Introduction
Lung cancer is one of the leading causes of
cancer-related mortality worldwide (1). Non-small cell lung cancer (NSCLC)
accounts for approximately 85% of all lung cancers (2). Although chemotherapy and radiotherapy
have been widely used in the treatment of advanced NSCLC, the
outcome remains unsatisfactory, with low long-term survival rates.
Thus, a more effective and safer therapy for lung cancer is
required (3). As increased
understanding of key cellular pathways involved in tumor growth,
progression and cell death has been achieved, molecular-targeted
therapies have been exploited.
A large number of studies have shown that vascular
endothelial growth factor (VEGF), a receptor tyrosine kinase, plays
an important role in tumorigenesis, and blocking the VEGF signaling
pathway can reduce tumor-associated angiogenesis and blood
vessel-dependent metastasis (4,5).
Therefore, a number of anti-angiogenic drugs blocking the VEGF
signaling pathway (ligand or the receptors) have been developed and
are currently in use in NSCLC therapy. Sorafenib (SOR), one of the
small-molecule inhibitors of receptor tyrosine kinase inhibitors
(RTKIs) targeting the VEGF receptor family, has been shown to have
significant antitumor effects in NSCLC (5). Although it has been shown that
blocking the VEGF signaling pathway inhibits angiogenesis in NSCLC,
the efficacy of these drugs is often limited by unfavorable
pharmacokinetics, low tumor accumulation and other adverse effects
(3). Studies have shown that
anti-VEGF monotherapy does not increase survival in cancer
patients, compared to standard chemotherapy, yet the combination of
these drugs with chemotherapy or other drugs may be an effective
strategy to increase the therapeutic effect in NSCLC (6,7).
Cyclooxygenase-2 (COX-2) signaling is involved in
multiple processes of tumor progression, including proliferation,
survival, angiogenesis and invasion (8). Overexpression of COX-2 has been found
in tissues and cell lines of NSCLC, and the COX-2 inhibitor
displays inhibitory effects in advanced NSCLC patients, although
its therapeutic effect is not more beneficial than chemotherapy
(6). Several combined EGFR and
COX-2 inhibition trials have been completed and have demonstrated
promising results for head and neck squamous cell carcinoma (HNSCC)
(9). Celecoxib (CXB) is a
COX-2-selective non-steroidal anti-inflammatory drug (NSAID), which
was found to exhibit therapeutic effects in various types of
cancer. At present, CXB is widely being tested in clinical trials
for its therapeutic activity against various types of cancers as a
single agent and also in combination with other agents (10,11).
Recently, it has been shown that the combination of SOR and a COX-2
inhibitor provides synergistic anti-proliferative and pro-apoptotic
effects in human liver cancer cells (12,13).
In this context, in the present study, we selected CXB as a COX-2
inhibitor in combination with SOR for suppressing VEGFR and COX-2
expression and simultaneously reducing the doses of both drugs for
treating NSCLC.
The objective of the present study was to evaluate
the feasibility of CXB in combination with SOR for inhibiting NSCLC
cell growth, proliferation and angiogenesis and to reveal the
underlying molecular mechanisms involved in SOR-induced apoptosis.
We also determined whether treatment of CXB, used as an adjuvant
agent, could allow the reduction of the dosage of SOR.
Materials and methods
Reagents
Celecoxib (CBX), one type of COX-2 inhibitor, was
purchased from Pfizer Inc. (New York, NY, USA). Sorafenib (SOR) was
purchased from Bayer Pharma AG (Wuppertal, Germany), and both drugs
were dissolved in dimethyl sulfoxide (DMSO). For western blot
analysis, the following antibodies were used: mouse monoclonal
anti-β-actin (Sigma Aldrich, St. Louis, MO, USA), mouse monoclonal
anti-Bcl2, mouse monoclonal anti-survivin, mouse monoclonal
anti-MEK, mouse monoclonal anti-phosphorylated (p-)MEK, mouse
monoclonal anti-ERK, mouse monoclonal anti-p-ERK and horseradish
peroxidase-conjugated goat anti-rabbit IgG and anti-mouse IgG
antibodies (Santa Cruz Biotechnology, Santa Cruz, CA, USA). Nonidet
P-40 lysis buffer, chemiluminescent peroxidase substrate, propidium
iodide (PI), 4′,6-diamidino-2-phenylindole (DAPI)and
3-(4,5-dimethylthiazol-2-yl)-2,5 diphenyltetrazolium bromide (MTT)
were from Sigma Aldrich; stock solutions of PI, DAPI and MTT were
prepared by dissolving 1 mg of each compound in 1 ml of
phosphate-buffered saline (PBS). The solution was protected from
light, stored at 4°C and used within 1 month.
Cell culture
The A549 human lung adenocarcinoma cells obtained
from the American Type Culture Collection (ATCC; Manassas, VA, USA)
were cultured in RPMI-1640 containing 10% fetal bovine serum (FBS)
in a 5% CO2 incubator and passaged with 0.25% trypsin
and 0.03% ethylenediamine tetraacetic acid (EDTA) solution.
Cell viability assay
A549 cells grown in monolayers were harvested and
dispensed in 96-well culture plates in 100 μl of RPMI-1640 at a
concentration of 5×103 cells/well. After 24 h,
differential drug concentrations of SOR (0–20 μM), CXB (0–40 μM) or
both (0–10 μM SOX plus 20 μM CXB) were added to the cells. Then,
the cells were incubated for another 4 h. At the end of the
treatment, 200 μl of DMSO was added to each well after removing the
supernatant. Then, cell viability was obtained by measuring the
absorbance at a wavelength of 490 nm by Thermo Multiskan MK3
microplate reader (Thermo Fisher Scientific Inc., Waltham, MA,
USA). This assay was carried out in triplicate. The growth
inhibition rate was calculated according to the following formula:
Inhibition rate (%) =[1 − (average absorbance of experimental
group/average absorbance of blank control group)] × 100%.
