Introduction
Endoplasmic reticulum (ER) stress (ERSR) is engaged
in many cellular functions such as protein synthesis, folding and
storage, as well as in calcium signaling. When disturbances in
these functions occur and levels of misfolded proteins in the ER
increase, the goal of ERSR is to enhance protein folding, reduce
new protein synthesis and clear misfolded proteins. In the case of
a default in this function, ERSR may trigger apoptotic cell death
for elimination of defective cells (1). There is much evidence that enhanced
ERSR, triggering apoptosis, is associated with many degenerative
diseases (reviewed in ref 2). An
opposite situation may be observed in tumor cells, which are
generally resistant to apoptotic stimuli. For example, malignant
glioma cells are in a condition of constant low ERSR, which
possibly contributes to their resistance to treatment (1). PC12 cells, derived from rat
pheochromocytoma, possess several mechanisms enabling them to
counteract apoptosis. Recently, a pathway mediated by heat shock
proteins, which prevents ERSR-induced apoptosis has been revealed
(3). Chaperone protein HSP72
enhances IRE1a-XBP1 signaling through a physical interaction
(4), and HSP90 is a key protein in
several pathways of cell proliferation and tumor progression
(3,5). Resistance to apoptosis is frequently
mediated by overexpression of anti-apoptotic or the absence of
pro-apoptotic proteins from the Bcl-2 family (6,7). This
pathway has been studied during the last decade of research, and
several substances modulating either expression or activity of
these proteins have been tested as new potential anticancer drugs
(6). Saito et al (8) induced apoptosis in PC12 cells by
multitargeted receptor tyrosine kinase inhibitor (sunitib), which
modulated the Bcl-2 and BAD pathways. The cytotoxic effect of this
drug was pronounced when autophagy in these cells was inhibited
(9).
A special group of potential anticancer drugs
include active substances from traditional medicinal plants.
Extracts from plants of the Phyllanthus species were shown
to interfere with multiple signaling cascades in human prostate
carcinoma PC-3 cells and were able to trigger apoptotic cell death
(10). Triptolide isolated from the
plant Trypterygium wilfordii was shown to inhibit
proliferation of a variety of cancer cells acting through the NF-κB
cascade (11,12). Several investigators have reported
the ability of capsaicin (Capsicum species) to prevent
tumorigenesis by triggering apoptotic pathways. Capsaicin
(8-methyl-N-vanillyl-6-nonenamide), a member of the vanilloid
family, binds to a receptor called the vanilloid receptor subtype 1
(TRPV1), which has been shown to be a member of the superfamily of
TRP ion channels and permits cations to pass through the cell
membrane and into the cell when activated. Ito et al
(13) showed that capsaicin induces
apoptosis in leukemic cells through oxidative stress. Experiments
more focused on receptor TRPV1 have shown its anti-oncogenic
effects in transitional urothelial cancer of the human bladder. A
progressive decrease in TRPV1 expression during the transitional
stage of cancer was found to trigger the development of a more
aggressive phenotype and invasiveness (14). When capsaicin was applied to
TRPV1-knockout urothelial cancer cells, an even more aggressive
form of tumor was observed (15).
It was also recently shown that the TRPV1 channel activated by
capsaicin caused an increase in intracellular calcium
concentrations in mammalian skeletal muscle (16).
In our previous studies we proved that two herbal
compounds, triptolide (TTL) and capsaicin (Caps), are inhibitors of
the nuclear transcription factor NF-κB in PC12 and MPC cells
(11). Inhibition of this factor
caused enhanced expression of norepinephrine transporter (NET) and
apoptosis (11). The aim of the
present study was to evaluate the mechanisms triggered by capsaicin
leading to the apoptosis in PC12 cells. We focused on primary
signals, which may be connected with calcium homeostasis and
calcium transporting proteins in the membrane of the endoplasmic
reticulum, as well as on typical factors of ERSR and apoptosis.
Materials and methods
Cell cultivation and treatment
PC12 cells (German Collection of Microorganisms and
Cell cultures, DSMZ, Braunschweig, Germany) derived from rat
pheochromocytoma were cultured in Dulbecco’s minimal essential
medium (Biochrom AG, Berlin, Germany) with high glucose (4.5 g/l)
supplemented with 15% fetal calf serum and penicillin and
streptomycin antibiotics. Cells were cultured in a water-saturated
atmosphere at 37°C with 5% CO2. Treatment was performed
by adding of 50, 100 and 500 μM (E)-capsaicin (Caps; Merck,
Germany) directly to the cultivation media for 24 h.
