Introduction
Laryngeal squamous cell carcinoma (LSCC) is a very
common malignant neoplasm of the head and neck. One of the key
challenges in the treatment of LSCC is the management of metastasis
to locoregional lymph nodes. Lymph node metastasis is common in
supraglottic carcinoma and is the main cause of mortality for these
patients. It has been reported that the incidence of lymph node
metastasis may be as high as 25–50% in supraglottic carcinoma
(1). Current treatments, including
surgical intervention, radiation therapy and chemotherapy, have a
moderate effect on early-stage cases, but are less effective in
more advanced cases. Five-year overall survival for supraglottic
cancer remains poor (2,3). Therefore, understanding the molecular
pathways of carcinogenesis or progression is key to improving
diagnosis, therapy and prevention of supraglottic carcinoma.
microRNAs (miRNAs) are a class of small, non-coding
RNAs that are endogenously expressed in animal and plant cells.
They regulate the expression of protein-coding genes at the
translational level. One strand of the mature double-stranded miRNA
is incorporated into the RNA-induced silencing complex, which
downregulates target mRNAs either by degradation or by
translational inhibition (4).
miRNAs play important roles in normal regulation of gene expression
for developmental timing, cell proliferation and apoptosis.
Moreover, altered miRNA expression is implicated in cancer.
Recently, miRNA genes were implicated in several types of cancer
(5–7). The expression of miRNAs varies between
cancer and normal cells and it also varies among different types of
cancer. It has been shown that some miRNAs are aberrantly expressed
in several different types of cancer (8–10),
suggesting that they may play a role as a novel class of oncogenes
or tumor-suppressor genes.
Several target genes have been experimentally
identified for some miRNAs in various LSCCs; however, the global
pattern of cellular functions and pathways affected by miRNAs in
supraglottic cancer remains elusive. In the present study,
expression profiling of miRNAs in clinical supraglottic carcinoma
tissue samples was carried out, revealing the relationship between
miRNA expression and supraglottic carcinoma. We also studied the
candidate expression of genes that regulate supraglottic carcinoma
miRNA processing and identified the possible gene ontology,
pathway, evolution relationship of these candidate target genes.
This information may be the basis of identifying a clinically
applicable diagnostic tool in the future.
Materials and methods
Laryngeal carcinoma specimens and RNA
isolation
Ten supraglottic carcinoma specimens and ten
adjacent normal tissues were collected by surgical resection from
the First Affiliated Hospital of Harbin Medical University between
January and December 2008. These specimens were from patients
between 42 to 78 years old, including 6 males and 4 females. Lymph
metastasis was found in three cases. Prior to surgery, no patients
were treated by radiotherapy or chemotherapy and they were in good
condition without cancer transmission found from other parts of
their bodies.
The samples were snap-frozen in liquid nitrogen and
stored at −80°C. Total RNA was isolated from adjacent normal tissue
and tumor tissues using the TRIzol reagent (Invitrogen, Carlsbad,
CA, USA) according to the manufacturer’s protocol. RNA was
quantified using the BioPhotometer (Eppendorf), aliquoted, and
stored at −80°C briefly until needed.
Microarray profiling
miRNA was synthesized, amplified and purified using
the Illumina TotalPrep RNA Amplification kit (Ambion Inc.),
following the manufacturer’s recommendations. Total RNA (500 ng)
was sent for miRNA profiling studies using the Human miRNA BeadChip
(V.12.0) in Illumina BeadStation 500GX (Illumina, Inc.), which is
single-channel format according to the standard operating
procedures of the company. RNA was reverse transcribed. After
second strand synthesis, the cDNA was transcribed in vitro
and ncRNA labelled with biotin-16-UTP. Labelled probe hybridization
to BeadChip was carried out. Briefly, poly-A tail was added, cDNA
synthesis (biotinylated) followed, combined with miRNA specific
oligo, then extension and ligation. PCR (Mastercycler 5333), lable,
hybridization (45°C, 14–20 h), wash and imaging were carried our.
Human miRNA BeadChip (V.12.0) contains 1,145 probes specific to
human miRNA assessed at ~30 different beads on average (~850 probes
from Sanger miRBase V.12.0 and 296 from literature or novel content
identified with Illumina Solexa sequencing). The Cy3 fluorescence
on the arrays was scanned at an excitation wavelength of 532 nm
using a BeadArray Reader GX scanner (Illumina). Illumina BeadStudio
software (version 1.5.0.34) was used for preliminary data analysis.
Several quality control procedures were implemented to assess the
quality of the whole experiment. Total RNA control samples were
analyzed in the process. The Illumina BeadStudio software was used
to view control summary reports and scatter plots. The scatter
plots indicated a reduction in assay performance and highlighted
samples that were of lower quality. The control summary report is
generated by the BeadStudio software, which evaluates the
performance of the built-in controls of the BeadChips in the
process. This allows the user to look for variations in signal
intensity, hybridization signal, background signal and the
background to noise ratio for all samples analyzed in that run.
