Introduction
Gastric cancer (GC) remains one of the leading
causes of cancer-related mortality worldwide, although therapeutic
outcomes for early GC have recently improved (1). Generally, GC can be classified into
two histological types: differentiated type and undifferentiated
type, according to the Japanese classification of gastric carcinoma
(2). Among the undifferentiated
type GCs, lesions in scirrhous carcinomas of the stomach display
rigid thickening of the gastric wall, leading to a leather
bottle-like appearance. Histopathologically, scirrhous cancer cells
do not form glands but cause diffuse infiltration. Due to these
pathological features, early clinical diagnosis of scirrhous GC
remains difficult (3) and it has a
poorer prognosis than other types of GC. This reflects the rapid
proliferation of these cancer cells, accompanied by progressive
invasion and a high frequency of metastasis to the peritoneum
(4), all of which distinguish it
from differentiated types of GC (5). In fact, there is currently no reliable
therapeutic target for this type of GC, thus the need for
elucidation remains.
Comprehensive gene expression analysis is a powerful
tool in the identification of potential therapeutic targets for GC
and to better understand the development of GC at the molecular
level. We previously performed several large-scale gene expression
studies using array-based hybridization (6) and serial analysis of gene expression
(SAGE) (7,8) enabling identification of several genes
including regenerating islet-derived family, member 4 (REG4, which
encodes REGIV) (9); olfactomedin 4
(OLFM4) (10); palate, lung and
nasal epithelium carcinoma-associated protein (PLUNC) (11); and GJB6 (encoding connexin 30)
(12). Our recent study of
Escherichia coli (E. coli) ampicillin secretion trap (CAST)
analysis on two scirrhous type GC tissues identified several
candidate cancer-specific genes. Among them, transmembrane 9
superfamily member 3 (TM9SF3) expression was associated with tumor
progression and was involved in the cancer cell invasion process.
Moreover, TM9SF3-positive status in GC significantly correlated
with depth of invasion and tumor stage, demonstrated by
immunohistochemical analysis (unpublished data).
We analyzed and verified several candidate genes
from previous CAST libraries. Among these genes, in the present
study, we focused on the zinc finger DHHC domain-containing protein
14 (ZDHHC14) gene expression, due to its frequent overexpression in
GC tissue in comparison to expression in corresponding
non-neoplastic mucosa, and we analyzed expression levels by
qRT-PCR. ZDHHC14 has been reported to be one of the genes
overexpressed in acute biphenotypic leukemia through chromosomal
translocation (13). However, to
our knowledge, the detailed function and expression profiles of the
ZDHHC14 gene in the majority of human cancers has not yet been
elucidated.
This is the first study of ZDHHC14 expression in
human GC; we examined the relationship between ZDHHC14 expression
and clinicopathologic characteristics, including tumor stage, TNM
grading and histological type. Here, we also clarified the
biological significance of ZDHHC14 mRNA expression, via qRT-PCR
analysis of surgically resected GC tissues. Furthermore, using an
siRNA knockdown system and forced expression vector, the biological
role of ZDHHC14 gene in cancer cell adhesion, migration and
invasion was examined in GC cell lines.
Materials and methods
Tissue samples
In total, 164 primary tumor samples were collected
from patients diagnosed with GC. For quantitative reverse
transcription-PCR (RT-PCR) analysis, 41 GC samples and
corresponding non-neoplastic mucosa samples were obtained during
surgery at the Hiroshima University Hospital, Japan. Informed
consent was obtained from all study subjects. The remaining 123 GC
samples used for mRNA analysis were obtained from patients who
underwent surgery at the Gastroenterological Center, Yokohama City
University Medical Center, and at the Department of Surgery,
Yokohama City University, Japan, from January 2002 to July 2007.
Informed consent was obtained from each participant, and ethics
approval was granted by the committee of Yokohama City University
Medical Center. Noncancerous samples were purchased from Clontech
(Palo Alto, CA, USA). The 164 cases were histologically classified
as differentiated type (papillary adenocarcinoma or tubular
adenocarcinoma) and undifferentiated type (poorly differentiated
adenocarcinoma, signet ring cell carcinoma or mucinous
adenocarcinoma), according to the Japanese Classification of
Gastric Carcinomas (2). Tumor
staging was assessed according to the International Union Against
Cancer TNM classification of malignant tumors.
Quantitative RT-PCR and western blot
analysis
Quantitative RT-PCR was performed with an ABI Prism
7900 Sequence Detection System (Applied Biosystems, Foster City,
CA, USA) as previously described (14). We calculated the ratio of target
gene mRNA expression levels between GC tissue (T) and corresponding
non-neoplastic mucosa (N). T/N ratios >2-fold were considered to
represent overexpression. β-actin (ACTB gene) was used as
housekeeping internal control. Western blot analysis was performed
as previously described (15).
Rabbit polyclonal anti-ZDHHC14 (83260; Abcam) antibody was used for
western blotting.
