Introduction
Ovarian cancer is the most lethal gynecologic
cancer, causing annually a large number of deaths throughout the
world. Early stages of ovarian cancer are often asymptomatic such
that the disease is usually diagnosed after it has disseminated
beyond the ovaries (1). In fact,
~70% of ovarian cancer patients present with advanced-stage disease
(2). Although patients usually
respond to initial therapy and cytoreductive surgery followed by
adjuvant chemotherapy with platinum and paclitaxel (3), ~70% of those with advanced-stage
ovarian cancer experience recurrence. In many of these cases, the
disease is incurable mainly due to the development of drug
resistance (4).
Drug resistance can be intrinsic or acquired and
arises from a variety of factors, including genetic differences in
somatic cell tumors and individual patient-specific variations
(5). In culture, cancer cells have
been shown to become resistant to cytotoxic anticancer drugs
through several different mechanisms, such as efflux pumps, cell
cycle defects, apoptosis, DNA damage, metabolic alterations,
changes in drug targets and compartmentalization (5). Some of the molecular mechanisms
underlying the development of resistance have been characterized in
cellular models of ovarian cancer. Johnson et al (6) described three general pathways;
increased DNA damage tolerance/repair, decreased cell-drug
associations and altered drug inactivation. All of these are
thought to be involved in platinum-based resistance mechanisms in
ovarian cancer. According to Sorrentino et al (7), drug resistance in ovarian cancer
reflects the deregulation of growth factor receptor expression,
defective DNA damage response, increased DNA repair activity and
the increased activity of anti-apoptotic regulators. A more recent
study suggested that targeting glycolytic, mitochondrial apoptotic
and necrotic pathways offers promising therapeutic strategies in
ovarian cancer (3). However,
despite the large number of studies and increasing insight into the
mechanisms of drug resistance in this disease, the 5-year survival
rate of patients is only ~40%, and only modest improvement has been
made over the past 40 years (2).
Tumor suppressor genes (TSGs) play regulatory roles
in diverse cellular activities, including cell cycle checkpoint
responses, mitogenic signaling, DNA damage repair, cell
proliferation, differentiation, migration, protein ubiquitination
and tumor angiogenesis (8,9). TSGs have also been implicated in the
drug resistance of ovarian cancer and their loss of function
contributes to the development of cancer in general (9,10). For
example, expression of tumor suppressor PTEN is lower in
drug-resistant than in drug-sensitive ovarian cancer cells. The
overexpression of PTEN by transfection was shown to reverse
chemoresistance to cisplatin, by inactivating the PI3K/AKT cell
survival pathway (11). The TSGs
BRCA1 and BRCA2 have been convincingly linked to ovarian cancer,
and tumors with BRCA1/2 mutations frequently develop platinum
resistance (12). In a literature
review, Yin et al (10)
identified 15 TSGs (such as BRCA1, PTEN and p53) that play critical
roles in drug resistance and that, consequently, may serve as
molecular targets for the treatment of chemoresistant ovarian
cancer. Therefore, the targeting of drug resistance-related TSGs
would provide the first steps in developing reasonable and feasible
methods to meet the challenges posed by drug resistance in ovarian
cancer.
TSGs may be of the class I or class II type. Class I
TSGs lose function by deletion, rearrangement or mutation (13); class II TSGs are structurally intact
in their sequence, yet are underexpressed or unexpressed due to
downregulation or silencing in transcription or translation
(14). Ribonuclease T2
(RNASET2) is a class II TSG associated with the suppression
of ovarian cancer (15); it belongs
to a group of genes encoding RNases and is highly conserved among
species (16). RNases, including
RNASET2, have many different functions, with a particularly
important one being the regulation of cell growth (17). Consistent with these properties is
the observation that in some human malignancies, such as lymphomas
and glioma, RNASET2 expression is downregulated at the
transcriptional level (18,19). Monti et al (20) showed an apparent decrease in the
antitumorigenic activity of RNASET2 in melanoma cancer cells. In
ovarian cancer, RNASET2 expression was shown to be
significantly reduced in 30% of primary ovarian tumors and in 75%
of ovarian tumor cell lines (15).
Conversely, the transfection of RNASET2 into ovarian tumor
cell lines suppressed their tumorigenicity in nude mice and induced
marked cell senescence during in vitro growth (15). Besides its antitumorigenic activity,
RNASET2 significantly reduces the metastatic potential of
ovarian cancer (21), in addition
to controlling ovarian tumorigenesis by modifying the cellular
microenvironment and by inducing immunocompetent cells of the
monocyte/macrophage lineage (22).
Collectively, these studies indicate RNASET2 is a poorly
explored member of the growing family of TSGs (22).
Gametogenetin binding protein 2 (GGNBP2), a
potential TSG, is a highly conserved gene localized to the
tumor-suppressor locus D17S800-D17S930 but also known to be
involved in spermatogenesis (23).
This gene is also known as ZNF403 and as laryngeal carcinoma
related gene 1 (LCRG1), in the latter case since it was
cloned from a laryngeal carcinoma (24). The expression of GGNBP2 is
significantly reduced in 40% (12/30) of primary laryngeal
carcinomas, where its primary tumor suppressive functions include
control of the cell proliferation rate and of soft agar growth,
tumor formation and tumor cell migration (24,25).
GGNBP2 knockdown alters the expression profiles of certain
genes, such as upregulating p21 and downregulating MCM2. This
participation in cell cycle regulatory pathways suggests that
GGNBP2 contributes to the regulation of the G2/M transition of the
cell cycle (25). In fact, cell
cycle analysis demonstrated that the downregulation of
GGNBP2 promotes G2/M cell cycle arrest in a dose-dependent
manner. Regarding the involvement of GGNBP2 in ovarian cancer, one
study reported downregulation of the gene in six of ten different
ovarian cancer cell lines (24).
Although RNASET2 and GGNBP2 are important TSGs and
disturbances in their expression have been implicated in the
development of ovarian and other types of cancer, the literature
contains no reports of their specific association with drug
resistance. A microarray analysis of carboplatin-sensitive and
-resistant ovarian cancer cells carried out in our laboratory
showed that the expression of RNASET2 and GGNBP2 is
significantly decreased in resistant cells (26), indicating the involvement of these
two genes in drug resistance. In the present study, on the basis of
the results of real-time quantitative polymerase chain reaction
(RT-qPCR) and comprehensive bioinformatics analysis, we were able
to show, for the first time, that the downregulation of
RNASET2 and GGNBP2 in drug-resistant ovarian cancer
tissues/cells contributes to the regulation of drug resistance in
ovarian cancer.