Data are expressed as log2 ratios of fluorescence intensities of
the experimental and the common reference sample. The Illumina data
were then normalized using the ‘normalize quantiles’ function in
the BeadStudio software.
miRNA expression analysis
Cluster analysis was performed using the metric
Euclidean distance to compute the distance matrix for supraglottic
carcinoma patients based on the expression of 1,145 differentially
expressed miRNAs. To agglomerate the patients in the hierarchical
cluster, we used the Ward method. Function heatmap.2 from R package
gplots was used for the graphical display of the dendrogram
(11).
Predicted miRNA target analysis
Potential miRNA targets were predicted and analyzed
using DIANA TarBase (12)
(http://diana.pcbi.upenn.edu/). Predicted
target lists were analyzed using DIANA mirPath for association with
molecular pathways potentially altered by the expression of single
or multiple miRNAs (13). Predicted
target lists were also analyzed for association with Gene Ontology
(GO) (14) terms using L2L
microarray analysis tools (15).
Evolution analysis
Potential miRNA target sequences were obtained from
the National Center for Biotechnology Information website (NCBI,
USA). The sequences were aligned by the ClustalW program
(http://www.ebi.ac.uk/clustalw/)
(16). The evolutionary tree of
protein sequences was constructed by the neighbor-joining (NJ)
method with protein p-distances by MEGA4 (Molecular Evolutionary
Genetics Analysis) software (http://www.megasoftware.net/index.html) (17) and the reliability of the tree
topology was assessed by 1,000 bootstrap replications.
Results
miRNA expression profile in supraglottic
carcinoma and normal tissues
Using the miRNA microarray containing 1,145 human
miRNA probe sets, we first assessed the miRNA profiles in three
paired supraglottic carcinoma and normal tissues.
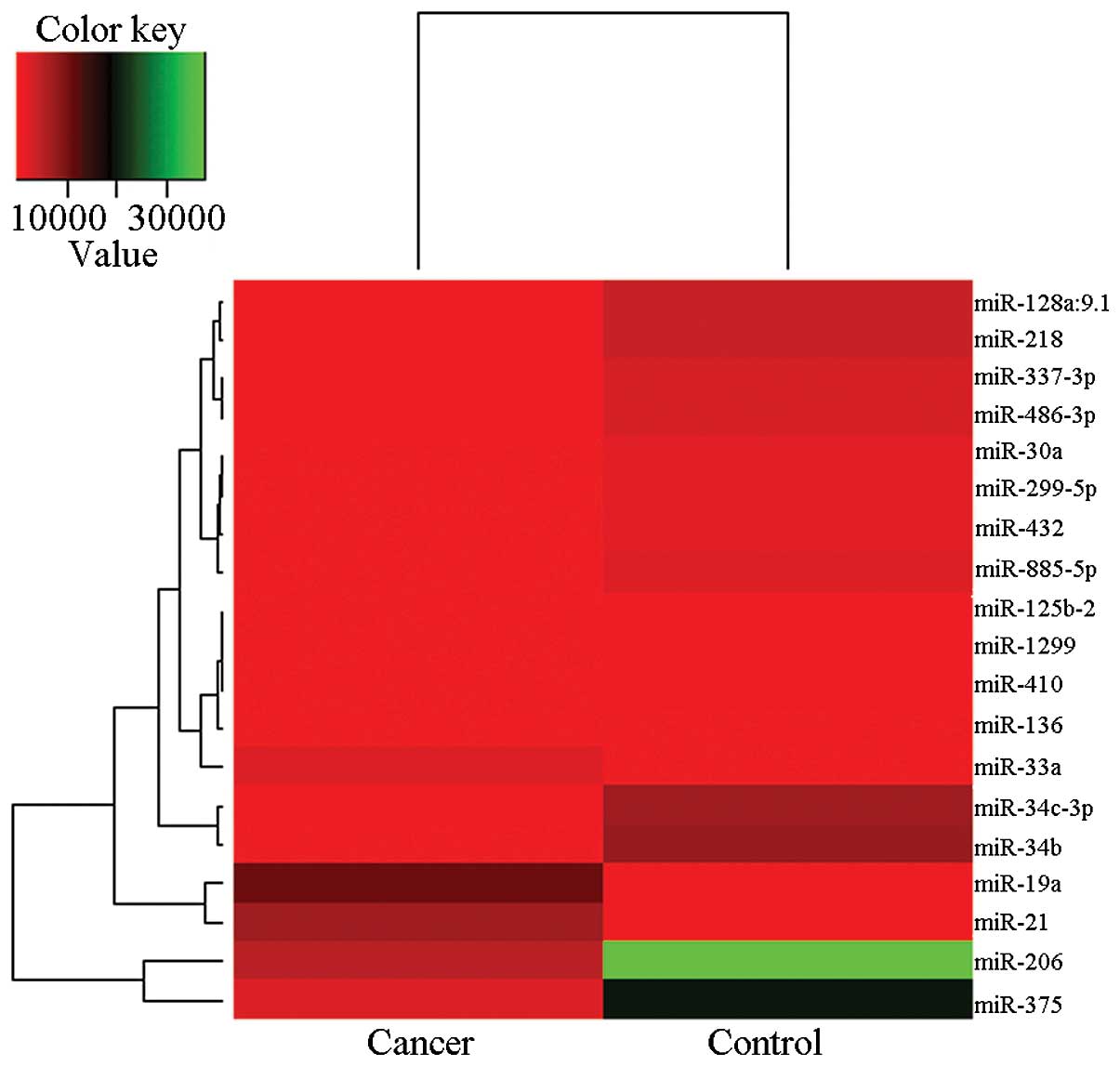
Unsupervised hierarchical clustering based on all
the miRNAs spotted on the chip revealed a marked, distinct
separation of the supraglottic carcinoma miRNA profiles compared
with those of normal tissues. Significance analysis of microarray
identified 85 significantly differentially expressed miRNAs
(P<0.01) between supraglottic carcinoma and normal tissues.