RNA interference (RNAi)
For knockdown of endogenous ZDHHC14, RNAi was
performed. siRNA oligonucleotides for ZDHHC14 and a negative
control were purchased from Invitrogen (Carlsbad, CA, USA). Three
independent oligonucleotides were used for ZDHC14 siRNA as follows:
ZDHHC14 siRNA1 sequence, 5′-CAAGCCTGATCGACAGAAGAGG GTA-3′; ZDHHC14
siRNA2 sequence, 5′-GACCAGTGCATT CAGAGCACCAAAT-3′ and ZDHHC14
siRNA3 sequence, 5′-AAGATGAGCACATGGGCCACCAGTT-3′. Transfection was
performed using Lipofectamine RNAiMAX Reagent (Invitrogen)
according to the manufacturer’s protocol. Briefly, 100 pmol siRNA
and 12 μl Lipofectamine RNAiMAX were mixed in 1 ml RPMI medium (10
nmol/l final siRNA concentration). After a 20-min incubation, the
mixture was added to the cells and these were plated on dishes for
each assay. In all experiments, the cells analyzed were incubated
for 48 h after transfection.
Cell lines and in vitro invasion
assays
Three cell lines derived from human GC were used.
The TMK-1 cell line was established in our laboratory from a poorly
differentiated adenocarcinoma (16). HSC-44PE and 44As3 cell lines were
established by one of the authors (K.Y., National Cancer Center
Hospital East) (3,17). All cell lines were maintained in
RPMI-1640 (Nissui Pharmaceutical Co, Ltd, Tokyo, Japan) containing
10% fetal bovine serum (Biowhittaker, Walkersville, MD, USA) in a
humidified atmosphere of 5% CO2 and 95% air at 37°C. The
HSC-44PE cells were seeded at a density of 2,000 cells/well in
96-well plates. Cell growth was monitored at day 0, 1, 2 and 4 for
the MTT assay, as mentioned elsewhere (18). Modified Boyden chamber assays were
carried out to examine invasiveness. Cells were suspended at
200,000 cells/well in RPMI-1640 medium plus 1% serum then plated
into the upper chamber of a Matrigel-coated Transwell insert (8 μm
pore diameter; Chemicon, Temecula, CA, USA). Medium containing 10%
serum was added in the bottom chamber using 24-well plate format.
On day 1 and 2, non-invading cells in the upper chamber were
removed by clean cotton swab and the cells attached on the lower
surface of the insert were stained with Cell Stain (Chemicon). The
invading cells were counted by light microscope on day 1 and 2.
Cell migration and adhesion assay
To evaluate cell motility, a wound-healing assay was
performed. Negative control and siRNA treated or overexpressing
cells were seeded after cell number adjustment performed via
automated cell counter (TC10; Bio-Rad). Cells grown to
subconfluence were scraped with the sharp edge of the pipette tip,
creating a cell-free area. Cells migrating into the scraped area
were observed and images were captured every 6 h (19) after mounting cells on fibronectin-
and collagen-coated glass slides. The cell adhesion assay was
performed by PBS wash at timepoints of 5, 15 and 30 min, and 1, 2
and 3 h post-seeding, in order to examine the remaining cells
attached to the glass surface in both control cells and RNAi
treated or overexpressed cells. Images were captured using an
inverted microscope (Eclipse TE300; Nikon, Japan).
Forced-expression vector construction and
generation of stable cell line
For constitutive expression of ZDHHC14 gene, cDNA
was PCR-amplified and subcloned into pcDNATM3.1(+)
vector (V790-20; Invitrogen) in-frame with EcoRI and
XhoI restriction enzymes for directional cloning. PCR
primers for cDNA amplification were also designed to include
EcoRI and XhoI sequence. Both transient and stable
transfections were carried out with the FuGENE6 Transfection
Reagent (Roche Diagnostics). The pcDNATM3.1(+) vector
contains the neomycin resistance gene for selection of stable cell
lines. Complete selection of vector construct containing ZDHHC14 or
empty vector was performed using G418/neomycin (Geneticin).
Statistical analysis
Correlations between clinicopathologic parameters
and ZDHHC14 expression were analyzed by Chi-square test. A P-value
of <0.05 was considered to indicate a statistically significant
difference. Statistical analyses were performed using JMP software
(version 9.0.2; SAS Institute, Carey, NC, USA).
