Introduction
Colorectal cancer is among the leading 25 causes of
global mortality (1). Age
standardized rates for colorectal cancer incidence and mortality
rank fourth as reported by the World Health Organization Globocan
Project. Although early detection and/or treatment have improved
the outcome substantially, a relatively high number of new cases
and deaths are still expected for 2013 (2).
Currently used antineoplastic drugs against
colorectal cancer include antimetabolites (3,4),
alkylating agents (5) and
camptothecin analogs (6). The
antimetabolites, 5-fluorouracil (5-FU) and capecitabine, are
utilized in various combination regimens. The response rate to 5-FU
was shown to increase following leucovorin (LV) modulation but the
survival rate remained unchanged. Addition of either oxaliplatin
(FOLFOX) (7) or irinotecan
(FOLFIRI) (8) to 5-FU and LV
combination led to prolongation of progression-free survival (PFS)
and higher response rates with an acceptable tolerability profile
in patients with metastatic colorectal cancer when used as
first-line therapy. Overall response and survival rates by FOLFOX
and FOLFIRI regimens are similar (9). Regimens in which the alkylating agent
oxaliplatin was added to fluorouracil (FUOX) or capecitabine
(CAPOX) also yielded no significant difference between the two
study arms, and the median survival rate remained between 18 and 21
months (10). Targeted therapies
with monoclonal antibodies against the epidermal growth factor
receptor (EGFR) were found to be correlated with prolonged survival
rates; however, therapeutic efficacy decreased significantly in
patients with K-RAS mutations (11,12).
Anti-VEGF agents such as bevacizumab and aflibercept also
demonstrated beneficial effects (13,14).
Bevacizumab was evaluated in combination with either capecitabine
plus oxaliplatin (XELOX) or with FOLFOX in patients with metastatic
colorectal cancer. Addition of bevacizumab significantly prolonged
PFS; however, overall survival and response rates were not improved
(15). Today the prognosis of
colorectal cancer is predicted by tumor staging. Overall survival
rate in stage IV and recurrent colorectal cancer ranges between 15
and 25 months depending on the combination regimen employed.
Therefore, new drugs or combinations are required to improve the
prognosis of colorectal cancer.
Alkylphosphocholines (APCs) are a novel class of
antineoplastic agents. APCs are structurally related to
alkyllysophospholipids (edelfosine and ilmofosine) with the
exception of the glycerol backbone, which was deemed unnecessary
for cytotoxic activity (16,17).
Unlike classical chemotherapeutics, APCs target the cell membrane
instead of DNA. At clinically relevant doses, they interfere with
phospholipid turnover; hence with cell signaling and survival
pathways (18). Such distinctive
features may facilitate their therapeutic efficacy, and render them
potential candidates for combination therapies (19).
The prototype of APCs, hexadecylphosphocholine
(miltefosine), did not meet expectations due to low response rates
and high gastrointestinal toxicity. Today, miltefosine use is
limited to topical treatment of skin metastases and oral
leishmaniasis treatment (18). By
structural modifications of APCs, many of these pending obstacles
could be overcome. The most recent APCs, erucylphosphocholine
(ErPC) and its homolog erufosine
(erucylphospho-N,N,N-trimethylpropanolamine, ErPC3) are
quite promising with favorable pharmacokinetic and pharmacodynamic
properties. They have significant cytotoxic efficacy but cause less
gastrointestinal toxicity. They possess the ability to cross the
blood-brain barrier and to accumulate in the brain tissue, which
indicates their potential for the treatment of brain tumors
(19). Owing to their structure
with a 22-carbon chain and a ω-9 cis-double bond, they have
reduced myelotoxic and hemolytic effects (20–22)
and because of the latter property, the agents are also the first
intravenously applicable APCs.
In vitro, the antiproliferative effect of
erufosine was demonstrated in various cell lines of human origin
such as chronic myeloid leukemia (CML; alone and when combined with
imatinib) (23), acute myeloid
leukemia (AML) (24,25), chronic lymphocytic leukemia (CLL)
(26), multiple myeloma (MM)
(20,27), bladder carcinoma (28), breast carcinoma (29) and oral squamous cell carcinoma
(30). Bladder carcinoma, AML and
multiple myeloma cell lines were more sensitive to erufosine in
vitro, and the IC50 values ranged between 4 and 14
μM (25,27,28).
However, higher IC50 values (22–41 μM) were detected in
CLL, breast and oral squamous carcinoma cell lines (26,29,30).
Erufosine also reduced colony formation in human MM, breast and
pancreatic carcinoma cells (22)
and inhibited migration in human MM cells (20).
Erufosine induced apoptosis in CLL (26), AML (24,25),
acute lymphocytic leukemia (ALL) (31), human glioblastoma (31–33),
prostate (34) and oral squamous
carcinoma (30) cell lines of human
origin. Its cytotoxic effect was decreased considerably by caspase
inhibitors (25,26). For this reason, part of its
antineoplastic activity was associated with apoptosis (35). Activation of executive procaspase-3
and cleavage of its substrate poly(ADP-ribose) polymerase (PARP)
are well documented time- and concentration-dependent effects of
erufosine (24–26,31–34).
Erufosine-induced apoptosis was modulated via the JNK 1/2,
Raf/MEK/ERK and PI3K/Akt/mTOR signaling pathways (25,27,30,34).
The antineoplastic effects of APCs on colorectal
cancer cell lines have been previously reported (36,37).
Miltefosine was more effective in the colon adenocarcinoma cell
line, HT29 (IC50, 3.1 μmol/l) as compared to mammary
carcinoma cell lines (IC50, 29.4–69.9 μmol/l) (36). Against this colorectal cell line,
miltefosine was more effective when compared to other APC congeners
such as octadecenyl-(trans-9.10)-phosphocholine,
octadecenyl-(cis-9.10)-phosphocholine and
octadecylphosphocholine (IC50, 5.8, 17.8 and 4.4 μmol/l,
respectively) (37). An
antineoplastic effect of erufosine in colorectal cancer cell lines
has not yet been reported. Therefore, the aim of the present study
was to investigate and compare the antiproliferative, antimigratory
and pro-apoptotic effects of erufosine in colorectal cell lines of
human (SW480) and rat (CC531) origin.
Materials and methods
Cell culture
The colon adenocarcinoma cell lines, SW480 (human)
and CC531 (rat), free of pathogenic contamination, were grown as
monolayers in RPMI-1640 medium supplemented with 10% fetal calf
serum (FCS) and L-glutamine (2 mM). The cell lines were maintained
in an incubator with a humidified atmosphere (5% CO2 in
air at 37°C). Cells were passaged two or three times a week to
maintain them in a logarithmic growth phase. For isolation and
propagation, the medium was discarded, and then the cells were
washed with phosphate-buffered saline (PBS), trypsinized (0.25%
trypsin/EDTA), pelleted at 1,500 rpm for 5 min and re-suspended at
the desired concentration in RPMI-1640 medium.
Cell proliferation assay
Cell proliferation was assessed by MTT
(3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide) dye
reduction assay as described by Mosmann, with some modifications
(38). In brief, MTT (Sigma,
Munich, Germany) solution (10 mg/ml in PBS) was added (10 μl/well).
Plates were further incubated for 3 h, and following removal of the
medium, the formazan crystals were dissolved by the addition of 100
μl solvent (0.04 N HCl acid in 2-propanol) per well and then by
thoroughly mixing. Optical density was measured at a 540-nm
wavelength (690 nm reference wavelength) using an ELISA plate
reader (Anthos Mikrosysteme GmbH, Krefeld, Germany). Cell doubling
time (DT) was calculated by using the Patterson formula: Td = T ×
lg2/(lgN2 - lgN1) where Td is the doubling time (in hours), N is
the number of cells and T is the time for cell growth from N1 to
N2. Cell growth rates (in hours) were calculated by the following
formula: Growth rate (μ) = ln
(N2/N1)/T2 - T1.