These human miRNAs further identified by significance analysis of
microarray method showed >5-fold difference between supraglottic
carcinoma and normal tissues. This method identified 19 miRNAs to
be significantly altered in their expression between supraglottic
carcinoma and non-diseased tissues, with 3 being upregulated
(miR-21, miR-19a, miR-33a) and 16 downregulated in supraglottic
carcinoma (Table I). Unsupervised
clustering analysis using these 19 miRNAs discriminated cancer from
non-cancer tissues (Fig. 1), thus
indicating that these miRNAs may be a valid diagnostic for
supraglottic carcinoma, although due to the small number of samples
analyzed, this requires further analysis.
 | Table IDifferentially expressed miRNAs in
supraglottic carcinoma tissues compared with normal tissues. |
Table I
Differentially expressed miRNAs in
supraglottic carcinoma tissues compared with normal tissues.
| miRNA | Chromosome | Expression fold
(cancer/normal tissue) | P-value |
|---|
| miR-34b | 11 | 0.061481 | 7.36E-38 |
| miR-885-5p | 3 | 0.107502 | 7.36E-38 |
| miR-218 | 4,5 | 0.133269 | 7.36E-38 |
| miR-34c-3p | 11 | 0.133316 | 7.36E-38 |
| miR-299-5p | 14 | 0.134148 | 7.36E-38 |
| miR-375 | 2 | 0.139267 | 7.36E-38 |
| miR-30a | 6 | 0.140864 | 3.62E-33 |
| miR-206 | 6 | 0.148176 | 2.83E-10 |
| miR-410 | 14 | 0.16099 | 2.15E-27 |
| miR-1299 | 9 | 0.16614 | 6.60E-25 |
| miR-128a:9.1 | - | 0.168493 | 7.36E-38 |
| miR-432 | 14 | 0.170929 | 6.93E-37 |
| miR-136 | 14 | 0.173921 | 6.49E-13 |
| miR-486-3p | 8 | 0.182158 | 1.16E-36 |
| miR-337-3p | 14 | 0.186334 | 7.86E-35 |