Detection of apoptosis
A549 cells were cultured in 6-well plates in
RPMI-1640 with 10% FBS medium and were treated with their
respective half maximal inhibitory concentration (IC50)
values of SOR, CXB or both for 48 h. The coverslips were washed
three times with PBS, and single cell suspensions were fixed in 1%
PBS. Cells were stained with 100 μg/ml acridine orange (AO) and 100
μg/ml ethidium bromide (EB) for 1 min. Then cells were observed
under a fluorescence microscope. At least 200 cells were counted,
and the percentage of apoptotic cells was determined. Triplicates
were performed in all experiments, and the experiments were
performed on five occasions. In addition, we also evaluated
survivin and Blc-2 protein expression by western blotting as an
additional indicator of apoptosis.
Cell cycle analysis
To determine the cell cycle distribution,
5×105 A549 cells were plated in 60-mm dishes and treated
with the respective IC50 values of SOR, CXB or both for
48 h. After treatment, the cells were collected by trypsinization,
fixed in 70% ethanol, and kept at −20°C overnight for fixation.
Cells were washed in PBS, resuspended in 1 ml of PBS containing 100
μg/ml RNase and 40 μg/ml PI and incubated in the dark for 30 min at
room temperature. The distribution of cells in the cell cycle
phases was analyzed from the DNA histogram with a FACSCalibur flow
cytometer (Becton-Dickinson, San Jose, CA, USA) and CellQuest
software (San Jose, CA, USA).
Measurement of prostaglandin E2 (PGE2)
production
PGE2 synthesis was determined by competitive
enzyme-linked immunosorbent assay (ELISA) as previously described
(14). In brief, A549 cells were
treated with their respective half maximal inhibitory concentration
(IC50) values of SOR, CXB or both for 48 h in 12-well
plates, and then these culture media were centrifuged to remove
cell debris. Cell-free culture media were collected at the
indicated times, and PGE2 levels were determined by competitive
ELISA as described by the kit manufacturer (Cayman Chemical, Ann
Arbor, MI, USA) using an ELISA reader (μQuant; Bio-Tek Instruments,
Inc., Winooski, VT, USA).
Tumor xenograft assay
All animal experiments were performed in accordance
with the institutional guidelines, following a protocol approved by
the Ethics Committees of the Disease Model Research Center, The
First Hospital of Jilin University. Female BALB mice, 6–7 weeks of
age, were maintained under specific pathogen-free (SPF) conditions
and provided with food and water ad libitum. All the animals
were fed with a normal pellet diet one week prior to the
experimentation.
Exponentially growing A549 cells were harvested, and
a tumorigenic dose of 2.5×106 cells was injected
intraperitoneally into the 6- to 7-week old female BALB mice.
Tumors were allowed to grow in the mice for 10 days, after which
the animals were randomly assigned into one of four treatment
groups (10 mice per group). The control group received 1%
polysorbate resuspended in deionized water. The other three groups
were treated with CXB (4.56 mg/kg body weight), SOX (80 mg/kg body
weight) or CXB plus SOX (2.5 and 40 mg/kg body weight,
respectively) intraperitoneally on alternative days for 3 weeks.
The doses were selected based on previous experiments (15,16).
Tumor weights were measured after the mice were sacrificed, and
tumor volumes were measured before the treatment injections were
administered and on day 7, 14 and 21 of the treatment. On day 22,
the animals were euthanized using chloroform, and their spleen
tissues were collected and cultured for a splenocyte surveillance
study. Furthermore, A549 cells were collected from the site of the
treatment injection for in vivo and ex vivo cell
cycle phase distribution studies.
Assay of splenocyte proliferation
Spleens from the treated mice were collected, and
single-cell spleen suspensions were pooled in serum-free RPMI-1640
by filtering the suspension through a sieve mesh with the aid of a
glass homogenizer to exert gentle pressure on the spleen fragments.
Samples were washed twice in PBS 0.1% (w/v) and bovine serum
albumin (BSA). After centrifugation at 200 × g for 10 min, the
cells were placed into 96-well flat-bottom microplates in
triplicate at 3×103 cells/well in RPMI-1640 supplemented
with 10% FBS. The cells were then incubated in a total volume of
100 μl/well. Serum-free RPMI-1640 was used as a control. After 24
h, cell proliferation was measured by the MTT assay.
Western blot analysis
A549 cells were treated with their respective
IC50 values of SOR, CXB or their combination for 48 h.
The cells were then homogenized in a lysis buffer (Tris-HCl 50
mmol/l, EDTA 5 mmol/l, NaCl 150 mmol/l, sodium deoxycholate 1%,
Na3VO4 500 μmol/l, Triton X-100 0.5%, AEBSF
10 μmol/l, NaF 10 mmol/l) on ice. The homogenates were then
centrifuged at 14,000 rpm at 4°C for 30 min, and the supernatants
were collected for protein concentration determination using the
Bradford reagent (Sigma). Cell extracts (50 μg of protein) were
separated on a sodium dodecyl sulfate-polyacrylamide
electrophoretic gel (SDS-PAGE) and transferred to nitrocellulose
membranes, which were blocked in 3% BSA for 2 h. After blocking,
the membranes were incubated with primary antibodies overnight at
4°C for 2 h, and then with a horseradish peroxidase-conjugated
secondary antibody for 2 h at room temperature. Proteins were
visualized by exposing the chemiluminescence substrate (Sigma) to
X-OMAT AR autoradiography film (Eastman Kodak, Rochester, NY, USA).
Blots were stripped and reprobed with anti-β-actin to control for
loading variations. Quantity One software (Bio-Rad Laboratories)
was used for quantification of the protein bands.
Statistical analysis
Statistical analyses were undertaken using GraphPad
Prism version 5.01 (GraphPad Software, San Diego, CA, USA) and the
SPSS® Statistical Package, version 19.0 (SPSS Inc.,
Chicago, IL, USA) for Windows®. Data are expressed as
the mean ± SD. The statistical significance was determined using
one-way analysis of variance (ANOVA). P<0.05 was considered to
indicate a statistically significant result.
