Introduction
Mounting evidence indicates that iron plays a
crucial role in malignant cell proliferation (1,2). A
dysregulated iron metabolism reveals a poor outcome in breast
cancer and esophageal adenocarcinoma patients (3,4).
Deferoxamine (DFX), a high affinity iron chelator, is known to
upregulate inflammatory mediators and it has been shown to have
significant and selective antitumor activity, although the
mechanisms of action remain to be completely elucidated (5–7). DFX
has the ability of hypoxia mimetics to induce a hypoxic state which
results in the upregulation or stabilization of hypoxia-inducible
factor-1 (HIF-1) (8,9). HIF-1 is a nuclear protein involved in
the transcriptional activation of a broad array of genes that
activate mitogenic, pro-invasive, pro-angiogenic and pro-metastatic
genes (10–12). Hence, DFX-induced hypoxia is a
complicated event, which may affect various pathways and gene
expression through the regulation of HIF-1 expression.
Epithelial-mesenchymal transition (EMT) is a
critical process for tumor invasion and metastasis (13–16).
It is characterized by loss of cell polarity and cell adhesion,
repression of E-cadherin expression and increased cell motility. At
the same time, the traits of epithelial cells such as long-lasting
morphological and molecular changes are altered to mesenchymal
characteristics, such as fibroblastic morphology, increased
vimentin and N-cadherin expression, enhanced migration and invasion
capacities, thus causing cancer cell metastasis (14,17).
It is important to note that the stabilization and activation of
HIF-1 are the most important mechanisms that promotes metastasis
and thereby, increase tumor aggressiveness (18,19).
For example, hypoxia or HIF-1 overexpression induces EMT and
metastatic phenotypes through the direct activation of Twist in
head and neck squamous cell carcinoma (HNSCC) (20). Hypoxia-induced EMT may be enhanced
by the formation of the HIF-1/β-catenin complex, which increases
the transcriptional activity of HIF-1α and consequently facilitates
EMT under hypoxic conditions in hepatocellular carcinoma (HCC)
(21).
In the present study, we treated colorectal cancer
(CRC) cells with DFX to create a DFX-induced hypoxic cell model,
and then explored EMT-associated features, such as the expression
of EMT-associated molecules, the cellular location of E-cadherin
and vimentin, cell morphological changes, adhesion, migration and
invasion abilities. Our study showed that DFX treatment increased
HIF-1α expression and enhanced EMT. Thus, we reported the potential
mechanism underlying DFX-induced hypoxia on cell migration and
invasion in CRC cells.
Materials and methods
Cell culture and cell treatments
HT29 and HCT116 cells were grown in RPMI-1640
(Gibco-BRL) supplemented with 10% fetal bovine serum (FBS) as we
previously described (22). For DFX
treatments, we utilized concentrations of 0, 50, 100 and 200 μM
respectively, of DFX supplemented with 10% FBS. Forty-eight hours
later, cell morphological changes were observed or cell lysates
were extracted to perform western blot analysis.
Western blotting, immunofluorescence
assays and antibodies
Western blotting and immunofluorescence assays were
carried out as previously described (23). Antibodies to HIF-1α (Novus
Biologicals, Littleton, CO, USA), E-cadherin and vimentin were
applied in the immunofluorescence staining (Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA). Vimentin (prediluted),
plakoglobin, N-cadherin and glyceraldehyde-3-phosphate
dehydrogenase (Abcam, Cambridge, UK) were all commercial
products.
Hematoxylin and eosin (H&E)
staining
Exponentially growing cells were seeded in
coverslips and then exposed to 100 μM of DFX. Forty-eight hours
later, coverslips were fixed in 4% paraformaldehyde for 15 min,
washed in phosphate-buffered saline (PBS) and running water,
followed by incubation with hematoxylin solution for 5 min and
washed under running water. Subsequently, the coverslips were
dipped in acid-ethanol, washed again, and then stained in
eosin-ethanol (1% eosin in 80% ethanol) for 3 min, subjected to
sequential dehydration and mounted. The coverslips were observed
under an Olympus microscope (Olympus Optical Co., Ltd., Tokyo,
Japan).