The optimal cell number to be seeded was determined
before assessment of the antiproliferative effects of erufosine in
both cell lines. For the growth curves, SW480 and CC531 cells were
seeded in 96-well microplates at final concentrations of
2×103, 4×103, 8×103 cells/well and
2×103, 4×103 cells/well, respectively. For
exposure to erufosine, cells were seeded into 96-well microplates
(2×103 cells/100 μl medium/well for SW480 and
4×103 cells/100 μl medium/well for CC531) and incubated
with increasing concentrations (3.1, 6.3, 12.5, 17.7, 25, 35.4, 50,
70.7 and 100 μM) for 24, 48 and 72 h. Cell survival rates were
expressed as the percentage of untreated controls at 24, 48 and 72
h. IC50 was calculated by the equation of logarithmic
regression trendline.
In vitro wound healing (scratch)
assay
SW480 and CC531 cells were seeded (10,000
cells/well) in 24-well plates and allowed to attach to the surface
under standard incubation conditions for 24 h. After 24 h, the
confluent cell monolayers were scratched in a straight line using a
200-μl sterile plastic pipette tip, as previously described
(39). The cells were then
carefully rinsed with culture medium to remove free-floating cells
and debris. Then, erufosine was added at final concentrations of
1.56, 3.125 and 6.5 μM/well, and the effect on wound healing was
monitored. Scratch zones representative for each cell line were
photographed at 12, 24, 36 and 48 h by the Axio Observer.Z1
microscope (Carl Zeiss AG, Oberkochen, Germany). Each experiment
was conducted in triplicate wells for each concentration of
erufosine and the control. AxioVision Rel. 4.8 software was used
for the measurements. For SW480 cells, cells that had migrated into
the scratched area were counted within a 400×400 μm frame, which
was created by the region of interest (ROI) function, enabling to
select 3 random regions in the scratched area. For the CC531 cells,
the distance between the wound edges was measured. For both cell
lines, three random measurements were made per photographed sample
at 12 h, which was used as baseline. Both cell lines were studied
in parallel, and the duration of the microscopic procedure was kept
the same to exclude environmental condition-related differences in
wound healing responses.
Caspase assay
Caspase-3/−7 enzymatic activity was measured by
Apo-ONE Homogeneous Caspase-3/7 assay (G7792 Promega, Germany),
according to the manufacturer’s instructions. Briefly, SW480 and
CC531 cells were seeded at a final concentration of 10,000
cells/well in a black 96-well plate, 24 h before drug treatment.
Erufosine was added to the wells at final concentrations of 50 and
100 μM and incubated for 5 h in an incubator (5% CO2/95%
O2 at 37°C). A standard assay (96-well, 200 μl final
reaction volume) was conducted in triplicate involving three
groups: blank [caspase reagent (CR) + cell culture medium without
cells], negative control (CR + vehicle-treated cell culture) and
assay (CR + treated cell culture). The contents of wells were
gently mixed using a plate-shaker at 300–500 rpm for 5 h at room
temperature (25°C). Immediately after this process, the
fluorescence of each well was measured using a spectrofluorometer
at an excitation wavelength range of 485±20 nm and an emission
wavelength range of 530±25 nm. Blank values were subtracted from
the experimental values to obtain the relative fluorescence units
(RFUs).
Gene expression analysis
CC531 cells were seeded at a density of 200,000
cells in 25-ml flasks. After 24 h, the medium was changed and the
cells were exposed to erufosine (12, 5 and 25 μM) for 48 h. For
osteonectin expression, an additional experiment was conducted as
follows. CC531 cells were seeded at a density of 400,000 cells/well
of a 6-well plate. Medium was changed after 24 h, and the cells
were incubated with erufosine (25 μM) for 24 and 48 h. Then, the
cells were harvested, and the cell pellets were stored at −20°C
until RNA isolation. Total RNA was isolated from CC531 cell pellets
by the RNeasy Mini kit (Qiagen GmBH, Hilden, Germany) and then
first strand cDNA was synthesized by Thermo Scientific Maxima
reverse transcriptase kit (Thermo Scientific GmbH, Schwerte,
Germany) according to the manufacturer’s protocols. Gene expression
was studied by basic PCR protocol (Invitrogen GmbH, Karlsruhe,
Germany). The following rat primers were used: ON,
gagtttggcagctcagagga (left) and tctgct tctgagatgggtca (right);
γ-tubulin, gatggcagtgacagcctagag (left) and gccgttccaagaggtagga
(right); COL1A1, catgttcagctttgtgga cct (left) and
gcagctgacttcagggatgt (right); COL1A2, cctggctctcgaggtgaac (left)
and caatgcccagaggaccag (right). Each gene expression experiment was
repeated twice. γ-tubulin served as the housekeeping gene. PCR
products were analyzed by polyacrylamide gel electrophoresis
(PAGE). Thermo Scientific pUC19 DNA/Mspl (Hpall) Marker was used
for sizing and approximate quantification of the PCR products
(Thermo Scientific). Lanes were framed automatically and
corresponding band intensities were calculated by Quantity One 1-D
Analysis software (Bio-Rad Laboratories Inc., Hercules, CA, USA).
For normalization, the band intensity value of the target gene was
divided by the band intensity of the housekeeping gene
γ-tubulin.
Statistical analysis
The data are presented as mean values ± SD. The
survival rates between the treatment groups were compared by ANOVA
Dunnett’s test. Statistical differences in wound healing and
caspase assays were calculated by one way, single factor ANOVA
test. P-values ≤0.05 were considered to indicate statistically
significant results.
Results
Growth curves of SW480 and CC531
cells
SW480 cells had a doubling time of 22.5±2.8,
20.0±4.1 and 24.3±9.3 h and growth rates of 0.031, 0.035 and 0.031
h−1 when seeded at a density of 2×103,
4×103 and 8×103 cells/well, respectively.
CC531 cells had a doubling time of 18.8±1.0 and 22.6±1.4 h and
growth rates of 0.037 and 0.031 h−1 when seeded at a
density of 2×103 and 4×103 cells/well,
respectively.
Antiproliferative effect of erufosine on
SW480 cells
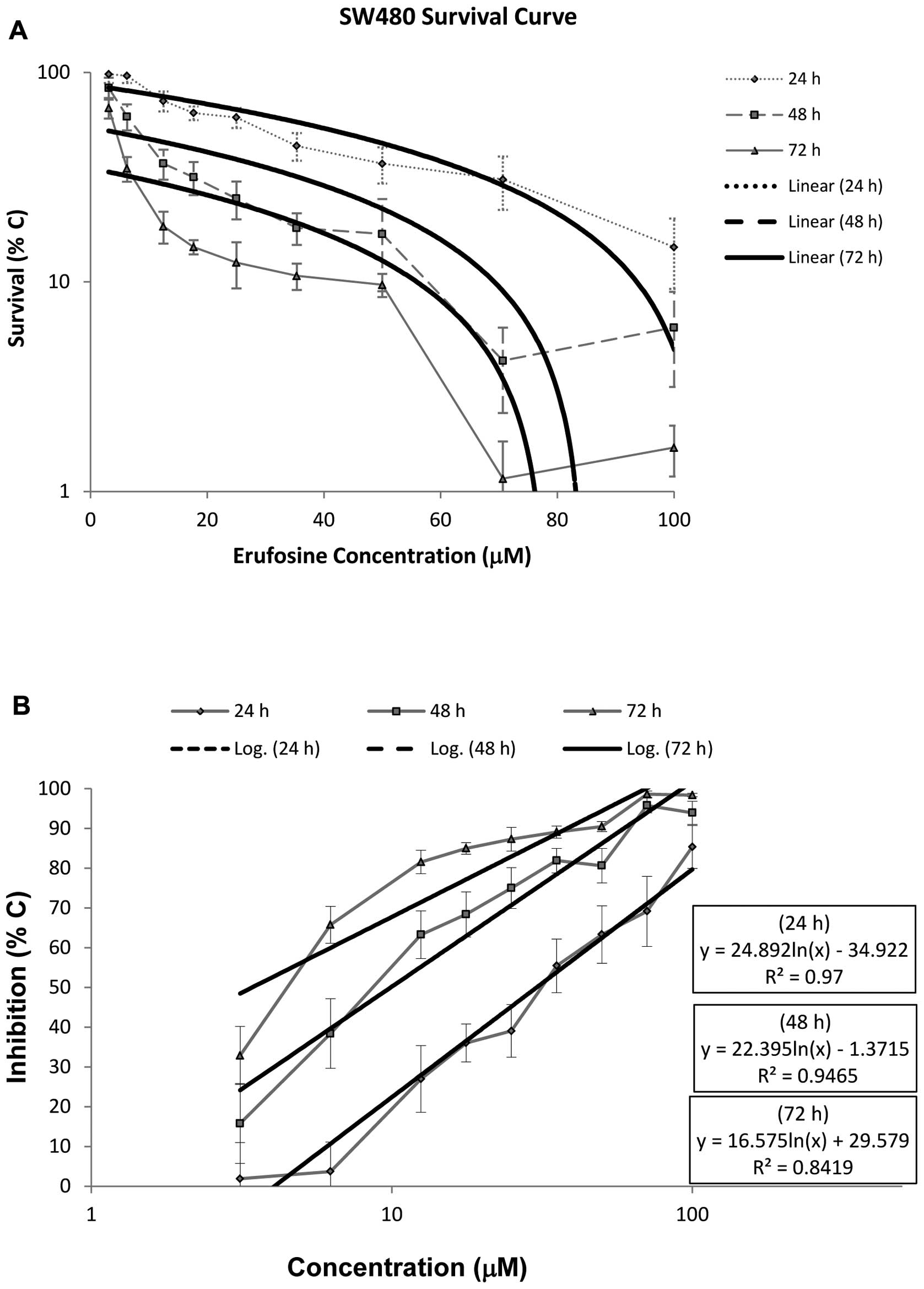
Survival rates of the SW480 cells following exposure
to erufosine are shown in Fig. 1A.
