Introduction
Paclitaxel is a plant alkaloid that was developed
from the bark of the Pacific yew tree, Taxus
brevifolia(1). Paclitaxel is a
taxane that stabilizes and disrupts microtubules required for cell
division, resulting in cell death (2,3).
Despite extensive clinical use in the treatment of patients with
lung, ovarian and breast cancer, long-term survival is frequently
compromised by the development of paclitaxel resistance, for which
the molecular basis remains to be fully delineated. microRNAs
(miRNAs) are non-coding, 21–25 nucleotide regulatory RNAs that
affect the stability and/or translational efficiency of messenger
RNA (mRNAs) (4). Thousands of
miRNAs are predicted to exist in the human genome (5) of which 1,100 human miRNAs have been
identified, collectively targeting more than 19,000 human genes
(www.microRNA.org). Deregulation of miRNAs has been
implicated in the development of many types of human cancers
(6,7), suggesting that some miRNAs function as
oncogenes or tumor suppressors (8,9). It
has been reported that loss of let7 may influence the
development of lung cancer as it negatively regulates let60/RAS
(10), whereas miRs-34a-c may play
an important role in the tumor-suppressor function of p53 (11,12)
and miR-181a was found to be related to a morphological subclass of
acute myeloid leukemia (13). Some
studies have suggested that miRNAs may also influence
chemosensitivity (14–16). It has been shown that miR-221/222
overexpression reduces p27Kip1 levels and induces
tamoxifen resistance due to cell cycle inhibition (17), whereas inhibition of miR-21
increases apoptosis in lung adenocarcinoma epithelial cell line
A549 after NSC 265450 (nogamycin) and NSC 670550 treatment by
downregulating Bcl2 protein (14).
In the present study, we integrated miRNA data for
lung, colon, breast, ovarian, kidney, skin (melanoma), prostate,
central nervous system (CNS), and hematologic (leukemia) cancer
cell lines with GI50 paclitaxel-sensitivity data in an
effort to identify miRNAs associated with paclitaxel response.
Furthermore, we evaluated the effect of in vitro targeted
modulation of these miRNAs on paclitaxel sensitivity.
Materials and methods
Cell culture and reagents
A subset of 40 of the NCI60 cancer cell line panel
was obtained from the National Cancer Institute (NCI) Developmental
Therapeutics Program (Table I).
Ovarian cancer (OVCA) cell lines in addition to those on the NCI60
panel were obtained from the American Type Culture Collection
(ATCC, Manassas, VA, USA; CAOV3, OV90, OVCAR3 and SKOV3), the
European Collection of Cell Cultures (Salisbury, UK; A2780CP and
A2780S), Kyoto University (Kyoto, Japan; M41, M41CSR, Tyknu, and
TyknuCisR), or as kind gifts from Dr Patricia Kruk, Department of
Pathology, College of Medicine, University of South Florida, Tampa,
FL, and Susan Murphy, Department of OBGYN/Division of Gynecologic
Oncology, Duke University, Durham, NC (HeyA8, IGR-OV1, IMCC3,
IMCC5, MCAS, OVCA420, OVCA429, OVCA432, OVCA433, FUOV1, PA1, PEO1,
PEO4, T8, TOV-112D, TOV-21-G, Dov13, OVCAR10, OVCAR8, OVCAR5,
OVCAR4 and OVCAR2). Human stem cell lines (H9) were obtained from
WiCell (Madison, WI, USA). All cancer cell lines were cultured in
RPMI-1640 medium supplemented with 1% non-essential amino acids, 1%
sodium pyruvate and 10% fetal bovine serum. H9 cells were cultured
according to the manufacturer’s protocol. Cells were cultured for
2–3 passages before experimentation. Paclitaxel was obtained from
Sequoia Research Products Ltd., (Oxfordshire, UK), dissolved in
DMSO at a concentration of 100 mM and stored at −20°C.
 | Table ICancer cell lines subjected to miRNA
expression analyses. |
Table I
Cancer cell lines subjected to miRNA
expression analyses.
| Origin of cancer
tissue | Cell lines |
|---|
| Lung | NCI-H460, NCI-H522,
NCI-H322M, HOP62, A549, EKVX, MALME-3M, NCI-H226 |
| Colon | HT29, HCT-116,
SE-620, HCT-15, HCC2998, COLO205 |
| Breast | HS-578T,
NCI/ADR-RES |
| Ovarian | OVCAR8, OVCAR4 |
| Renal | ACHN, SN-12C,
786-O, CAKI-1, UO-31, TK-10, A498 |
| Melanoma | SK-MEL-28,
UACC-257, M14, UACC-62, SK-MEL-2, LOX-IMVI |
| Prostate | DU-145, PC-3 |
| CNS | SF-295, SF-539,
SNB-75, U251 |
| Leukemia | HL-60, RPMI8226,
K562 |
miRNA extraction and expression
profiling
Total RNA was extracted from 1×106
log-phase cells using the mirVana miRNA isolation kit (Life
Technologies) according to the manufacturer’s instructions. The
yield and quality of total RNA for each cell line were determined
using an Agilent Bioanalyzer. The RNA integrity number (RIN) of all
samples was in a range of 8–10 and no genomic DNA contamination was
detected. Total RNA (10 μg) from each cell line was then subjected
to miRNA expression profiling. The cell line samples were
co-hybridized to printed arrays that contained a 562 Ambion mirVana
miRNA probe set (Ambion, Austin TX, USA) and 632 of Invitrogen’s
NCode Multi-Species miRNA probes (Invitrogen, Carlsbad, CA, USA),
which contain >800 unique human miRNAs. The hybridized arrays
were scanned on a GenePix 4000B scanner, and expression data were
generated using the GenePix Pro software (Molecular Devices,
Sunnyvale, CA, USA).
