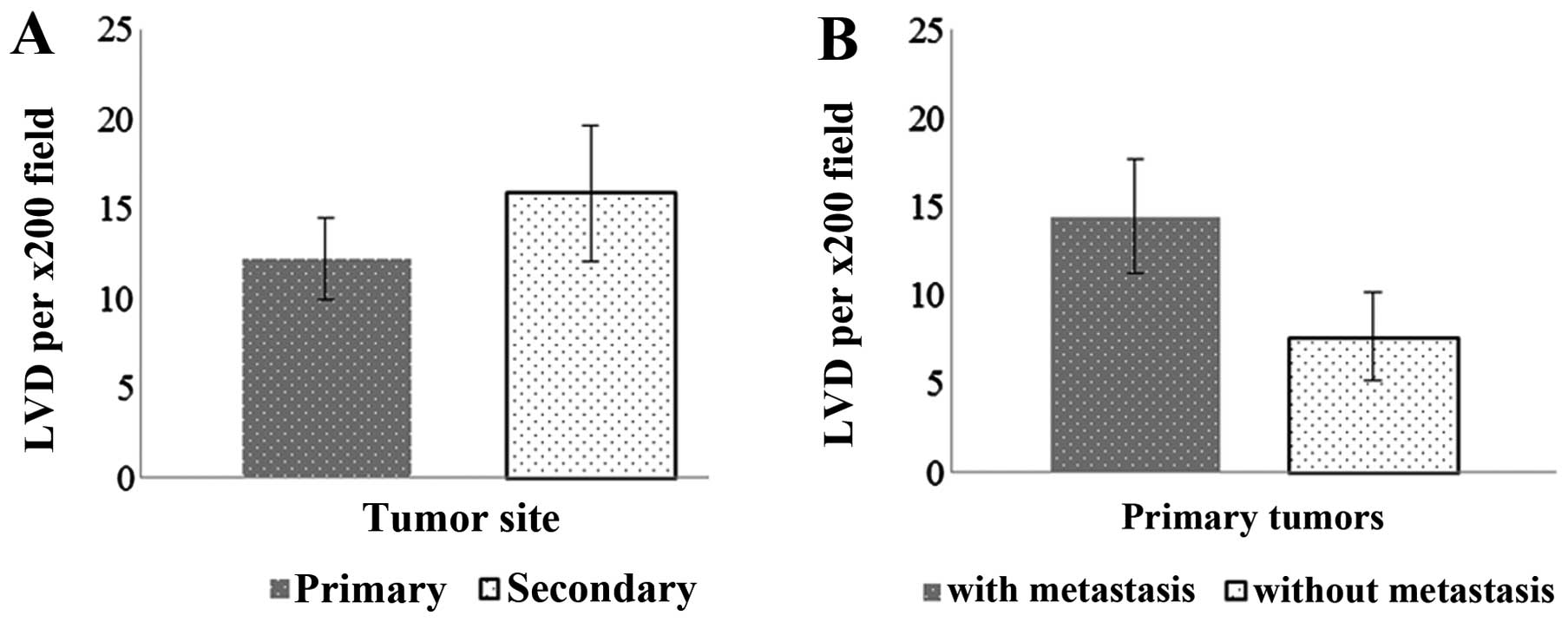
Lymphatic vessel density (LVD) was not significantly
different between primary and secondary lesions (Fig. 6) but there was a similar trend
(P=0.19) toward increased LVD in primary xenografts forming
metastases compared to those that did not form metastasis.
MVD and LVD relationships with CD151
expression levels and tumour growth
The level of CD151 expression, MVD and LVD was not
significantly different between primary and secondary lesions. The
relationships between CD151 staining, MVD and LVD were examined in
primary lesions (Table I). CD151
expression was positively correlated with LVD (LYVE-1 staining)
with statistical significance (r=0.62; P=0.03). The association
between CD151 and MVD (CD31 staining) did not reach statistical
significance (r=0.48; P=0.058) and MVD and LVD were not correlated.