Erufosine exerted a concentration- and time-dependent
antiproliferative effect. This effect was observed after 24 h with
concentrations ≥12.5 μM. However, a significant difference from the
control group was found at all concentrations at 48 and 72 h. The
antiproliferative effect of erufosine significantly increased with
longer exposure times at all concentrations. The IC50
values of erufosine in the SW480 cell line were 30.3, 9.9 and 3.4
μM following 24, 48, and 72 h of incubation, respectively. The
relevant regression equations and their corresponding R2
values are provided in Fig. 1B. In
the dose-response curve, a steep decline in the survival rate was
observed at high erufosine concentrations (70.7 and 100 μM),
preceded by a gradual decrease at low concentrations (Fig. 1A). The shoulder width and the
amplitude became smaller in a time-dependent manner.
Antiproliferative effect of erufosine on
CC531 cells
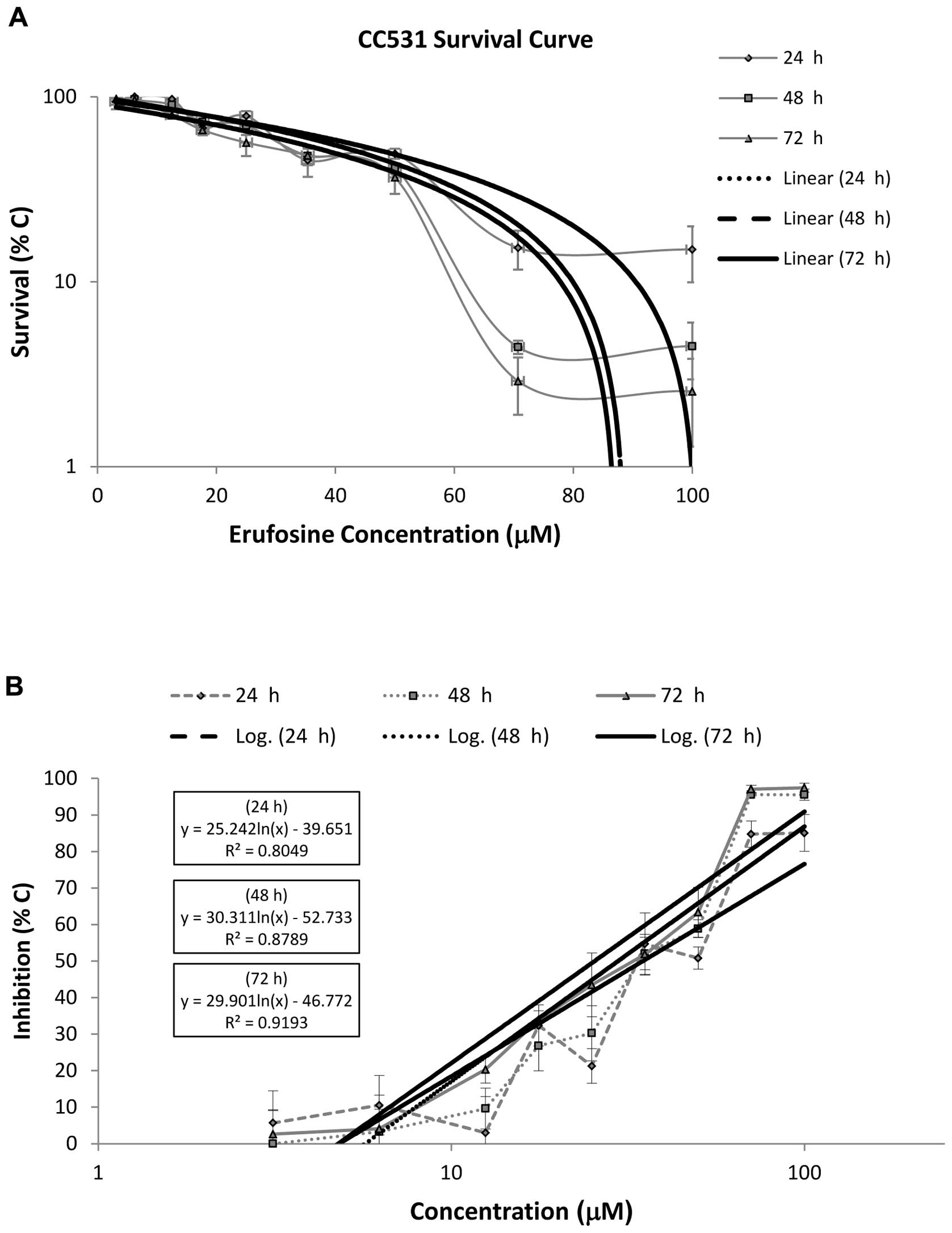
Survival rates of the CC531 cells following exposure
to erufosine are shown in Fig. 2A.
An antiproliferative effect was observed at concentrations ≥25 μM
after 24 h (P=0.003) and 48 h (P<0.0001) of incubation. After 72
h, no significant cytotoxic effect was demonstrated at the
concentrations of 3.125, 6.25 and 12.5 μM (P>0.05). At the
effective concentrations, the survival rates decreased
significantly as the incubation time was prolonged. The
IC50 values of erufosine in the CC531 cell line were
34.9, 29.7 and 25.4 μM following 24, 48 and 72 h of incubation,
respectively. The relevant regression equations and their
corresponding R2 values are provided in Fig. 2B. In the dose-response curve, a
steep decrease in the survival rate was observed at high erufosine
concentrations (70.7 and 100 μM), preceded by a gradual decrease at
low concentrations (Fig. 2A). This
shoulder effect was concentration- and time-dependent. The
amplitude and the width were higher at 48 and 72 h when compared to
the survival curve of the SW480 cells. The shoulder width became
smaller after 48 h. There was no significant difference between the
survival rates for 70.7 and 100 μM after 48 and 72 h. The survival
curves after 48 and 72 h nearly overlapped with each other.
Wound healing assay in SW480 and CC531
cells
The wound healing assay was followed over 48 h for
the low concentrations used in the cytotoxicity assay. Both cell
lines displayed different migratory profiles during wound healing.