Materials and methods
Cell culture and tissue samples
The human epithelial ovarian cancer cell line A2780
was maintained in our laboratory and propagated in vitro by
serial passage in RPMI-1640 supplemented with 10% fetal bovine
serum. The A2780-CBP and A2780-DDP cell lines were developed by
sequential exposure of A2780 cells to increasing concentrations of
carboplatin and cisplatin, respectively. The resistance index of
A2780-CBP and -DDP ovarian cancer cells was 2.0 and 1.8,
respectively.
Ovarian cancer tissues were collected from patients
surgically treated for primary epithelial ovarian cancer at the
Department of Gynecologic Oncology, Affiliated Tumor Hospital of
Guangxi Medical University. The tissue samples were evaluated by
pathologists. All patients underwent conventional chemotherapy with
6–8 postoperative cycles of paclitaxel plus cisplatin (TP) or
paclitaxel plus carboplatin (TC) after optimal cytoreductive
surgery. They were followed up for a minimum of 1 year after the
completion of chemotherapy. In addition, the patients were assigned
to either the chemotherapy-resistant group (n=10) or the
chemotherapy-sensitive group (n=10) in accordance with the 2013
guidelines National Comprehensive Cancer Network (NCCN). The median
age of the patients was 50.8 years (range, 35–73 years). All of the
malignant tissues were from stage II-III tumors, according to the
International Federation of Gynecology and Obstetrics (FIGO)
classification. The study was endorsed by the Ethics Committee of
Guangxi Medical University. All patients received an explanation
concerning the aims of the study and provided signed informed
consent. Samples were collected from primary lesions during surgery
and stored in a liquid nitrogen tank until they were used for mRNA
isolation.
RT-qPCR analysis
Total RNA from ovarian cancer cells/tissues was
isolated using TRIzol reagent and digested with amplification-grade
DNase I (both from Invitrogen, Carlsbad, CA, USA) prior to cDNA
synthesis. The first-strand cDNA was synthesized using the
Transcriptor First Strand cDNA Synthesis kit (Roche, Germany).
RT-qPCR was performed with the FastStart Universal SYBR-Green
Master kit (Roche). Data were collected with the StepOnePlus
Real-Time PCR System (ABI, Foster City, CA, USA) in accordance with
the manufacturer’s instructions. The RT-qPCR gene-specific primers
for RNASET2 were: (forward primer)
5′-ACCCTCCGGATTACTGGACA-3′ and (reverse primer)
5′-TGCGATTGGGAAACGAGTGA-3′; for GGNBP2, the forward primer
was 5′-ATCACGCGAAGTCCTGAGTG-3′ and the reverse primer was
5′-GGGAAAAGAGACGCTCCACA-3′. For GAPDH, used as the control,
the forward primer was 5′-GAAGGTGAAGGTCGGAGT-3′ and the reverse
primer 5′-GAAGATGGTGATGGGATTT-3′.
Bioinformatics analysis
Microarray data on gene expression were retrieved