Transfection of miRNA
Cell lines were transfected with pre-miR-367,
pre-miR-30a-5p, anti-miR-367 or anti-miR-30a-5p (Life Technologies)
using siPORT NeoFX transfection reagent, according to the
manufacturer’s protocol. The concentrations of miRNA and the
transfection reagent were optimized by the manufacturer’s
recommendations prior to experimentation. Log-phase cells (~60%
confluence) were incubated with 6.25 μM precursor or inhibitor
miRNAs in the presence of 50 nM transfection reagent for 4 h.
pre-miR negative control 1 precursor and anti-miR negative control
1 were used as controls for miRNA overexpression and depletion
experiments, respectively. Control miRNAs were transfected under
the same condition as the experimental miRNAs. Both control miRNAs
have mechanisms similar to the pre-miRNA precursors and anti-miRNA
inhibitors but have no observed effects on known miRNA
functions.
Comparative CT RT-PCR
Real-time comparative CT RT-PCR was used
to determine relative miRNA levels. miRNAs were converted into cDNA
using miRNA sequence-specific primers and the TaqMan®
microRNA reverse transcription (Applied Biosystems). Total RNA (10
ng) was used for each 15-μl reverse transcription reaction.
Comparative CT RT-PCR was performed on the Applied
Biosystems StepOne Real-Time PCR system, according to the
manufacturer’s protocols. Briefly, 1.33 μl of miRNA-specific cDNA
was combined with sequence-specific TaqMan miRNA assays and TaqMan
Universal PCR Master Mix, no AmpErase UNG in a total reaction
volume of 20 μl. RNU44 was used as the endogenous miRNA control for
normalization. RNU44 was considered a qualified candidate of
endogenous control based on preliminary experimentation (data not
shown). miRNA expression levels were analyzed using StepOne
Real-Time PCR instrument software.
Growth inhibition assay
Cells were seeded in 96-well plates (Perkin-Elmer)
at a density of 6×104 cells/ml and incubated overnight
at 37°C. Cells were incubated with the indicated concentrations of
paclitaxel for 72 h, and cell viability was assessed using the
CellTiter-Glo™ luminescent cell viability assay kit (MTS kit;
Promega). Luminescence was recorded using Wallace Victor2™ 1420
Multilabel Counter (Perkin-Elmer). Wells containing medium without
cells were used to obtain background luminescence. All experimental
wells and controls were set up in triplet. Paclitaxel
log10 (IC50) values for transfected cell
lines were calculated using the sigmoidal dose-response (variable
slope) curve equation (Prism 5). Dose-curve graphs were generated
using GraphPad (Prism 5). Cells cultured with transfected medium
without miRNA precursors or inhibitors were used as controls. The
effects of increased pre-miR and anti-miR levels due to
transfection on cell viability were assessed 48 h post-transfection
using MTS assays.
Statistical analysis
Two color spotted array data for miRNA levels were
generated from the GenePix Pro software. Background subtraction
(18) and Loess normalization
(19) were performed using Limma
software. Replicate probe sets were averaged by their design
(Ambion or Invitrogen). Processed data were then analyzed using SAM
(significance analysis of microarrays) software (20). Missing values were imputed with
SAM’s nearest neighbor imputer, where k was set to 10. For
each drug, Pearson’s correlation test was used to identify those
miRNAs with expression values associated with sensitivity measured
by GI50.
Pathway analysis
The miRanda database was used to identify the mRNA
targets of miRNAs found to be associated with in vitro
sensitivity to chemotherapy. The identified mRNA targets were
subjected to GeneGo MetaCore analysis to determine biological
signaling pathway representation. P<0.05 represented statistical
significance of the association between the mRNA targets of the
miRNAs and the biological pathways.
Results
Correlation of miRNA expression and
paclitaxel sensitivity/resistance
Paclitaxel sensitivity (GI50) data for
the subset of 40 cancer cell lines of the NCI60 cell panel (3
leukemia, 6 melanoma, 8 non-small cell lung, 6 colon, 4 central
nervous system, 2 ovarian, 7 renal, 2 prostate and 2 breast cancer
cell lines) was obtained from NCI Website (http://dtp.nci.nih.gov/dtpstandard/cancerscreeningdata/index.jsp).
Based on miRNA expression and GI50 data, Pearson’s
correlation test identified 35 miRNAs associated with in
vitro paclitaxel sensitivity (P<0.05). miR-367 and
miR-30a-5p demonstrated the highest level of statistical
significance in association with sensitivity to paclitaxel
(P<0.0003) (Table II).
| Table IImicroRNAs associated with paclitaxel
sensitivity. |
Table II
microRNAs associated with paclitaxel
sensitivity.