SW480 cells migrated to the scratched area and closed the wound by
forming colonies during the ‘healing’ process. In the CC531 cell
culture, the wound edges approached each other to close the
‘wound.’ Untreated SW480 cells showed a time-dependent ability to
close the gap and increasingly more cells migrated into the
scratched area (Fig. 3A). This
difference in migration was statistically significant at 24
(P=0.003), 36 and 48 h (P<0.0001). In SW480 cells, erufosine at
a concentration of 1.56 μM inhibited wound healing, and this effect
was significantly different from the control group after 12
(P=0.001), 24, (P=0.007), 36, (P=0.02) and 48 h (P<0.0001)
(Fig. 3C). This effect observed
following erufosine treatment (1.56 μM) indicated 53.8±13.3%
inhibition of wound healing as compared to the control group at 48
h. In the SW480 cells, apoptotic morphological changes were noted
at the 6.3 μM concentration even after 12 h of incubation and these
signs were also observable, although less distinct, at the 3.1 μM
concentration. Apoptotic cells lost contact with their neighbors,
became rounded and detached from the surface displaying pronounced
membrane blebbing. At a concentration of 1.56 μM, SW480 cells
displayed no apparent signs of apoptosis. CC531 cells also had a
good capacity to fill the gap (Fig.
3B). In the untreated group, the distance between the two edges
of the wound progressively decreased. This decrease became
statistically significant at 36 h (P<0.002), and was more
pronounced at 48 h (P<0.0001), which corresponds to a wound
healing of 62.2%. In the CC531 cells, erufosine inhibited wound
healing capacity, which became evident by 48 h at all
concentrations tested when compared with their corresponding
baseline values (1.56, 3.125 and 6.25 μM; P<0.0001). As compared
to the control following 48 h, the gap widths in the
erufosine-treated cells were 183.5±75.2 (P=0.0025), 207.7±81.6
(P<0.0001) and 221.0±67.2% (P<0.0001) of the untreated cells
following treatment with 1.56, 3.125 and 6.25 μM, respectively
(Fig. 3D). Thus, erufosine-treated
CC531 cells had an ~2-fold wider wound width after 48 h, as
compared to the control group. A significant
concentration-dependent effect was observed at erufosine
concentrations between 1.56 and 6.25 μM at 24 and 36 h,
respectively (P=0.0002). However, at 48 h this
concentration-dependent effect could not be observed any longer
between the treatment groups: 1 vs. 3 μM (P=0.277), 1 vs. 6 μM
(P=0.064) and 3 vs. 6 μM (P=0.168). No morphological signs of
apoptosis were detected in the CC531 cells at test
concentrations.
Caspase assay
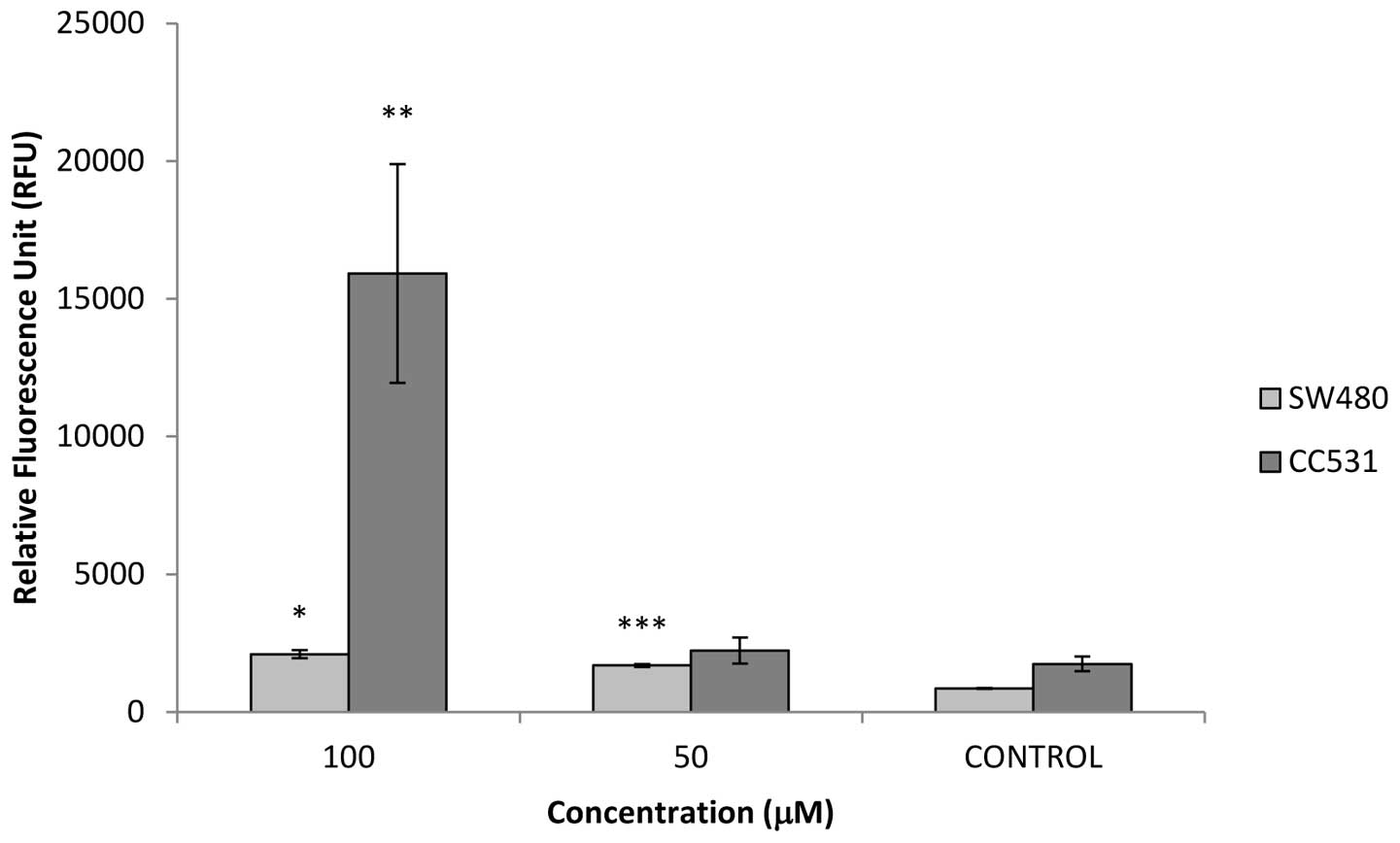
In both cell lines, erufosine induced caspase-3/−7
enzymatic activity (Fig. 4). In the
SW480 cells, erufosine had a concentration-dependent effect on
caspase release. The fluorescence readings (RFU) were 851±18,
1686±55 and 2093±144 for the untreated, 50 and 100 μM
erufosine-treated groups, respectively. This
concentration-dependent response was significantly different from
the control group (P=0.035 for 50 μM and P<0.001 for 100 μM) and
also between both concentrations (P=0.0001). In the CC531 cells,
RFU values were 1746±274, 2232±471 and 15916±3973 for the untreated
and erufosine (50 and 100 μM)-treated groups, respectively. In the
CC531 cells, there was no significant difference between the 50 μM
and the untreated control group, whereas the difference between 100
μM and the untreated group was statistically significant
(P=0.0035).