Results
Effects of SOR and CXB alone or in
combination on NSCLC cell growth
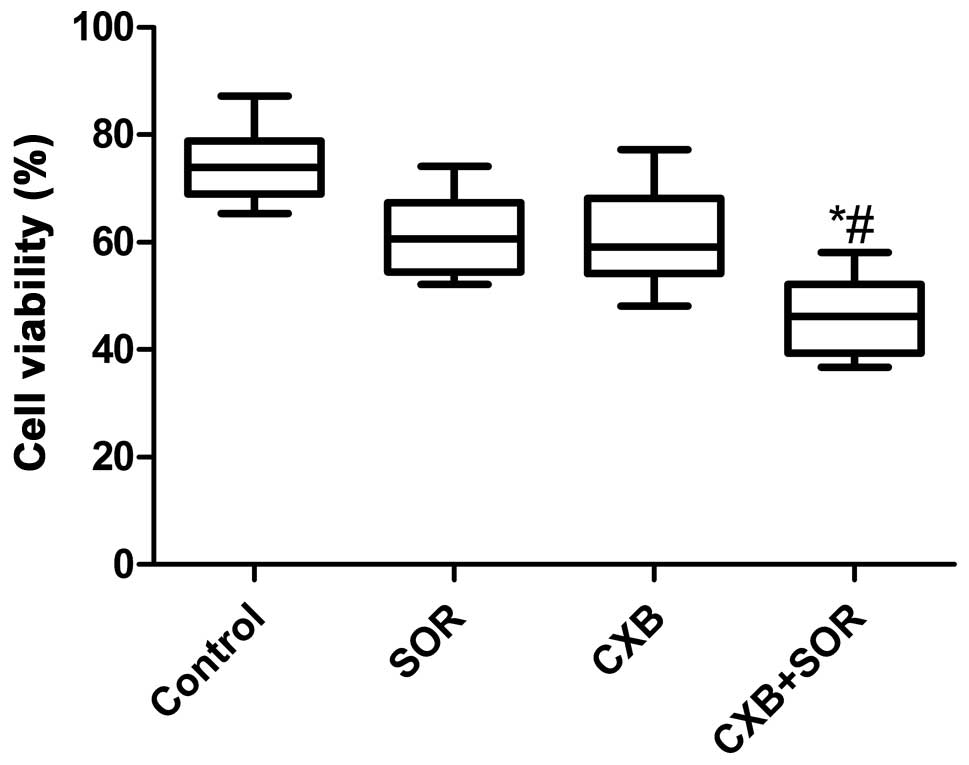
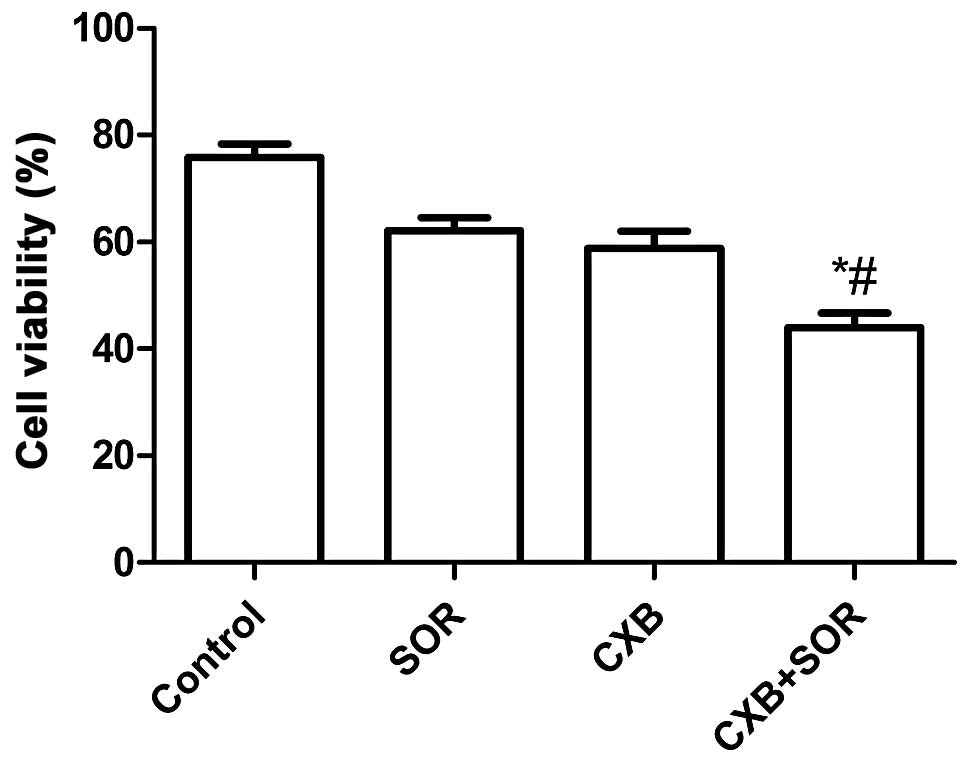
To evaluate the effect of SOR, CXB and their
combination on the cell viability of NSCLC cells in vitro,
A549 cells were treated with increasing concentrations of CXB (0–40
μM) or SOR (0–20 μM). SOR inhibited cell proliferation of A549
cells dose-dependently with an IC50 of 4.4±0.18 μM. CXB
also reduced cell viability of A549 cells in a dose-dependent
manner with an IC50 of 25.5±0.79 μM. Combination
treatment (0–10 μM in the presence of 20 μM CXB) reduced the cell
viability of A549 cells in a dose-dependent manner with an
IC50 of 2.8±0.79 μM. Based on these results, we chose
the respective IC50 values of the drugs for further
treatments throughout the study.
We next examined whether the combination of
relatively low concentrations of SOR and CXB could additively or
synergistically inhibit HCC cell growth in vitro using the
respective IC50 values of SOR and CXB. As shown in
Fig. 1, the inhibitory rate of the
combination treatment was higher than the rates after single drug
treatments (P<0.01). There was no significance different between
the SOR-treated group and CXB-treated group (P>0.05).