Results
Evaluation of ZDHHC14 mRNA level in GC
tissue and corresponding non-neoplastic mucosa
Our previous CAST analysis showed that ZDHHC14 was
expressed in the scirrhous type GC tissue CAST library but not in
the normal stomach tissue CAST library (unpublished data). To
confirm whether the ZDHHC14 gene is cancer-specific, quantitative
RT-PCR for ZDHHC14 was performed in 41 GC tissues and their
corresponding non-neoplastic gastric mucosa. A ratio of target gene
mRNA expression levels between GC tissue (T) and corresponding
non-neoplastic mucosa (N) (T/N ratios) of >2-fold was considered
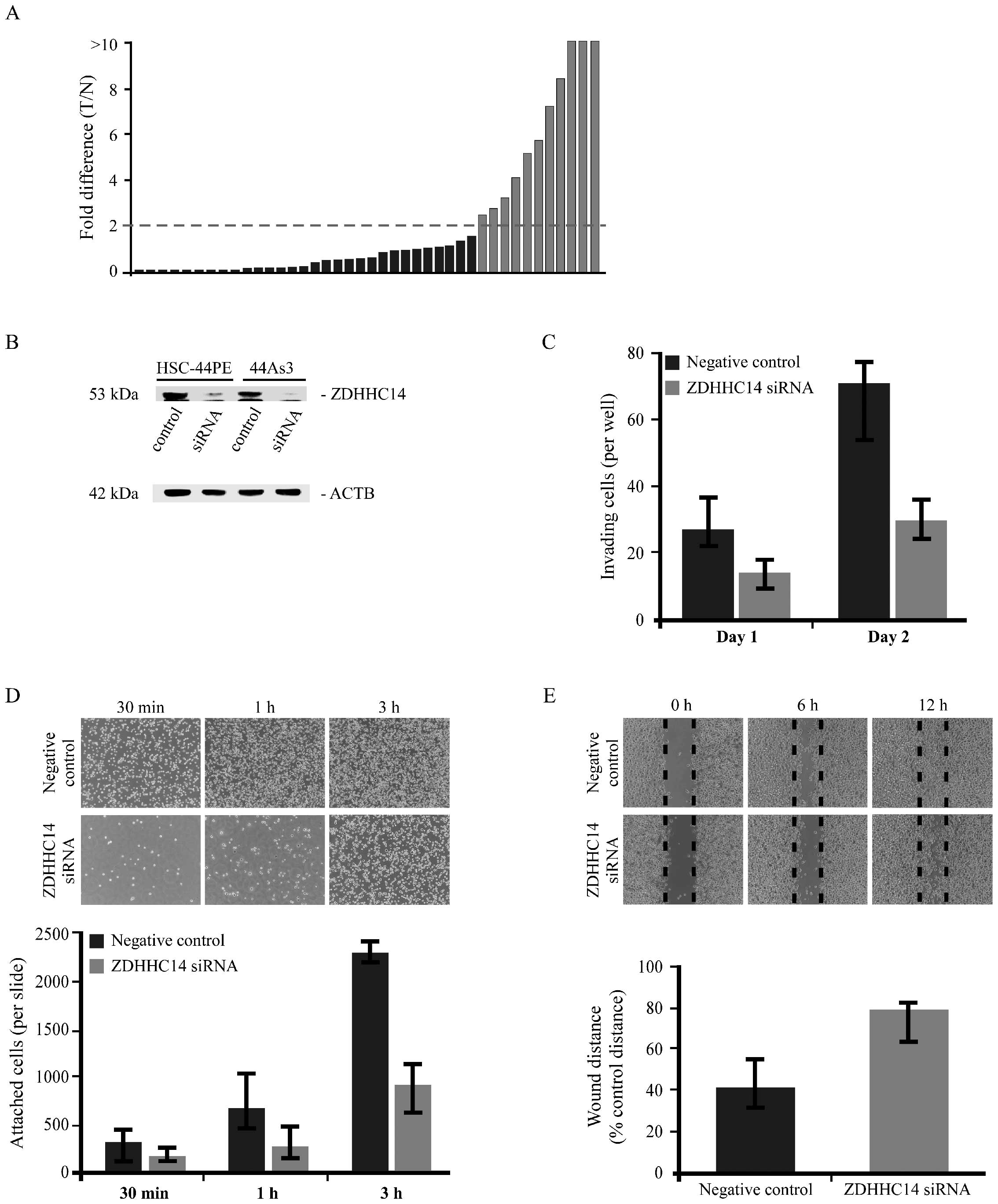
to represent overexpression. ZDHHC14 mRNA was overexpressed in
11/41 GC cases (26.8%) (Fig.
1A).
Relationship between high ZDHHC14
expression and clinicopathologic parameters
To analyze the relationship between
clinicopathologic parameters and ZDHHC14 expression in GC, we
performed qRT-PCR analysis of 123 surgically resected GC tissues
(Table I). ZDHHC14 expression was
overexpressed in 62/123 (50%) of GCs. ZDHHC14 highly-expressed GC
cases showed a significant correlation with the depth of invasion
(P=0.0275), and undifferentiated histological subtype (P=0.0169).
There was a tendency towards an association between later
pathological stage and high ZDHHC14 expression (data not shown).
Grouping scirrhous type GC cases and non-scirrhous type GC cases
separately revealed a strong correlation between scirrhous type GC
and high expression of ZDHHC14 (P=0.0247). There was no significant
association between ZDHHC14 expression and other parameters (age,
gender, nodal or stage).
 | Table IRelationship between high ZDHHC14
expression and clinicopathologic parameters in 123 GC cases,
evaluated by qRT-PCR. |
Table I
Relationship between high ZDHHC14
expression and clinicopathologic parameters in 123 GC cases,
evaluated by qRT-PCR.