| miR-125b-2 | 21 | 0.187213 | 3.62E-20 |
| miR-21 | 17 | 8.453834 | 0.008087 |
| miR-19a | 13 | 13.15016 | 0.0054576 |
| miR-33a | 22 | 13.33146 | 0.0057371 |
Bioinformatics analysis of supraglottic
carcinoma miRNA target genes
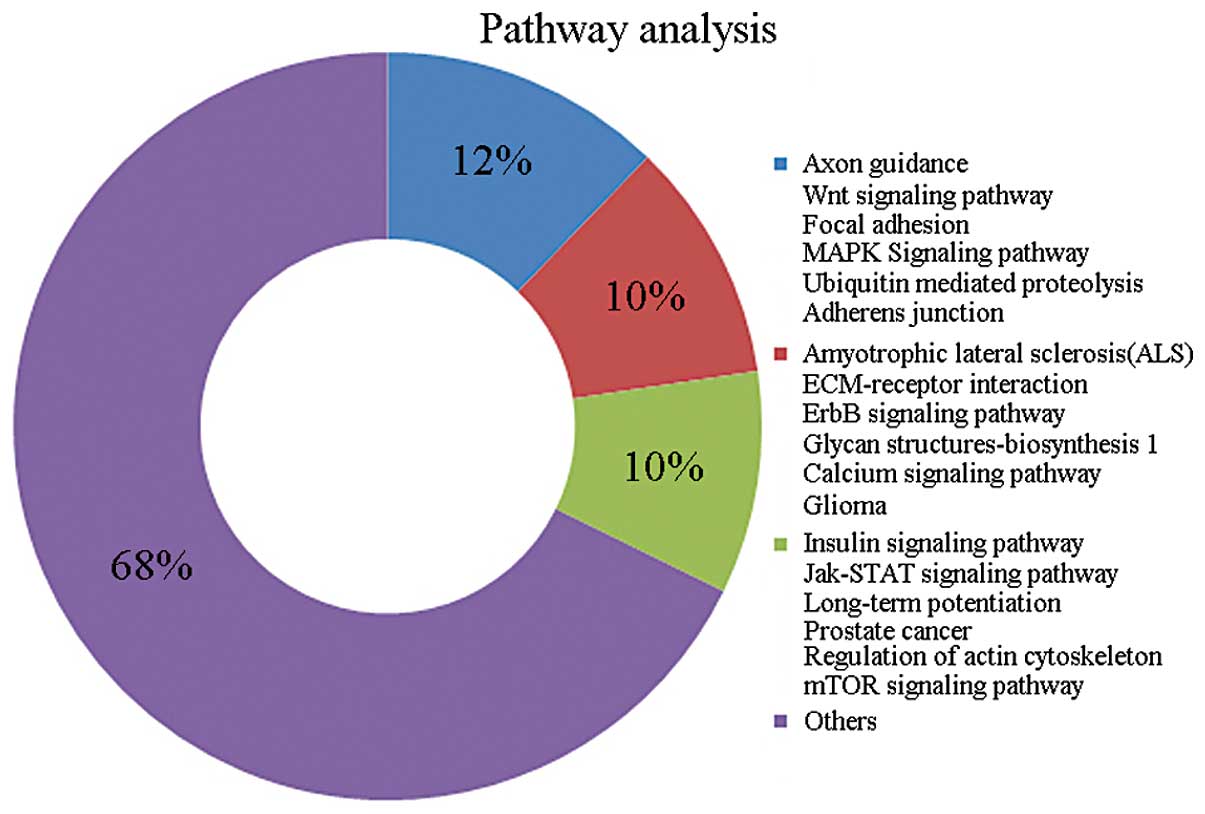
To investigate the involvement of miRNAs in
supraglottic carcinoma, we performed systemic bioinformatics
analysis to identify potential gene targets, including pathway
analysis (Fig. 2). Several pathways
were enriched in the target genes. Notably, multiple genes were
related to signaling pathways. The list of regulated gene targets
was also used to perform a GO (14)
analysis to evaluate the specific functional categories of genes
from broad GO categories (Table
II). The results indicated that seven biological processes, six
molecular functions and two cellular components were statistically
enriched with these miRNA targets. Furthermore, we found that 25
potential target genes related to 12 miRNAs which were strongly
supported by references to avoid the false negative predictions
(Table III). As a result, 3
target genes were involved in mRNA cleavage and 13 target genes
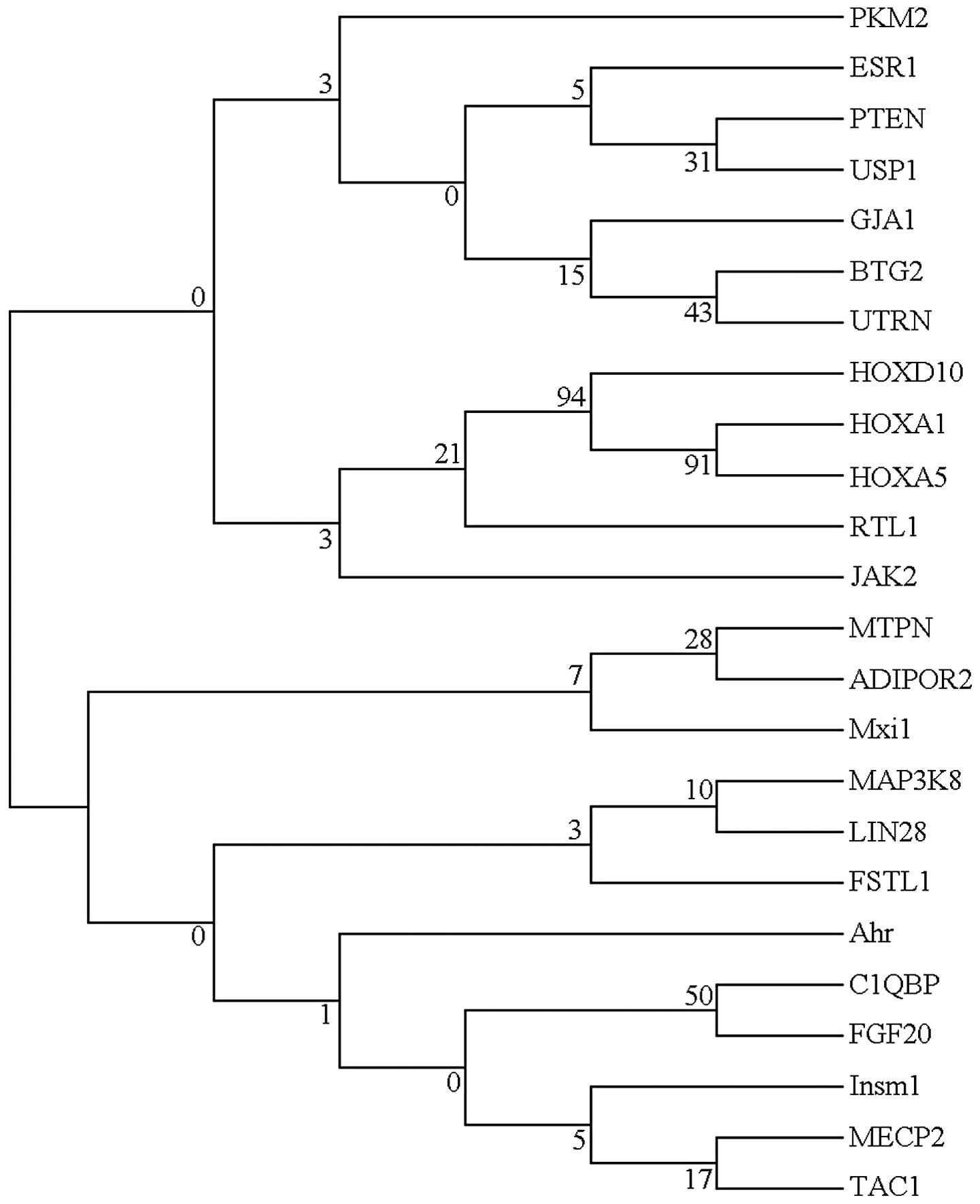
were involved in mRNA repression. The evolutionary tree of miRNA
target genes was further analyzed to illustrate the function of
these genes (Fig. 3). These genes
showed a high level of sequence divergence. It is reasonable to
infer that these genes are members of a multigene family, since
they shared low sequence similarity. Generally, a node with >70%