Effects of SOR and CXB alone or in
combination on NSCLC cell apoptosis and cell cycle
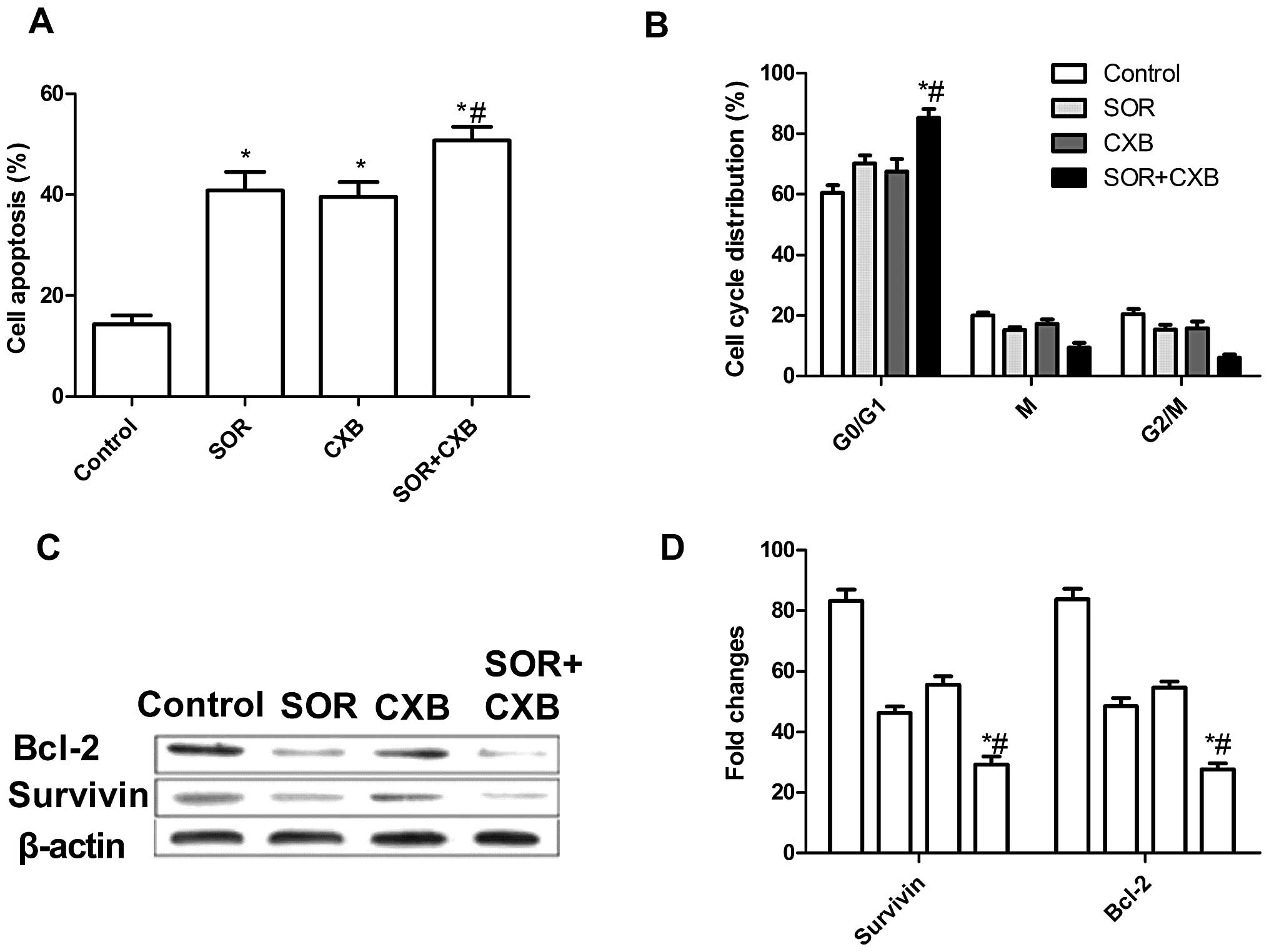
To investigate whether SOR and CXB alone or their
combination could induce apoptosis, we analyzed the apoptosis after
treatment with SOR and/or CXB. Treatment with the combination of
these drugs led to a marked increase in apoptotic cells when
compared to the extent of apoptosis following treatment with the
single drugs (P<0.05) (Fig. 2A).
In addition, there was no significance different between the
SOR-treated group and CXB-treated group in regards to induction of
NSCLC cell apoptosis.
The effects of SOR and CXB on the cell cycle
distribution of A549 cells were then analyzed by flow cytometry.
A549 cells treated with SOR or CXB demonstrated an increased
percentage of apoptotic cells (cell cycle arrest at the G0/G1
phase) compared with the untreated cells (Fig. 2B). The low-dose combination resulted
in an even greater percentage of apoptotic cells when compared with
the percentage of apoptosis following treatment with higher doses
of either drug alone (P<0.01). These data are consistent with
the results from the acridine orange (AO) staining assay. In
conclusion, these results indicate an additive mechanism of SOR and
CXB in inducing cell death through apoptosis.
In order to explore the possible mechanism of the
pro-apoptotic effect of the combination treatment with SOR and CXB,
expression patterns of survivin and Bcl-2 were determined by
western blot analysis. The results (Fig. 2C and D) showed that treatment with
the combination of CXB and SOR significantly decreased the
expression of inhibitor of apoptosis genes, survivin and Bcl-2, in
A549 cells, when compared to the expression levels in cells treated
with CXB or SOR alone (P<0.01).