Immunofluorescence
Cells growing on coverslips were fixed with 4%
paraformaldehyde at room temperature for 15 min, washed with PBS
and permeabilized with 0.1% Triton X-100 for 5 min. After blocking
with 1% bovine serum albumin (BSA) for 30 min, cells were incubated
with E-cadherin and vimentin antibodies for 2 h. After washing with
PBS, IgG Texas Red- (TR-) and IgG-FITC-labeled secondary antibodies
were added and incubated for another 1 h. Cells were then rinsed
with PBS followed by Hoechst 33258 staining to reveal nuclei.
Fluorescence images were captured and analyzed using the Olympus
CKX41 fluorescence microscope (Olympus Optical Co., Ltd.).
Adhesion assay
Substrates for adhesion assays included 25 μg/ml
fibronectin or 10 mg/ml heat-inactivated BSA. After overnight
coating, wells were rinsed and blocked with 10 mg/ml
heat-inactivated BSA as the negative control. Cells were treated
with or without 100 μM DFX for 48 h, harvested, resuspended in
serum-free medium, and 5×104 live cells were seeded into
single wells of a 96-well plate. Cells were then incubated for 2 h
at 37°C. Subsequently, the detached cells were washed away, and
attached cells were determine by MTT using a microplate reader.
Migration, invasion and wound healing
assays
Cell migration and invasion assays were performed
using Transwell inserts (BD Biosciences, Bedford, MA, USA) in a
similar manner. The difference was that the upper surface of the
chambers in the invasion assay was pre-coated with Matrigel (BD
Biosciences), while not in the migration assay. Standard protocol
was followed. In brief, cells were pre-treated with or without 100
μM DFX for 24 h. Then cells were digested, washed and seeded into
the inner chamber (5,000 cells/chamber) in medium without FBS.
RPMI-1640 plus 15% FBS was added to the lower chamber, and the
plate was incubated for 24 h. Cells on the upper surface of the
filter were scraped away with a rubber scraper, while cells on the
lower surface were fixed with 4% paraformaldehyde and stained with
0.5% crystal violet, and then counted under a light microscope. For
the wound healing assay, DFX-treated cells were seeded in 6-well
plates. After scratching the monolayer, cells were washed with PBS,
cultured in RPMI-1640 plus 10% FBS, and photographed under a ×10
objective len at the indicated time points. All of the above assays
were carried out in duplicate, and the results presented are the
means of 5 random fields from each well.
Cell growth in soft agar
Cells that were treated with or without DFX (100 μM)
were harvested and pipetted well to become a single-cell
suspension, and then mixed with regular medium containing 0.7% agar
resulting in a final agar concentration of 0.35%. The above cell
suspensions were immediately plated in 6-well plates coated with
0.6% agar in regular medium and cultured for 14 days. Tumor cell
colonies were observed and counted under a dissecting
microscope.
Statistical methods
Statistical analysis was carried out using SPSS 13.0
for Windows (SPSS, Inc.). Quantitative variables were analyzed
using the t-test. P<0.05 was considered to indicate a
statistically significant result.
Results
DFX treatment induces HIF-1α
expression
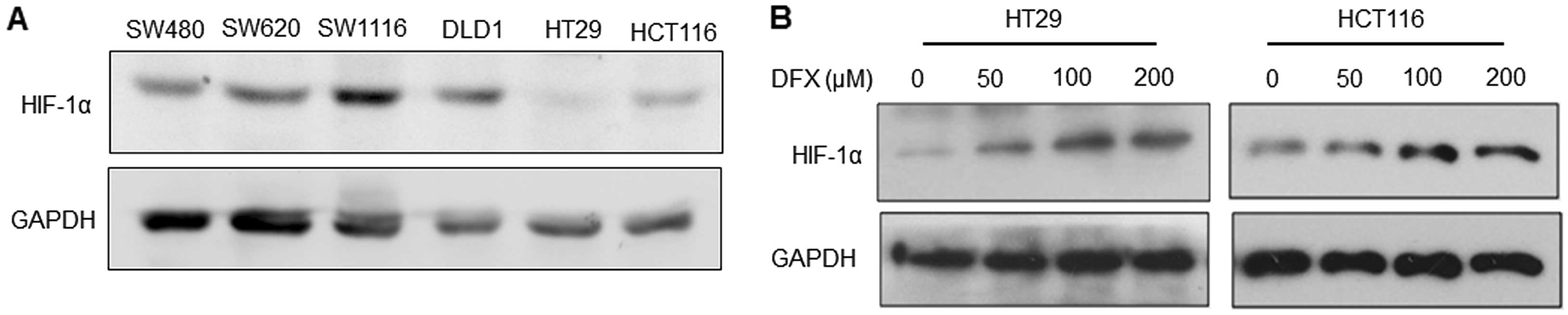
To select a suitable cell candidate to induce HIF-1α
expression and EMT, we first detected the endogenous expression
level of HIF-1α in 6 colorectal cancer cell lines. As shown in
Fig. 1A, the level was much lower
in the HT29 and HCT116 cells when compared with that in the SW480,
SW620, SW1116 and DLD1 cells. Therefore, HT29 and HCT116 cells were
applied in our further experiments.