from Gene Expression Omnibus (GEO) Profiles (http://www.ncbi.nlm.nih.gov/geoprofiles/). The
gene/protein-gene/protein interaction network was generated with
GeneMANIA (http://www.genemania.org/) (25); the annotation of biological
processes was analyzed with the Coremine Medical online tool
(http://www.coremine.com/medical/). The
miRWalk online tool (http://www.umm.uni-heidelberg.de/apps/zmf/mirwalk/)
was used for miRNA target-gene prediction (27). Data on genes co-expressed with
RNASET2 and GGNBP2 according to the TCGA Ovarian 2
database were obtained from the Oncomine online database
(https://www.oncomine.org/resource/login.html). The
pathway enrichment was performed using the DAVID online tool
(http://david.abcc.ncifcrf.gov/)
(28,29). The pathways in which the genes were
involved were analyzed using the online database Kyoto Encyclopedia
of Genes and Genomes (KEGG) (http://www.genome.jp/kegg/).
Results
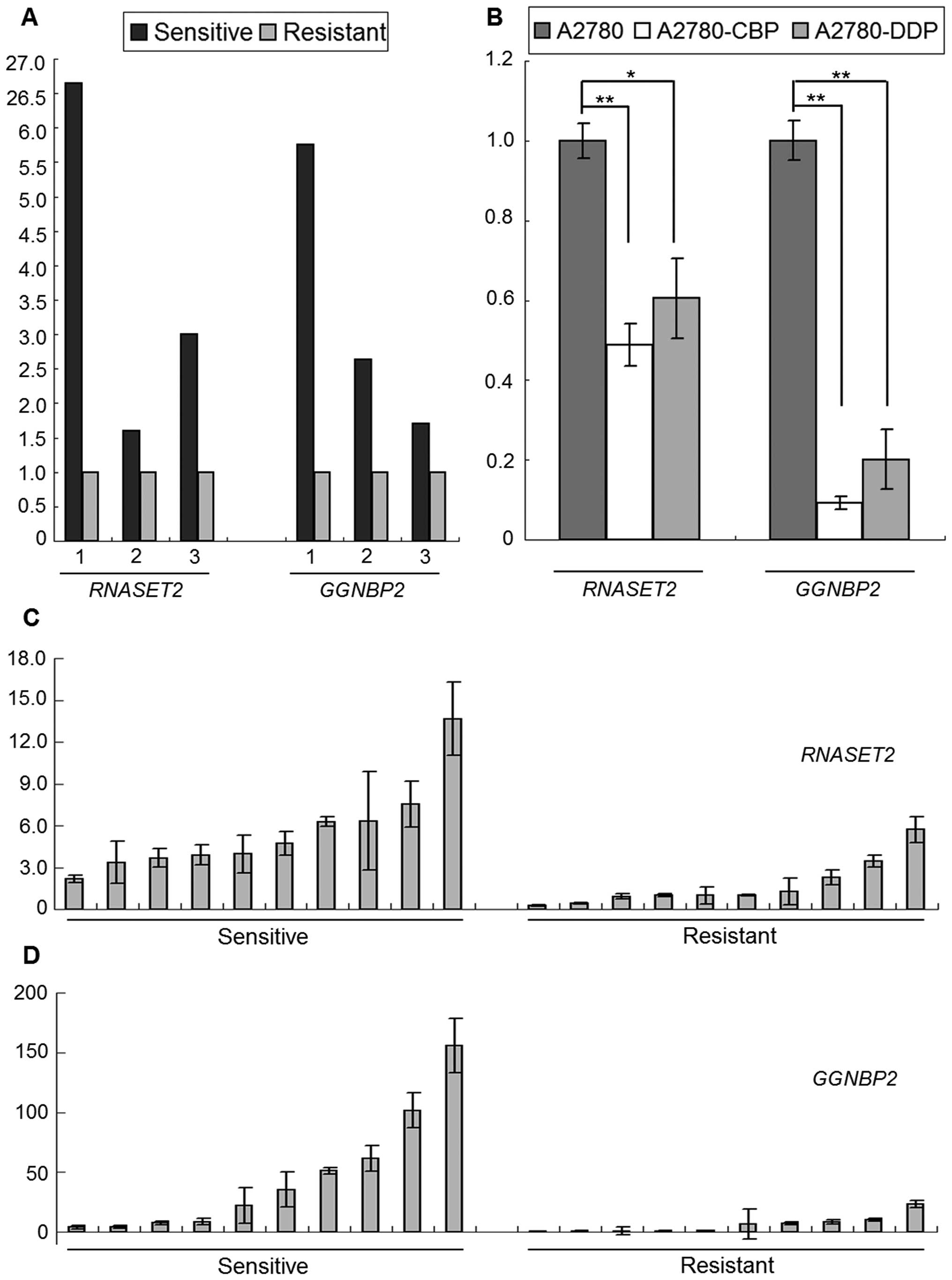
RNASET2 and GGNBP2 mRNA expression is
significantly downregulated in drug-resistant ovarian cancer
cells/tissues
Based on the microarray data obtained in our
laboratory (26) and the data
retrieved from the GEO database (Fig.
1A), the expression of RNASET2 and GGNBP2 was
significantly lower (at least 1.6-fold) in platinum- and
vincristine-resistant ovarian cancer cells than in their
drug-sensitive counterparts. This result was confirmed by measuring
the expression of the two genes in all three ovarian cancer cell
lines. Specifically, the RT-qPCR results showed large decreases in
RNASET2 and GGNBP2 mRNA levels in A2780-CBP and -DDP
cancer cells when compared with A2780 cells. Indeed, GGNBP2
expression was reduced by at least 5-fold in resistant cells
(Fig. 1B). Further study of these
genes in ovarian cancer tissues revealed that RNASET2 and
GGNBP2 mRNA levels were significantly lower in
drug-resistant than in drug-sensitive tissues (Fig. 1C and D). Compared with the average
expression levels in ten sensitive cancers, those of RNASET2
and GGNBP2 in ten resistant cancers were decreased 3.2- and
7.4-fold, respectively. Collectively, the data on drug-resistant
cells as provided in the GEO database, the results of our
experiments in cultured drug-resistant-A2780 cells, and our
analyses of drug resistance in ovarian cancer tissues consistently
showed that the expression of RNASET2 and GGNBP2 mRNA
is downregulated in ovarian cancer, indicating a role for these
genes in the development of drug-resistant variants of these
tumors.
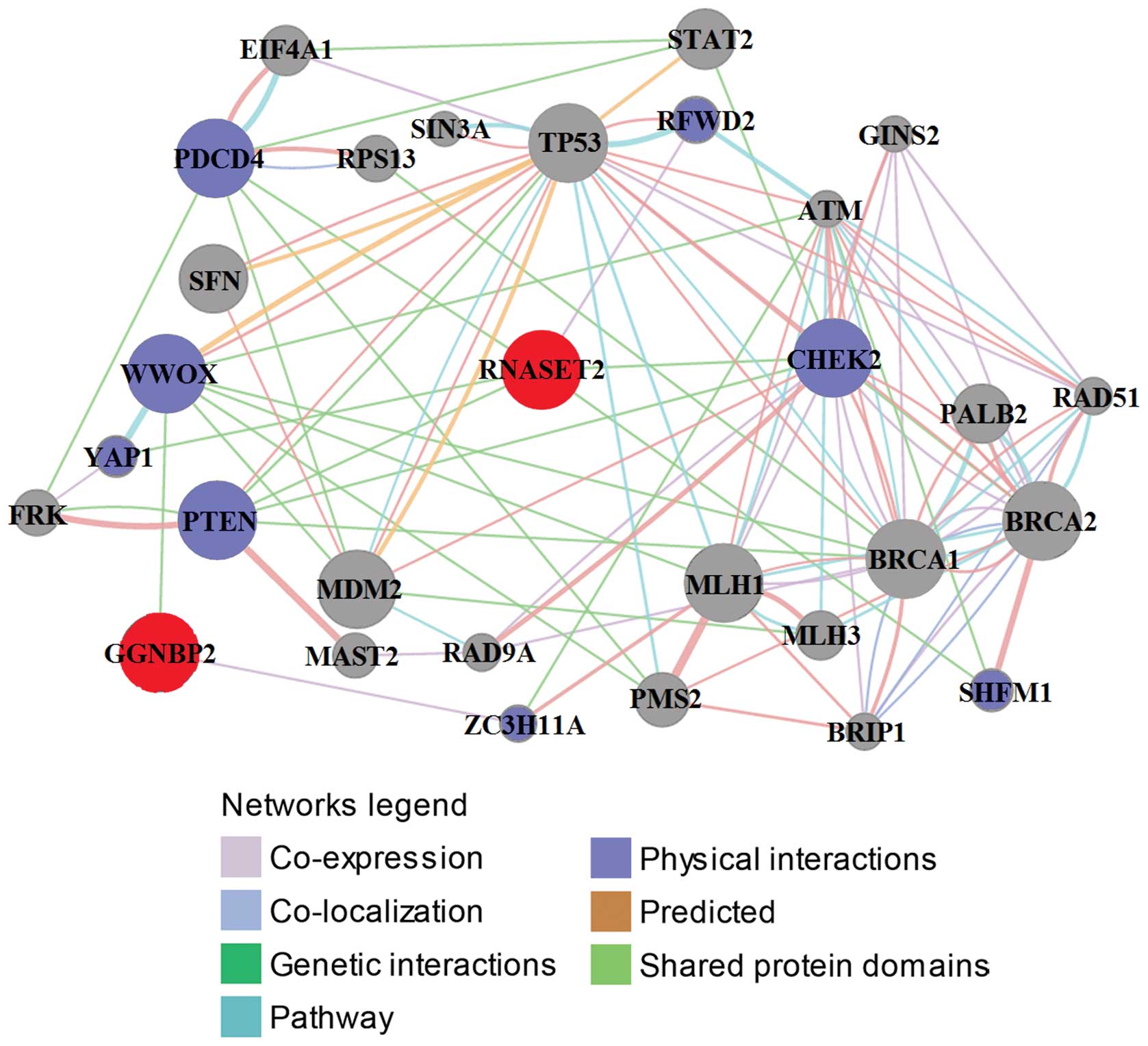
Prediction and analysis of function based
on gene/protein-gene/protein interactions
The drug resistance-related functions of RNASET2 and
GGNBP2 were analyzed using GeneMANIA (Fig. 2). RNASET2 was co-expressed,
co-localized, physically interacted and shared protein domains and
pathways directly/indirectly with a number of proteins.