RNA isolation and relative quantification
of mRNA levels by RT-PCR and qPCR
Total RNA was isolated by TRI reagent (MRC Ltd.,
Cincinnati, OH, USA). Briefly, cells were scraped and homogenized
by a pipette tip in sterile water and afterwards TRI reagent was
added. After 5 min the homogenate was extracted by chloroform. RNAs
in the aqueous phase were precipitated by isopropanol. RNA pellet
was washed with 75% ethanol and stored in 96% ethanol at −70°C. The
purity, quantity and integrity of isolated RNAs were assessed using
GeneQuant Pro spectrophotometer (Amersham Biosciences,
Buckinghamshire, UK). Reverse transcription was performed using 1.5
μg of total RNAs and Ready-To-Go You-Prime First-Strand beads with
pd(N)6 primer (both from GE Healthcare Life Sciences, USA). PCR
specific for the unspliced form of X-box binding protein 1 (XBP1)
(GI: 51259532) was performed with primers: XBP1 forward,
5′-AGCGCTGCCGCTCATGCTTC-3′ and reverse, 5′-TCTCGCGCAGTCTGTGCTGC-3′;
for ATF4 (GI: 165971604) forward, 5′-GGCCACCATGGCGTATTAAGA-3′ and
reverse, 5′-GACATTAAGTCCCCCGGCCAA-3′; and for CHOP (GI: 2660765)
forward, 5′-AGGGCTAGCTTGGTCCTAGA-3′ and reverse,
5′-CCCCAAGTCCTGAACTCCAC-3′. For anti-apoptotic Bcl-2 (GI: 408946)
primers were: Bcl-2 forward, 5′-ACTTCTCTCGTCGCTACCGT-3′ and
reverse, 5′-GTTCCACAAAGGCATCCCAG-3′; for Bax (GI: 3320116) forward,
5′-GAAGCTGAGCGAGTCTCTCC-3′ and reverse, 5′-GATCAGCTCGGGCACTTTAG-3′;
for SERCA2 (GI: 8392934) forward, 5′-ATTGTTCGAAGTCTGCCTTCTGTG-3′
and reverse, 5′-CATAGGTTGATCCAGTATGGTAAA-3′. Primers for rat IP3R1
(GI: 1055286) were: IP3R1 forward, 5′-GTGGAGGTTTCATCTGCAAGC-3′ and
reverse, 5′-GCTTTCGTGGAATACTCGGTC-3′; for IP3R2 (GI: 13752805)
IP3R2 forward, 5′-GCTCTTGTCCCTGACATTG-3′ and reverse,
5′-CCCATGTCTCCATTCTCATAGC-3′; and for IP3R3 (GI: 6981109) forward,
5′-CTGCCCAAGAGGAGGAGGAAG-3′ and reverse,
5′-GAACAGCGCGGCAATGGAGAAG-3′. Primers for ryanodine receptor type 2
(RyR2) (GI: 2305245) were forward, 5′-CATCGGTGAAATTGAAGA-3′ and
reverse, 5′-AGCATCAATGATCAAACCTTG-3′. As housekeeping genes rat
β-actin A (GI: 42475962) was used with primers BA forward,
5′-AGTGTGACGTTGACATCCGT-3′ and reverse, 5′-GACTGATCGTACTCCTGCTT-3′
or cyclophilin (GI: 203701) with primers CYCLO forward,
5′-CGTGCTCTGAGCACTGGGGAGAAA-3′ and reverse,
5′-CATGCCTTCTTTCACCTTCCCAAAGAC-3′. The same primers were used for
RT-PCR and also for real-time quantitative PCR. Products of RT-PCR
were analyzed on a 2% agarose gel and signals were evaluated by
PCBAS 2.0 software. Real-time PCR was performed on PikoReal 96
cycler with DyNAmo Color Flash SYBR-Green Master Mix (both from
Thermo Fisher Scientific, Hampshire, UK). Results were evaluated by
PikoReal software 2 as a peak area for every well and quantified
relatively from Cq values according to the formula ΔΔCq =
ΔCq(sample) − ΔCq(housekeeper), where the rat
β-actin A was used as the housekeeping gene.
[Ca2+]free
measurement in the reticular fraction with Rhod-5N dye
We used the method as was described in our previous
study (17). Briefly, cells were
scraped from wells, sedimented by centrifugation and washed with
phosphate-buffered saline (PBS) solution. Gentle lysis was
performed with 100 μl of cell lysis buffer provided in the kit for
cytoplasmic and nuclear protein isolation (ProteoJet™; Fermentas,
Germany) in the presence of dithiothreitol (DTT) (10 mM).
Post-mitochondrial fractions with ER cisternae were isolated as
described in a study by Kalén et al (18). Pellets from the post-mitochondrial
fraction were homogenized in nuclear lysis buffer from the
ProteoJet™ kit and pipetted to wells in a 24-well plate. To each
sample, Rhod-5N fluorescent dye was added to a final concentration
of 20 μM. Measurements were captured using the BioTek (BioTek,
Germany) fluorescence reader (excitation, 551 nm/emission, 576 nm).
The fluorescent (F) signal was saturated by adding EGTA solution,
pH 7.0, to the final concentration of 2.5 mM (Fmin). Fmax value was
measured by adding CaCl2 to the final concentration of
0.5 mM. Final values of [Ca2+]free were
calculated according to the formula:
[Ca2+]free = Kd [(Fmax − F)/(F − Fmin)],
where Kd for Rhod-5N is 320 nM. Results were calculated relative to
protein levels, which were determined in the samples by the method
of Lowry et al (19).
Cytofluorometric analysis of the
mitochondrial membrane potential
Analysis of mitochondrial membrane potential via
Ψm was performed as previously described (20). Briefly, cells were collected by
centrifugation at 1000 × g for 5 min and washed twice with cold
PBS. Incubation was performed in 200 μl of PBS/0.2% BSA containing
4 μM JC-1 fluorescent dye
(5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethylbenzimidazolyl
carbocyanine iodide) and 7-aminoactinomycin D (7-AAD) (5 ng/μl)
both from Invitrogen Life Technologies, USA for 30 min at 37°C in
the dark. 7-AAD was used to exclude the population of necrotic
cells. Cell data were acquired using the EPICS Altra (Beckman
Coulter) flow cytometer equipped with a 488-nm excitation laser,
and fluorescence emission of JC-1 green, JC-1 red, and 7-AAD was
measured using a band pass filter set at 525, 575 and 675 nm,
respectively. Forward and side light scattering characteristics
were used to exclude cell debris from the analysis. For each
analysis, 1×104 cells were acquired, and the ratio of
JC-1 red/JC-1 green fluorescence of viable cells (7-AAD negative)
was used to calculate the decrease in mitochondrial membrane
potential (ΔΨm). Data were analyzed by FCS4 software (De
Novo Software, Los Angeles, CA, USA).
Detection of apoptosis with
Annexin-V-FLUOS
Cells were sedimented and washed with 1Χ PBS
solution. The cell pellet from each well was labeled using the
Annexin V-FLUOS/propidium iodide labeling kit (Roche Diagnostics,
Germany) according to the protocol of the producer and incubated at
room temperature in the dark for 20 min. After the incubation was
completed, the reaction was halted by adding three volumes of ice
cold PBS. Measurements were performed on an Accuri C6 flow
cytometer, and results were evaluated by the C-Flow sampler 2.1 (BD
Accuri Cytometers, Ann Arbor, MI, USA).