Next, we incubated HT29 and HCT116 cells with DFX at
doses of 0, 50, 100 and 200 μM, respectively, for 48 h, to
determine the maximum efficacy at inducing EMT in both cell lines.
Treatment with DFX resulted in marked HIF-1α expression in a
dose-dependent manner at the range of 0 to 100 μM as shown in
Fig. 1B. No significant difference
between cells was noted at 100 and 200 μM. Thus, the concentration
of 100 μM was used in the following experiments.
DFX treatment enhances cell adhesion,
migration and invasion abilities
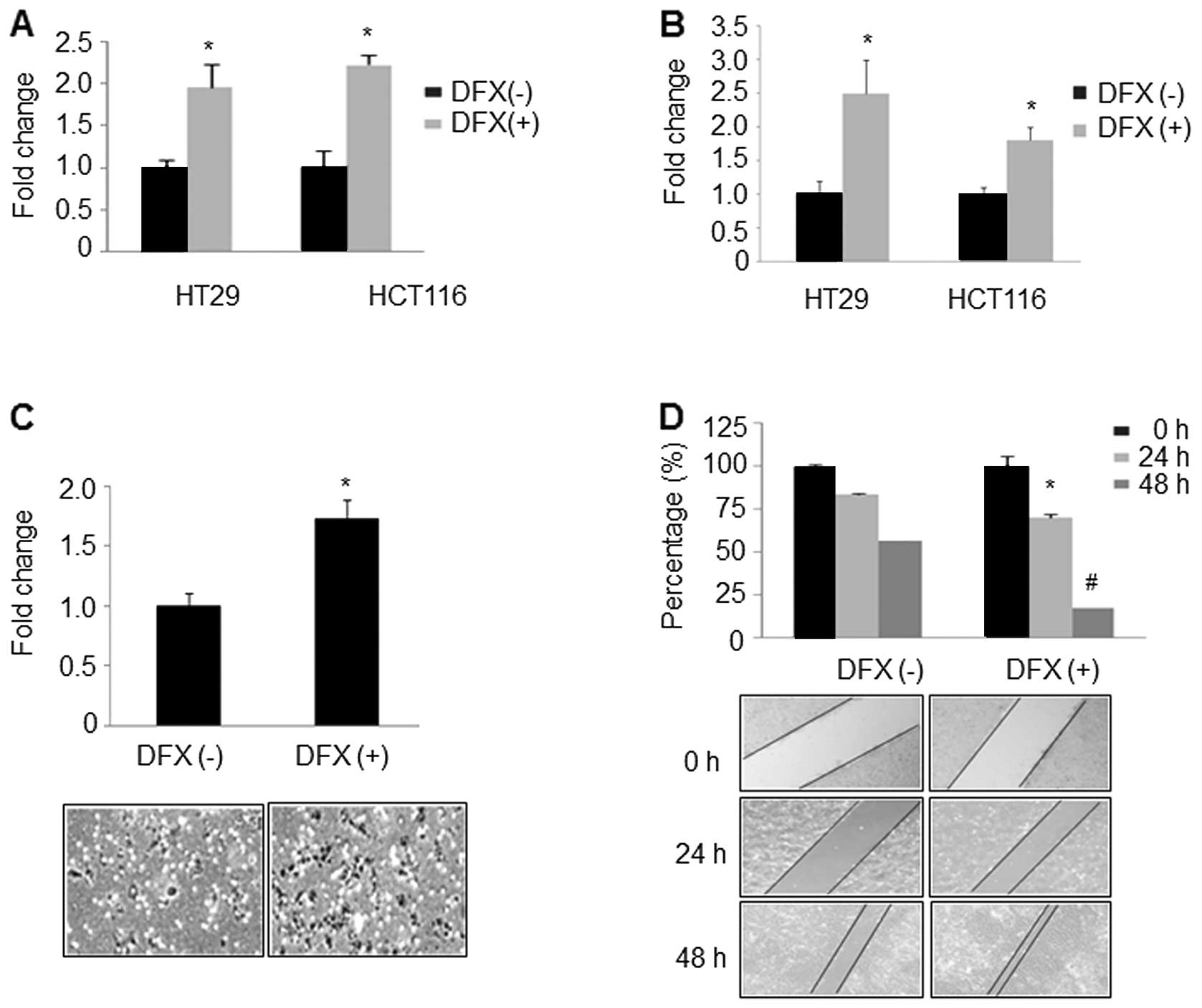
Considering that the adhesion of cancer cells to the
extracellular matrix (ECM) is the first step in tumor metastasis
(24,25), we first evaluated the effect of DFX
on cell adhesion. HT29 and HCT116 cells with or without DFX
treatment were incubated with one major ECM component in
fibronectin-coated 96-well plates for 2 h. We found that DFX
treatment increased cancer cell adhesion to fibronectin by 1.96-
and 2.23-fold in HT29 and HCT116 cells, respectively, when compared
to the cells without DFX treatment (Fig. 2A). Next, we detected the effect of
DFX on cell migration and invasion. As shown in Fig. 2B and C, DFX significantly increased