bootstrap supporting value is a stable node. The NJ-tree of miRNA
target genes exhibited low bootstrap supporting values on each deep
branch node and, therefore, few strong conclusions may be drawn
from the evolutionary tree.
| Table IIGene Ontology analysis in
supraglottic carcinoma. |
Table II
Gene Ontology analysis in
supraglottic carcinoma.
| GO type | GO term | Description | P-value |
|---|
| Process | GO: 0006725 | Cellular aromatic
compound metabolic process | 2.79E-4 |
| Process | GO: 0050776 | Regulation of
immune response | 4.64E-4 |
| Process | GO: 0001919 | Regulation of
receptor recycling | 4.94E-4 |
| Process | GO: 0003015 | Heart process | 5E-4 |
| Process | GO: 0060047 | Heart
contraction | 5E-4 |
| Process | GO: 0019882 | Antigen processing
and presentation | 5.76E-4 |
| Process | GO: 0006066 | Alcohol metabolic
process | 8.35E-4 |
| Process | GO: 0008152 | Metabolic
process | 9.43E-4 |
| Process | GO: 0031943 | Regulation of
glucocorticoid metabolic process | 9.83E-4 |
| Function | GO: 0005488 | Binding | 5.05E-7 |
| Function | GO: 0003824 | Catalytic
activity | 1.08E-5 |
| Function | GO: 0004423 |
Iduronate-2-sulfatase activity | 5.37E-4 |
| Function | GO: 0046872 | Metal ion
binding | 5.9E-4 |
| Function | GO: 0000062 | Acyl-CoA
binding | 9.21E-4 |
| Function | GO: 0043169 | Cation binding | 9.35E-4 |
| Component | GO: 0044424 | Intracellular
part | 3.63E-6 |
| Component | GO: 0044444 | Cytoplasmic
part | 8.87E-5 |
| Table IIIComparison of pathways associated
with miRNA and mRNA signatures. |
Table III
Comparison of pathways associated
with miRNA and mRNA signatures.