Western blot analysis
Cells were scraped and suspended in 10 mM Tris-HCl,
pH 7.5, 1 mM phenylmethylsulfonyl fluoride (PMSF) (Serva, Germany),
protease inhibitor cocktail tablets (Complete EDTA-Free; Roche
Diagnostics) and subjected to centrifugation for 10 min at 10,000 ×
g at 4°C. The pellet was resuspended in Tris-buffered saline (TBS)
containing 50 μM CHAPS
[3[(3-cholamidopropyl)dimethylammonio]-l-propanesulfonate)] (Sigma,
USA) and then incubated for 10 min at 4°C. The lysate was
centrifuged for 10 min at 10,000 × g at 4°C. Protein concentration
of the supernatants was determined by the method of Lowry et
al (19). Protein extract (20
μg) from each sample was separated by electrophoresis on 10% SDS
polyacrylamide gels and proteins were transferred to the Hybond-P
membrane using semi-dry blotting (Owl, Inc., Portsmouth, NH, USA).
Membranes were blocked in 5% non-fat dry milk in TBS with Tween-20
(TBS-T) overnight at 4°C and then incubated for 1 h with XBP1/TREB5
monoclonal antibody (Antibodies-online GmbH, Aachen, Germany),
CREB2/ATF4 (sc-200) polyclonal antibody, GADD153/CHOP (sc-575)
polyclonal antibody, Bax (sc-7480) polyclonal antibody (all from
Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA) and the Bcl-2
polyclonal antibody (Abcam, Cambridge, UK). Following washing,
membranes were incubated with secondary antibodies to rabbit and
mouse IgG-conjugated to horseradish peroxidase for 1 h at room
temperature. For relative quantification, each membrane was
reprobed for housekeeper GAPDH mouse monoclonal antibody (Abcam).
An enhanced chemiluminiscence detection system (ECL Plus; Amersham
Biosciences) was used to detect the bound antibody. The optical
density of individual bands was quantified using PCBAS 2.0
software.
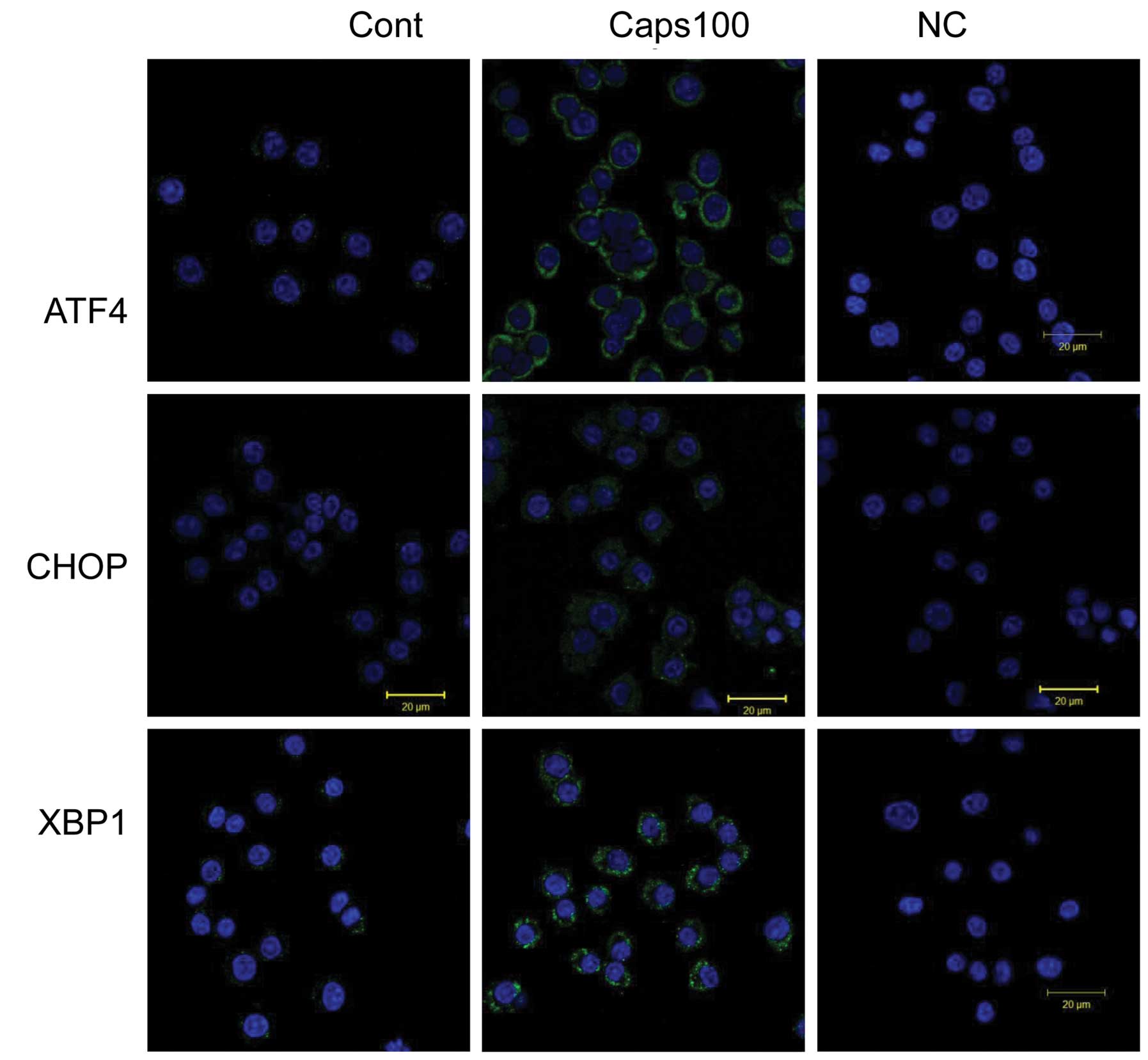
Immunofluorescence
For immunofluorescence, cells were fixed with
ice-cold methanol. In these experiments, the same antibodies
against XBP1, ATF4 and CHOP were used as in western blot analysis.