migration and invasion of HT29 (2.5±0.5 vs. 1±0.2; P<0.05) and
HCT116 cells (1.8±0.2 vs. 1±0.1, P<0.05), when compared with the
untreated control cells. Similarly, the wound healing assay also
indicated that DFX significantly induced cell migration at 24 and
48 h, respectively (Fig. 2D,
P<0.05). Collectively, these observations indicate that DFX
induced cell adhesion, migration and invasion abilities in HT29 and
HCT116 CRC cells.
DFX treatment results in altered cell
morphology and cell growth
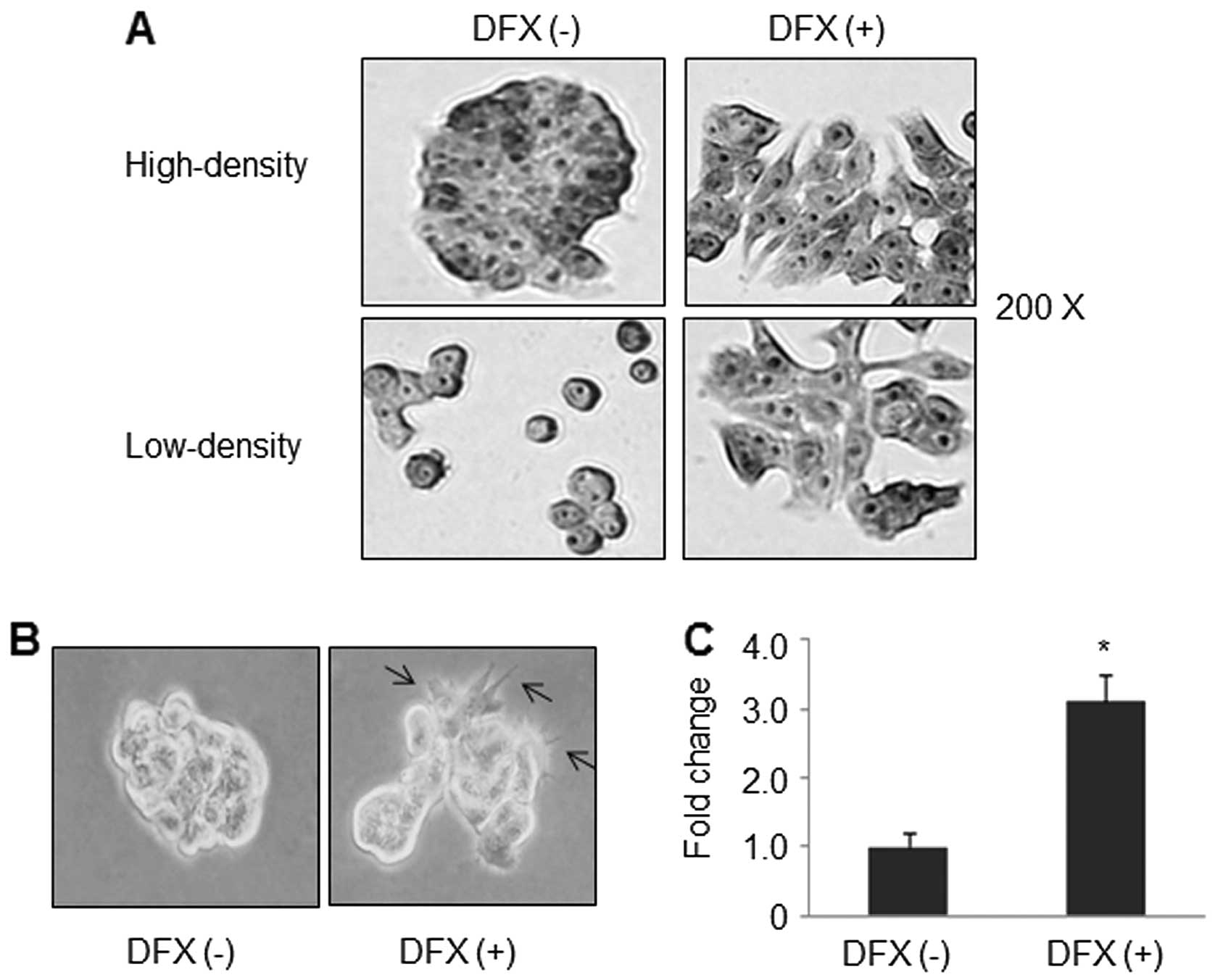
Morphological change plays an important role in many
cellular processes such as migration, differentiation and apoptosis
(22,23,26).
We then investigated whether the increased motility and invasive
ability of CRC cells was coupled with any morphological change.
H&E staining results revealed that treatment with DFX resulted
in dramatic morphological alterations in HT29 cells at both
high-density and low-density as shown in Fig. 3A. In fact, cells treated with DFX
displayed flattened, spread morphology. Cells became more isolated
and looked like spindle-shaped and fibroblast-like cells. In
contrast the untreated HT29 cells were much more round and had very