| ZDHHC14 expression
(%) | |
|---|
|
| |
|---|
| High (n=62) | Low (n=61) | P-value |
|---|
| Age (years) |
| ≤65 (n=35) | 13 (37) | 22 | NS |
| >65 (n=88) | 49 (56) | 39 | |
| Gender |
| Male (n=87) | 42 (48) | 45 | NS |
| Female (n=36) | 20 (56) | 16 | |
| T grade |
| T1/T2 (n=39) | 14 (36) | 25 | 0.0275 |
| T3/T4 (n=84) | 48 (57) | 36 | |
| N grade |
| N0 (n=50) | 23 (46) | 27 | NS |
| N1/N2/N3 (n=73) | 39 (53) | 34 | |
| Stage |
| Stage I/II
(n=65) | 28 (43) | 37 | NS |
| Stage III/IV
(n=58) | 34 (59) | 24 | |
| Histology |
| Differentiated
(n=64) | 28 (44) | 36 | 0.0169 |
| Undifferentiated
(n=59) | 34 (58) | 25 | |
| Type |
| Scirrhous
(n=19) | 14 (74) | 5 | 0.0247 |
| Non-scirrhous
(n=104) | 48 (46) | 56 | |
Effect of ZDHHC14 downregulation on cell
adhesion, motility and invasive activity
To analyze the biological significance of ZDHHC14
protein in GC, we performed siRNA knockdown on this gene, using
scirrhous type GC cell lines. Gene silencing in HSC-44PE and 44As3
scirrhous type GC cell lines was confirmed by western blot analysis
(Fig. 1B). Very similar western
blot results were obtained from both cell lines; therefore Fig. 1C (measuring GC invasiveness)
displays results obtained using the 44As3 cell line.
To determine the possible role of ZDHHC14 in the
invasiveness of GC cells, a Transwell invasion assay was performed
in the 44As3 GC cell line. Invasion ability was significantly
abrogated in ZDHHC14 knockdown GC cells compared to negative
control siRNA-transfected GC cells (Fig. 1C). We also investigated the possible
proliferative effect of ZDHHC14. We performed an MTT assay 2 days
after negative control siRNA and ZDHHC14-siRNA transfection in
44As3 cells; there was no significant difference in proliferation
rate between control and knockdown cells (data not shown).
Furthermore, we observed that the knockdown cells were abnormally
loosely attached to the 96-well plate, leading to our closer
analysis of the effect of ZDHHC14 on cancer cell adhesion and
migration. We investigated cell adhesion via an assay using
fibronectin- and collagen-coated culture slides. In the adhesion
assay, ZDHHC14-knockdown cells showed significantly impaired
attachment to the culture glass compared to negative control cells
(Fig. 1D). This effect was more
significant on fibronectin-coated culture glass. Next, we performed
a cell migration assay, which showed significantly impaired
motility in ZDHHC14-knockdown cells compared to negative control