Cells grown on glass coverslips were fixed in ice-cold methanol at
−20°C for 5 min. Non-specific binding was blocked by incubation
with PBS containing 3% BSA for 60 min at 37°C. Cells were then
incubated with primary antibody diluted 1:500 in PBS with 1% BSA
(PBS-BSA) for 1 h at 37°C, washed three times with PBS-BSA for 10
min, incubated with CF Fluor® 488 goat anti-rabbit IgG
(Biotium, Hayward, CA, USA) diluted 1:1,000 in PBS-BSA for 1 h at
37°C and washed as previously described. Finally, cells were
mounted onto slides in mounting medium with CitiFluor (Agar
Scientific Ltd., Essex, UK), and analyzed using the LSM 510 Meta
microscope (Carl Zeiss, Jena, Germany) with EC Plan-Neofluar ×40
objective. Cellular nuclei were stained with
4′,6-diamidino-2-phenylindole (DAPI) (Life Technologies Carlsbad,
CA, USA). All images were captured using the same camera and
microscope.
Statistical analysis
Each value was the average of 24-wells in at least 4
independent cultivations of PC12 cells. Results are presented as
means ± SEM. Statistical differences between the groups were
determined by ANOVA. p<0.05 was considered to indicate a
statistical significant result. An adjusted t-test with p-values
corrected by the Bonferroni’s method was used for multiple
comparisons (Instat; GraphPad Software).
Results
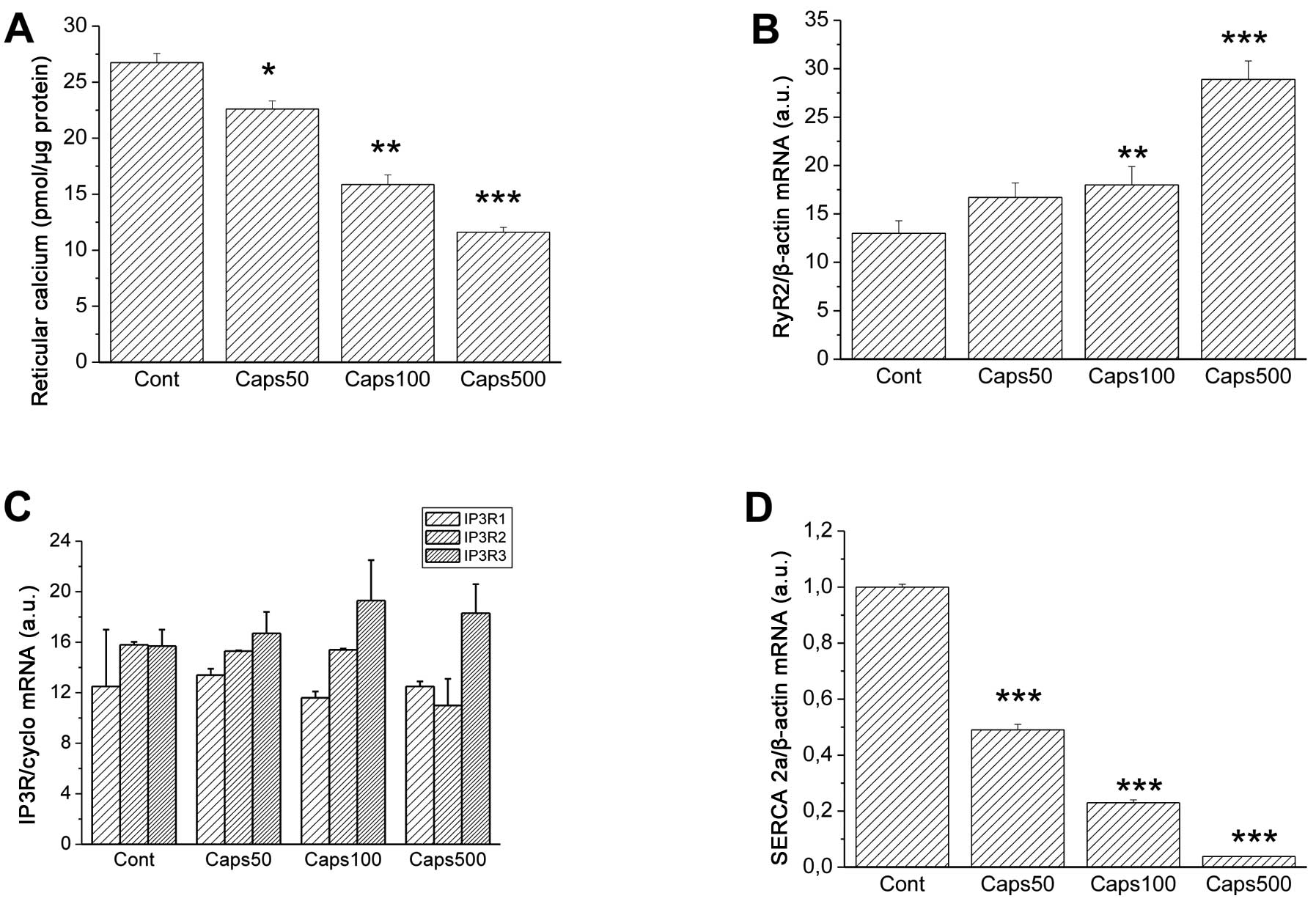
Capsaicin (Caps) modulated the calcium turnover in
the reticular fraction of PC12 cells in a concentration-dependent
manner. Whereas 50 μM Caps moderately lowered the calcium content
when compared to the control, 100 and 500 μM Caps caused a
substantial and significant decrease (Fig. 1A) [from 26.74±0.82 (control) to
15.86±0.86 and 11.60±0.43 pmol/μg of protein]. This decrease was
accompanied by changes in the expression of reticular calcium
transport systems. Expression of calcium release channel RyR2 was
elevated mainly at 100 and 500 μM Caps [Fig. 1B; from 13.0±1.3 (control) to
18.0±1.9 and 28.9±2.8 a.u., respectively]. In contrast, expression
of SERCA2, which is responsible for transport of calcium to
endoplasmic reticulum, was significantly reduced at all three Caps
concentrations tested [Fig. 1D;
from 1.0 (control) to 0.038 a.u. (at 500 μM Caps)]. Notably, no
significant effect by Caps was noted on the expression of other
calcium release channels, IP3 receptors (IP3R)1, 2 and 3 (Fig. 1C).