| miRNA | P-value | Up/down |
|---|
| hsa_miR_367 | 0.00022 | Down |
| hsa_miR_30a_5p | 0.00028 | Up |
| hsa_miR_141 | 0.00115 | Down |
| hsa_miR_30a_3p | 0.00306 | Up |
| hsa_miR_516_3p | 0.00318 | Up |
| hsa_miR_377 | 0.00350 | Down |
| hsa_miR_134 | 0.00371 | Up |
| hsa_miR_142_5p | 0.00767 | Down |
| hsa_let_7e | 0.01000 | Up |
| hsa_miR_29c | 0.01017 | Down |
| hsa_miR_218 | 0.01105 | Down |
| hsa_miR_17_3p | 0.01123 | Down |
| hsa_miR_17_5p | 0.01288 | Down |
| hsa_miR_130a | 0.01289 | Up |
| hsa_miR_195 | 0.01939 | Down |
| hsa_miR_99b | 0.02003 | Up |
| hsa_miR_338 | 0.02508 | Up |
| hsa_miR_106a | 0.02611 | Down |
| hsa_miR_193b | 0.02720 | Up |
| hsa_miR_515_3p | 0.02884 | Up |
| hsa_miR_374 | 0.02889 | Up |
| hsa_miR_125a | 0.02993 | Up |
| hsa_miR_192 | 0.03023 | Down |
| hsa_miR_30c | 0.03039 | Up |
| hsa_miR_95 | 0.03684 | Down |
| hsa_miR_452_AS | 0.03703 | Down |
| hsa_miR_489 | 0.03774 | Up |
| hsa_miR_32 | 0.03897 | Down |
| hsa_miR_373* | 0.03938 | Up |
| hsa_miR_130b | 0.04136 | Down |
| hsa_miR_19b | 0.04385 | Down |
| hsa_miR_126 | 0.04543 | Down |
| hsa_miR_148a | 0.04650 | Down |
| hsa_miR_376b | 0.04835 | Up |
| hsa_miR_7 | 0.04835 | Up |
Selection of cell lines
The OVCA cell lines PA1 and OVCAR4 were selected for
further evaluation based on associations between paclitaxel
sensitivity and miR-367/miR-30a-5p expression. An evaluation of the
OVCA cell lines showed PA1 to have a higher relative expression
value of miR-367 (2.997), a lower relative expression value of
miR-30a-5p (−0.32), and relative sensitivity to paclitaxel-induced
cell growth arrest (IC50, 1.69 nM). In contrast, OVCAR4
showed a lower relative expression value of miR-367 (−0.64), a
higher relative expression values of miR-30a-5p (3.27), and was
more resistant to paclitaxel (IC50, 17.8 nM). The
differential expression of miR-367 and miR-30a-5p in PA1 and OVCAR4
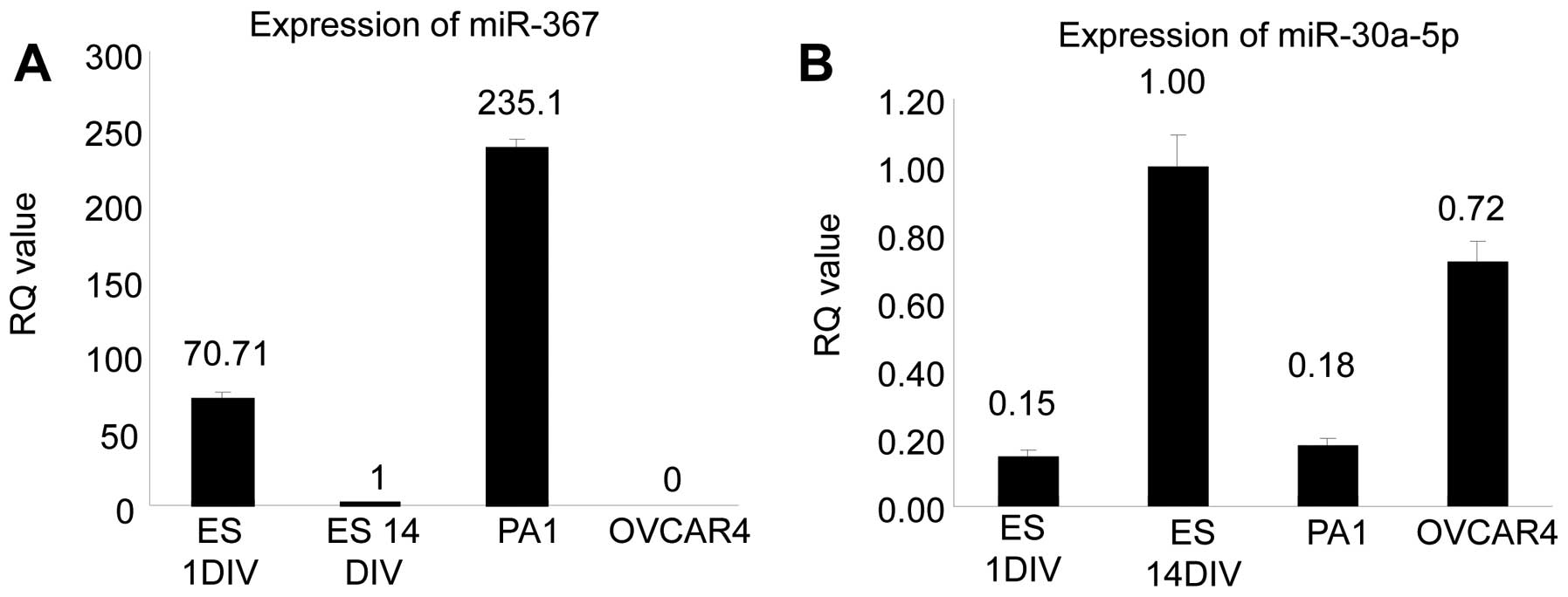
cells was confirmed by quantitative RT-PCR (Fig. 1). As a reference, miRNA expression
was compared to embryonic stem cells, which reportedly express
miR-367 in the early stage and reduced miR-367 expression in the
late stage (21). Embryonic stems
cells were defined as early vs. late stage based on culture
duration of 1 (ES1DIV) vs. 14 (ES14DIV) days, respectively. As
shown in Fig. 1A, using ES14DIV as
the reference [relative expression (RQ)=1], the relative expression
(RQ value) of miR-367 PA1 cells was 235.1, compared to RQ=70.71 for
ES1DIV (positive control) and RQ=0 for OVCAR4 cells. In contrast,
normalized to ES14DIV (RQ=1), OVCAR4 had the highest expression of
miR-30a-5p with an RQ=0.72, compared to RQ=0.18 for PA1 cells
(Fig. 1B).
Evaluation of miR-367 and miR-30a-5p as
therapeutic targets
To determine the value of miR-367 and miR-30a-5p as
therapeutic targets in OVCA cells, miR-367 was overexpressed or
depleted in the paclitaxel-resistant cell line PA1 (high miR-367,
low miR-30a-5p), through transient transfection of the miRNA
precursor (pre-miR-367) and inhibitor (anti-miR-367), respectively.