Specifically, direct genetic interactions were established between
RNASET2 and checkpoint kinase 2 (CHEK2), programmed cell death 4
(PDCD4), phosphatase and tensin homolog (PTEN), split hand/foot
malformation (ectrodactyly) type 1 (SHFM1) and Yes-associated
protein 1 (YAP1). In addition, RNASET2 was found to be co-expressed
directly with ring finger and WD repeat domain 2 (RFWD2). These
results suggested that these genes/proteins and RNASET2 are
functionally related.
CHEK2 is one of the critical kinases governing cell
apoptosis, DNA damage repair and cell cycle checkpoint control in
response to DNA damage signals. It is degraded at the protein level
in response to cisplatin, through a ubiquitin-proteasome pathway.
In ovarian cancer, the expression of CHEK2 is decreased in
cisplatin-resistant ovarian cancer cell lines, suggesting a role
for this kinase in chemoresistance (30).
PDCD4 is another TSG and its expression was shown to
enhance chemosensitivity in renal and prostate cancer cell lines
(31,32). In ovarian cancer, PDCD4 promotes
cisplatin-induced apoptosis, mainly by activating the death
receptor pathway and PDCD4 gene transfer in combination with
cisplatin therapy may break the chemotherapeutic resistance of
ovarian cancer cells (33).
PTEN is a well-known TSG associated with cancer
development through the ERK1/2 signaling (34) and PI3K/Akt/mTOR (35,36)
pathways. It interacts with TSGs (such as p53 and BRCA1) that
contribute to the development of drug resistance in several types
of cancer (37). In ovarian cancer,
PTEN contributes to multidrug resistance through cell cycle
regulation, apoptosis and the PI3K/Akt pathway (38,39).
Yuan et al (40) demonstrated that YAP1 is a TSG.
Knockdown of the gene in breast cell lines resulted in several
abnormal cell behaviors, including inhibition of the response of
these cells to taxol. Accordingly, the authors concluded that YAP1
plays a role in drug resistance.
SHFM1 is a TSG required for the stability of BRCA2
(41), yet it has also been
implicated in acquired drug resistance in ovarian cancer (42).
Finally, RFWD2, also known as constitutive
photomorphogenic 1 (COP1), is a newly discovered TSG (43,44).
As reported by Migliorini et al (44), it exerts its tumor suppressor
function in part by antagonizing c-Jun oncogenic activity. A recent
study suggested RFWD2 as a target for gene therapy in the treatment
of gastric cancer, suggesting the involvement of this protein in
drug resistance in cancer (45).
GGNBP2 is associated with fewer genes/proteins than
RNASET2. As shown in Fig. 2, GGNBP2
is co-expressed only with zinc finger CCCH-type containing 11A
(ZC3H11A) and genetically interacts with WW domain containing
oxidoreductase (WWOX). ZC3H11A is a poorly characterized protein
that belongs to a large family of zinc finger proteins, with 58
known members (46). Zinc finger
proteins are known to mediate the epithelial-mesenchymal
transition, which in lung cancer is related to drug resistance
(47). Thus, the function of
ZC3H11A, as a member of the zinc finger protein family, may also be
drug resistance-related. As a TSG, WWOX plays important roles in
ovarian and many other types of cancers (48). For example, the overexpression of
WWOX preferentially inhibits the viability of human glioblastoma
cells expressing mutant p53 and induces apoptosis via a mechanism
independent of the intrinsic apoptotic pathway (49). Conversely, WWOX suppression by RNA
interference reverses platinum resistance in DDP-resistant SKOV3
ovarian cancer cells (50).
In the present study, we determined direct
interactions of six genes/proteins with RNASET2 and two with
GGNBP2, and in all cases these were linked to drug resistance in
ovarian and other types of cancers. In addition, indirect
interactions between several TSGs, such as TP53, MLH1, MLH3, BRCA1,
and BRCA2, and RNASET2, and GGNBP2 were demonstrated. All of these
TSGs are linked to drug resistance in ovarian cancer (10), a relationship that supports our
conclusion that RNASET2 and GGNBP2 are associated with drug
resistance in ovarian cancer.