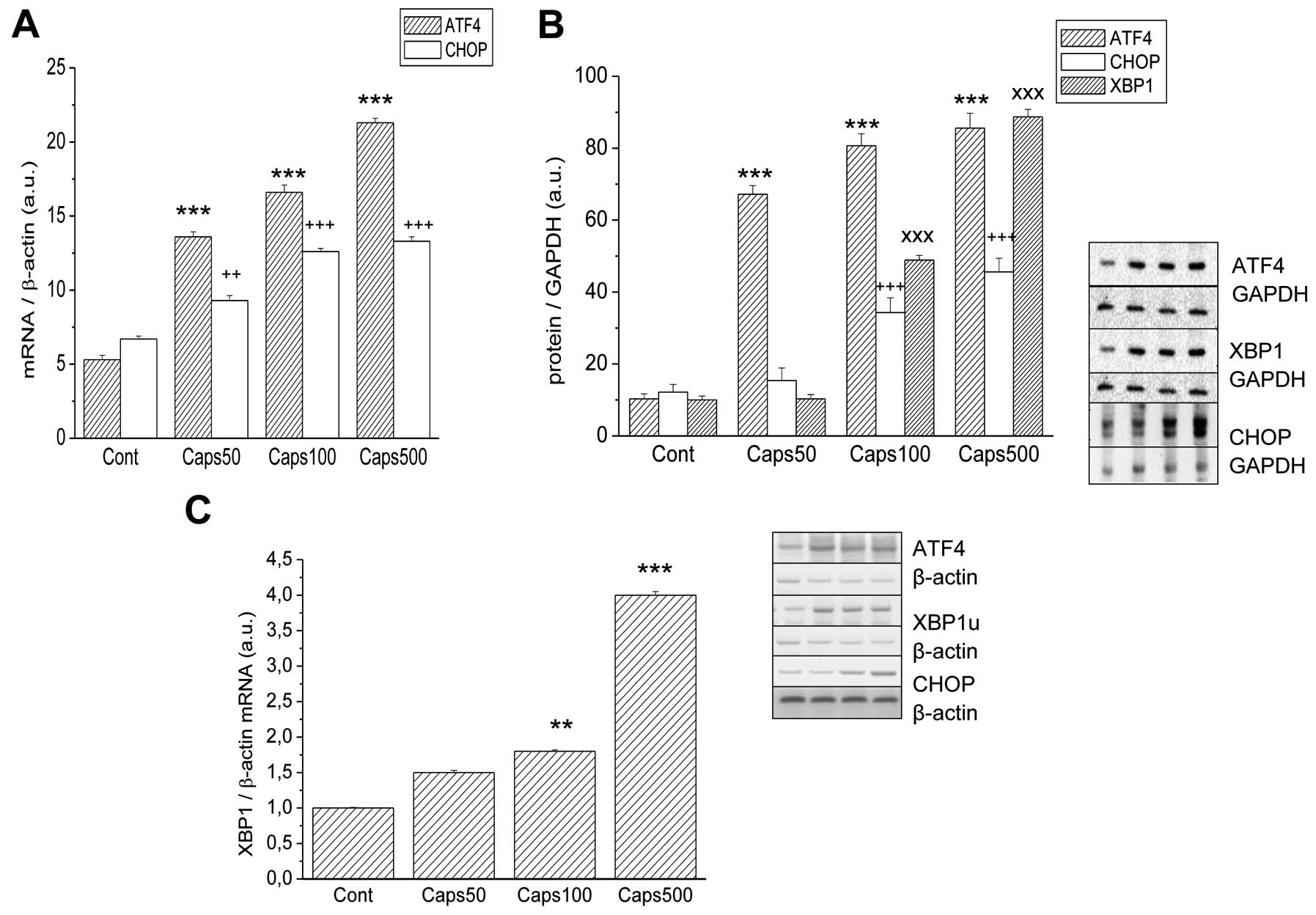
Gene and protein expression of the ER stress markers
ATF4, CHOP and XBP1 also exhibited a concentration-dependence on
capsaicin. Significantly elevated mRNA signals were noted for ATF4
and CHOP (Fig. 2A) and XBP1
(Fig. 2C). CHOP mRNA was
significantly elevated at 100 and 500 μM Caps [from 6.7±0.2
(control) to 12.6±0.21 and 13.3±0.3 a.u., respectively]. A
different pattern of expression was observed for ER stress marker
ATF4, where the mRNA signal was elevated all three capsaicin
concentrations (50, 100, 500 μM) [from 5.3±0.3 (control) to
13.6±0.33; 16.6±0.5 and 21.3±0.3 a.u., respectively]. Relative
quantification of the unspliced form of the XBP1 by real-time PCR
revealed its mRNA elevation at 100 and 500 μM Caps. Particularly,
500 μM Caps caused a 4-fold elevated expression of XBP1 (Fig. 2C). An increase in mRNA signals was
accompanied by an increase in protein signals as determined by
western blotting (Fig. 2B). Levels
of CHOP and XBP1 proteins were increased significantly at 100 and
500 μM Caps [CHOP: 12.2±2.1 (control) to 34.3±4.1 and 45.6±3.8
a.u.; XBP1: from 10.0±1.1 (control) to 48.9±1.3 and 88.7±2.1 a.u.,
respectively]. ATF4 protein signal was elevated at all three
concentrations of capsaicin [from 10.3±1.4 (control) to 67.2±2.4;
80.7±3.3 and 85.6±4.1 a.u., respectively]. A strong increase in
immunofluorescent signals at the ER for ATF4, CHOP and unspliced
XBP1 were observed in the PC12 cells treated with 100 μM Caps at 24
h (Fig. 3).