| miRNA | Gene | Type | Author/(Ref.) |
|---|
| miR-10a | HOXA1 | mRNA
repression | Garzon et al
(29) |
| miR-10b | HOXD10 | mRNA
repression | Ma et al
(33) |
| miR-125b | LIN28 | mRNA cleavage | Wu and Belasco
(39) |
| miR-133a | PKM2 | Unknown | Wong et al
(38) |
| miR-133b | PKM2 | Unknown | Wong et al
(38) |
| miR-136 | RTL1 | mRNA cleavage | Davis et al
(28) |
| miR-19a | MECP2 | Unknown | Lewis et al
(32) |
| miR-19a | PTEN | mRNA
repression | Lewis et al
(32) |
| miR-19a | HOXA5 | Unknown | Lewis et al
(32) |
| miR-206 | FSTL1 | mRNA
repression | Rosenberg et
al (36) |
| miR-206 | UTRN | mRNA
repression | Rosenberg et
al (36) |
| miR-206 | GJA1 | mRNA
repression | Anderson et
al (27) |
| miR-206 | ESR1 | mRNA cleavage | Adams et al
(26) |
| miR-206 | TAC1 | mRNA
repression | Greco and Rameshwar
(30) |
| miR-370 | MAP3K8 | Unknown | Meng et al
(34) |
| miR-375 | MTPN | mRNA
repression | Poy et al
(35) |
| miR-375 | Ahr | Unknown | Krek et al
(31) |
| miR-375 | C1QBP | mRNA
repression | Krek et al
(31) |
| miR-375 | Insm1 | Unknown | Krek et al
(31) |
| miR-375 | ADIPOR2 | mRNA
repression | Krek et al
(31) |
| miR-375 | JAK2 | mRNA
repression | Krek et al
(31) |
| miR-375 | Mxi1 | Unknown | Krek et al
(31) |
| miR-375 | USP1 | mRNA
repression | Krek et al
(31) |
| miR-433 | FGF20 | Unknown | Wang et al
(37) |
| miR-21 | BTG2 | mRNA
repression | Liu et al
(22) |
Discussion
The present study demonstrated that multiple
microRNAs (miRNAs) are upregulated or downregulated in supraglottic
carcinoma, including an extensively validated subset that may be
potential clinical biomarkers of disease. Microarray profiling of
more than 1,000 miRNAs identified 85 miRNAs that were significantly
differentially expressed in tumor tissues compared with normal
tissues. Of the 85 human miRNAs, three upregulated miRNAs (miR-21,
miR-19a, miR-33a) and two downregulated miRNAs (miR-206, miR-375)
were highlighted. Twenty-five target genes were identified to help
characterize the diverse functions of miRNAs. Furthermore, biology
process and evolution were explored to find the function of
miRNAs.
miRNA expression profiles can be used to classify
human cancer (14,18). Distinct signatures for several
epithelial cancers have been reported, such as breast, lung,
pancreatic and gastric cancer (9,19–21).
Our study explored miRNA expression in supraglottic carcinoma.
Consistent with previous reports of miRNA expression in head and
neck cancer (22–25), some miRNAs were significantly
deregulated, although changes in individual miRNAs did not match
completely. For example, miR-21 was significantly upregulated in
all head and neck cancer analysis (22–25).
Tran et al found miR-21 and miR-205 were highly expressed in
head and neck cancer cell lines (25). Chang et al found miR-21,
let-7, 18, 29c, 142-3p, 155 and 16b were significantly
overexpressed in primary head and neck squamous cell carcinoma
(24). Avissar et al found
miR-221 and miR-375 were significantly altered and upregulated in
head and neck squamous cell carcinoma (23). Liu et al identified 13 miRNAs
that were differentially expressed, 7 miRNAs were downregulated and
6 miRNAs (let-7a-1, miR-203, miR-205, miR-21, miR-98 and miR-16-1)
were upregulated in laryngeal carcinoma tissues based on 210 human
miRNA probe sets (22). This
indicated that the molecular biology of head and neck squamous cell
carcinoma is complex. It is important to analyze the miRNA
mechanism on a larger scale.
miRNAs modulate gene expression by targeting mRNAs
for translational suppression or mRNA cleavage and it is well known
that miRNAs regulate a variety of cellular activities through their
effect on the expression of multiple target genes (4). The identification of target genes
regulated by a specific miRNA has been proved difficult despite the
development of computational approaches to predict miRNA targets.
The ability to find target genes is further complicated by the fact
that target selectivity of miRNAs may depend on the cellular
microenvironment. Our studies identified 25 reliable target genes,
supported by previous studies (22,26–39).
The evolutionary tree of these target genes showed a low sequence
similarity and it indicated that miRNAs may play different roles in
supraglottic carcinoma. Also, these results provided a possible way
to address biological meaning from the global pattern of cellular
functions and pathways that are affected by miRNAs in supraglottic
carcinoma. Gene Ontology (GO) (14)
was developed into three structured controlled vocabularies
(ontologies) that describe gene products in components and
molecular functions in a species-independent manner (40). GO enrichment analysis was used to
reduce the number of targets of a large group of co-expressed
miRNAs and to find biological functions potentially affected by
multiple miRNAs in our research. We performed a statistical
enrichment analysis of GO categories to find categories that are
enriched with targets of co-expressed miRNAs. It allowed us to
reduce a very large raw list of predicted target genes to a smaller
set of target genes from significantly enriched GO categories. We
assumed that filtering GO categories on the total number of hits by
miRNAs targeting the same category would reduce the number of false
positive target predictions and at the same time would allow
narrowing down of the large target lists and determining those
biological functions.