Effects of SOR and CXB alone or in
combination on the PGE2 production in NSCLC cells
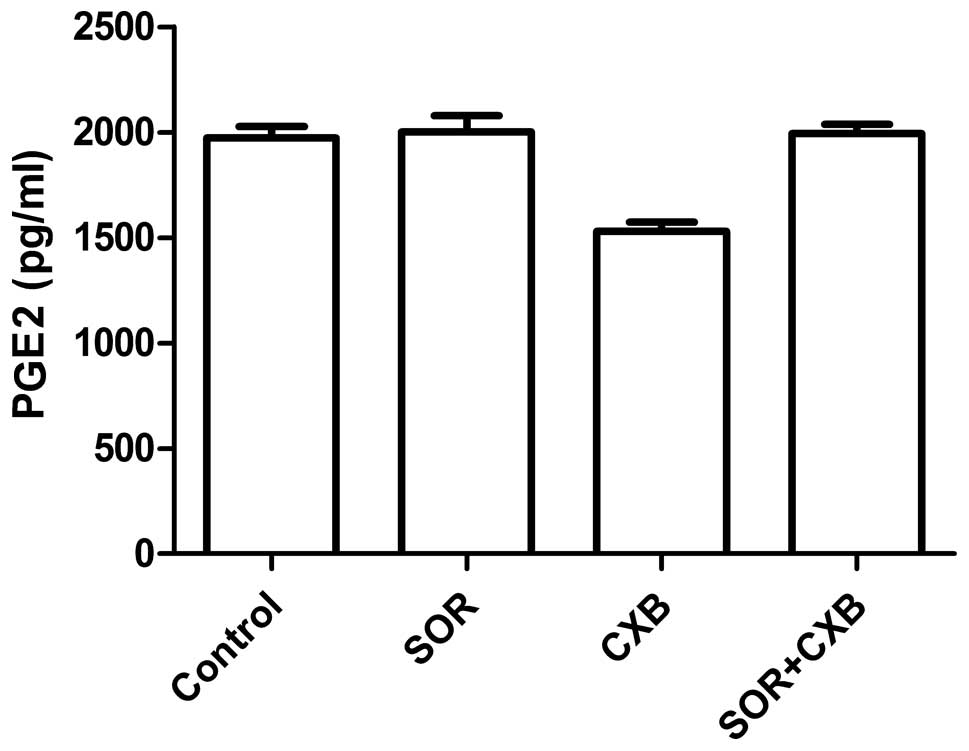
To examine the effect of SOR and CXB on PGE2
production in A549 cell, ELISA was performed. As shown in Fig. 3, CXB inhibited PGE2 production;
however, CXB in combination with SOR or SOR alone did not inhibit
PGE2 production, suggesting that the synergy between CXB and SOR
did not extend to COX-2-dependent PGE2 production in NSCLC
cells.
Effects of SOR and CXB on the RAF/MEK/ERK
signaling pathway in A549 cells
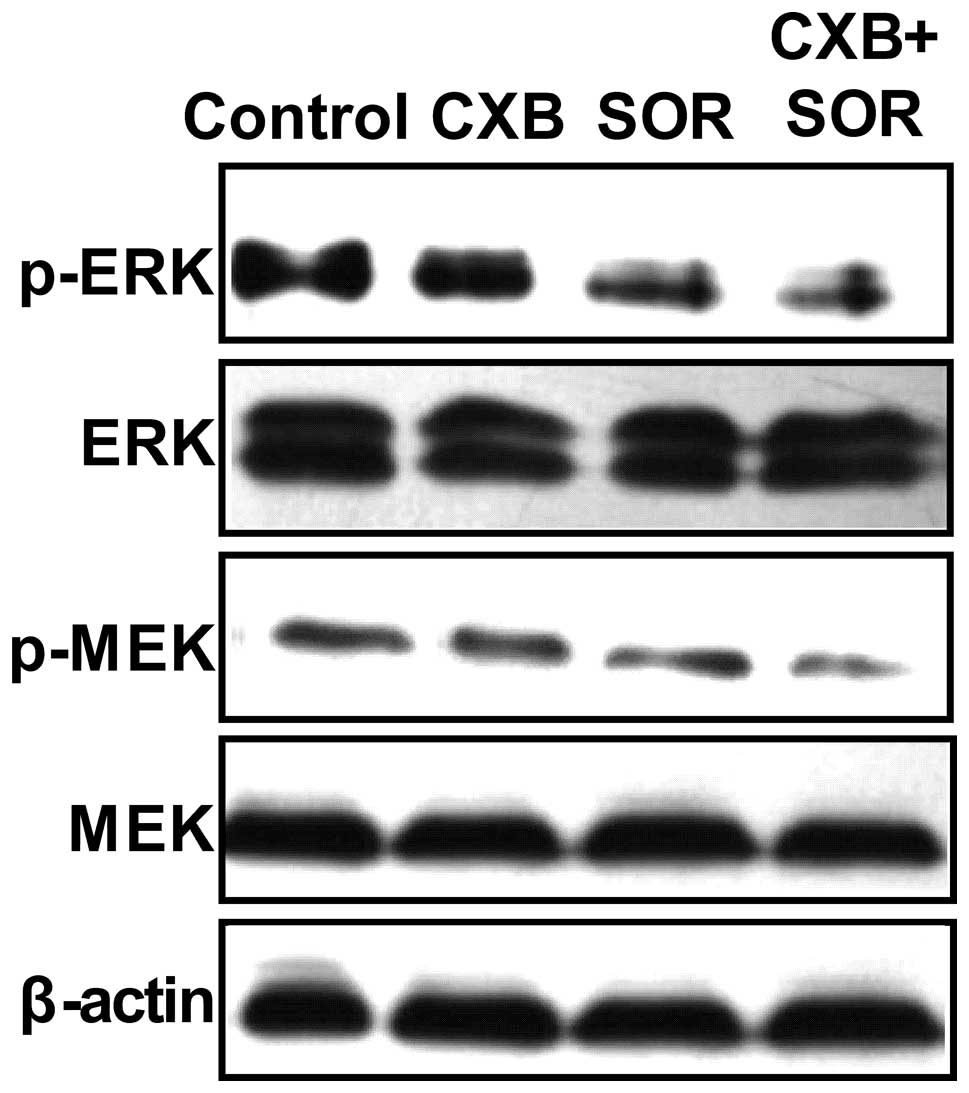
The RAF/MEK/ERK pathway is downstream of Ras
activation, and tyrosine phosphorylation of these proteins is
essential for cancer cell proliferation (17). Therefore, we evaluated the effect of
SOR and CXB alone and in combination on the phosphorylation of
these proteins by western blotting (Fig. 4). We compared the phosphorylation of
these proteins in cells treated with the respective IC50
values of SOR and CXB alone and in combination for 2 h. It was
found that SOR and CXB alone or in combination inhibited the
tyrosine phosphorylation of MEK and ERK (Fig. 4). In addition, following treatment
with the combination of SOR and CXB, a greater reduction in
phospho-MEK and ERK was caused than what was observed by either
agent alone.