cells. Following cell scraping, cells were examined from 0 to 12 h
at intervals, until confluent (Fig.
1E).
Alteration of gene expression as a result
of ZDHHC14 knockdown
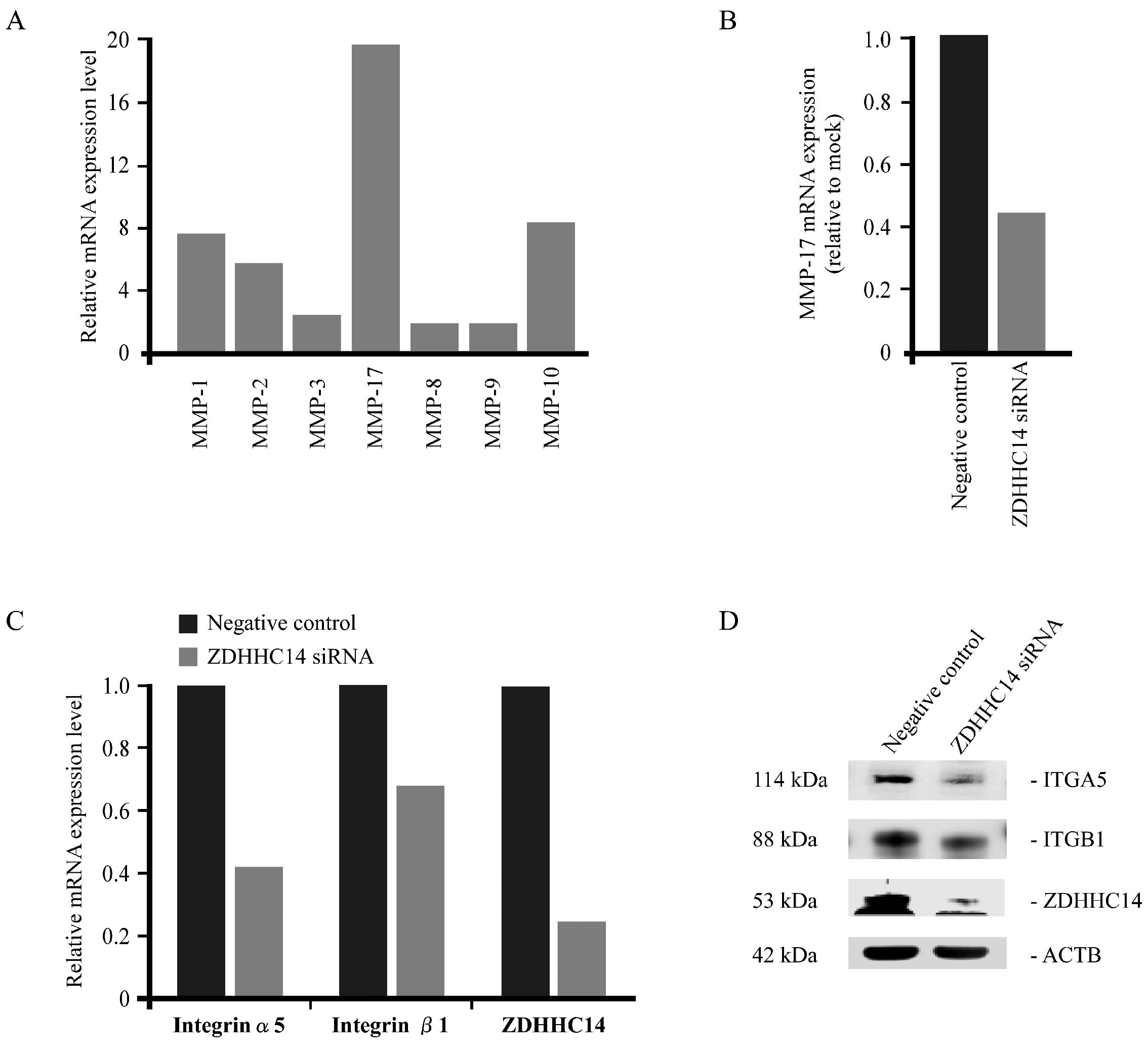
Based on these results, we analyzed the alteration
of gene expression in cancer invasion-associated genes and those
related with cell attachment after ZDHHC14-siRNA treatment. In
semi-quantitative RT-PCR analysis, MT4-MMP (MMP-17) showed the
highest expression among all other MMPs after normalization
(Fig. 2A). Significant
downregulation of MMP-17 expression was observed in ZDHHC14-siRNA
transfected 44As3 cells, compared with negative control (mock)
transfection (Fig. 2B). We also
analyzed the expression of several subunits of integrin α and β by
qRT-PCR (Fig. 2C) and by western
blot analysis (Fig. 2D); these
analyses both showed a downregulation of integrin α5 and integrin
β1 expression post-ZDHHC14-siRNA treatment.
Forced expression of ZDHHC14 promotes GC
cell migration and invasion
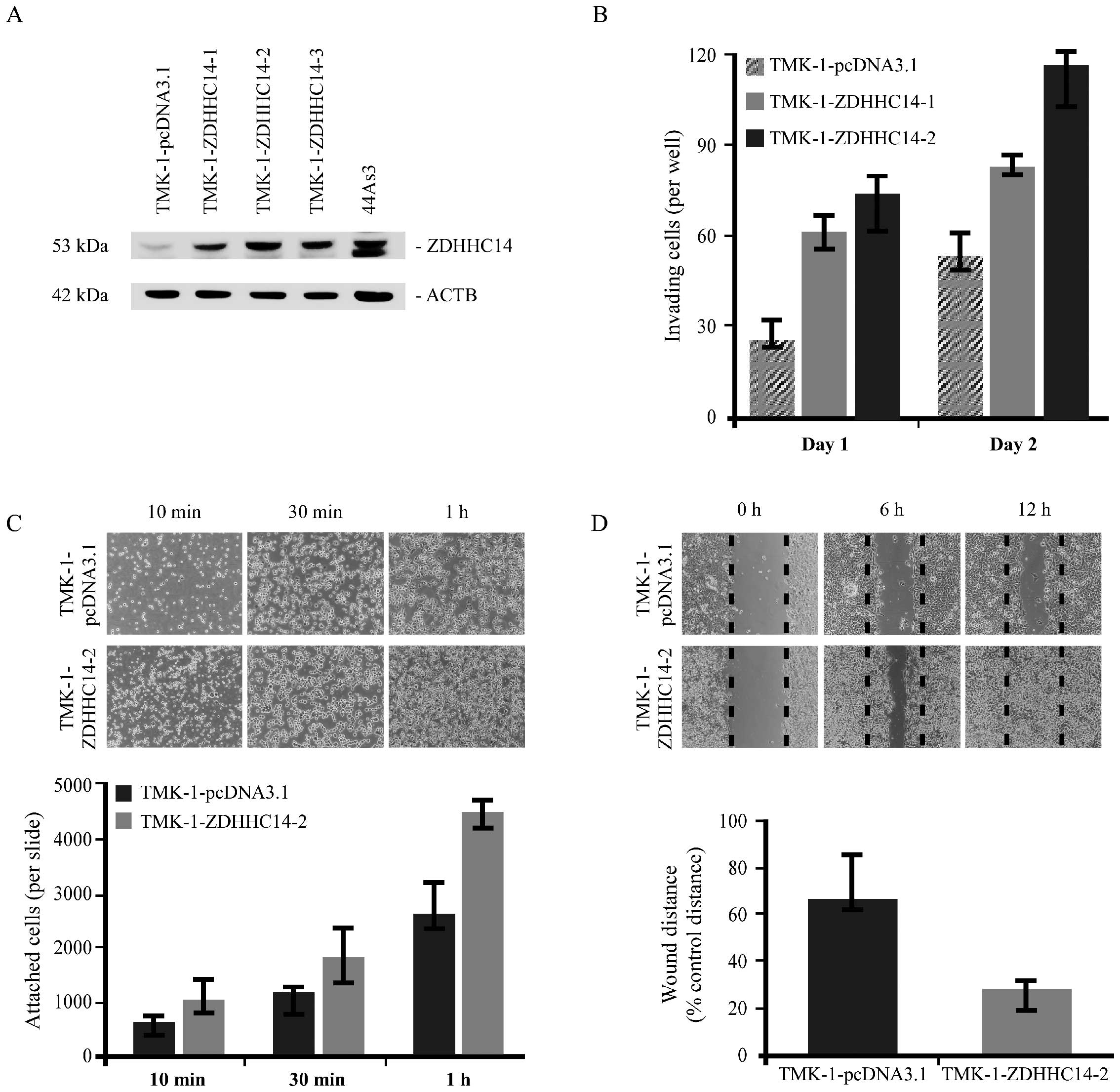
TMK-1 cells were selected for forced-expression
experiments from our screening of 8 different GC cell lines, due to
their low ZDHHC14 protein expression (data not shown). The TMK-1 GC
cell line was stably transfected with pcDNA3.1-ZDHHC14. For the
transfection experiments, clones were selected in G418 antibiotic
and examined for ZDHHC14 expression by western blotting (Fig. 3A). TMK-1 cells demonstrating stable
expression were designated as TMK-1-ZDHHC14-1, TMK-1-ZDHHC14-2 and
TMK-1-ZDHHC14-3. Endogenous and exogenous levels of ZDHHC14 protein
were also investigated by western blotting with anti-ZDHHC14
antibody to demonstrate the relative overexpression of ZDHHC14. As
shown in Fig. 3A, TMK-1-ZDHHC14-1,
TMK-1-ZDHHC14-2 and TMK-1-ZDHHC14-3 expressed ZDHHC14 protein at
significantly higher levels than TMK-1 cells transfected with empty
vector (TMK-1-pcDNA3.1). Next, Boyden chamber invasion assays were
performed. ZDHHC14-transfected TMK-1 cells were more invasive than
cells transfected with control vector (Fig. 3B). To determine the effect of
ZDHHC14 overexpression on cell adhesion and migration in
vitro, we performed cell adhesion and migration assays. In the
adhesion assay, ZDHHC14-overexpressed cells showed significant
attachment to culture glass slides compared to empty
vector-transfected cells (Fig. 3C).
In the cell migration assay, ZDHHC14-overexpressed cells migrated
significantly more rapidly compared to empty vector-transfected