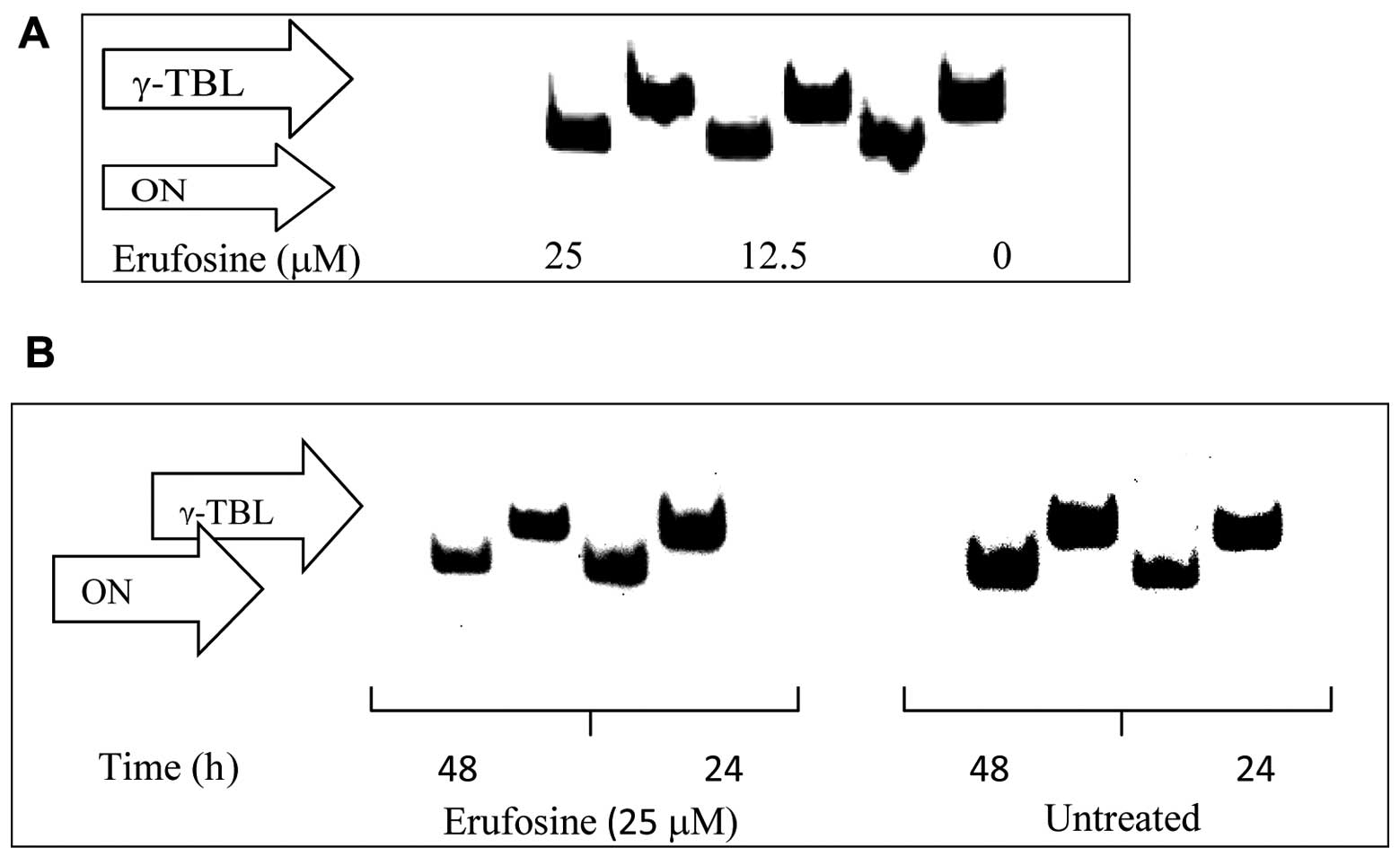
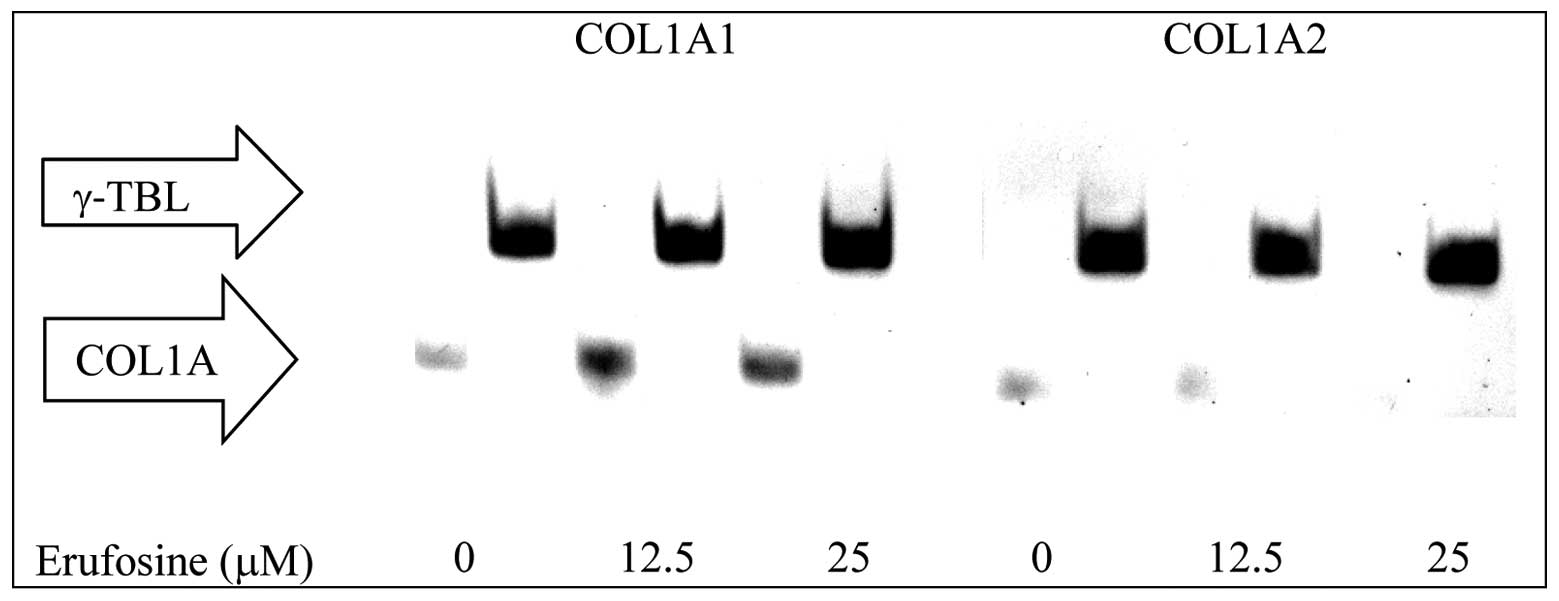
Osteonectin, COL1A1 and A2 expression in
CC531 cells
Erufosine (25 μM) incubation attenuated osteonectin
expression by 12 and 11% following 24 and 48 h of incubation
(Fig. 5). Erufosine (25 μM)
abolished COL1A2 expression after 48 h (Fig. 6) but no change was observed for
COL1A1 (Fig. 6).
Discussion
This is the first report on the activity of
erufosine in colorectal cell lines, SW480 and CC531. Our in
vitro results showed concentration- and time-dependent
antiproliferative effects in both cell lines. After 72 h, similar
growth inhibitory effects were observed at a concentration of 100
μM in both cell lines, but at lower concentrations, erufosine was
significantly more effective in SW480 cells. The IC50
value decreased progressively with prolonged incubation times in
the SW480 cells but such a decline was not observed in the CC531
cells. Following 72 h of exposure to erufosine, the IC50
value for CC531 cells was nearly 4-fold higher than that of the
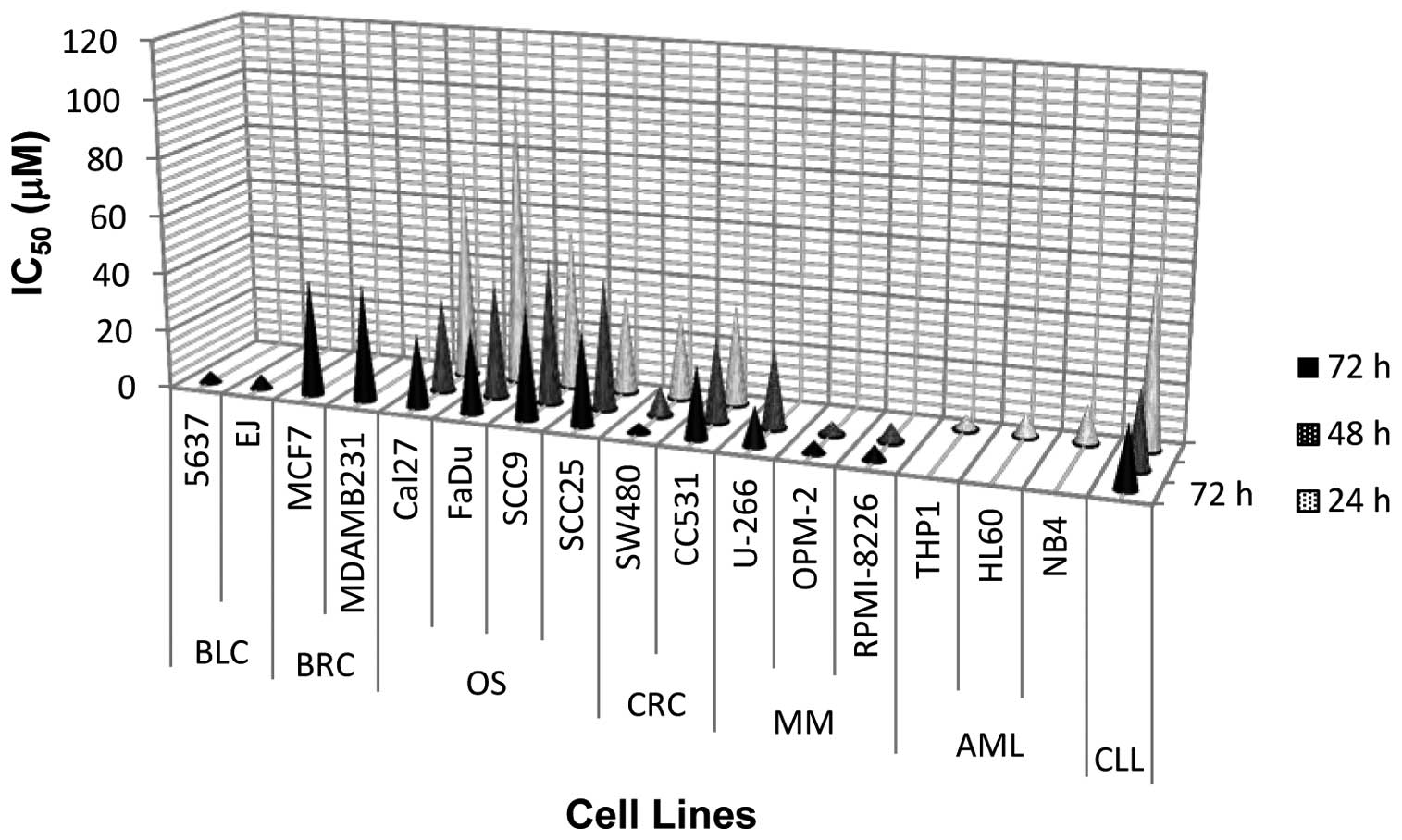
SW480 cells. An antiproliferative effect of erufosine has been
demonstrated in various human-derived cell lines, as outlined in
Fig. 7. When compared to other
types of human cancers, erufosine exhibits marked antiproliferative
activity in SW480 cells, similar to bladder carcinoma and several
(OPM-2 and RPMI-8226) MM cell lines. However, erufosine failed to
show the same degree of efficacy in CC531 cells, and its efficacy
in SW480 cells was only comparable to oral squamous carcinoma and
CLL cells. CC531 cells were also shown to be less sensitive to the
alkylating agent melphalan, when compared to SW480 cells (40).
For SW480 and CC531 cells, shoulder-type survival
curves were demonstrated (Fig. 3A
and 4A). Survival rates declined
gradually between 3.1 and 50 μM but a steep decrease was observed
at high concentrations (70.7 and 100 μM). Shoulders on survival
curves have been explained by two models. In a multi-hit target
model, shoulders are due to the need to hit more than one target
for cell-killing and zero or non-zero initial slopes at low doses
become exponential as higher concentrations are reached. In the
repair model, shoulders indicate that cells can proficiently repair
the damage (41). If the repair
process is saturated with some damage left, the dying process is
initiated (42). Our results
demonstrated marked differences between shoulder-type survivals in
both cell lines. The shoulder had a higher magnitude and a broader
width in CC531 cells. The shoulder magnitudes and final slopes of
the survival curves may be altered by dose-dependent DNA repair
mechanisms and/or interdependence of lethal/mutational responses
(41). Cells display differential
capacities to accumulate and repair sublethal damage. For example,
the survival curve of HeLa cells had a small initial shoulder and a
modest dose-rate effect. However, Chinese hamster cells displayed a
broad shoulder and a large dose-rate effect. Such a difference was
suggested to indicate the dominance of apoptotic cell death in HeLa
cells (43). The shoulders on the
survival curves of CC531 and SW480 cells (Figs. 1A and 2A) may indicate the synthesis of
anti-apoptotic or pro-survival factor(s), which can help to repair
erufosine-induced cellular damage(s). For example, high expression
of the inhibitor of apoptosis protein (IAP) family (survivin, XIAP
and other members) has been associated with colon carcinogenesis
and resistance to chemotherapeutic agents (44). Other pro-survival factors such as
insulin-like growth factor binding proteins, IGFBP3 and 7, CXCL5
and sirtuin 1 were also reported for colorectal cancer cell lines
(45–47). Anti-apoptotic or pro-survival