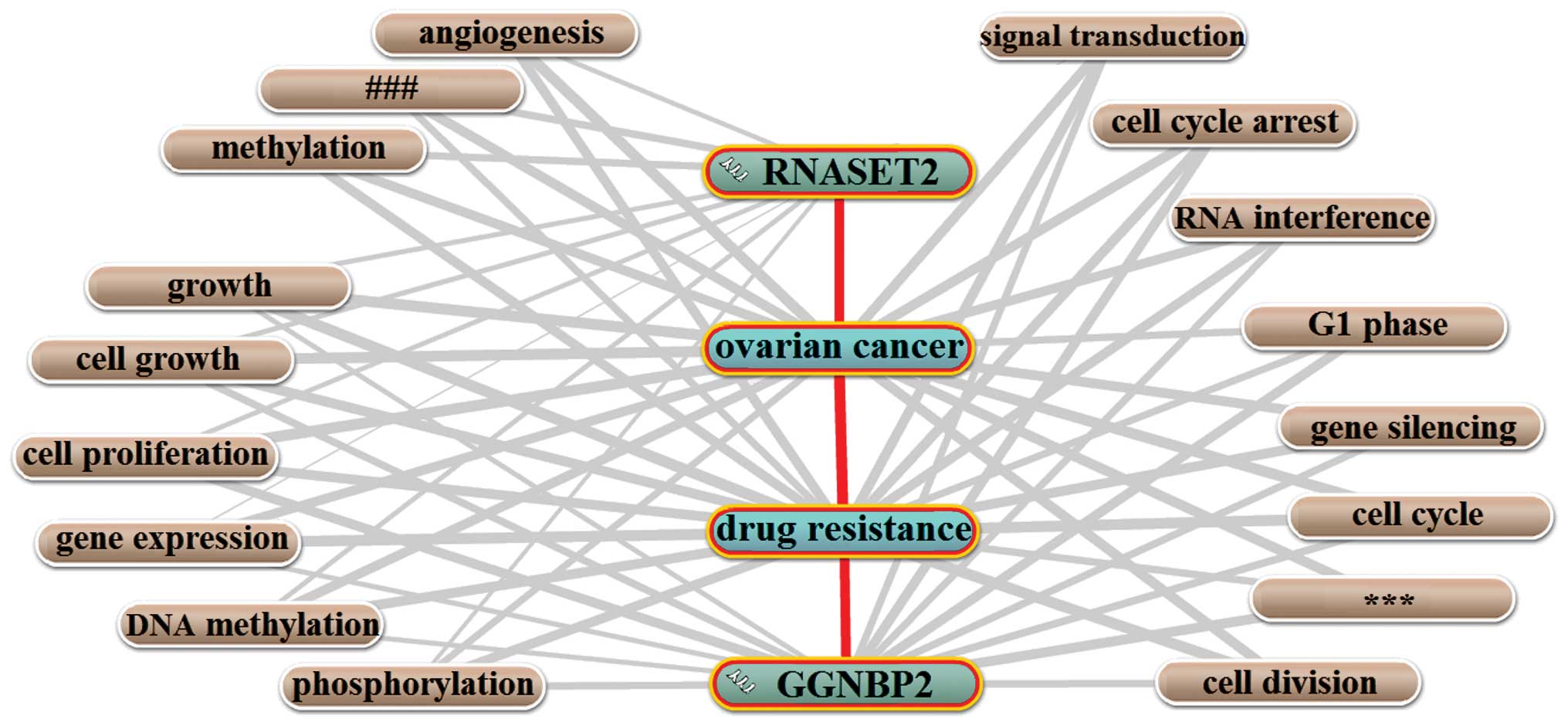
Functional prediction based on the
annotated biological process
In Coremine Medical (51), gene and protein names are
cross-referenced to terms relevant to an understanding of their
biological function and importance in disease. We therefore used
the Coremine online tool and probabilistic scoring (p<0.009) to
annotate RNASET2 and GGNBP2 with respect to biological processes.
As shown in Fig. 3, 17 biological
processes closely associated with ovarian cancer, drug resistance,
and the two genes were annotated, suggesting their mutual
involvement in drug resistance in ovarian cancer. Among the nine
processes annotated to RNASET2, three were related to growth, cell
growth and cell proliferation, consistent with the theory that the
drug resistance-related functions of RNASET2 mainly proceed through
the regulation of cell growth and the RNase activity of RNASET2
(17). GGNBP2 annotated with 14 of
the 17 processes: three (growth, cell growth and cell
proliferation) were cell-growth related, four (gene expression, DNA
methylation, RNA interference and gene silencing) were
gene-expression related, and three (cell cycle arrest, G1 phase and
cell cycle) were cell cycle related. Of these, the relationship
between the cell cycle-related function of GGNBP2 and drug
resistance is in agreement with a previous study in which GGNBP2
was shown to be involved in cell cycle regulation during cancer
development (25).
Functional prediction based on the
functionality of the miRNAs that target them
Among the transcriptional targets of GGNBP2 and
RNASET2 were 228 and 179 miRNAs, respectively, as predicted through
miRWalk, which is made up of ten miRNA-mRNA prediction tools. Eight
miRNAs, i.e., those yielding the highest score for each gene, were
selected for subsequent analysis (Table
I). As shown in Table I, except
for hsa-miR-323-3p, all the miRNAs that targeted GGNBP2 influenced
drug resistance in ovarian and other types of cancers. For RANSET2,
although only hsa-miR-758 and hsa-miR-22 were specifically involved
in drug resistance in ovarian and breast cancer, three other miRNAs
(hsa-miR-320a, hsa-miR-320d and hsa-miR-1257) contributed to drug
resistance-related processes, such as cell proliferation, tumor
invasion and cell differentiation. Collectively, among eight miRNAs
most strongly targeting RNASET2 and especially GGNBP2, the majority
were involved in drug resistance in ovarian and other cancers,
suggesting that the two genes also mediate drug resistance.
 | Table IThe 8 miRNAs most strongly targeting
GGNBP2 and RNASET2, and their functions in cancer. |
Table I
The 8 miRNAs most strongly targeting
GGNBP2 and RNASET2, and their functions in cancer.
| Gene name | MicroRNA | miRNA-gene
interaction prediction tools | Drug resistance and
related functions in cancer (ref.) |
|---|
|
|---|
| A | B | C | D | E | F | G | H | I |
|---|
| GGNBP2 | hsa-miR-135a | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | Drug resistance in
breast cancer (52) |
| hsa-miR-23b | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | Drug resistance in
ovarian cancer (53) and breast
cancer (52) |
| hsa-miR-135b | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | Drug resistance in
esophageal cancer (54) |
| hsa-miR-23a | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 0 | 1 | Drug resistance in
ovarian cancer (55) |
| hsa-miR-150 | 1 | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 1 | Drug resistance in
breast cancer (52) |
| hsa-miR-323-3p | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 0 | 1 | - |
| hsa-miR-210 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 0 | 1 | Drug resistance in
breast cancer (56) |
| hsa-miR-224 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 0 | 1 | Drug resistance in
lung cancer cells (57) and gastric
cancer cells (58) |
| RNASET2 | hsa-miR-758 | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 1 | Drug resistance in
breast cancer (52) |
| hsa-miR-320c | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 1 | - |
| hsa-miR-320a | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 1 | Cell proliferation
in colon cancer cell (59) and
tumor invasion in colorectal cancer (60) |
| hsa-miR-320d | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 1 | Regulation of stem
cell differentiation in colon cancer stem cells (61) |
| hsa-miR-320b | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 1 | - |
| hsa-miR-635 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | - |
| hsa-miR-22 | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | Drug resistance in
ovarian cancer (62) |
| hsa-miR-1257 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | Tumor
dedifferentiation and/or progression in liposarcoma (63) |
Potential pathways by which RNASET2 and
GGNBP2 contribute to drug resistance
Pathway enrichment of the genes co-expressed with
RNASET2 and GGNBP2 in drug-resistant ovarian cancer cells was
performed to mine the possible pathways in which these two TSGs may
exercise their drug resistance-related functions in ovarian cancer.