MVD, LVD and CD151 expression did not correlate with tumour size
(data not shown). Table II
demonstrated that LVD may have a prognostic value; 83% of mice with
high LVD progressed to form lymph node metastases.
 | Table IPearson correlation analysis of
CD151, CD31 and LYVE-1 staining of primary PC-3 xenograft
tumours. |
Table I
Pearson correlation analysis of
CD151, CD31 and LYVE-1 staining of primary PC-3 xenograft
tumours.
| Primary tissue
staining comparison | Pearson correlation
(r-value) | P-value
(two-tailed) |
|---|
| CD151 vs. MVD | 0.48 | 0.058 |
| CD151 vs. LVD | 0.62 | 0.03a |
| MVD vs. LVD | 0.50 | 0.09 |
| Table IILVD in relation to metastasis
formation in PC-3 orthotopic xenograft mouse model of prostate
cancer. |
Table II
LVD in relation to metastasis
formation in PC-3 orthotopic xenograft mouse model of prostate
cancer.
| LVD in primary
lesions | n | Lymph node
metastasis (%) |
|---|
| High (> mean
value of LVD) | 6 | 5 (83) |
| Low (< mean
value of LVD) | 6 | 3 (50) |
Discussion
Tumour metastasis is a multi-factorial process and
tumour cells are sensitive to local microenvironmental cues that
regulate normal physiological functions such as wound healing and
epithelial morphogenetic changes leading to malignant behaviour
(22). CD151 is heterogeneously
expressed across prostate cancer cell lines and the level of
expression was noted to be higher in more highly aggressive cell
lines. However, it has been shown that both LNCaP and PC-3 cell
lines express integrin α3β1, which is a major partner of CD151 in
the promotion of motility and metastasis (23). Integrin expression profiles between
LNCaP and PC-3 cells differ, especially α3 integrin where LNCaPs
have reduced expression compared to PC-3 cells which may explain
the altered level of tumourigenicity between the two cell lines
(24). This suggests the importance
of integrins in invasive behaviour of prostate cancer. CD151-α3β1
association is found in most tissues that express α3β1 (25,26),
which reinforces the importance of CD151 in the various functions
of integrin α3β1. Our finding suggests that CD151 correlates with
aggressiveness of the disease but whether CD151 has a role in
transformation of androgen-responsive to androgen-irresponsive
states cannot be concluded and is being explored in our
laboratory.
While it is evident that CD151 expression correlates
more with malignant types of prostate cancer, CD151 may have a role
in angiogenesis as well as tumour motility and invasion. Our
finding indicates that angiogenesis predicts tumour progression,
which is in agreement with other studies (20,27–29).
Although the exact mechanism is not known, there has been some
evidence that suggests the role of CD151 in blood vessel formation
in animal models. CD151 is expressed in vascular endothelial cells
and associated with integrin β1, β3, β4, α2, α3, α5 and α6 chains
(30). A study using CD151-null
mice has shown a possible role of CD151 in tumour angiogenesis
(31). These mice were viable and
no phenotypic change was reported, however, pathological
angiogenesis was greatly affected, supporting a pro-angiogenic role
for CD151 specifically in pathological conditions (31,32).
Indeed, in animal models of myocardial ischaemia, introduction of
CD151 via gene delivery improved capillary densities (33,34).
In vitro studies using endothelial cells have shown that
CD151-integrin complexes have a role in endothelial cell
proliferation, morphogenesis and migration, which are important in
the angiogenesis process (30,35,36).
In prostate cancer, angiogenesis is associated with progression and
prognosis (17,20,37–39).
CD151 involvement in angiogenesis has been
demonstrated through the discovery of CD151-integrin complex
localisation within endothelial cells and communication between
tumour and endothelial cells via CD151, both essential for the
formation of tumour neovascularisation (8,36,40).
Absence of CD151 in mouse lung endothelial cells led to alterations
in angiogenic ability which was found to be through disruption in
association with laminin-binding integrins and CD151-null mice also
had defective pathological angiogenesis (31). Importantly, CD151 may have a role in
regulation of tumour cell-endothelial cell dynamics and migrating
behaviour of the endothelial cell which in turn promotes
angiogenesis (40). In order to
investigate the possible link between CD151 and angiogenesis, the
changes in an angiogenesis marker were observed in a mouse model of
prostate cancer. In prostate cancer, angiogenesis (using CD31 as a
marker) was found to be an independent prognostic predictor
(17). When angiogenesis was
examined in the initiation and progression of prostate cancer using
the spontaneous autochthonous transgenic adenocarcinoma of the
mouse prostate (TRAMP) mouse model, CD31 staining revealed changes
in angiogenesis patterns from low-grade prostate intra-epithelial
neoplasia (PIN), high-grade PIN and amongst different prostate
cancer grades (27). Our results
indicate that CD151 expression and angiogenesis was a prognostic
indicator in the PC-3 orthotopic xenograft mouse model and this is
not correlated to tumour size.