factor(s) against erufosine-induced cytotoxicity remain to be
identified.
Cell survival depends on the balance between
anti-apoptotic [Ras-Raf-MAPK/ERK and phosphatidylinositol 3′-kinase
(PI3K)/Akt] and pro-apoptotic (SAPK/JNK) signaling mechanisms.
Inhibition of survival pathways is a well-documented effect of APCs
(19). Accordingly, the
pro-apoptotic effect of erufosine has been demonstrated in various
cell lines (24–27,30–35).
Erufosine-induced apoptosis was correlated with
hypo-phosphorylation (activation) of the retinoblastoma (Rb)
protein, which inhibits Abl and JNK kinases as well as E2F
transcription factors (35,48). The mitochondrial death pathway is
another mechanism in erufosine-induced apoptosis (31–33,49).
Caspase-3 was also suggested as one of the major determinants of
erufosine-induced apoptosis (50).
A concentration-dependent activation of caspase-3 and cleavage of
the caspase-3 substrate PARP were detected in prostate cancer (PC3
and LNCaP) cells in response to erufosine treatment (12.5–25 μM)
(34). In another study, erufosine
(10 μM) treatment for 6–12 h resulted in depletion of procaspases
and induced PARP cleavage (92%) in MM cells (OPM-2) (27). In freshly isolated CLL cells,
erufosine (1–100 μM) cleaved PARP totally after 24 h in a
concentration-dependent manner and a pan-caspase inhibitor
completely abrogated apoptosis (26). Erufosine at concentrations of 30 and
50 μM enhanced caspase-3/−7 activity and cleavage of PARP in a
concentration-dependent manner in oral squamous carcinoma cells
(30). Our results showing enhanced
caspase-3/−7 activity in both cell lines confirm these previous
reports. Erufosine-induced caspase-3/−7 activity was
concentration-dependent in SW480 cells, but was observed only at a
high (100 μM) concentration in CC531 cells. Additionally, control
caspase-3/−7 activity was higher (2-fold) in the CC531 than this
activity in the SW480 cells, and this difference grew markedly
(8-fold) following erufosine exposure (100 μM). However, erufosine
(100 μM, 72 h) eventually exhibited the same antiproliferative
effect in both cell lines. Caspase activation is not necessarily
related to apoptosis in colorectal cancer cells (51). Beyond cell death, caspases are
involved in many functions such as cellular formation and
differentiation. Cytokines, tyr protein kinases, Ser/Thr protein
kinases, protein phosphatases and G-proteins have been shown to be
substrates of caspases. For this reason, caspase activation and
cleavage of their substrates cannot be solely interpreted as
irreversible processing of programmed cell death (52). Furthermore, caspase activation is
not the unique determinant in programmed cell death.
Caspase-independent cell death programs include autophagy, mitotic
catastrophe, slow cell death and paraptosis (53). For example, APC-induced cell death
was shown to be BCL-XL-sensitive and caspase-independent in human
malignant glioma cells (54).
According to our results, apoptosis is unlikely to be the major
pathway for programmed cell death in CC531 cells at test
concentrations, as supported by its characteristic survival curve
(broad shoulder, high amplitude and steep slope) and lack of
apoptotic signs during the wound healing assay (43).
APCs were shown to inhibit migration of human
retinal pigment epithelial (RPE) cells by >90% in a
concentration-dependent manner. Inhibition of migration was
correlated with carbon chain length of the APC. For APCs longer
than 20 carbon atoms (>C20), the IC50 value of RPE
cell migration ranged between 0.1 and 1 μM, but increased to 10 μM
for a chain length of 18C (55).
Our wound healing assay showed that the IC50 of
erufosine (22C) ranged from 1.56 to 3.1 μM in both cell lines.