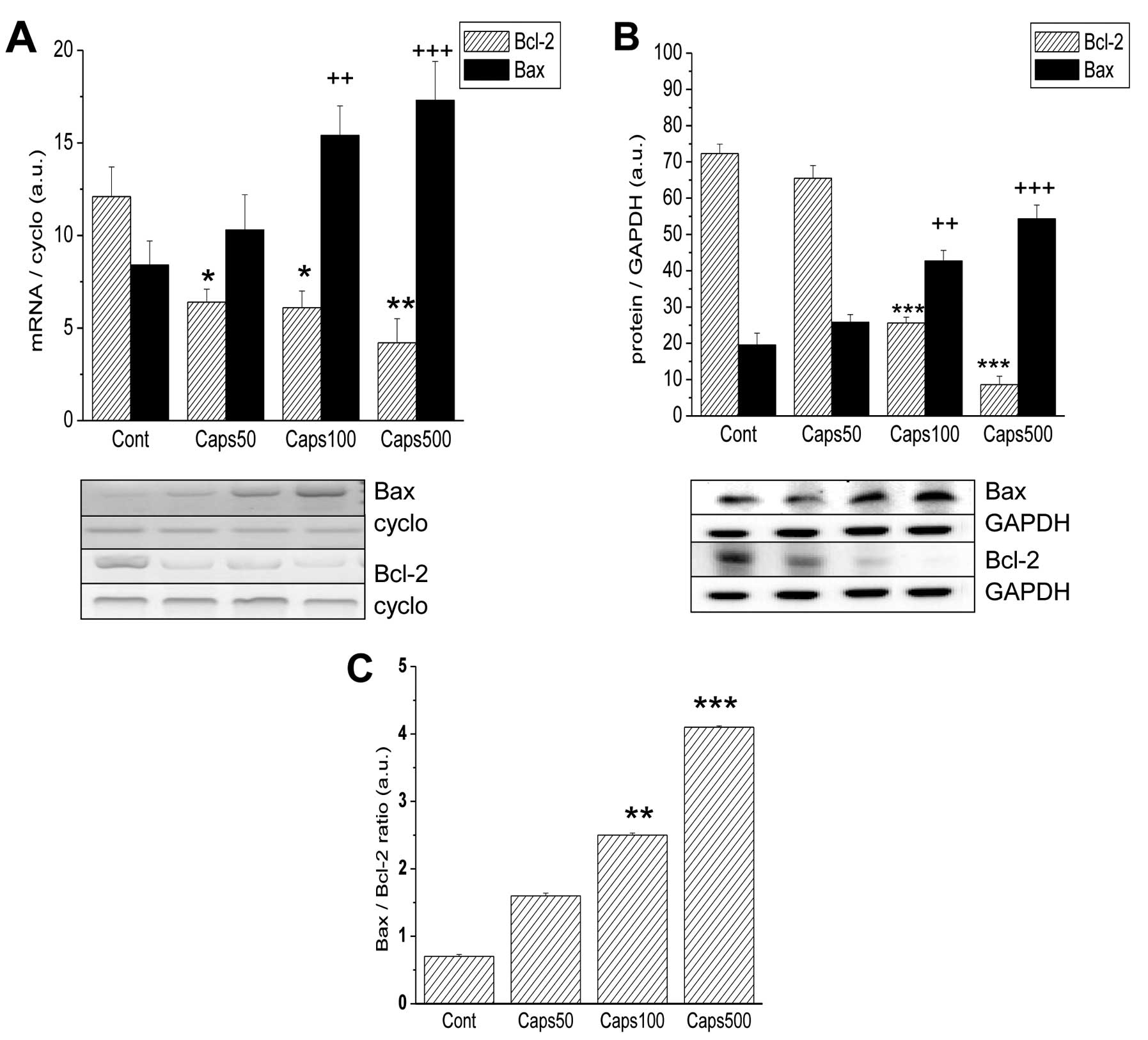
Decreased reticular calcium content is associated
mainly with changes in the expression and function of calcium
rheostat proteins from the Bcl-2 family. We monitored two typical
members of this family, Bcl-2 known as an anti-apoptotic protein
and Bax as a representative pro-apoptotic protein acting on the ER
and mitochondria. Fig. 4 shows that
100 and 500 μM capsaicin caused a significant elevation in mRNA and
protein levels of pro-apoptotic Bax [Fig. 4A; from 8.4±1.3 (control) to 15.4±2.1
a.u. (at 100 μM) and Fig. 4B from
19.54±3.3 (control) to 42.74±2.9 and 54.34±3.8 a.u. (at 100 and 500
μM, respectively)]. In contrast, levels of anti-apoptotic Bcl-2
were dramatically decreased at both the mRNA [Fig. 4A; from 12.1±1.6 (control) to 6.4±0.7
6.1±0.9 and 4.2±1.3 a.u. (at 50, 100 and 500 μM, respectively)] and
protein levels [Fig. 4B; from
72.34±2.6 (control) to 25.64±1.6 and 8.64±2.3 a.u. (at 100 and 500
μM, respectively)] This effect resulted in a change in the ratio of
Bax/Bcl-2 content, which was shifted in favor of Bax (Fig. 4C).
Possible induction of apoptosis by capsaicin in PC12
cells was tested by a decline in mitochondrial membrane potential
ΔΨm [Fig. 5B;
ΔΨm decreased from 92.5±1.15 (control) to 84.4±1.13 and
77.38±1.63 a.u. (at 100 and 500 μM, respectively)] and by measuring
cytoplasmic membrane phosphatidylserine translocation by Annexin
V-FLUOS (Fig. 5A). The percentage
of Annexin-positive cells in the population increased from
10.64±1.3 (control) to 19.6±2.5, 28.74±3.1 and 44.3±3.6 at 50, 100
and 500 μM, respectively.