In contrast, the paclitaxel-sensitive OVCA cell line OVCAR4 (low
miR-367, high miR-30a-5p) was transfected with the miR-30a-5p
precursor (pre-miR-30a-5p) and inhibitor (anti-miR-30a-5p).
Forty-eight hours after transfection, cells were incubated with
increasing doses of paclitaxel for 72 h and evaluated for cell
viability using the CellTiter-Glo™ luminescent assay.
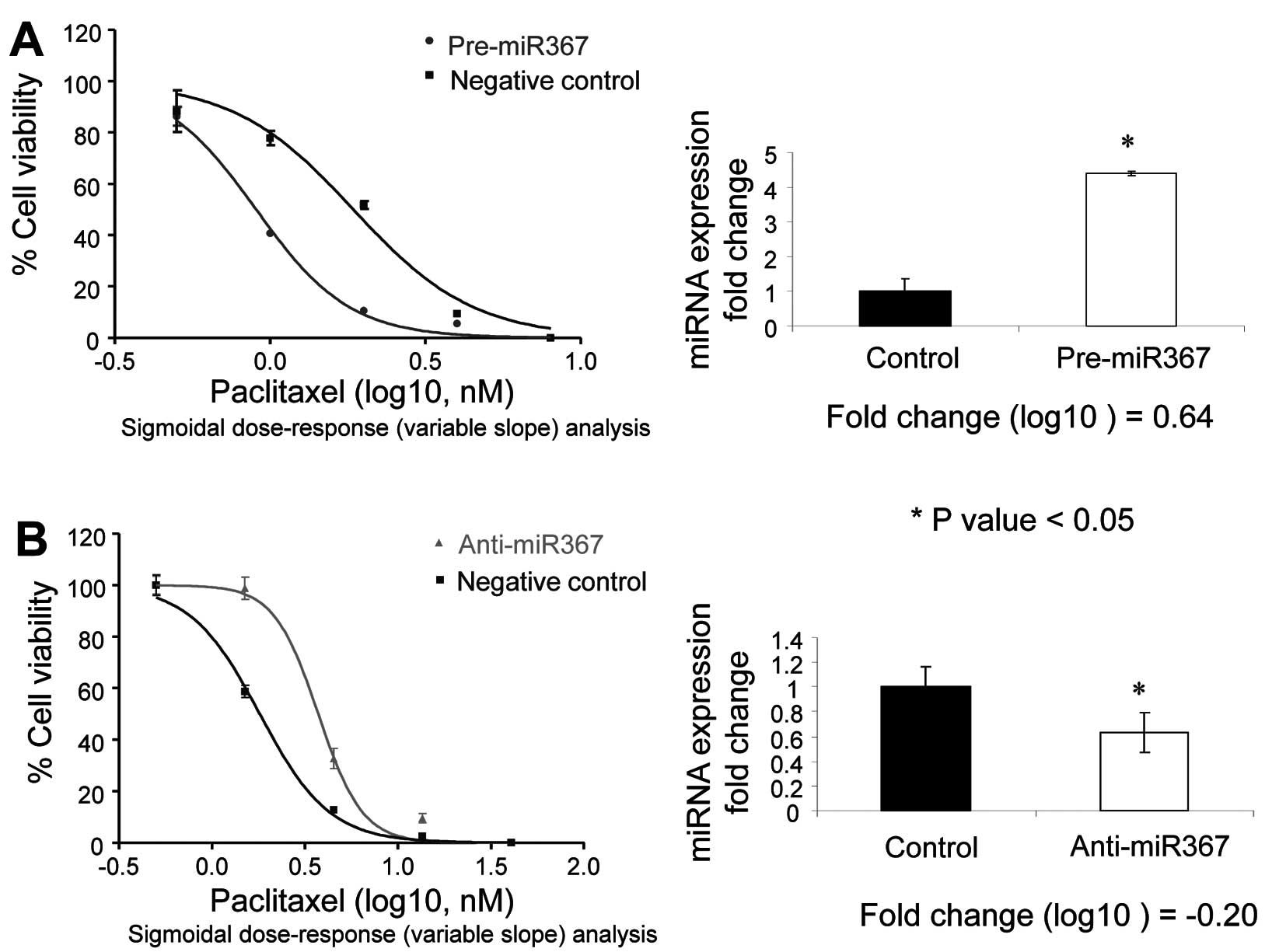
Compared to the mock transfection controls, PA1
cells transfected with pre-miR-367 showed a 57% decrease in cell
survivability 48 h after transfection, whereas transfection of
anti-miR-367 did not affect cell survival. Transfection of
pre-miR-30a-5p and anti-miR-30a-5p had no effect on the
survivability of PA1 cells (data not shown). Despite the decrease
in cell survival, transfection of pre-miR-367 in PA1 cells resulted
in a 0.64 log10-fold-change in miR-367 expression and an
increase in paclitaxel sensitivity when compared to the precursor
negative control (Fig. 2A). In
contrast, transfection of PA1 cells with anti-miR-367 resulted in a
−0.2 log10-fold-change and a decrease in paclitaxel
sensitivity (Fig. 2B).