Retarding effects of antineoplastic drugs on wound healing are well
known, but they vary in their efficacy to inhibit collagen
synthesis. Only the DNA alkylating agent cisplatin strongly and
specifically inhibited collagen synthesis in colon fibroblasts
(56). Similarly, the alkylating
agent ET-743 was shown to reduce COL1A1 mRNA levels up to 80% in
scleroderma fibroblasts (57). We
studied the effect of erufosine on COL1A1 and COL1A2 expression in
CC531 cells, which was more prone to the retarding effect of
erufosine on wound healing. Our results demonstrated that erufosine
(25 μM) reduced the COL1A2 band intensity to an undetectable level
but did not have any influence on COL1A1 expression. Osteonectin, a
member of the ‘secreted protein acidic and rich in cysteine’
(SPARC) family of proteins is also involved in wound repair. SPARC
silencing in human tendon fibroblasts did not interfere with cell
proliferation but displayed an antifibrotic effect by reducing
collagen I expression (58). Our
results showed that erufosine (25 μM) caused a slight (12%)
reduction in osteonectin expression.
In light of our findings, erufosine appears to be a
promising chemotherapeutic agent in colorectal cancer. We observed
marked differences in the antiproliferative, delayed wound healing
and apoptotic effects between SW480 and CC531 cells. CC531, an
immortalized cell line from a DMH-induced adenocarcinoma in rats,
was less sensitive to erufosine than SW480 cells of human origin.
Since a single human colorectal cancer cell line is inadequate to
reflect the molecular heterogeneity of clinical colorectal tumors
(59), future research should
include various cell lines of different origins to further
elucidate their survival mechanisms against erufosine.
References
|
1
|
Lozano R, Naghavi M, Foreman K, et al:
Global and regional mortality from 235 causes of death for 20 age
groups in 1990 and 2010: a systematic analysis for the Global
Burden of Disease Study 2010. Lancet. 380:2095–2128. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel R, Naishadham D and Jemal A: Cancer
Statistics 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar
|
|
3
|
Buyse M, Thirion P, Carlson RW,
Burzykowski T, Molenberghs G and Piedbois P: Relation between
tumour response to first-line chemotherapy and survival in advanced
colorectal cancer: a meta-analysis. Lancet. 356:373–378. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hoff PM, Ansari R, Batist G, et al:
Comparison of oral capecitabine versus intravenous fluorouracil
plus leucovorin as first-line treatment in 605 patients with
metastatic colorectal cancer: results of a randomized phase III
study. J Clin Oncol. 19:2282–2292. 2001.
|
|
5
|
Porschen R, Arkenau HT, Kubicka S, et al:
Phase III study of capecitabine plus oxaliplatin compared with
fluorouracil and leucovorin plus oxaliplatin in metastatic
colorectal cancer: a final report of the AIO Colorectal Study
Group. J Clin Oncol. 25:4217–4223. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fuchs CS, Marshall J, Mitchell E, et al:
Randomized, controlled trial of irinotecan plus infusional, bolus,
or oral fluoropyrimidines in first-line treatment of metastatic
colorectal cancer: results from the BICC-C Study. J Clin Oncol.
25:4779–4786. 2007. View Article : Google Scholar
|
|
7
|
de Gramont A, Figer A, Seymour M, et al:
Leucovorin and fluorouracil with or without oxaliplatin as
first-line treatment in advanced colorectal cancer. J Clin Oncol.
18:2938–2947. 2000.
|
|
8
|
Saltz LB, Cox JV, Blanke C, et al:
Irinotecan plus fluorouracil and leucovorin for metastatic
colorectal cancer. N Engl J Med. 343:905–914. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Colucci G, Gebbia V, Paoletti G, et al:
Phase III randomized trial of FOLFIRI versus FOLFOX4 in the
treatment of advanced colorectal cancer: a multicenter study of the
Gruppo Oncologico Dell’Italia Meridionale. J Clin Oncol.
23:4866–4875. 2005.PubMed/NCBI
|
|
10
|
Díaz-Rubio E, Tabernero J, Gómez-España A,
et al: Phase III study of capecitabine plus oxaliplatin compared
with continuous-infusion fluorouracil plus oxaliplatin as
first-line therapy in metastatic colorectal cancer: final report of
the Spanish Cooperative Group for the Treatment of Digestive Tumors
Trial. J Clin Oncol. 25:4224–4230. 2007.
|
|
11
|
Tol J, Koopman M, Cats A, et al:
Chemotherapy, bevacizumab, and cetuximab in metastatic colorectal
cancer. N Engl J Med. 360:563–572. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Van Cutsem E, Peeters M, Siena S, et al:
Open-label phase III trial of panitumumab plus best supportive care
compared with best supportive care alone in patients with
chemotherapy-refractory metastatic colorectal cancer. J Clin Oncol.
25:1658–1664. 2007.PubMed/NCBI
|
|
13
|
Kubicka S, Greil R, André T, et al:
Bevacizumab plus chemotherapy continued beyond first progression in
patients with metastatic colorectal cancer previously treated with
bevacizumab plus chemotherapy: ML18147 study KRAS subgroup
findings. Ann Oncol. 24:2342–2349. 2013. View Article : Google Scholar
|
|
14
|
Van Cutsem E, Tabernero J, Lakomy R, et
al: Addition of aflibercept to fluorouracil, leucovorin, and
irinotecan improves survival in a phase III randomized trial in
patients with metastatic colorectal cancer previously treated with
an oxaliplatin-based regimen. J Clin Oncol. 30:3499–3506. 2012.
|
|
15
|
Saltz LB, Clarke S, Díaz-Rubio E, et al:
Bevacizumab in combination with oxaliplatin-based chemotherapy as
first-line therapy in metastatic colorectal cancer: a randomized
phase III study. J Clin Oncol. 26:2013–2019. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hilgard P, Klenner T, Stekar J and Unger
C: Alkylphosphocholines: a new class of membrane-active anticancer
agents. Cancer Chemother Pharmacol. 32:90–95. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Berkovic D: Cytotoxic ether-phospholipid
analogues. Gen Pharmacol. 31:511–517. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Vink SR, van Blitterswijk WJ, Schellens
JHM and Verheij M: Rationale and clinical application of
alkylphospholipid analogues in combination with radiotherapy.
Cancer Treat Rev. 33:191–202. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Van Blitterswijk WJ and Verheij M:
Anticancer mechanisms and clinical application of
alkylphospholipids. Biochim Biophys Acta. 1831:663–674.
2013.PubMed/NCBI
|
|
20
|
Yosifov DY, Todorov PT, Zaharieva MM,
Georgiev KD, Pilicheva BA, Konstantinov SM and Berger MR:
Erucylphospho-N,N,N-trimethylpropylammonium (erufosine) is a
potential antimyeloma drug devoid of myelotoxicity. Cancer
Chemother Pharmacol. 67:13–25. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Georgieva MC, Konstantinov SM,
Topashka-Ancheva M and Berger MR: Combination effects of
alkylphosphocholines and gemcitabine in malignant and normal
hematopoietic cells. Cancer Lett. 182:163–174. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bagley RG, Kurtzberg L, Rouleau C, Yao M
and Teicher BA: Erufosine, an alkylphosphocholine, with
differential toxicity to human cancer cells and bone marrow cells.
Cancer Chemother Pharmacol. 68:1537–1546. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zaharieva MM, Konstantinov SM, Pilicheva
B, Karaivanova M and Berger MR: Erufosine - a membrane targeting
antineoplastic agent with signal transduction modulating effects.
Ann NY Acad Sci. 1095:182–192. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fiegl M, Lindner LH, Juergens M, Eibl H,
Hiddemann W and Braess J: Erufosine, a novel alkylphosphocholine,
in acute myeloid leukemia: single activity and combination with
other antileukemic drugs. Cancer Chemother Pharmacol. 62:321–329.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Martelli AM, Papa V, Tazzari PL, et al:
Erucylphosphohomocholine, the first intravenously applicable
alkylphosphocholine, is cytotoxic to acute myelogenous leukemia
cells through JNK- and PP2A-dependent mechanisms. Leukemia.
24:687–698. 2010. View Article : Google Scholar
|
|
26
|
Königs SK, Pallascha CP, Lindnerb LH, et
al: Erufosine, a novel alkylphosphocholine, induces apoptosis in
CLL through a caspase-dependent pathway. Leuk Res. 34:1064–1069.