In conclusion, our global analysis of miRNA array in
human supraglottic carcinoma tissues demonstrated that co-expressed
miRNAs may collectively provide systemic compensatory response to
the abnormal functional and phenotypic changes in supraglottic
carcinoma by targeting a broad range of functional categories and
abnormally activated pathways known to be affected in supraglottic
carcinoma. Such system biology based approach may provide new
avenues for biological interpretation of miRNA profiling data as
well as generation of experimentally testable hypotheses regarding
collective regulatory functions of miRNAs in supraglottic carcinoma
biology.
Acknowledgements
This study was supported by grants from the
Heilongjiang Postdoctoral Foundation (LBHZ12194), and the Research
Foundation of Heilongjiang Provincial Department of Education.
References
|
1
|
Rudolph E, Dyckhoff G, Becher H, Dietz A
and Ramroth H: Effects of tumour stage, comorbidity and therapy on
survival of laryngeal cancer patients: a systematic review and a
meta-analysis. Eur Arch Otorhinolaryngol. 268:165–179. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cosetti M, Yu GP and Schantz SP: Five-year
survival rates and time trends of laryngeal cancer in the US
population. Arch Otolaryngol Head Neck Surg. 134:370–379.
2008.PubMed/NCBI
|
|
3
|
Petrakos I, Kontzoglou K, Nikolopoulos TP,
Papadopoulos O and Kostakis A: Glottic and supraglottic laryngeal
cancer: epidemiology, treatment patterns and survival in 164
patients. J BUON. 17:700–705. 2012.PubMed/NCBI
|
|
4
|
Bartel DP: MicroRNAs: genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar
|
|
5
|
Ferretti E, De Smaele E, Po A, et al:
MicroRNA profiling in human medulloblastoma. Int J Cancer.
124:568–577. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Khoshnaw SM, Green AR, Powe DG and Ellis
IO: MicroRNA involvement in the pathogenesis and management of
breast cancer. J Clin Pathol. 62:422–428. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Raponi M, Dossey L, Jatkoe T, et al:
MicroRNA classifiers for predicting prognosis of squamous cell lung
cancer. Cancer Res. 69:5776–5783. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Akao Y, Nakagawa Y and Naoe T: MicroRNAs
143 and 145 are possible common onco-microRNAs in human cancers.
Oncol Rep. 16:845–850. 2006.PubMed/NCBI
|
|
9
|
Iorio MV, Ferracin M, Liu CG, et al:
MicroRNA gene expression deregulation in human breast cancer.
Cancer Res. 65:7065–7070. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Michael MZ, O’ Connor SM, van Holst
Pellekaan NG, Young GP and James RJ: Reduced accumulation of
specific microRNAs in colorectal neoplasia. Mol Cancer Res.
1:882–891. 2003.PubMed/NCBI
|
|
11
|
Gentleman RC, Carey VJ, Bates DM, et al:
Bioconductor: open software development for computational biology
and bioinformatics. Genome Biol. 5:R802004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Papadopoulos GL, Reczko M, Simossis VA,
Sethupathy P and Hatzigeorgiou AG: The database of experimentally
supported targets: a functional update of TarBase. Nucleic Acids
Res. 37:D155–D158. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Papadopoulos GL, Alexiou P, Maragkakis M,
Reczko M and Hatzigeorgiou AG: DIANA-mirPath: integrating human and
mouse microRNAs in pathways. Bioinformatics. 25:1991–1993. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lu J, Getz G, Miska EA, et al: MicroRNA
expression profiles classify human cancers. Nature. 435:834–838.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Newman JC and Weiner AM: L2L: a simple
tool for discovering the hidden significance in microarray
expression data. Genome Biol. 6:R812005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Thompson JD, Higgins DG and Gibson TJ:
CLUSTAL W: improving the sensitivity of progressive multiple
sequence alignment through sequence weighting, position-specific
gap penalties and weight matrix choice. Nucleic Acids Res.