tight connections. Similar results were found in soft agar assay
experiments. As shown in Fig. 3B,
cell-cell contacts through the ‘pseudopod’ were present in
DFX-treated cells. The edges of these cells commonly showed several
finger-like protrusions which may aid in cell motility. In
contrast, cells without DFX treatment had a clear boundary and
resembled scattered spheres. At the same time, DFX significantly
increased anchorage-independent cell growth by 3.1-fold (Fig. 3C).
Effect of DFX on EMT markers
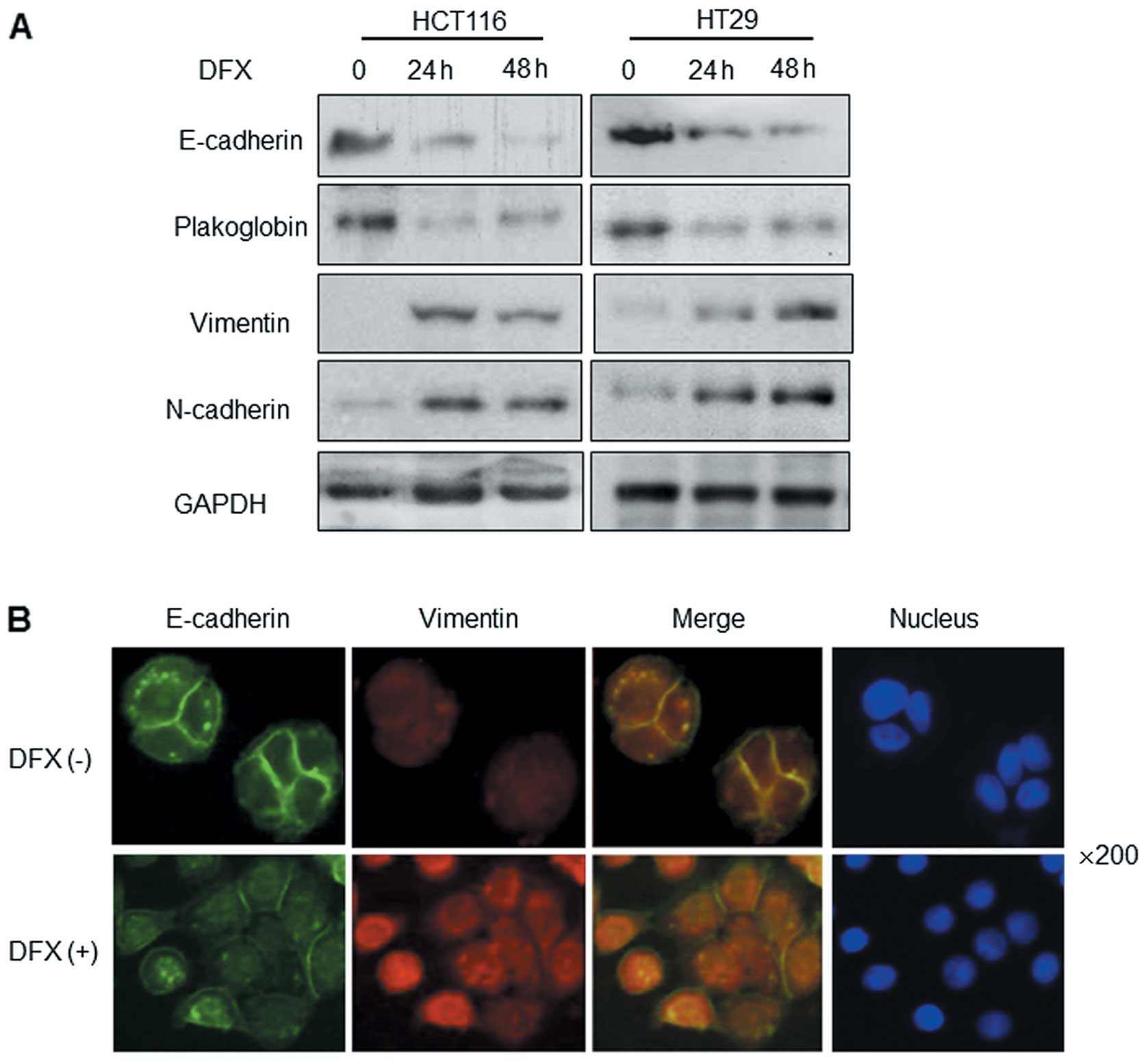
In addition to alterations in the functions
(adhesion, migration and invasion) and cell morphology, the altered
expression of EMT markers is another typical feature of EMT and
cancer metastasis. We evaluated the effect of DFX on EMT epithelial
markers (E-cadherin and plakoglobin) and mesenchymal markers
(vimentin and N-cadherin). Western blot analysis indicated that
E-cadherin and plakoglobin expression was significantly decreased,
while vimentin and N-cadherin expression was increased after DFX
treatment for 24 or 48 h (Fig. 4A).
As a critical adhesion molecular, E-cadherin is reported as the
most important epithelial marker of EMT. We then observed the
expression and localization of E-cadherin and vimentin in cells
with or without DFX treatment by immunofluorescence staining. As
shown in Fig. 4B, the expression of
E-cadherin in the cell membrane was markedly decreased in the
DFX-treated HT29 cells, which indicated lower cell adhesion
ability. At the same time, vimentin expression was upregulated.
Furthermore, DFX-treated cells were able to spread much wider on
coverslips which was consistent with the H&E staining results
(Fig. 3A). These findings suggest
that DFX treatment promotes EMT progression in CRC.
Discussion
Accumulating evidence indicates that hypoxia or
overexpression of HIF-1α are linked to the genesis, progression and
metastasis of human cancers, suggesting HIF-1α as an emerging
biomarker for cancer prognosis and as a promising therapeutic
target (27–29). Using a DFX-induced hypoxia model, we
revealed that DFX promoted HIF-1α expression and enhanced cell
adhesion, migration and invasion abilities in CRC cells, through
modulating cell morphology and expression of EMT markers.
Hypoxia is a major pathophysiological condition for
the induction of angiogenesis, which is a common and crucial aspect
of cell growth in solid tumors (10–12,30).