Genes co-expressed with RNASET2 and GGNBP2 in eight
irinotecan-resistant, four paclitaxel-resistant and five
topotecan-resistant ovarian cancer cells were retrieved from the
Oncomine online database. Of the co-expressed genes, the 50 genes
with the highest correlation efficiency with GGNB2 and RNASET2 were
selected for further analysis (Table
II). For GGNBP2, two pathways (cytokine-cytokine receptor
interaction and chemokine signaling pathway) were found to be
enriched, as determined using DAVID (28,29),
whereas with RNASET2 no enriched pathway was identified. Thus, a
basic search in the KEGG database was performed to retrieve the
pathways to which the co-expressed genes belonged. This resulted in
the identification of 12 RNASET2-associated genes and 19
GGNBP2-associated genes as a member of certain pathways, several of
which could be related to drug resistance in ovarian and other
types of cancers (Table II).
| Table IIThe 50 genes most strongly
co-expressed with RNASET2 and GGNBP2 in drug-resistant ovarian
cancer cells and their drug resistance-associated pathways. |
Table II
The 50 genes most strongly
co-expressed with RNASET2 and GGNBP2 in drug-resistant ovarian
cancer cells and their drug resistance-associated pathways.
| Genes co-expressed
in drug-resistant ovarian cancer cellsa (correlation efficiency >0.873) | Annotated
pathwayb | Genes belong to
certain pathway based on the KEGG databasec and the pathways were involved in the
drug resistance in ovarian and other cancers (ref.) |
|---|
| RNASET2 |
| FGFR1OP,
CCR6, T, PRR18, SFT2D1,
BRP44L, | - | CCR6, I
(69) and II (71) |
| RPS6KA2,
C6orF118, PDE10A, UNC93A, TTLL2,
TCP10, | | PARK2, III
(77) |
| GPR31,
TCP10L2, QKI, PACRG, PARK2,
SMOC2, | | THBS2, IV
(77) and V (39,78) |
| DACT2,
PSMB1, TBP, PDCD2, FAM120B,
DLL1, THBS2, | | TCP1, V
(39,78) |
| PHF10,
WDR27, C6orf120, TCTE3, C6orf70,
HGC6.3, | | PSMB1, VI
(30,79) |
| KIF25,
FRMD1, C6orf124, MLLT4, MAP3K4,
AGPAT4, | | MAP3K4, VII
(80–83) |
| SOD2,
WTAP, TCP1, ACAT2, PNLDC1,
MRPL18, MAS1, | | |
| IGF2R,
LPA, LPAL2, SLC22A3, SLC22A2,
SLC22A1 | | |
| GGNBP2 |
| DHRS11,
MYO19, PIGW, MRM1, ZNHIT3, AATF,
LHX1, | I | PIGW, VIII
(3) |
| SLFN13,
SLFN11, SLFN12, CCL3, CCL23,
CCL18, | II | CCL3,
CCL5, CCL7, CCL1, CCL8 and
CCL11, I (69), |
| MMP28,
TAF15, C17orf50, CCL5, C17orf66,
GAS2L2, | | II (71) |
| RASL108,
LYZL6, RDM1, CCL16, CCL14,
AP2B1, | | CCL23,
CL18, CCL16 and CCL14, I (69) |
| SLFN12L, PEX12,
SLFN14, SLFN5, AMAC1, C17orf75, | | CCL2, I
(69) and IX (73,74) |
| ZNF207,
RHBDL3, PSMD11, CDK5R1, MYO1D, | | CCL13, IX
(73,74) |
| TMEM98,
LRRC37B, RHOT1, UTP6, C17orf79,
SUZ12, | | AP2B1, X
(84) |
| C17orf102,
TMEM132E, CCL7, CCL1, CCL13,
CCL8, | | |
| CCL11,
CCL2 | | PSMD11, VI
(30,79) and XI (85) |
For example, the genes THBS2 and TCP1,
co-expressed with RNASET2, are part of the PI3K/Akt signaling
pathway, which suggested that it is among the pathways used by this
TSG to regulate drug resistance. In fact, a strong association
between PI3K/Akt signaling and drug resistance has been shown in
ovarian cancer (11,39). Pathway enrichment analysis suggested
that the hypomethylation-mediated activation of cell
growth-promoting pathways, including PI3K/Akt, contributes to the
onset of chemoresistance in ovarian cancer cells (64). Studies on the genes involved in the
activation of this pathway identified two mechanisms, PTEN
reduction and PIK3CA mRNA amplification, that finally contribute to
cisplatin resistance in an ovarian cancer cell line (39). Similarly, the chemoresistance caused
by interleukin (IL)-6 and -8 expression is associated with the
activation of several pathways, including PI3K/Akt (65,66).
Accordingly, altered RNASET2 expression in drug-resistant ovarian
cancers may well contribute to chemoresistance by activating the
PI3K/Akt signaling pathway.
The cytokine-cytokine receptor interaction and
chemokine signaling pathways were enriched in the 50 genes most
highly co-expressed with GGNBP2 in drug-resistant ovarian cancer
cells, suggesting that GGNBP2 contributes to drug resistance via
these two pathways. In tumorous spleens, downregulated genes that
have been linked to transformation are mainly associated with the
cytokine-cytokine receptor interaction pathway (67); similarly, most of the genes
associated with lung carcinogenesis are annotated to this same
pathway (68). Cytokine-cytokine
receptor interactions with drug resistance are thought to proceed
via tumor-stroma communication, as shown in primary tumors with a
high risk of relapse (69). In a
non-small cell lung cancer cell line, functional annotation showed
that genes with altered expression induced by docetaxel are
strongly co-related with several pathways, including
cytokine-cytokine receptor interactions (70). These results provide evidence of the
interaction between the cytokine-cytokine receptor pathway and the
drug resistance of several types of tumor.