The role of CD151 in angiogenesis is perhaps through
assisting communication between tumour cells and endothelial cells
(40). Many studies have found that
CD151-integrin (especially α3β1 and α6β4) complexes are localised
at the tumour cell and endothelial cell contact area (8,36,40).
Expression of CD151 is required for integrin distribution within
the endothelial intercellular contacts, which in turn promotes
angiogenesis (31). While
cross-talk between tumour cells and endothelial cells assisting in
transendothelial migration of tumour cells is important for
angiogenesis, CD151 has been found to be a membrane linker, through
which other signalling proteins stimulate the important regulator
of endothelial cell function, endothelial nitric oxide synthase
(41). Direct CD151 gene delivery
into endothelial cells results in increased proliferation,
migration and tube formation (42).
Integrins, especially α3β1, have been found to be involved in the
induction of angiogenesis by endothelial cells (43–46).
Based on this information, it is likely that CD151 promotes
pathological angiogenesis, via its association with integrin α3β1
and this association could be important at tumour or endothelial
cells. Thus, inhibition of this complex via anti-CD151 agents might
disrupt angiogenesis formation at either tumour or endothelial
cells.
Our findings indicated statistically positive
relationships between CD151 and LVD. Although LVD was not found to
have prognostic value in this mouse model, the CD151 and LVD
correlation suggest a possible role of CD151 in lymphangiogenesis
in prostate cancer. This is particularly relevant in the PC-3
orthotopic xenograft model as the lymph nodes are the most common
sites of metastases (15). This
provides a basis for further exploration.
In conclusion, the involvement of CD151 in prostate
cancer migration and metastasis is underscored and demonstrated in
the present study. Because cell motility and invasion are decisive
events for tumour metastasis, inhibiting CD151 is an attractive
approach for inhibition of motility and invasion signalling pathway
in prostate cancer. The present study supports further exploration
into the role of CD151 in prostate cancer angiogenesis and
lymphangiogenesis.
Acknowledgements
The present study was supported by grant funding to
S.D. and A.G.F. from the Austin Medical Research Foundation and to
E.D.W. from the National Health & Medical Research Council of
Australia and the Victorian Government’s Operational Infrastructure
Support Program. We thank Professor Leonie K. Ashman for supplying
CD151 antibody for our studies.
Abbreviations:
|
CD
|
cluster designation
|
|
ECM
|
extracellular matrix
|
|
TM4SF
|
transmembrane-4-superfamily
|
|
SCID
|
severe combined immunodeficiency
|
|
MVD
|
microvessel density
|
|
LVD
|
lymphatic vessel density
|
|
PIN
|
prostate intra-epithelial
neoplasia
|
References
|
1
|
Liu AY, Roudier MP and True LD:
Heterogeneity in primary and metastatic prostate cancer as defined
by cell surface CD profile. Am J Pathol. 165:1543–1556. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Liu AY and True LD: Characterization of
prostate cell types by CD cell surface molecules. Am J Pathol.
160:37–43. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Liu AY: Differential expression of cell
surface molecules in prostate cancer cells. Cancer Res.
60:3429–3434. 2000.PubMed/NCBI
|
|
4
|
Hemler ME: Tetraspanin functions and
associated microdomains. Nat Rev Mol Cell Biol. 6:801–811. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Berditchevski F: Complexes of tetraspanins
with integrins: more than meets the eye. J Cell Sci. 114:4143–4151.
2001.PubMed/NCBI
|
|
6
|
Levy S and Shoham T: Protein-protein
interactions in the tetraspanin web. Physiology. 20:218–224. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Testa JE, Brooks PC, Lin JM and Quigley
JP: Eukaryotic expression cloning with an antimetastatic monoclonal
antibody identifies tetraspanin (PETA-3/CD151) as an effector of
human tumor cell migration and metastasis. Cancer Res.
59:3812–3820. 1999.
|
|
8
|
Longo N, Yanez-Mo M, Mittelbrunn M, et al:
Regulatory role of tetraspanin CD9 in tumor-endothelial cell
interaction during transendothelial invasion of melanoma cells.
Blood. 98:3717–3726. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nishiuchi R, Sanzen N, Nada S, et al:
Potentiation of the ligand-binding activity of integrin α3β1 via
association with tetraspanin CD151. Proc Natl Acad Sci USA.