SOR plus CXB causes significant
inhibition of tumor growth
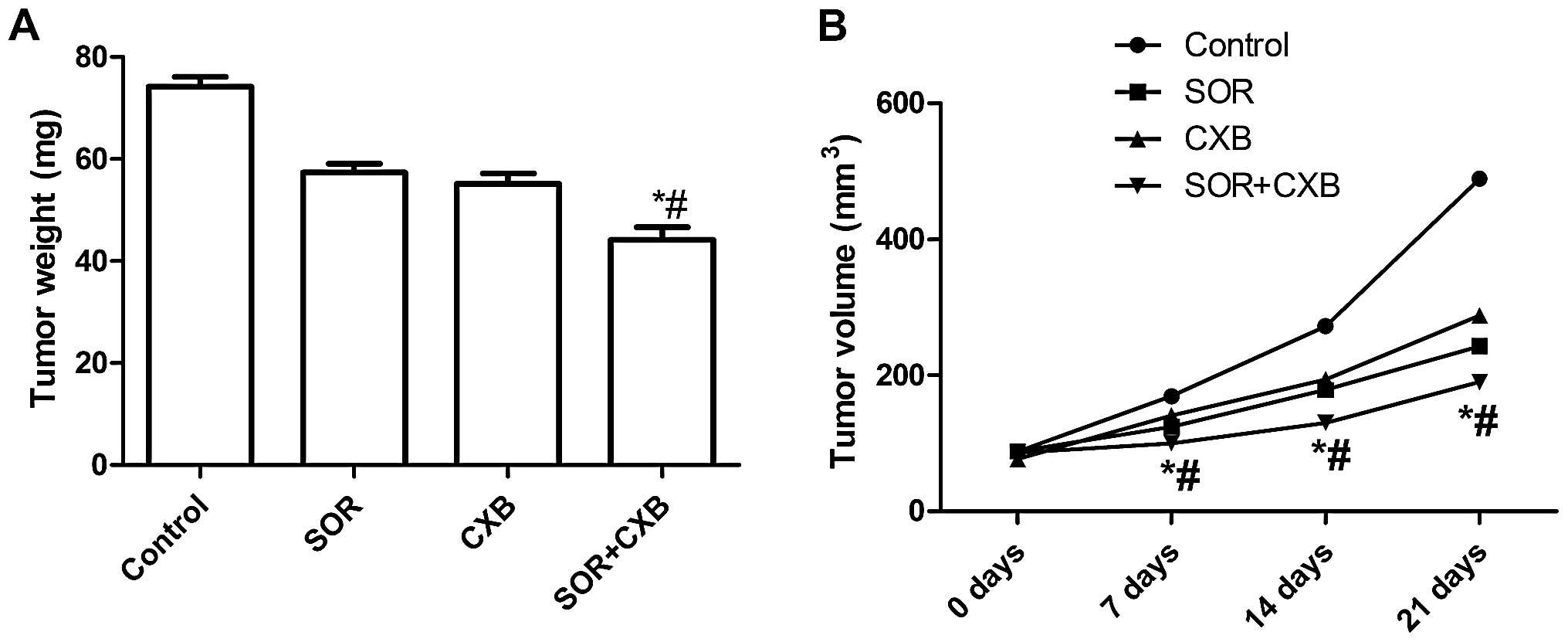
We assessed the in vivo therapeutic efficacy
of SOR and CXB in female BALB mice bearing A549 cell tumors. Mice
were sacrificed and tumor tissue was obtained 21 days after
treatment. The tumor weight of each animal was measured. It was
found that the tumor weight in the mice treated with the
combination of SOR and CXB was lower than the tumor weight of the
untreated group and single drug-treated groups (P<0.01)
(Fig. 5A). In addition, we found
that the tumor growth rate after treatment with the combination of
SOR and CXB was significantly reduced when compared with the single
drug-treated groups and the untreated group (P<0.01) (Fig. 5B). These results indicate that the
combination treatment of SOR and CXB suppressed the tumorigenicity
of A549 cell-derived tumors in mice.
Combination of SOR and CXB inhibits
splenocyte proliferation
To assess the efficacy of SOR and CXB in modulating
splenocyte proliferation, spleen cells of the treated A549 mice
were isolated and cultured in RPMI-1640 containing 10% fetal bovine
serum for 24 h and subjected to in vitro proliferation
assays using MTT assay. As shown in Fig. 6, the inhibitory growth rate of cell
proliferation following treatment with the combination of SOR and
CXB was higher than this rate in the single drug-treated groups,
which demonstrated that the combination treatment inhibited A549
cell proliferation.
Discussion
Recently, studies have shown that VEGF plays an
important role in angiogenesis of NSCLC, and blocking the VEGF
signaling pathway is an effective means to inhibit angiogenesis in
animal models of NSCLC and in NSCLC patients. Although targeting
the VEGF signaling pathway has been shown to be a promising
antitumor strategy, treatment effectiveness remains unsatisfactory
(18). COX-2 inhibitors have been
demonstrated to have an antitumor effect in NSCLC cells and
patients (6). Recently, studies
have shown that a combination with different drugs to treat tumor
patients may increase the efficiency of the antitumor response. In
the present study, we hypothesized that treatment with the
combination of CBX and SOR could enhance the inhibitory effect on
NSCLC cells. The results showed that the combination of CBX and SOR
significantly increased the growth inhibition rate and the
apoptosis rate of A549 cells.