control cells after cell scraping (Fig.
3D).
Discussion
In the present study, we focused on the novel target
gene ZDHHC14 among genes specifically upregulated in our previously
reported scirrhous type GC CAST library, derived from surgically
resected GC tissue samples (unpublished data). Our rationale for
the in-depth analysis of ZDHHC14 was 2-fold: firstly, quantitative
RT-PCR analysis revealed that ZDHHC14 was more frequently
overexpressed in GC tissue than in non-neoplastic gastric mucosa.
Secondly, the biological role of ZDHHC14 in the majority of human
cancers has not been elucidated. We showed that ZDHHC14 expression
was associated with undifferentiated type GC, scirrhous type GC
variant and greater depth of invasion by qRT-PCR analysis on 123
human GC cases. Forced expression of ZDHHC14 in GC cell lines
promoted cancer cell migration and cell attachment in vitro,
and also stimulated cancer cell invasion. Furthermore, knockdown of
ZDHHC14 by RNAi inhibited integrin α5 and β1 expression and exerted
effects opposite to those observed in the stably-overexpressing GC
cell line. Taken together, these results suggest that ZDHHC14
constitutes a potential therapeutic target in GC.
ZDHHC14 encodes zinc finger DHHC domain-containing
protein 14, also known as DHHC14 protein or NEW1CP. DHHC proteins
are evolutionarily conserved genes that can be found in multiple
eukaryotes (20). There are 23
human DHHC proteins (DHHC1–DHHC24) encoded by 23 human ZDHHC genes,
named ZDHHC1–ZDHHC24 (ZDHHC10 is omitted) (21,22).
Besides the DHHC domain, these proteins also contain four or more
transmembrane domains. Although the role of DHHC14 remains to be
characterized in mammals, its Drosophila homolog has
recently been confirmed as DHHC palmitoyltransferase, which is
involved in the regulation of Fat signaling (23). Protein palmitoylation is a classical
and reversible lipid modification that has been shown to regulate
protein stability and intracellular localization, in addition to
protein-protein and protein-lipid interactions (24,25).
To our knowledge, ZDHHC14 was the first palmitoyltransferase
discovered to be upregulated in hematological cancer, through the
chromosomal translocation in patients with acute biphenotypic
leukemia. This suggests the potential regulatory role of ZDHHC14 in
cellular differentiation of leukemogenesis (13). However, the biological function and
role of ZDHHC14 gene in solid tumors is poorly understood.