102:1939–1944. 2005.
|
|
10
|
Ang J, Lijovic M, Ashman LK, Kan K and
Frauman AG: CD151 protein expression predicts the clinical outcome
of low-grade primary prostate cancer better than histologic
grading: a new prognostic indicator? Cancer Epidemiol Biomarkers
Prev. 13:1717–1721. 2004.
|
|
11
|
Ang J, Fang BL, Ashman LK and Frauman AG:
The migration and invasion of human prostate cancer cell lines
involves CD151 expression. Oncol Rep. 24:1593–1597. 2010.PubMed/NCBI
|
|
12
|
Sincock PM, Mayrhofer G and Ashman LK:
Localization of the transmembrane 4 superfamily (TM4SF) member
PETA-3 (CD151) in normal human tissues: comparison with CD9, CD63
and α5β1 integrins. J Histochem Cytochem. 45:515–525.
1997.PubMed/NCBI
|
|
13
|
Geary SM, Cambareri AC, Sincock PM, Fitter
S and Ashman LK: Differential tissue expression of epitopes of the
tetraspanin CD151 recognised by monoclonal antibodies. Tissue
Antigens. 58:141–153. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bradford MM: Rapid and sensitive method
for quantitation of microgram quantities of protein utilizing
principle of protein-dye binding. Anal Biochem. 72:248–254. 1976.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zeng Y, Opeskin K, Goad J and Williams ED:
Tumor-induced activation of lymphatic endothelial cells via
vascular endothelial growth factor receptor-2 is critical for
prostate cancer lymphatic metastasis. Cancer Res. 66:9566–9575.
2006. View Article : Google Scholar
|
|
16
|
Tomayko MM and Reynolds CP: Determination
of subcutaneous tumor size in a athymic (nude) mice. Cancer
Chemother Pharmacol. 24:148–154. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mehta R, Kyshtoobayeva A, Kurosaki T, et
al: Independent association of angiogenesis index with outcome in
prostate cancer. Clin Cancer Res. 7:81–88. 2001.PubMed/NCBI
|
|
18
|
Wroel T, Mazur G, Dziegiel P, et al:
Density of intranodal lymphatics and VEGF-C expression in B-cell
lymphoma and reactive lymph nodes. Folia Histochem Cytobiol.
44:43–47. 2006.PubMed/NCBI
|
|
19
|
Weidner N, Semple JP, Welch WR and Folkman
J: Tumor angiogenesis and metastasis - correlation in invasive
breast carcinoma. N Engl J Med. 324:1–8. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Weidner N, Carroll PR, Flax J, Blumenfeld
W and Folkman J: Tumor angiogenesis correlates with metastasis in
invasive prostate carcinoma. Am J Pathol. 143:401–409.
1993.PubMed/NCBI
|
|
21
|
Weidner N: Current pathologic methods for
measuring intratumoral microvessel density within breast carcinoma
and other solid tumors. Breast Cancer Res Treat. 36:169–180. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Quaranta V: Motility cues in the tumor
microenvironment. Differentiation. 70:590–598. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Schmelz M, Cress AE, Scott KM, et al:
Different phenotypes in human prostate cancer: α6 or α3 integrin in
cell-extracellular adhesion sites. Neoplasia. 4:243–254. 2002.
|
|
24
|
Witkowski CM, Rabinovitz I, Nagle RB,
Affinito KS and Cress AE: Characterization of integrin subunits,
cellular adhesion and tumorgenicity of 4 human prostate cell-lines.
J Cancer Res Clin Oncol. 119:637–644. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yauch RL, Kazarov AR, Desai B, Lee RT and
Hemler ME: Direct extracellular contact between integrin
α3β1 and TM4SF protein CD151. J Biol Chem.
275:9230–9238. 2000.
|
|
26
|
Kazarov AR, Yang X, Stipp CS, Sehgal B and
Hemler ME: An extracellular site on tetraspanin CD151 determines α3
and α6 integrin-dependent cellular morphology. J Cell Biol.
158:1299–1309. 2002.PubMed/NCBI
|
|
27
|
Huss WJ, Hanrahan CF, Barrios RJ, Simons
JW and Greenberg NM: Angiogenesis and prostate cancer:
identification of a molecular progression switch. Cancer Res.