2010.PubMed/NCBI
|
|
27
|
Yosifov DY, Reufsteck C, Konstantinov SM
and Berger MR: Interleukin-6, osteopontin and Raf/MEK/ERK signaling
modulate the sensitivity of human myeloma cells to
alkylphosphocholines. Leuk Res. 36:764–772. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Konstantinov SM and Berger MR: Human
urinary bladder carcinoma cell lines respond to treatment with
alkylphosphocholines. Cancer Lett. 144:153–160. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Dineva IK, Zaharieva MM, Konstantinov SM,
Eibl H and Berger MR: Erufosine suppresses breast cancer in vitro
and in vivo for its activity on PI3K, c-Raf and Akt proteins. J
Cancer Res Clin Oncol. 138:1909–1917. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kapoor V, Zaharieva MM, Das SN and Berger
MR: Erufosine simultaneously induces apoptosis and autophagy by
modulating the Akt-mTOR signaling pathway in oral squamous cell
carcinoma. Cancer Lett. 319:39–48. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lemeshko VV and Kugler W: Synergistic
inhibition of mitochondrial respiration by anticancer agent
erucylphosphohomocholine and cyclosporin A. J Biol Chem.
282:37303–37307. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rübel A, Handrick R, Lindner LH, et al:
The membrane targeted apoptosis modulators erucylphosphocholine and
erucylphosphohomocholine increase the radiation response of human
glioblastoma cell lines in vitro. Radiat Oncol. 1:62006.
|
|
33
|
Veenman L, Alten J, Linnemannstöns K, et
al: Potential involvement of F0F1-ATP(synth)ase and reactive oxygen
species in apoptosis induction by the antineoplastic agent
erucylphosphohomocholine in glioblastoma cell lines: a mechanism
for induction of apoptosis via the 18 kDa mitochondrial
translocator protein. Apoptosis. 15:753–768. 2010.
|
|
34
|
Rudner J, Ruiner CE, Handrick R, Eibl HJ,
Belka C and Jendrossek V: The Akt-inhibitor erufosine induces
apoptotic cell death in prostate cancer cells and increases the
short term effects of ionizing radiation. Radiat Oncol. 5:1082010.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Berger MR, Tsoneva I, Konstantinov SM and
Eibl H: Induction of apoptosis by
erucylphospho-N,N,N-trimethylammonium is associated with
changes in signal molecule expression and location. Ann NY Acad
Sci. 1010:307–310. 2003.PubMed/NCBI
|
|
36
|
Sobottka SB and Berger MR: Assessment of
antineoplastic agents by MTT assay: partial underestimation of
antiproliferative properties. Cancer Chemother Pharmacol.
30:385–393. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sobottka SB, Berger MR and Eibl H:
Structure-activity relationships of four anti-cancer
alkylphosphocholine derivatives in vitro and in vivo. Int J Cancer.
53:418–425. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Mosmann T: Rapid colorimetric assay for
cellular growth and survival: application to proliferation and
cytotoxicity assays. J Immunol Methods. 65:55–63. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Liang CC, Park AY and Guan JL: In vitro
scratch assay: a convenient and inexpensive method or analysis of
cell migration in vitro. Nat Protoc. 2:329–333. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Rothbarth J, Koevoets C, Tollenaar RA,
Tilby MJ, van de Velde CJ, Mulder GJ and Kuppen PJ:
Immunohistochemical detection of melphalan-DNA adducts in colon
cancer cells in vitro and human colorectal liver tumours in
vivo. Biochem Pharmacol. 67:1771–1778. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Haynes RH, Eckardt F and Kunz BA: The DNA
damage-repair hypothesis in radiation biology: comparison with
classical hit theory. Br J Cancer. 49:81–90. 1984.PubMed/NCBI
|
|
42
|
Orr JS: Concepts, problems and the role of
modifying agents in the relationship between recovery of cells’
survival ability and mechanisms of repair of radiation lesions. Br
J Cancer (Suppl). 49:1–6. 1984.PubMed/NCBI
|
|
43
|
Hall EJ and Brenner DJ: Radiobiology of
low- and high-dose-rate brachytherapy. Technical Basis of Radiation
Therapy. 4th edition. Levitt SH, Purdy JA, Perez CA and Vijayakumar
S: Springer Verlag; Berlin: pp. 291–307. 2008
|
|
44
|
Miura K, Fujibuchi W, Ishida K, et al:
Inhibitor of apoptosis protein family as diagnostic markers and
therapeutic targets of colorectal cancer. Surg Today. 41:175–182.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Georges RB, Adwan H, Hamdi H, Hielscher T,
Linnemann U and Berger MR: The insulin-like growth factor binding
proteins 3 and 7 are associated with colorectal cancer and liver
metastasis. Cancer Biol Ther. 12:69–79. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Kawamura M, Toiyama Y, Tanaka K, et al:
CXCL5, a promoter of cell proliferation, migration and invasion, is
a novel serum prognostic marker in patients with colorectal cancer.
Eur J Cancer. 48:2244–2251. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Kabra N, Li Z, Chen L, et al: SirT1 is an
inhibitor of proliferation and tumor formation in colon cancer. J
Biol Chem. 284:18210–18217. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Yosifov DY, Dineva IK, Zaharieva MM,
Konstantinov SM and Berger MR: The expression level of the tumor
suppressor retinoblastoma protein (Rb) ınfluences the antileukemic
efficacy of erucylphospho-N,N,N-trimethylpropylammonium (ErPC3).
Cancer Biol Ther. 6:930–935. 2007.
|
|
49
|
DeGregori J: The Rb network. J Cell Sci.
117:3411–3413. 2004. View Article : Google Scholar
|
|
50
|
Kugler W, Buchholz F, Köhler F, Eibl H,
Lakomek M and Erdlenbruch B: Downregulation of Apaf-1 and caspase-3
by RNA interference in human glioma cells: consequences for
erucylphosphocholine-induced apoptosis. Apoptosis. 10:1163–1174.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Yang SY, Bolvin C, Sales KM, Fuller B,
Seifalian AM and Winslet MC: IGF-I activates caspases 3/7, 8 and 9
but does not induce cell death in colorectal cancer cells. BMC
Cancer. 9:1582009. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Nhan TQ, Liles WC and Schwartz SM:
Physiological functions of caspases beyond cell death. Am J Pathol.
169:729–737. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Bröker LE, Kruyt FAE and Giaccone G: Cell
death independent of caspases: a review. Clin Cancer Res.
11:3155–3162. 2005.
|
|
54
|
Naumann U, Wischhusen J, Weit S, Rieger J,
Wolburg H, Massing U and Weller M: Alkylphosphocholine-induced
glioma cell death is BCL-XL-sensitive, caspase-independent and
characterized by massive cytoplasmic vacuole formation. Cell Death
Differ. 11:1326–1341. 2004. View Article : Google Scholar
|
|
55
|
Eibl KH, Kook D, Priglinger S, Haritoglou
C, Yu A, Kampik A and Welge-Lussen U: Inhibition of human retinal
pigment epithelial cell attachment, spreading, and migration by
alkylphosphocholines. Invest Ophthalmol Vis Sci. 47:364–370. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Hendriks T, Martens MF, Huyben CM and
Wobbes T: Inhibition of basal and TGFJ-induced fibroblast collagen
synthesis by antineoplastic agents. Implications for wound healing.
Br J Cancer. 67:545–550. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Louneva N, Saitta B, Herrick DJ and
Jimenez SA: Transcriptional inhibition of type I collagen gene
expression in scleroderma fibroblasts by the antineoplastic drug
ecteinascidin 743. J Biol Chem. 278:40400–40407. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Seet LF, Su R, Toh LZ and Wong TT: In
vitro analyses of the anti-fibrotic effect of SPARC silencing in
human Tenon’s fibroblasts: comparisons with mitomycin C. J Cell Mol
Med. 16:1245–1259. 2012.PubMed/NCBI
|
|
59
|
Auman JT and McLeod HL: Colorectal cancer
cell lines lack the molecular heterogeneity of clinical colorectal
tumors. Clin Colorectal Cancer. 9:40–47. 2010. View Article : Google Scholar : PubMed/NCBI
|