DFX-induced hypoxia is a complicated event, which may affect a
variety of cellular pathways and gene expression. Particularly, DFX
is capable of inducing the activation of HIF-1α and NF-κB (31). HIF-1α represents the most important
transcription factor regulating gene expression under hypoxic
conditions. It acts by binding to hypoxia-responsive elements
(HREs) in promoters, thereby regulating gene expression and playing
a central role in cancer tumors. Yang et al(20) reported that HIF-1α promotes tumor
progression, EMT and metastasis by direct regulation of Twist, a
key transcriptional regulator of EMT. In the present study, we
found that DFX increased the HIF-1α protein level in a
dose-dependent manner, which may be one of the mechanisms of
DFX-induced cell migration and invasion.
As an important adhesion molecule, membrane
E-cadherin is an important hallmark of EMT (15,19,32).
We demonstrated that incubation of HT29 cells with DFX led to the
loss of E-cadherin in the cell membrane. Furthermore, the
expression of EMT mesenchymal marker, vimentin, was increased
following DFX treatment, which are both consistent with the EMT
event.
In conclusion, the present study demonstrated that
DFX increased HIF-1α protein, decreased membrane-associated
E-cadherin expression and increased vimentin expression.
Consequently, less E-cadherin combines with other adhesion
molecules on the membrane to decrease cell-cell contact and
increase migration and invasion. At the same time, due to the
transition from an epithelial-like to a mesenchymal-like cell
morphology, cell motility was facilitated. These findings explain,
in part, the marked role of DFX in colorectal cancer
metastasis.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (nos. 81172057 and 81272761),
the ‘President Foundation of Nanfang Hospital, Southern Medical
University’ (2012B009), and a ‘high-level topic matching funds’ of
Nanfang Hospital (G201227).
References
|
1
|
Kalinowski DS and Richardson DR: The
evolution of iron chelators for the treatment of iron overload
disease and cancer. Pharmacol Rev. 57:547–583. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mantovani A, Evans R and Alexander P:
Non-specific cytotoxicity of spleen cells in mice bearing
transplanted chemically induced fibrosarcomas. Br J Cancer.
36:35–40. 1977. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Boult J, Roberts K, Brookes MJ, et al:
Overexpression of cellular iron import proteins is associated with
malignant progression of esophageal adenocarcinoma. Clin Cancer
Res. 14:379–387. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Miller LD, Coffman LG, Chou JW, et al: An
iron regulatory gene signature predicts outcome in breast cancer.
Cancer Res. 71:6728–6737. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Follézou JY and Bizon M: Cancer
chemotherapy induces a transient increase of serum-iron level.
Neoplasma. 33:225–231. 1986.PubMed/NCBI
|
|
6
|
Yu Y, Gutierrez E, Kovacevic Z, et al:
Iron chelators for the treatment of cancer. Curr Med Chem.
19:2689–2702. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Recalcati S, Locati M, Gammella E,
Invernizzi P and Cairo G: Iron levels in polarized macrophages:
regulation of immunity and autoimmunity. Autoimmun Rev. 11:883–889.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kaczmarek M, Cachau RE, Topol IA, Kasprzak
KS, Ghio A and Salnikow K: Metal ions stimulated iron oxidation in
hydroxylases facilitates stabilization of HIF-1α protein. Toxicol
Sci. 107:394–403. 2009.PubMed/NCBI
|
|
9
|
Martínez-Romero R, Martínez-Lara E,
Aguilar-Quesada R, Peralta A, Oliver FJ and Siles E: PARP-1
modulates deferoxamine-induced HIF-1α accumulation through the
regulation of nitric oxide and oxidative stress. J Cell Biochem.
104:2248–2260. 2008.PubMed/NCBI
|
|
10
|
Yang Y, Sun M, Wang L and Jiao B: HIFs,
angiogenesis, and cancer. J Cell Biochem. 114:967–974. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Simiantonaki N, Taxeidis M, Jayasinghe C,
Kurzik-Dumke U and Kirkpatrick C: Hypoxia-inducible factor 1 alpha
expression increases during colorectal carcinogenesis and tumor
progression. BMC Cancer. 8:3202008. View Article : Google Scholar
|
|
12
|
Semenza GL: Hypoxia, clonal selection, and
the role of HIF-1 in tumor progression. Crit Rev Biochem Mol Biol.
35:71–103. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Vincent-Salomon A and Thiery JP: Host
microenvironment in breast cancer development:
epithelial-mesenchymal transition in breast cancer development.