Previous studies have shown that SOR alone
significantly inhibits the growth of NSCLC cells, and its
combinations with other drugs, such as gemcitabine and erlotinib,
could increase the inhibitory effect on NSCLC cells or models
(19,20). COX-2 inhibitors, CBX or N398, have
also been shown to promote the apoptosis of NSCLC and other cancer
cells (21,22). Since COX-2 inhibition promotes
interferon (IFN)-γ-dependent enhancement of antitumor responses,
the role of COX-2 inhibitors in advanced NSCLC has been evaluated
in numerous studies (6,21). However, COX-2 inhibition monotherapy
did not enhance the treatment effectiveness, while its combination
with chemotherapy or other target drugs exhibited a synergistic
antitumor effect (6). To the best
of our knowledge, the results of the present study first
demonstrated that CBX and SOR provide synergistic
anti-proliferative and pro-apoptotic effects in NSCLC cells.
Recently, the synergistic anti-proliferative and pro-apoptotic
effects of CBX and SOR were confirmed in human liver cancer cells
(10). Although the results are
encouraging, the antitumor effects must be further investigated in
NSCLC models and patients.
In order to explore the possible mechanism of the
synergistic effects of CBX and SOR against NSCLC cells, the
expression of survivin and Bcl-2 was assessed using western blot
analysis. The results showed that the combination of CBX and SOR
significantly decreased the expression of these critical inhibitor
of apoptosis proteins. It is well known that multiple genetic
pathways are involved in the regulation of cell apoptosis. Survivin
and Bcl-2 are critical inhibitors of apoptosis proteins. Survivin
inhibits cell apoptosis by acting on caspase directly or
indirectly. Previous studies have demonstrated the expression of
COX-2 and its prognostic significance in NSCLC (23). The COX-2 inhibitor alone was found
to inhibit the expression of survivin and Bcl-2 in NSCLC cells
(22). Similarly, our results
showed that CBX or SOR alone decreased the expression of these two
proteins, and the combination of CBX and SOR further decreased the
protein expression. This result is consistent with the results of
cell proliferation and apoptosis. In addition, the synergistic
effect of CBX and SOR may be due to the enhancement of inhibitory
action on the VEGF signaling pathway by the COX-2 inhibitor. It has
been shown that treatment with a COX-2 inhibitor leads to
restricted angiogenesis and decreased production of VEGF (24). RNAi-mediated knockdown of COX-2 was
found to inhibit the growth and expression of VEGF in human
osteosarcoma (25).
Accumulating evidence shows that the role of CBX
when administered in combination with other drugs in cancer therapy
is modulatory rather than therapeutic, and the efficacy of this
approach has been reported for various types of cancers (13,26–28).
Our results indicated that PGE2 was not involved in the combination
effect of CBX with SOR in the present study, which further
confirmed this concept.
MAPKs (mitogen-activated protein kinases) including
extracellular signal-regulating kinase (ERK), p38 MAPK and c-Jun
N-terminal protein kinase (JNK) play important roles in cell
proliferation, apoptosis, and many other nuclear events (29). Accumulating evidence indicates that
alterations of the activities of MAPKs are involved in the effects
of antitumor agents in various cancer cell lines (17,30–32).
Therefore, targeting of the MAPK pathways is a promising strategy
for NSCLC treatment. In the present study, to the best of our
knowledge, we first examined the intracellular signaling pathway in
A549 cells following treatment with the combination of SOR and CXB.
Our results showed that SOR combined with CXB inhibited the
Ras/Raf/MAPK pathway in A549 cells, which is in agreement with
previous studies that SOR inhibits cancer cell growth through
phosphorylation of p-MEK (12,13,33,34).
Thus, the combination of SOR with CXB may target the Ras/Raf/MAPK
pathway and this targeted approach may underlie the synergistic
effects revealed here.
In conclusion, the present study showed that SOR in
combination with CXB enhances the anti-proliferative and
pro-apoptotic effects on NSCL cells via actions on the
anti-apoptotic RAS/RAF signaling pathways. Celecoxib strengthens
the anti-proliferative action of sorafenib, promoting NSCLC cell
apoptosis and allowing for the use of lower doses of sorafenib than
those currently used. Therefore, it is worthwhile to consider this
combination treatment for NSCLC and warrants further evaluation in
clinical trials.
Acknowledgements
The present study was supported by the Science and
Technology Research and Innovation team funded by Jilin Province
(JL2012058).
References
|
1
|
Mendez M, Custodio A and Provencio M: New
molecular targeted therapies for advanced non-small-cell lung
cancer. J Thorac Dis. 3:30–56. 2011.PubMed/NCBI
|
|
2
|
Jemal A, Siegel R, Ward E, Hao Y, Xu J and
Thun MJ: Cancer statistics, 2009. CA Cancer J Clin. 59:225–249.
2009. View Article : Google Scholar
|
|
3
|
Roy M, Luo YH, Ye M and Liu J: Nonsmall
cell lung cancer therapy: insight into multitargeted small-molecule
growth factor receptor inhibitors. Biomed Res Int.
2013:9647432013.PubMed/NCBI
|
|
4
|
Majeti BK, Lee JH, Simmons BH and Shojaei
F: VEGF is an important mediator of tumor angiogenesis in malignant
lesions in a genetically engineered mouse model of lung
adenocarcinoma. BMC Cancer. 13:2132013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen CT and Hung MC: Beyond anti-VEGF:
dual-targeting antiangiogenic and antiproliferative therapy. Am J
Transl Res. 5:393–403. 2013.PubMed/NCBI
|
|
6
|
Dempke WC, Suto T and Reck M: Targeted
therapies for non-small cell lung cancer. Lung Cancer. 67:257–274.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li J, Pan YY and Zhang Y: Sorafenib
combined with gemcitabine in EGFR-TKI-resistant human lung cancer
cells. Oncol Lett. 5:68–72. 2013.PubMed/NCBI
|
|
8
|
Khan Z, Khan N, Tiwari RP, Sah NK, Prasad
GB and Bisen PS: Biology of Cox-2: an application in cancer
therapeutics. Curr Drug Targets. 12:1082–1093. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kao J, Sikora AT and Fu S: Dual EGFR and
COX-2 inhibition as a novel approach to targeting head and neck
squamous cell carcinoma. Curr Cancer Drug Targets. 9:931–937. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wei D, Wang L, He Y, Xiong HQ, Abbruzzese
JL and Xie K: Celecoxib inhibits vascular endothelial growth factor
expression in and reduces angiogenesis and metastasis of human
pancreatic cancer via suppression of Sp1 transcription factor
activity. Cancer Res. 64:2030–2038. 2004. View Article : Google Scholar
|
|
11
|
Jeon YW and Suh YJ: Synergistic apoptotic
effect of celecoxib and luteolin on breast cancer cells. Oncol Rep.