Cell migration plays an important role in cancer
cell invasion and is generally presumed to be regulated by cell
adhesion molecules. For example, integrins bind extracellular
matrix (ECM) and link the ECM to intracellular signaling pathways;
the turnover of integrin focal contacts may promote cell migration
(26,27). It has previously been reported that
integrin subunits are the substrates of palmitoylation enzymes
(28). Increasing evidence
indicates that during progression from tumor growth to metastasis,
specific integrin signals enable cancer cells to detach from
neighboring cells, re-orientate their polarity during migration,
and survive and proliferate in foreign microenvironments (29). In the present study, ZDHHC14
transient knockdown revealed alteration in both mRNA and protein
expression of integrin α5 and β1 subunits. Integrin α5β1 regulates
the function of integrin αvβ3 on endothelial cells during their
migration in vitro or angiogenesis in vivo (30). However, the mechanism by which
ZDHHC14 transcriptionally downregulates these integrin subunits
remains unclear. On the other hand, ZDHHC14 knockdown affects
MMP-17 expression. Reports that the stem region of MMP-17 contains
two cysteine residues of unknown function suggest that these sites
might play an important role in regulation of protease function
since cysteine residues are known sites for a variety of
post-translational modifications including palmitoylation,
disulfide bond formation and prenylation (31). ZDHHC14 may therefore have a critical
role in cell migration, although the precise molecular mechanism
needs to be clarified. The data in the present study verified that
ZDHHC14 is associated with cancer progression and invasion of
cancer cells by mediating cell adhesion and migration activity.
Results of our clinicopathologic parameter analyses in surgically
resected GC tissues are consistent with the cell adhesion and
migration data.
Collectively, ZDHHC14 is a promising therapeutic
target for management of stomach cancer, particularly scirrhous
type GC. Identification of the substrates associated with ZDHHC14
via use of strategies such as siRNA knockdown and overexpression in
stable GC cell lines will provide further insights into GC
pathobiology and open new therapeutic avenues for more effective
treatment of GC. Subsequent studies examining the specific
substrates of the ZDHHC14 and other novel candidate genes may
further elucidate the molecular mechanisms of the genes
contributing to GC and its progression.
Acknowledgements
We thank Mr. Shinichi Norimura for his excellent
technical assistance and advice. This study was carried out with
the kind cooperation of the Research Center for Molecular Medicine
and Analysis Center of Life Sciences, Hiroshima University. This
study was supported in part by Grants-in-Aid for Cancer Research
from the Ministry of Education, Culture, Science, Sports, and
Technology of Japan, in part by a Grant-in-Aid for the Third
Comprehensive 10-year Strategy for Cancer Control and for Cancer
Research from the Ministry of Health, Labor and Welfare of Japan,
and in part by the National Cancer Center Research and Development
Fund (23-A-9).
References
|
1
|
Yasui W: Future perspectives of gastric
cancer treatment: from bench to bedside. Pathobiology. 78:293–294.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Japanese Gastric Cancer Association.
Japanese classification of gastric carcinoma: 3rd English edition.
Gastric Cancer. 14:101–112. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yanagihara K, Takigahira M, Tanaka H,
Komatsu T, Fukumoto H, Koizumi F, Nishio K, Ochiya T, Ino Y and
Hirohashi S: Development and biological analysis of peritoneal
metastasis mouse models for human scirrhous stomach cancer. Cancer
Sci. 96:323–332. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hippo Y, Yashiro M, Ishii M, Taniguchi H,
Tsutsumi S, Hirakawa K, Kodama T and Aburatani H: Differential gene
expression profiles of scirrhous gastric cancer cells with high
metastatic potential to peritoneum or lymph nodes. Cancer Res.
61:889–895. 2001.PubMed/NCBI
|
|
5
|
Jass JR, Sobin LH and Watanabe H: The
World Health Organization’s histologic classification of
gastrointestinal tumors. A commentary on the second edition.
Cancer. 66:2162–2167. 1990.
|
|
6
|
Lockhart DJ, Dong H, Byrne MC, Follettie
MT, Gallo MV, Chee MS, Mittmann M, Wang C, Kobayashi M, Horton H
and Brown EL: Expression monitoring by hybridization to
high-density oligonucleotide arrays. Nat Biotechnol. 14:1675–1680.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Velculescu VE, Zhang L, Vogelstein B and
Kinzler KW: Serial analysis of gene expression. Science.
270:484–487. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Oue N, Hamai Y, Mitani Y, Matsumura S,
Oshimo Y, Aung PP, Kuraoka K, Nakayama H and Yasui W: Gene
expression profile of gastric carcinoma: identification of genes
and tags potentially involved in invasion, metastasis, and
carcinogenesis by serial analysis of gene expression. Cancer Res.
64:2397–2405. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mitani Y, Oue N, Matsumura S, Yoshida K,
Noguchi T, Ito M, Tanaka S, Kuniyasu H, Kamata N and Yasui W: Reg
IV is a serum biomarker for gastric cancer patients and predicts
response to 5-fluorouracil-based chemotherapy. Oncogene.