61:2736–2743. 2001.PubMed/NCBI
|
|
28
|
Steiner I, Jung K, Miller K, Stephan C and
Erbersdobler A: Expression of endothelial factors in prostate
cancer: A possible role of caveolin-1 for tumour progression. Oncol
Rep. 27:389–395. 2012.PubMed/NCBI
|
|
29
|
Gray DR, Huss WJ, Yau JM, et al:
Short-term human prostate primary xenografts: an in vivo model of
human prostate cancer vasculature and angiogenesis. Cancer Res.
64:1712–1721. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sincock PM, Fitter S, Parton RG, Berndt
MC, Gamble JR and Ashman LK: PETA-3/CD151, a member of the
transmembrane 4 superfamily, is localised to the plasma membrane
and endocytic system of endothelial cells, associates with multiple
integrins and modulates cell function. J Cell Sci. 112:833–844.
1999.
|
|
31
|
Takeda Y, Kazarov AR, Butterfield CE, et
al: Deletion of tetraspanin CD151 results in decreased pathological
angiogenesis in vivo and in vitro. Blood. 109:1524–1532. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wright MD, Geary SM, Fitter S, et al:
Characterization of mice lacking the tetraspanin superfamily member
CD151. Mol Cell Biol. 24:5978–5988. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zuo HJ, Liu ZX, Liu XC, et al: Assessment
of myocardial blood perfusion improved by CD151 in a pig myocardial
infarction model. Acta Pharmacol Sin. 30:70–77. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lan RF, Liu ZX, Liu XC, Song YE and Wang
DW: CD151 promotes neovascularization and improves blood perfusion
in a rat hind-limb ischemia model. J Endovasc Ther. 12:469–478.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang XA, Kazarov AR, Yang X, Bontrager
AL, Stipp CS and Hemler ME: Function of the tetraspanin CD151-α6β1
integrin complex during cellular morphogenesis. Mol Biol Cell.
13:1–11. 2002.
|
|
36
|
Yáñez-Mó M, Alfranca A, Cabanas C, et al:
Regulation of endothelial cell motility by complexes of tetraspan
molecules CD81/TAPA-1 and CD151/PETA-3 with α3β1 integrin localized
at endothelial lateral junctions. J Cell Biol. 141:791–804.
1998.PubMed/NCBI
|
|
37
|
Offersen BV, Borre M and Overgaard J:
Immunohistochemical determination of tumor angiogenesis measured by
the maximal microvessel density in human prostate cancer. APMIS.
106:463–469. 1998. View Article : Google Scholar
|
|
38
|
Trojan L, Thomas D, Knoll T, Grobholz R,
Alken P and Michel MS: Expression of pro-angiogenic growth factors
VEGF, EGF and bFGF and their topographical relation to
neovascularisation in prostate cancer. Urol Res. 32:97–103. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhu W and Dahut WL: Tumor angiogenesis as
an early marker of long-term prostate cancer mortality. Future
Oncol. 6:341–345. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Sadej R, Romanska H, Baldwin G, et al:
CD151 regulates tumorigenesis by modulating the communication
between tumor cells and endothelium. Mol Cancer Res. 7:787–798.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zheng ZZ and Liu ZX: CD151 gene delivery
increases eNOS activity and induces ECV304 migration, proliferation
and tube formation. Acta Pharmacol Sin. 28:66–72. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zuo HJ, Lin JY, Liu ZY, et al: Activation
of the ERK signaling pathway is involved in CD151-induced
angiogenic effects on the formation of CD151-integrin complexes.
Acta Pharmacol Sin. 31:805–812. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Mitchell K, Svenson KB, Longmate WM, et
al: Suppression of integrin α3β1 in breast cancer cells reduces
cyclooxygenase-2 gene expression and inhibits tumorigenesis,
invasion, and cross-talk to endothelial cells. Cancer Res.
70:6359–6367. 2010.
|
|
44
|
Wang X, Ferreira AM, Shao Q, Laird DW and
Sandig M: β3 integrins facilitate matrix interactions during
transendothelial migration of PC3 prostate tumor cells. Prostate.
63:65–80. 2005.
|
|
45
|
Mitchell K, Szekeres C, Milano V, et al:
α3β1 integrin in epidermis promotes wound angiogenesis and
keratinocyte-to-endothelial-cell crosstalk through the induction of
MRP3. J Cell Sci. 122:1778–1787. 2009.
|
|
46
|
Dominguez-Jimenez C, Yanez-Mo M, Carreira
A, et al: Involvement of α3 integrin/tetraspanins complexes in the
angiogenic response induced by angiotensin II. FASEB J.
15:1457–1459. 2001.
|