Discussion
Tumor cells are known to be resistant to apoptotic
stimuli by several mechanisms. Tumors overexpress anti-apoptotic
proteins such Bcl-2 and have decreased expression of pro-apoptotic
proteins such as Bax or BH3 (21),
have impaired signals from death receptors (22), or activated nuclear factor NF-κB
which prevents activation of caspase-mediated cleavage (23). In our research, we showed that
impaired calcium signals induced by capsaicin may result in ER
stress and apoptosis. We observed significant calcium leakage from
ER, which was associated with the overexpression of RyR2 calcium
release channels. This depletion was apparently irreversible, as
expression of sarco-endoplasmic ATPase, responsible for reload of
calcium back to the ER, was substantially decreased. It is known
that overexpression of RyR2 increases susceptibility of cells to
apoptosis. For example, it was shown that overexpression of a
splice variant of RyR2 may trigger apoptosis in the heart (24). SERCA2 has 3 binding sites for NF-κB
in the promoter region and a decrease in expression may be
explained by the inhibitory effect of capsaicin on NF-κB. It seems
that capsaicin has a double impact on calcium homeostasis in the ER
of PC12 cells. The first effect is caused by activation of the
TRPV1 channel with possible changes in cationic fluxes. The second
effect is modulation of the expression of calcium transporting
systems RyR2 and SERCA2 with an impact on calcium content in the
ER. Functional connection of these structures was recently
described as TRPV1/RyR1 crosstalk in mouse skeletal muscle, where
TRPV1 was expressed at ER membranes in the proximity of SERCA1
pumps (25). In our research,
capsaicin was also found to modulate calcium rheostat proteins at
the ER, and the Bcl-2/Bax ratio, which showed an expression shift
to pro-apoptotic Bax. Several authors have shown that cells with
overexpressed Bax, as well as cells having Bcl-2 deficiency, show
decreased ER calcium content (7).
Decrease in the expression of Bcl-2 protein caused by capsaicin is
in accordance with our previous study (11), where we showed that capsaicin is an
inhibitor of NF-κB. This transcription factor promotes upregulation
of anti-apoptotic Bcl-2 (1).
ER stress-mediated apoptosis through the activation
of CHOP has been intensively studied (26,27).
CHOP (also called growth arrest DNA damage-inducible gene 153,
GADD153) transcriptionally regulates genes that participate in the
apoptotic pathway and it has been shown that increased levels of
this protein are associated with inhibition of Bcl-2 which triggers
the effect of the Bax/Bad systems in mitochondria (24). This effect may be antagonized by
activation of NF-κB and upregulation of Bcl-2 (26,27).
We also observed, in addition to an altered Bax/Bcl-2 ratio,
elevated levels of CHOP suggesting that capsaicin modulates only
this pathway. We also found enhanced expression of ATF4, which
indicated an elevated ratio of more signals typical for ER stress,
such as protein folding and degradation. Very similar results were
obtained by Sánchez et al in prostate tumor cells. They
showed by microarray, real-time PCR and western blotting techniques
that capsaicin upregulates the CHOP and ATF4 pathways (28).
The pivotal role of IRE1α/XBP1 signaling in
tumorigenicity has been well recognized (29). The spliced active form of XBP1
(XBP1s) acts as a transcription factor in the nucleus and activates
genes for protein folding and restoration of ER homeostasis. In
contrast, the unspliced form of XBP1 (XBP1u) functions as the
dominant-negative form that antagonizes the function of XBP1s
(30). We observed that capsaicin
caused an increase in the XBP1 at ER. In connection to all these
evidence for induced ER stress, we also found typical markers for
apoptosis in the PC12 cells. Loss of mitochondrial membrane
potential (ΔΨm) and binding of Annexin V-FLUOS indicated
that ER stress induced by capsaicin resulted in apoptosis. Taken
together, capsaicin acts in PC12 cells by triggering ER stress in a
concentration-dependent manner and this stress results in
apoptosis.
This is the first study demonstrating that the
primary signal induced by capsaicin is calcium release from the ER
which consequently triggers a cascade of events leading to ERSR and
apoptosis. These data may aid to the identification of new targets
and pathways for cancer treatment.
Acknowledgements
The present study was supported by grant
APVV-0045-11, grant VEGA 2/0074 and funding from the Center of
Excellence for Studying Metabolic Aspects of Development,
Diagnostics and Treatment of Oncology Diseases (CEMAN).
References
|
1
|
Johnson GG, White MC and Grimaldi M:
Stressed to death: targeting endoplasmic reticulum stress
response-induced apoptosis in gliomas. Curr Pharm Des. 17:284–292.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chakrabarti A, Chen AW and Varner JD: A
review of the mammalian unfolded protein response. Biotechnol
Bioeng. 108:2777–2793. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Giubellino A, Sourbier C, Lee MJ,
Scroggins B, Bullova P, Landau M, Ying W, Neckers L, Trepel JB and
Pacak K: Targeting heat shock protein 90 for the treatment of
malignant pheochromocytoma. PLoS One. 8:e560832013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gupta S, Deepti A, Deegan S, Lisbona F,
Hetz C and Samal IA: HSP72 protects cells from ER stress-induced
apoptosis via enhancement of IRE1α-XBP1 signaling through a
physical interaction. PLoS Biol. 8:e10004102010.PubMed/NCBI
|
|
5
|
Jhaveri K, Taldone T, Modi S and Chiosis
G: Advances in the clinical development of heat shock protein 90
(Hsp90) inhibitors in cancers. Biochim Biophys Acta. 1823:742–755.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Labi V, Grespi F, Baumgartner F and
Villunger A: Targeting the Bcl-2-regulated apoptosis pathway by BH3
mimetics: a breakthrough in anticancer therapy? Cell Death Differ.