Crosstalk networks among pathways associated with
prostate cancer (primary and metastatic), by integrating
protein-protein interactions and KEGG pathways, point to the
regulation of multiple processes, including drug resistance, by the
chemokine signaling pathway during cancer progression (71). Oxaliplatin-induced chemokine
signaling was shown to reinforce NF-κB transcriptional activity,
thus potentiating the resistance of prostate cancer cells to this
platinum-based drug (72). The
NF-κB signaling pathway contains two genes (CCL2 and
CCL13) that are co-expressed with GGNBP2 in drug-resistant
ovarian cancer cells. Many studies have shown that this pathway
plays a critical role in anti-apoptosis and the drug resistance of
tumor cells during chemotherapy (73). For example, curcumin downregulates
the multidrug-resistance mdr1b gene by inhibiting the
PI3K/Akt and NF-κB signaling pathways in multidrug-resistant
L1210/Adr cells (74). Pre-clinical
models demonstrated that many chemotherapy drugs, such as
platinum-based agents, anthracyclines and taxanes, promote NF-κB
pathway activation, which finally leads to chemoresistance through
the subsequent activation of a multitude of mediators including
anti-apoptotic genes (75). In
ovarian cancer, NF-κB functions as an oncogene in drug-resistant
ovarian cancer cells and exerts its anti-apoptotic effects by
repressing mitogen-activated protein kinase phosphorylation in
aggressive chemoresistant cell lines (76).
Thus, some of the genes co-expressed with RNASET2
and GGNBP2 in drug-resistant ovarian cancer cells belong to
pathways associated with drug resistance, suggesting the similar
involvement of these two TSGs. In particular, GGNBP2 may
participate in the regulation of drug resistance in ovarian cancer
via cytokine-cytokine receptor interactions, chemokine signaling
and NF-κB signaling, whereas for RNASET2, the PI3K/Akt pathway is
relevant.
Discussion
Drug resistance is an important obstacle to
successful chemotherapeutic treatment in ovarian cancer and causes
treatment failure in over 90% of patients with metastatic disease
(86). Several mechanisms,
including DNA damage repair and disruptions in the cell cycle,
apoptosis and metabolism, are associated with the rise of drug
resistance in ovarian cancer (3,7),
mostly through aberrant gene expression. As key regulators of genes
related to drug resistance, the mining and exploration of TSGs can
provide insights into meeting the therapeutic challenge of drug
resistance in ovarian cancer. The prediction of gene function based
on comprehensive bioinformatics analysis offers a valuable tool for
gene function mining. For example, on the basis of a comprehensive
bioinformatics analysis, Yin et al (87) reported the association of SPARCL1
and CCL21 with drug resistance in ovarian cancer. In another study,
15 TSGs associated with drug resistance in ovarian cancer were
analyzed by comprehensive bioinformatics to better understand the
relationships of these genes to drug resistance (10).
The Gene Expression Omnibus (GEO), a public
repository at the National Center for Biotechnology Information
(NCBI), stores tens of millions of expression profiles, including
those obtained from microarray-based experiments measuring mRNA
abundance (88). The identification
of large-scale networks of molecular interactions within the cell
has highlighted the need to go beyond one-dimensional approaches to
study gene/protein function (89).
One such alternative is GeneMANIA, a fast and highly accurate
web-based database and tool for the prediction of gene function on
the basis of multiple networks, as determined from varied genomic
or proteomic data/sources (90).
Coremine Medical is a product of the PubGene Company, which itself
grew out of the PubGene online tool, a gene/protein database and
web-based tool for literature mining. By the automated analysis of
titles and abstracts in over 10 million MEDLINE records, Coremine
Medical creates a gene-to-gene co-citation network for millions of
named human genes. The identified relationship networks provide
both an overview of related terms and an intuitive system for
exploring the information space (91). In the present study, the GEO
database was used to retrieve data on RNASET2 and
GGNBP2 expression in drug-sensitive and drug-resistant
ovarian cancer cells; GeneMANIA and Coremine Medical were then used
to predict the functions of RNASET2 and GGNBP2 through both their
interaction networks and annotated biological processes.
MicroRNAs (miRNAs) are a class of small (22 bp)
endogenous non-coding RNAs that regulate gene expression and thus
many cellular processes, both pathological and physiological, by
binding to the 3′-UTR of the target mRNA, causing mRNA cleavage,
destabilization or translational repression (92–95).
This method of post-transcriptional gene regulation has been shown
to play a crucial role in drug resistance in many types of cancers,
including ovarian cancer (7).
However, the specificity of miRNAs for their target genes also
makes them of great interest in diagnostics, prognosis and therapy
(96) and in predicting gene
function. miRWalk is a comprehensive database on miRNA-mRNA
interactions that gathers predicted and validated miRNA binding
sites on all mRNAs, mitochondrial genes, and the 10-kb upstream
flanking regions of all known genes in humans, mice and rats. More
importantly, it is a real-time database in which the ‘Validated
Target’ module is updated every month and the ‘Predicted Target’
module every 6 months (27). As a
result, the functional prediction of genes as determined by their
miRNAs through miRWalk is both feasible and reliable.
In the present study, RNASET2 and
GGNBP2 were shown to be notably and consistently
downregulated in drug-resistant ovarian cancers and ovarian cancer
cells, which suggested the involvement of these TSGs in drug
resistance. Support for this hypothesis came from a network
analysis based on gene/protein-gene/protein interaction networks
and biological processes annotation. The former indicated that
RNASET2 and GGNBP2 directly interact with many genes/proteins
(Fig. 2), all of which are
associated with drug resistance in ovarian cancer. Similarly, the
annotated biological processes analysis related RNASET2 and GGNBP2
mainly to processes closely linked to drug resistance (Fig. 3). In addition, determinations of
miRNA-mRNA interaction showed that among the eight miRNAs most
strongly targeting RNASET2 and GGNBP2 (Table I), the majority were involved in the
regulation of drug resistance in ovarian and other cancers,
suggesting roles for the two target genes in drug resistance.
Further support of an association between RNASET2 and GGNBP2 and
drug resistance came from a bioinformatics analysis of the drug
resistance-related pathways in which their co-expressed genes were
involved (Table II).