29:819–825. 2013.PubMed/NCBI
|
|
12
|
Cervello M, Bachvarov D, Lampiasi N, et
al: Novel combination of sorafenib and celecoxib provides
synergistic anti-proliferative and pro-apoptotic effects in human
liver cancer cells. PLoS One. 8:e655692013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Morisaki T, Umebayashi M, Kiyota A, et al:
Combining celecoxib with sorafenib synergistically inhibits
hepatocellular carcinoma cells in vitro. Anticancer Res.
33:1387–1395. 2013.PubMed/NCBI
|
|
14
|
Tai MH, Weng CH, Mon DP, Hu CY and Wu MH:
Ultraviolet C irradiation induces different expression of
cyclooxygenase 2 in NIH 3T3 cells and A431 cells: the roles of
COX-2 are different in various cell lines. Int J Mol Sci.
13:4351–4366. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fulzele SV, Chatterjee A, Shaik MS,
Jackson T and Singh M: Inhalation delivery and anti-tumor activity
of celecoxib in human orthotopic non-small cell lung cancer
xenograft model. Pharm Res. 23:2094–2106. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Murakami M, Zhao S, Zhao Y, et al:
Increased intratumoral fluorothymidine uptake levels following
multikinase inhibitor sorafenib treatment in a human renal cell
carcinoma xenograft model. Oncol Lett. 6:667–672. 2013.
|
|
17
|
Boutros T, Chevet E and Metrakos P:
Mitogen-activated protein (MAP) kinase/MAP kinase phosphatase
regulation: roles in cell growth, death, and cancer. Pharmacol Rev.
60:261–310. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Das M and Wakelee H: Targeting VEGF in
lung cancer. Expert Opin Ther Targets. 16:395–406. 2012. View Article : Google Scholar
|
|
19
|
Pasqualetti G, Ricciardi S, Mey V, Del
Tacca M and Danesi R: Synergistic cytotoxicity, inhibition of
signal transduction pathways and pharmacogenetics of sorafenib and
gemcitabine in human NSCLC cell lines. Lung Cancer. 74:197–205.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Giovannetti E, Labots M, Dekker H, et al:
Molecular mechanisms and modulation of key pathways underlying the
synergistic interaction of sorafenib with erlotinib in
non-small-cell-lung cancer (NSCLC) cells. Curr Pharm Des.
19:927–939. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ferrario A, Fisher AM, Rucker N and Gomer
CJ: Celecoxib and NS-398 enhance photodynamic therapy by increasing
in vitro apoptosis and decreasing in vivo
inflammatory and angiogenic factors. Cancer Res. 65:9473–9478.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Qiu R, Chen J, Sima J, Shen X, Liu D and
Shen J: NS398 induces apoptosis in non-small cell lung cancer
cells. J Cancer Res Clin Oncol. 138:119–124. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang LQ, Wang J, Jiang F, Xu L, Liu FY
and Yin R: Prognostic value of survivin in patients with non-small
cell lung carcinoma: a systematic review with meta-analysis. PLoS
One. 7:e341002012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kirkpatrick K, Ogunkolade W, Elkak A, et
al: The mRNA expression of cyclo-oxygenase-2 (COX-2) and vascular
endothelial growth factor (VEGF) in human breast cancer. Curr Med
Res Opin. 18:237–241. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhao Q, Wang C, Zhu J, et al:
RNAi-mediated knockdown of cyclooxygenase2 inhibits the growth,
invasion and migration of SaOS2 human osteosarcoma cells: a case
control study. J Exp Clin Cancer Res. 30:262011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Falandry C, Canney PA, Freyer G and Dirix
LY: Role of combination therapy with aromatase and cyclooxygenase-2
inhibitors in patients with metastatic breast cancer. Ann Oncol.
20:615–620. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Choe MS, Zhang X, Shin HJ, Shin DM and
Chen ZG: Interaction between epidermal growth factor receptor- and
cyclooxygenase 2-mediated pathways and its implications for the
chemoprevention of head and neck cancer. Mol Cancer Ther.
4:1448–1455. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Buchanan FG, Holla V, Katkuri S, Matta P
and DuBois RN: Targeting cyclooxygenase-2 and the epidermal growth
factor receptor for the prevention and treatment of intestinal
cancer. Cancer Res. 67:9380–9388. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chong H, Vikis HG and Guan KL: Mechanisms
of regulating the Raf kinase family. Cell Signal. 15:463–469. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Anjum R and Blenis J: The RSK family of
kinases: emerging roles in cellular signalling. Nat Rev Mol Cell
Biol. 9:747–758. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
31
|
Roberts PJ and Der CJ: Targeting the
Raf-MEK-ERK mitogen-activated protein kinase cascade for the
treatment of cancer. Oncogene. 26:3291–3310. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cuevas BD, Abell AN and Johnson GL: Role
of mitogen-activated protein kinase kinase kinases in signal
integration. Oncogene. 26:3159–3171. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Peng CL, Guo W, Ji T, et al: Sorafenib
induces growth inhibition and apoptosis in human synovial sarcoma
cells via inhibiting the RAF/MEK/ERK signaling pathway. Cancer Biol
Ther. 8:1729–1736. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lu X, Tang X, Guo W, Ren T and Zhao H:
Sorafenib induces growth inhibition and apoptosis of human
chondrosarcoma cells by blocking the RAF/ERK/MEK pathway. J Surg
Oncol. 102:821–826. 2010. View Article : Google Scholar : PubMed/NCBI
|