22:4673–4680. 1994. View Article : Google Scholar
|
|
17
|
Kumar S, Nei M, Dudley J and Tamura K:
MEGA: a biologist-centric software for evolutionary analysis of DNA
and protein sequences. Brief Bioinform. 9:299–306. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Volinia S, Calin GA, Liu CG, et al: A
microRNA expression signature of human solid tumors defines cancer
gene targets. Proc Natl Acad Sci USA. 103:2257–2261. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Petrocca F, Visone R, Onelli MR, et al:
E2F1-regulated microRNAs impair TGFβ-dependent cell-cycle arrest
and apoptosis in gastric cancer. Cancer Cell. 13:272–286. 2008.
|
|
20
|
Bloomston M, Frankel WL, Petrocca F, et
al: MicroRNA expression patterns to differentiate pancreatic
adenocarcinoma from normal pancreas and chronic pancreatitis. JAMA.
297:1901–1908. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yanaihara N, Caplen N, Bowman E, et al:
Unique microRNA molecular profiles in lung cancer diagnosis and
prognosis. Cancer Cell. 9:189–198. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu M, Wu H, Liu T, et al: Regulation of
the cell cycle gene, BTG2, by miR-21 in human laryngeal
carcinoma. Cell Res. 19:828–837. 2009. View Article : Google Scholar
|
|
23
|
Avissar M, Christensen BC, Kelsey KT and
Marsit CJ: MicroRNA expression ratio is predictive of head and neck
squamous cell carcinoma. Clin Cancer Res. 15:2850–2855. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chang SS, Jiang WW, Smith I, et al:
MicroRNA alterations in head and neck squamous cell carcinoma. Int
J Cancer. 123:2791–2797. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tran N, McLean T, Zhang X, et al: MicroRNA
expression profiles in head and neck cancer cell lines. Biochem
Biophys Res Commun. 358:12–17. 2007. View Article : Google Scholar
|
|
26
|
Adams BD, Furneaux H and White BA: The
micro-ribonucleic acid (miRNA) miR-206 targets the human estrogen
receptor-α (ERα) and represses ERα messenger RNA and protein
expression in breast cancer cell lines. Mol Endocrinol.
21:1132–1147. 2007.PubMed/NCBI
|
|
27
|
Anderson C, Catoe H and Werner R: MIR-206
regulates connexin43 expression during skeletal muscle development.
Nucleic Acids Res. 34:5863–5871. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Davis E, Caiment F, Tordoir X, et al:
RNAi-mediated allelic trans-interaction at the imprinted
Rtl1/Peg11 locus. Curr Biol. 15:743–749. 2005.PubMed/NCBI
|
|
29
|
Garzon R, Pichiorri F, Palumbo T, et al:
MicroRNA fingerprints during human megakaryocytopoiesis. Proc Natl
Acad Sci USA. 103:5078–5083. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Greco SJ and Rameshwar P: MicroRNAs
regulate synthesis of the neurotransmitter substance P in human
mesenchymal stem cell-derived neuronal cells. Proc Natl Acad Sci
USA. 104:15484–15489. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Krek A, Grün D, Poy MN, et al:
Combinatorial microRNA target predictions. Nat Genet. 37:495–500.
2005. View
Article : Google Scholar
|
|
32
|
Lewis BP, Shih IH, Jones-Rhoades MW,
Bartel DP and Burge CB: Prediction of mammalian microRNA targets.
Cell. 115:787–798. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ma L, Teruya-Feldstein J and Weinberg RA:
Tumour invasion and metastasis initiated by microRNA-10b in breast
cancer. Nature. 449:682–688. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Meng F, Wehbe-Janek H, Henson R, Smith H
and Patel T: Epigenetic regulation of microRNA-370 by interleukin-6
in malignant human cholangiocytes. Oncogene. 27:378–386. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Poy MN, Eliasson L, Krutzfeldt J, et al: A
pancreatic islet-specific microRNA regulates insulin secretion.
Nature. 432:226–230. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Rosenberg MI, Georges SA, Asawachaicharn
A, Analau E and Tapscott SJ: MyoD inhibits Fstl1 and Utrn
expression by inducing transcription of miR-206. J Cell Biol.
175:77–85. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang G, van der Walt JM, Mayhew G, et al:
Variation in the miRNA-433 binding site of FGF20 confers risk for
Parkinson disease by overexpression of α-synuclein. Am J Hum Genet.
82:283–289. 2008.PubMed/NCBI
|
|
38
|
Wong TS, Liu XB, Chung-Wai Ho A, et al:
Identification of pyruvate kinase type M2 as potential oncoprotein
in squamous cell carcinoma of tongue through microRNA profiling.
Int J Cancer. 123:251–257. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wu L and Belasco JG: Micro-RNA regulation
of the mammalian lin-28 gene during neuronal differentiation
of embryonal carcinoma cells. Mol Cell Biol. 25:9198–9208.
2005.PubMed/NCBI
|
|
40
|
Ashburner M, Ball CA, Blake JA, et al:
Gene ontology: tool for the unification of biology. The Gene
Ontology Consortium. Nat Genet. 25:25–29. 2000. View Article : Google Scholar : PubMed/NCBI
|