26:4383–4393. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Oue N, Sentani K, Noguchi T, Ohara S,
Sakamoto N, Hayashi T, Anami K, Motoshita J, Ito M, Tanaka S,
Yoshida K and Yasui W: Serum olfactomedin 4 (GW112, hGC-1) in
combination with Reg IV is a highly sensitive biomarker for gastric
cancer patients. Int J Cancer. 125:2383–2392. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sentani K, Oue N, Sakamoto N, Arihiro K,
Aoyagi K, Sasaki H and Yasui W: Gene expression profiling with
microarray and SAGE identifies PLUNC as a marker for hepatoid
adenocarcinoma of the stomach. Mod Pathol. 21:464–475. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sentani K, Oue N, Sakamoto N, Anami K,
Naito Y, Aoyagi K, Sasaki H and Yasui W: Upregulation of connexin
30 in intestinal phenotype gastric cancer and its reduction during
tumor progression. Pathobiology. 77:241–248. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yu L, Reader JC, Chen C, Zhao XF, Ha JS,
Lee C, York T, Gojo I, Baer MR and Ning Y: Activation of a novel
palmitoyltransferase ZDHHC14 in acute biphenotypic leukemia and
subsets of acute myeloid leukemia. Leukemia. 25:367–371. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kondo T, Oue N, Yoshida K, Mitani Y, Naka
K, Nakayama H and Yasui W: Expression of POT1 is associated with
tumor stage and telomere length in gastric carcinoma. Cancer Res.
64:523–529. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yasui W, Ayhan A, Kitadai Y, Nishimura K,
Yokozaki H, Ito H and Tahara E: Increased expression of p34cdc2 and
its kinase activity in human gastric and colonic carcinomas. Int J
Cancer. 53:36–41. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ochiai A, Yasui W and Tahara E:
Growth-promoting effect of gastrin on human gastric carcinoma cell
line TMK-1. Jpn J Cancer Res. 76:1064–1071. 1985.PubMed/NCBI
|
|
17
|
Yanagihara K, Tanaka H, Takigahira M, Ino
Y, Yamaguchi Y, Toge T, Sugano K and Hirohashi S: Establishment of
two cell lines from human gastric scirrhous carcinoma that possess
the potential to metastasize spontaneously in nude mice. Cancer
Sci. 95:575–582. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Alley MC, Scudiero DA, Monks A, Hursey ML,
Czerwinski MJ, Fine DL, Abbott BJ, Mayo JG, Shoemaker RH and Boyd
MR: Feasibility of drug screening with panels of human tumor cell
lines using a microculture tetrazolium assay. Cancer Res.
48:589–601. 1988.PubMed/NCBI
|
|
19
|
Sentani K, Oue N, Naito Y, Sakamoto N,
Anami K, Oo HZ, Uraoka N, Aoyagi K, Sasaki H and Yasui W:
Upregulation of HOXA10 in gastric cancer with the intestinal mucin
phenotype: reduction during tumor progression and favorable
prognosis. Carcinogenesis. 33:1081–1088. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ohno Y, Kihara A, Sano T and Igarashi Y:
Intracellular localization and tissue-specific distribution of
human and yeast DHHC cysteine-rich domain-containing proteins.
Biochim Biophys Acta. 1761:474–483. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Linder ME and Deschenes RJ: Model
organisms lead the way to protein palmitoyltransferases. J Cell
Sci. 117:521–526. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mitchell DA, Vasudevan A, Linder ME and
Deschenes RJ: Protein palmitoylation by a family of DHHC protein
S-acyltransferases. J Lipid Res. 47:1118–1127. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Matakatsu H and Blair SS: The DHHC
palmitoyltransferase approximated regulates Fat signaling and Dachs
localization and activity. Curr Biol. 18:1390–1395. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Dunphy JT and Linder ME: Signalling
functions of protein palmitoylation. Biochim Biophys Acta.
1436:245–261. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Resh MD: Fatty acylation of proteins: new
insights into membrane targeting of myristoylated and palmitoylated
proteins. Biochim Biophys Acta. 1451:1–16. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hynes RO: Integrins: Versatility,
modulation, and signaling in cell adhesion. Cell. 69:11–25. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Moissoglu K and Schwartz MA: Integrin
signalling in directed cell migration. Biol Cell. 98:547–555. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang X, Kovalenko OV, Tang W, Claas C,
Stipp CS and Hemler ME: Palmitoylation supports assembly and
function of integrin-tetraspanin complexes. J Cell Biol.
167:1231–1240. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Guo W and Giancotti FG: Integrin
signalling during tumour progression. Nat Rev Mol Cell Biol.
5:816–826. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Dormond O and Rüegg C: Regulation of
endothelial cell integrin function and angiogenesis by COX-2, cAMP
and Protein Kinase A. Thromb Haemost. 90:577–585. 2003.PubMed/NCBI
|
|
31
|
Sohail A, Sun Q, Zhao H, Bernardo MM, Cho
JA and Fridman R: MT4-(MMP17) and MT6-MMP (MMP25), a unique set of
membrane-anchored matrix metalloproteinases: properties and
expression in cancer. Cancer Metastasis Rev. 27:289–302. 2008.
View Article : Google Scholar : PubMed/NCBI
|