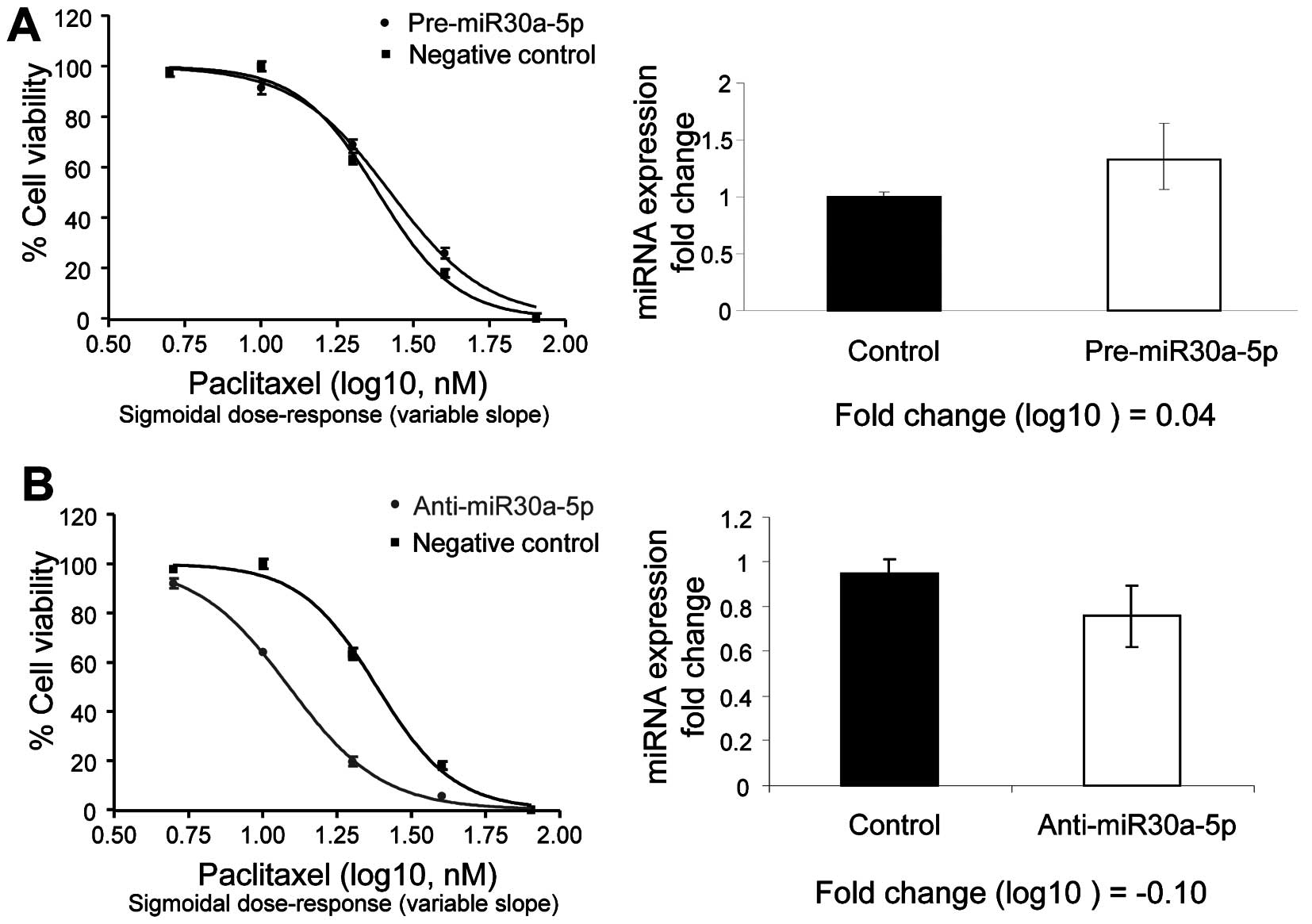
In the intrinsically paclitaxel-resistant cell line
(OVCAR4), which has high relative expression of miR-30a-5p and low
relative expression of miR-367, overexpression of the miR-30a-5p
precursor (0.04 log10-fold change) slightly decreased
paclitaxel-induced sensitivity, whereas depletion of miR-30a-5p
(−0.10 log10-fold change) increased paclitaxel-induced
growth arrest (Fig. 3).
Pathways involved in the deregulation of
miRNAs
To evaluate the potential influence of miR-367 and
miR-30-5p on various cellular processes, we identified the
predicted mRNA target genes of these miRNAs using the miRanda
database (22). This database hosts
target sites for 1,100 human miRNAs and 16,228,619 predicted miRNA
targets in 34,911 distinct 3′UTRs of 19,898 human genes. The
miRanda database predicted 1,536 and 2,320 target genes associated
with miR-367 and miR-30a-5p, respectively (mirSVR score >−0.15,
mirSVR score ranged from −0.1 to −1.35) (20). These predicted miRNA targets were
further analyzed for biologic signaling pathway representation
using GeneGo MetaCore software. Pathway modeling identified 16
pathways represented among the predicted target genes for miR-367
(P<0.0001) and 20 pathways were represented among the target
genes for miR-30a (P<0.0001) (Table III).
| Table IIImiR-367/miR-30a-5p target
gene-involved pathways (P<0.0001). |
Table III
miR-367/miR-30a-5p target
gene-involved pathways (P<0.0001).