15:977–987. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hetz AC: ER stress signaling and the BCL-2
family of proteins: from adaptation to irreversible cellular
damage. Antioxid Redox Signal. 9:2345–2355. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Saito Y, Tanaka Y, Aita Y, Ishii KA, Ikeda
T, Isobe K, Kawakami Y, Shimano H, Hara H and Takekoshi K:
Sunitinib induces apoptosis in pheochromocytoma tumor cells by
inhibiting VEGFR2/Akt/mTOR/S6K1 pathways through modulation of
Bcl-2 and BAD. Am J Physiol Endocrinol Metab. 302:E615–E625. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ikeda T, Ishii KA, Saito Y, Miura M,
Otagiri A, Kawakami Y, Shimano H, Hara H and Takekoshi K:
Inhibition of autophagy enhances sunitinib-induced cytotoxicity in
rat pheochromocytoma PC12 cells. J Pharmacol Sci. 121:67–73. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tang YQ, Jaganath I, Manikam R and Sekaran
SD: Phyllanthus suppresses prostate cancer cell, PC-3,
proliferation and induces apoptosis through multiple signalling
pathways (MAPKs, PI3K/Akt, NFκB, and hypoxia). Evid Based
Complement Alternat Med. 2013:6095812013.PubMed/NCBI
|
|
11
|
Pacak K, Sirova M, Giubellino A, Lencesova
L, Csaderova L, Laukova M, Hudecova S and Krizanova O: NF-κB
inhibition significantly upregulates the norepinephrine transporter
system, causes apoptosis in pheochromocytoma cell lines and
prevents metastasis in an animal model. Int J Cancer.
131:2445–2455. 2012.
|
|
12
|
Ou CC, Chen YW, Hsu SC, Sytwu HK, Loh SH,
Li JW and Liu JY: Triptolide transcriptionally represses HER2 in
ovarian cancer cells by targeting NF-κB. Evid Based Complement
Alternat Med. 2012:3502392012.PubMed/NCBI
|
|
13
|
Ito K, Nakazato T, Yamato K, Myiakawa Y,
Yamada T, Hozumi N, Segawa K, Ikeda Y and Kizaki M: Induction of
apoptosis in leukemic cells by homovanillic acid derivative,
capsaicin, through oxidative stress: implication of phosphorylation
of p53 at Ser-15 residue by reactive oxygen species. Cancer Res.
64:1071–1078. 2004. View Article : Google Scholar
|
|
14
|
Santoni G, Caprodossi S, Farfariello V,
Liberati S, Gismondi A and Amantini C: Antioncogenic effects of
transient receptor potential vanilloid 1 in the progression of
transitional urothelial cancer of human bladder. ISRN Urol.
2012:4582382012.
|
|
15
|
Caprodossi S, Amantini C, Nabissi M,
Morelli MB, Farfariello V, Santoni M, Gismondi A and Santoni G:
Capsaicin promotes a more aggressive gene expression phenotype and
invasiveness in null-TRPV1 urothelial cancer cells. Carcinogenesis.
32:686–694. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ito N, Ruegg UT, Kudo A, Miyagoe-Suzuki Y
and Takeda S: Capsaicin mimics mechanical load-induced
intracellular signaling events: involvement of TRPV1-mediated
calcium signaling in induction of skeletal muscle hypertrophy.
Channels. 7:221–224. 2013. View Article : Google Scholar
|
|
17
|
Lencesova L, Hudecova S, Csaderova L,
Markova J, Soltysova A, Pastorek M, Sedlak J, Wood ME, Whiteman M,
Ondrias K and Krizanova O: Sulphide signalling potentiates
apoptosis through the up-regulation of IP3 receptor types 1 and 2.
Acta Physiol. 208:350–361. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kalén A, Appelkvist EL and Dallner G: The
effects of inducers of the endoplasmic reticulum, peroxisomes and
mitochondria on the amounts and synthesis of ubiquinone in rat
liver subcellular membranes. Chem Biol Interact. 73:221–234.
1989.PubMed/NCBI
|
|
19
|
Lowry OH, Rosebrough NJ, Farr AL and
Randall RJ: Protein measurement with the Folin phenol reagent. J
Biol Chem. 193:265–275. 1951.PubMed/NCBI
|
|
20
|
Jakubikova J, Bao Y and Sedlak J:
Isothiocyanates induce cell cycle arrest, apoptosis and
mitochondrial potential depolarization in HL-60 and
multidrug-resistant cell lines. Anticancer Res. 25:3375–3386.
2005.PubMed/NCBI
|
|
21
|
Teijido O and Dejean L: Upregulation of
Bcl2 inhibits apoptosis-driven BAX insertion but favors BAX
relocalization in mitochondria. FEBS Lett. 584:3305–3310. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fulda S: Tumor resistance to apoptosis.
Int J Cancer. 124:511–515. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Levkau B, Garton KJ, Ferri N, Kloke K,
Nofer JR, Baba HA, Raines EW and Breithardt G: xIAP induces
cell-cycle arrest and activates nuclear factor-κB: new survival
pathways disabled by caspase-mediated cleavage during apoptosis of
human endothelial cells. Circ Res. 88:282–290. 2001.PubMed/NCBI
|
|
24
|
Valdivia HH: One gene, many proteins:
alternative splicing of the ryanodine receptor gene adds novel
functions to an already complex channel protein. Circ Res.
100:761–763. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lotteau S, Ducreux S, Romestaing C,
Legrand C and Van Coppenolle F: Characterization of functional
TRPV1 channels in the sarcoplasmic reticulum of mouse skeletal
muscle. PLoS One. 8:e586732013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ma Y and Hendershot LM: The role of
unfolded protein response in tumor development: friend or foe? Nat
Rev Cancer. 4:966–977. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yamaguchi H and Wang HG: CHOP is involved
in endoplasmic reticulum stress-induced apoptosis by enhancing DR5
expression in human carcinoma cells. J Biol Chem. 279:45495–45502.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sánchez AM, Martínez-Botas J,
Malagarie-Cazenave S, Olea N, Vara D, Lasunción MA and Díaz-Laviada
I: Induction of the endoplasmic reticulum stress protein
GADD153/CHOP by capsaicin in prostate PC-3 cells: a microarray
study. Biochem Biophys Res Commun. 372:785–791. 2008.PubMed/NCBI
|
|
29
|
Shajahan AN, Riggins RB and Clarke R: The
role of X-box binding protein-1 in tumorigenicity. Drug News
Perspect. 22:241–246. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang FM, Galson DL, Roodman GD and Ouyang
H: Resveratrol triggers the pro-apoptotic endoplasmic reticulum
stress response and represses the pro-survival XBP1 signaling in
human multiple myeloma cells. Exp Hematol. 39:999–1006. 2011.
View Article : Google Scholar : PubMed/NCBI
|