Breast Cancer Res. 5:101–106. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang W, Jiang B, Guo Z, et al:
Four-and-a-half LIM protein 2 promotes invasive potential and
epithelial-mesenchymal transition in colon cancer. Carcinogenesis.
31:1220–1229. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang W, Wang J, Zou B, et al: Four and a
half LIM protein 2 (FHL2) negatively regulates the transcription of
E-cadherin through interaction with Snail1. Eur J Cancer.
47:121–130. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kahlert UD, Nikkhah G and Maciaczyk J:
Epithelial-to-mesenchymal(-like) transition as a relevant molecular
event in malignant gliomas. Cancer Lett. 331:131–138. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Roy N, Bommi PV, Bhat UG, et al: DDB2
suppresses epithelial-to-mesenchymal transition in colon cancer.
Cancer Res. 73:3771–3782. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Higgins DF, Kimura K, Bernhardt WM, et al:
Hypoxia promotes fibrogenesis in vivo via HIF-1 stimulation of
epithelial-to-mesenchymal transition. J Clin Invest. 117:3810–3820.
2007.PubMed/NCBI
|
|
19
|
Jiang J, Tang YL and Liang XH: EMT: a new
vision of hypoxia promoting cancer progression. Cancer Biol Ther.
11:714–723. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yang MH, Wu MZ, Chiou SH, et al: Direct
regulation of TWIST by HIF-1α promotes metastasis. Nat Cell Biol.
10:295–305. 2008.
|
|
21
|
Zhang Q, Bai X, Chen W, et al:
Wnt/β-catenin signaling enhances hypoxia-induced
epithelial-mesenchymal transition in hepatocellular carcinoma via
crosstalk with hif-1α signaling. Carcinogenesis. 34:962–973.
2013.
|
|
22
|
Wang J, Yang Y, Xia HH, et al: Suppression
of FHL2 expression induces cell differentiation and inhibits
gastric and colon carcinogenesis. Gastroenterology. 132:1066–1076.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hollestelle A, Peeters JK, Smid M, et al:
Loss of E-cadherin is not a necessity for epithelial to mesenchymal
transition in human breast cancer. Breast Cancer Res Treat.
138:47–57. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lester BR and McCarthy JB: Tumor cell
adhesion to the extracellular matrix and signal transduction
mechanisms implicated in tumor cell motility, invasion and
metastasis. Cancer Metastasis Rev. 11:31–44. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bartolazzi A, Cerboni C, Nicotra MR,
Mottolese M, Bigotti A and Natali PG: Transformation and tumor
progression are frequently associated with expression of the alpha
3/beta 1 heterodimer in solid tumors. Int J Cancer. 58:488–491.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wei L, Yang Y, Zhang X and Yu Q: Altered
regulation of Src upon cell detachment protects human lung
adenocarcinoma cells from anoikis. Oncogene. 23:9052–9061. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zheng Y, Ni Y, Huang X, Wang Z and Han W:
Overexpression of HIF-1α indicates a poor prognosis in tongue
carcinoma and may be associated with tumour metastasis. Oncol Lett.
5:1285–1289. 2013.
|
|
28
|
Cheli Y, Giuliano S, Fenouille N, et al:
Hypoxia and MITF control metastatic behaviour in mouse and human
melanoma cells. Oncogene. 31:2461–2470. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Martinez-Outschoorn UE, Trimmer C, Lin Z,
et al: Autophagy in cancer associated fibroblasts promotes tumor
cell survival: role of hypoxia, HIF1 induction and NFκB activation
in the tumor stromal microenvironment. Cell Cycle. 9:3515–3533.
2010.PubMed/NCBI
|
|
30
|
Tanaka T and Nangaku M: Angiogenesis and
hypoxia in the kidney. Nat Rev Nephrol. 9:211–222. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Haddad JJ, Olver RE and Land SC:
Antioxidant/pro-oxidant equilibrium regulates HIF-1α and NF-κB
redox sensitivity. Evidence for inhibition by glutathione oxidation
in alveolar epithelial cells. J Biol Chem. 275:21130–21139.
2000.PubMed/NCBI
|
|
32
|
Schmalhofer O, Brabletz S and Brabletz T:
E-cadherin, β-catenin, and ZEB1 in malignant progression of cancer.
Cancer Metastasis Rev. 28:151–166. 2009.
|