| P-value |
Objects/networks |
|---|
| miR-367 target
gene-involved pathways (P<0.0001) | | |
| Signal
transduction_PKA signaling | 1.84E-07 | 14/15 |
| Signal
transduction_cAMP signaling | 2.58E-07 | 12/38 |
|
Development_Thrombopoietin-regulated cell
processes | 2.63E-07 | 13/45 |
| Development_A2A
receptor signaling | 1.16E-06 | 12/43 |
| Immune
response_IL-23 signaling pathway | 2.44E-06 | 9/25 |
|
Development_TGF-β-dependent induction of
EMT via RhoA, PI3K and ILK | 2.54E-06 | 12/46 |
| Signal
transduction_Activation of PKC via G-Protein coupled receptor | 1.01E-05 | 12/52 |
| Neurophysiological
process_Glutamate regulation of Dopamine D1A receptor
signaling | 1.32E-05 | 11/45 |
| Development_Role
of HDAC and calcium/calmodulin-dependent kinase (CaMK) | 1.53E-05 | 12/54 |
| in control of
skeletal myogenesis |
| Development_PACAP
signaling in neural cells | 2.09E-05 | 10/39 |
|
Translation_Insulin regulation of
translation | 4.19E-05 | 10/42 |
| Apoptosis and
survival_BAD phosphorylation | 4.19E-05 | 10/42 |
| Development_IGF-1
receptor signaling | 4.68E-05 | 11/51 |
| Signal
transduction_AKT signaling | 5.22E-05 | 10/43 |
|
Transport_Clathrin-coated vesicle
cycle | 6.08E-05 | 13/71 |
| Neurophysiological
process_ACM regulation of nerve impulse | 9.64E-05 | 10/46 |
| miR30a-5p target
gene-involved pathways (P<0.0001) |
| Cytoskeleton
remodeling_TGF, WNT and cytoskeletal remodeling | 8.72E-08 | 25/111 |
| Cytoskeleton
remodeling_Cytoskeleton remodeling | 2.81E-07 | 23/102 |
| Cell
adhesion_Ephrin signaling | 1.02E-06 | 14/45 |
|
Development_Thrombopoietin-regulated cell
processes | 1.02E-06 | 14/45 |
| Development_HGF
signaling pathway | 1.84E-06 | 14/47 |
| Development_WNT
signaling pathway. Part 2 | 8.82E-06 | 14/53 |
| Muscle
contraction_Regulation of eNOS activity in endothelial cells | 2.05E-05 | 15/64 |
|
Development_Regulation of
epithelial-to-mesenchymal transition (EMT) | 2.05E-05 | 15/64 |
| Apoptosis and
survival_FAS signaling cascades | 2.12E-05 | 12/43 |
|
Development_Membrane-bound ESR1:
interaction with growth factor signaling | 2.12E-05 | 12/43 |
| Cardiac
hypertrophy_NF-AT signaling in cardiac hypertrophy | 2.5E-05 | 15/65 |
| Immune
response_ETV3 affect on CSF1-promoted macrophage
differentiation | 2.58E-05 | 10/31 |
| Development_Role
of IL-8 in angiogenesis | 2.7E-05 | 14/58 |
|
Development_Ligand-independent activation
of ESR1 and ESR2 | 2.73E-05 | 12/44 |
| Cell
adhesion_Chemokines and adhesion | 4.13E-05 | 19/100 |
|
Translation_Regulation of EIF4F
activity | 4.36E-05 | 13/53 |
| Apoptosis and
survival_Caspase cascade | 4.75E-05 | 10/33 |
| Development_PIP3
signaling in cardiac myocytes | 5.59E-05 | 12/47 |
| PGE2 pathways in
cancer | 6.61E-05 | 13/55 |
| DNA damage_Role of
SUMO in p53 regulation | 7.41E-05 | 7/17 |
Discussion
The efficacy of cancer treatment is frequently
limited by intrinsic and acquired resistance to chemotherapy.
Despite progress in delineating the molecular determinants of
cancer chemo-response, a comprehensive understanding of the factors
that underlie drug resistance remains elusive.
Evidence is accumulating to support a role for
miRNAs in the development and progression of human cancer (7,23,24).
Moreover, recent data also suggest that miRNAs may influence cancer
cell response to chemotherapy (14,16,25) by
mechanisms that may be both cancer cell-type or drug specific.
Previous studies have shown that paclitaxel sensitivity may be
associated with the expression of miR-200c in both ovarian
(26) and gastric cancer (27), miR-148 in prostate cancer cells
(28), miR-337, miR-34 and miR-135a
in lung cancer (29–31), miR-22 in colon cancer (32), and miR-125b and miR-21 in breast
cancer (33,34).
In the present study, miRNA expression data
integrated with publicly available chemosensitivity data for 40
human cancer cell lines (representing 9 different cancer cell
types) identified 35 miRNAs to be associated with in vitro
paclitaxel sensitivity (P<0.05). Two of these miRNAs, miR-367
and miR-30a-5p, were selected for further experimentation based on
associations between paclitaxel sensitivity and miR-367/miR-30a-5p
expression. The effects of miR-367 and miR-30a-5p expression on
chemosensitivity were investigated in OVCA cell lines shown to have
differential expression of these miRNAs. The OVCA cell line PA1 was
found to be relatively sensitive to paclitaxel-induced cell death
and have relatively high expression of miR-367 and low expression
of miR-30a-5p. In contrast, OVCAR4 cells were found to have almost
no expression of miR-367 and relatively high expression of
miR-30a-5p and were relatively resistant to paclitaxel. In PA1
cells, the overexpression and depletion of miR-367 increased the
sensitivity and resistance of these cell lines to
paclitaxel-induced growth arrest and cell death, respectively. In
contrast, in OVCAR4 cells, an increase in miR-30a-5p expression was
associated with decreased paclitaxel sensitivity, whereas a
depletion of miR-30a-5p was associated with an increase in
paclitaxel sensitivity. The mechanism by which these miRNAs affect
chemosensitivity was not determined. However, miR-367, which
belongs to the miR302 cluster, is only present in embryonic stem
cells and is significantly decreased after cells differentiate
(21). miR-302 and miR-367 not only
participate in the processes of maintaining cell self-renewal and
pluripotency in embryonic stem cells but are also overexpressed in
various cancer cells (21,35–38)
and may play a role in chemosensitivity (39). Similarly, miR-30a-5p has been
reported to be differentially expressed in various malignancies,
including lung, thyroid, anaplastic and gastric cancer (40–44)
and has been associated with survival of patients with cancer
(45,46).
Bioinformatic analyses of the predicted miR-367
target genes indicated that miR-367 may have an important
regulatory role in the expression or activity of 16 biologic
signaling pathways. Notably, the majority of these
miR-367-influenced pathways, such as signal transduction/PKA
signaling, signal transduction/AKT signaling, cAMP signaling, and
apoptosis and survival/BAD phosphorylation, influence cell survival
through regulating cell cycle and cell apoptosis and maintaining
cell self-renewal and stemness (Table
III), although the role of miR-367 in carcinogenesis and
chemosensitivity is largely unknown.
miR-30a-5p was predicted to influence the expression
of 20 biologic signaling pathways, several of which are also known
to influence cellular survival, such as apoptosis and survival/FAS
signaling cascades and apoptosis and survival/caspase cascade.
However, the majority of pathways under the influence of
miR-30a-5p’s appears to involve cytoskeletal remodeling and cell
migration (Table III).
The present study demonstrated that the integration
of miRNA expression data with existing chemosensitivity data from
the NCI40 cell line set may provide insight into miRNAs that
influence in vitro paclitaxel sensitivity. However, it
should be acknowledged that such an approach may preferentially
identify those miRNAs that were influential in determination of
chemosensitivity across tumor types and may not identify those
miRNAs that have a cancer-specific influence on the response to
paclitaxel. Although data such as these provide an important
contribution to our knowledge of the underpinnings of cancer cell
response to therapeutic agents, it should be recognized that a
comprehensive understanding of the biologic determinants of
chemo-response will ultimately require us to incorporate
information on additional variables such as DNA sequence and copy
number, mRNA expression (vs. predicted mRNA targets), and protein
levels and post-translational modifications. Our data contribute to
the growing body of evidence suggesting that miRNAs have potential
utility as personalized medicine biomarkers of cancer cell response
to therapy and, moreover, may also represent viable therapeutic
targets to increase cancer cell chemosensitivity.
Acknowledgements
This study was supported (in part) by Moffitt/USF
Anna Valentine Fund and the Cancer Center Support Grant P30-
CA76292-14. We thank Rasa Hamilton (Moffitt Cancer Center) for her
editorial assistance.
References
|
1
|
Wani MC, Taylor HL, Wall ME, Coggon P and
McPhail AT: Plant antitumor agents. VI The isolation and structure
of Taxol, a novel antileukemic and antitumor agent from Taxus
brevifolia. J Am Chem Soc. 93:2325–2327. 1971. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Schiff PB and Horwitz SB: Taxol assembles
tubulin in the absence of exogenous guanosine 5′-triphosphate or
microtubule-associated proteins. Biochemistry. 20:3247–3252.
1981.PubMed/NCBI
|
|
3
|
Schiff PB and Horwitz SB: Taxol stabilizes
microtubules in mouse fibroblast cells. Proc Natl Acad Sci USA.
77:1561–1565. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lim LP, Lau NC, Garrett-Engele P, et al:
Microarray analysis shows that some microRNAs downregulate large
numbers of target mRNAs. Nature. 433:769–773. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cummins JM and Velculescu VE: Implications
of micro-RNA profiling for cancer diagnosis. Oncogene.
25:6220–6227. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
He H, Jazdzewski K, Li W, et al: The role
of microRNA genes in papillary thyroid carcinoma. Proc Natl Acad
Sci USA. 102:19075–19080. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lu J, Getz G, Miska EA, et al: MicroRNA
expression profiles classify human cancers. Nature. 435:834–838.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen CZ: MicroRNAs as oncogenes and tumor
suppressors. N Engl J Med. 353:1768–1771. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hwang HW and Mendell JT: MicroRNAs in cell
proliferation, cell death, and tumorigenesis. Br J Cancer.
94:776–780. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Johnson SM, Grosshans H, Shingara J, et
al: RAS is regulated by the let-7 microRNA family. Cell.
120:635–647. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Corney DC, Flesken-Nikitin A, Godwin AK,
Wang W and Nikitin AY: MicroRNA-34b and MicroRNA-34c are targets of
p53 and cooperate in control of cell proliferation and
adhesion-independent growth. Cancer Res. 67:8433–8438. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chang TC, Wentzel EA, Kent OA, et al:
Transactivation of miR-34a by p53 broadly influences gene
expression and promotes apoptosis. Mol Cell. 26:745–752. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Debernardi S, Skoulakis S, Molloy G,
Chaplin T, Dixon-McIver A and Young BD: MicroRNA miR-181a
correlates with morphological sub-class of acute myeloid leukaemia
and the expression of its target genes in global genome-wide
analysis. Leukemia. 21:912–916. 2007.PubMed/NCBI
|
|
14
|
Blower PE, Chung JH, Verducci JS, et al:
MicroRNAs modulate the chemosensitivity of tumor cells. Mol Cancer
Ther. 7:1–9. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zheng T, Wang J, Chen X and Liu L: Role of
microRNA in anticancer drug resistance. Int J Cancer. 126:2–10.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Boren T, Xiong Y, Hakam A, et al:
MicroRNAs and their target messenger RNAs associated with ovarian
cancer response to chemotherapy. Gynecol Oncol. 113:249–255. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zheng A, Kallio A and Harkonen P:
Tamoxifen-induced rapid death of MCF-7 breast cancer cells is
mediated via extracellularly signal-regulated kinase signaling and
can be abrogated by estrogen. Endocrinology. 148:2764–2777. 2007.
View Article : Google Scholar
|
|
18
|
Ritchie ME, Silver J, Oshlack A, et al: A
comparison of background correction methods for two-colour
microarrays. Bioinformatics. 23:2700–2707. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Smyth GK and Speed T: Normalization of
cDNA microarray data. Methods. 31:265–273. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tusher VG, Tibshirani R and Chu G:
Significance analysis of microarrays applied to the ionizing
radiation response. Proc Natl Acad Sci USA. 98:5116–5121. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bar M, Wyman SK, Fritz BR, et al: MicroRNA
discovery and profiling in human embryonic stem cells by deep
sequencing of small RNA libraries. Stem Cells. 26:2496–2505. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang X and El Naqa IM: Prediction of both
conserved and nonconserved microRNA targets in animals.
Bioinformatics. 24:325–332. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sontheimer EJ and Carthew RW: Silence from
within: endogenous siRNAs and miRNAs. Cell. 122:9–12. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
O’Donnell KA, Wentzel EA, Zeller KI, Dang
CV and Mendell JT: c-Myc-regulated microRNAs modulate E2F1
expression. Nature. 435:839–843. 2005.PubMed/NCBI
|
|
25
|
Blower PE, Verducci JS, Lin S, et al:
MicroRNA expression profiles for the NCI-60 cancer cell panel. Mol
Cancer Ther. 6:1483–1491. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cittelly DM, Dimitrova I, Howe EN, et al:
Restoration of miR-200c to ovarian cancer reduces tumor burden and
increases sensitivity to paclitaxel. Mol Cancer Ther. 11:2556–2565.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen Y, Zuo J, Liu Y, Gao H and Liu W:
Inhibitory effects of miRNA-200c on chemotherapy-resistance and
cell proliferation of gastric cancer SGC7901/DDP cells. Chin J
Cancer. 29:1006–1011. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fujita Y, Kojima K, Ohhashi R, et al:
MiR-148a attenuates paclitaxel resistance of hormone-refractory,
drug-resistant prostate cancer PC3 cells by regulating MSK1
expression. J Biol Chem. 285:19076–19084. 2010. View Article : Google Scholar
|
|
29
|
Du L, Subauste MC, DeSevo C, et al:
miR-337-3p and its targets STAT3 and RAP1A modulate
taxane sensitivity in non-small cell lung cancers. PLoS One.
7:e391672012.PubMed/NCBI
|
|
30
|
Catuogno S, Cerchia L, Romano G, Pognonec
P, Condorelli G and de Franciscis V: miR-34c may protect lung
cancer cells from paclitaxel-induced apoptosis. Oncogene.
32:341–351. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Holleman A, Chung I, Olsen RR, et al:
miR-135a contributes to paclitaxel resistance in tumor cells both
in vitro and in vivo. Oncogene. 30:4386–4398. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li J, Zhang Y, Zhao J, Kong F and Chen Y:
Overexpression of miR-22 reverses paclitaxel-induced
chemoresistance through activation of PTEN signaling in p53-mutated
colon cancer cells. Mol Cell Biochem. 357:31–38. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhou M, Liu Z, Zhao Y, et al:
MicroRNA-125b confers the resistance of breast cancer cells to
paclitaxel through suppression of pro-apoptotic Bcl-2 antagonist
killer 1 (Bak1) expression. J Biol Chem. 285:21496–21507. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mei M, Ren Y, Zhou X, et al:
Downregulation of miR-21 enhances chemotherapeutic effect of taxol
in breast carcinoma cells. Technol Cancer Res Treat. 9:77–86. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Huang YW, Liu JC, Deatherage DE, et al:
Epigenetic repression of microRNA-129-2 leads to overexpression of
SOX4 oncogene in endometrial cancer. Cancer Res. 69:9038–9046.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mialon A, Sankinen M, Soderstrom H, et al:
DNA topoisomerase I is a cofactor for c-Jun in the regulation of
epidermal growth factor receptor expression and cancer cell
proliferation. Mol Cell Biol. 25:5040–5051. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Barroso-delJesus A, Romero-Lopez C,
Lucena-Aguilar G, et al: Embryonic stem cell-specific miR302–367
cluster: human gene structure and functional characterization of
its core promoter. Mol Cell Biol. 28:6609–6619. 2008.PubMed/NCBI
|
|
38
|
Lavon I, Zrihan D, Granit A, et al:
Gliomas display a microRNA expression profile reminiscent of neural
precursor cells. Neuro Oncol. 12:422–433. 2010.PubMed/NCBI
|
|
39
|
Sokilde R, Kaczkowski B, Podolska A, et
al: Global microRNA analysis of the NCI-60 cancer cell panel. Mol
Cancer Ther. 10:375–384. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Greither T, Grochola LF, Udelnow A,
Lautenschlager C, Wurl P and Taubert H: Elevated expression of
microRNAs 155, 203, 210 and 222 in pancreatic tumors is associated
with poorer survival. Int J Cancer. 126:73–80. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Levati L, Alvino E, Pagani E, et al:
Altered expression of selected microRNAs in melanoma:
antiproliferative and proapoptotic activity of miRNA-155. Int J
Oncol. 35:393–400. 2009.PubMed/NCBI
|
|
42
|
Pan X, Zhao J, Zhang WN, et al: Induction
of SOX4 by DNA damage is critical for p53 stabilization and
function. Proc Natl Acad Sci USA. 106:3788–3793. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Baraniskin A, Birkenkamp-Demtroder K,
Maghnouj A, et al: MiR-30a-5p suppresses tumor growth in colon
carcinoma by targeting DTL. Carcinogenesis. 33:732–739. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yanaihara N, Caplen N, Bowman E, et al:
Unique microRNA molecular profiles in lung cancer diagnosis and
prognosis. Cancer Cell. 9:189–198. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Marchini S, Cavalieri D, Fruscio R, et al:
Association between miR-200c and the survival of patients with
stage I epithelial ovarian cancer: a retrospective study of two
independent tumour tissue collections. Lancet Oncol. 12:273–285.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Li X, Zhang Y, Zhang Y, Ding J, Wu K and
Fan D: Survival prediction of gastric cancer by a seven-microRNA
signature. Gut. 59:579–585. 2010. View Article : Google Scholar : PubMed/NCBI
|