Currently, there are no data on the genetic pathways
related to RNASET2 and GGNBP2, either in the literature or in the
KEGG database. Instead, in the present study, examination of the
pathways enriched in the expression of genes co-expressed with
RNASET2 and GGNBP2 in drug-resistant ovarian cancer
cells suggested that the downregulation of the former resulted in
drug resistance through the activation of the PI3K-Akt signaling
pathway, whereas for the latter, cytokine-cytokine receptor
interactions, chemokine signaling pathways and NF-κB signaling were
implicated.
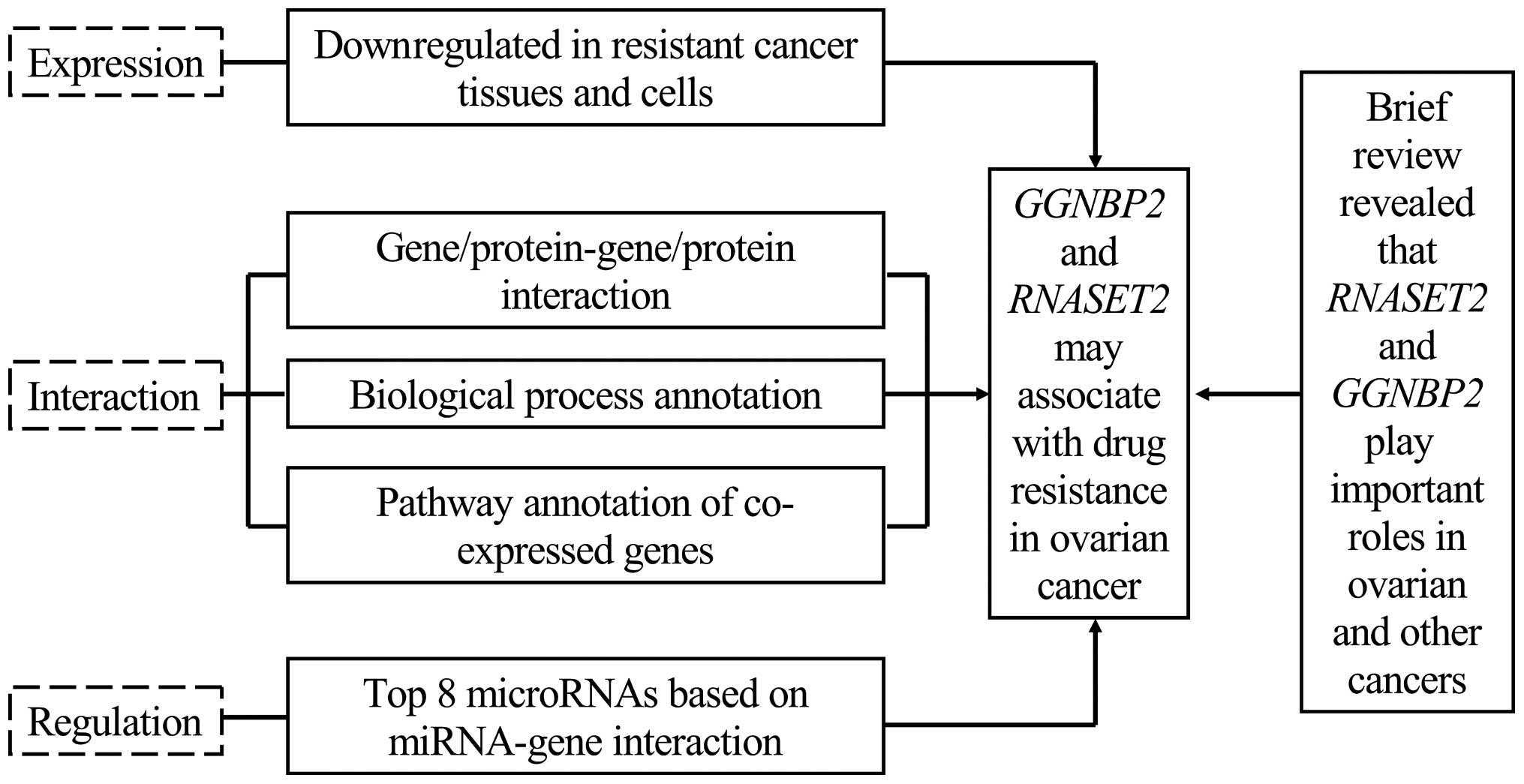
Collectively, on the basis of our studies of gene
expression at the transcriptional level, protein interactions,
annotated biological processes, miRNA-gene interactions and pathway
analysis (Fig. 4), we were able to
attribute the downregulation of RNASET2 and GGNBP2 in
drug-resistant ovarian cancers and ovarian tumor cells to drug
resistance in ovarian cancer. The present study sets the stage for
further experimental investigations into the associations between
RNASET2 and GGNBP2 and drug resistance.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (grant no. 81302283), and the Youth
Science Foundation of Guangxi Medical University (no.
GXMUYSF201205).
References
|
1
|
Balch C, Huang TH, Brown R and Nephew KP:
The epigenetics of ovarian cancer drug resistance and
resensitization. Am J Obstet Gynecol. 191:1552–1572. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2012. CA Cancer J Clin. 62:10–29. 2012. View Article : Google Scholar
|
|
3
|
Suh DH, Kim MK, No JH, Chung HH and Song
YS: Metabolic approaches to overcoming chemoresistance in ovarian
cancer. Ann N Y Acad Sci. 1229:53–60. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Matsuo K, Eno ML, Im DD, Rosenshein NB and
Sood AK: Clinical relevance of extent of extreme drug resistance in
epithelial ovarian carcinoma. Gynecol Oncol. 116:61–65. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gottesman MM: Mechanisms of cancer drug
resistance. Annu Rev Med. 53:615–627. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Johnson SW, Ozols RF and Hamilton TC:
Mechanisms of drug resistance in ovarian cancer. Cancer.
71:644–649. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sorrentino A, Liu CG, Addario A, Peschle
C, Scambia G and Ferlini C: Role of microRNAs in drug-resistant
ovarian cancer cells. Gynecol Oncol. 111:478–486. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sager R: Tumor suppressor genes: the
puzzle and the promise. Science. 246:1406–1412. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sherr CJ: Principles of tumor suppression.
Cell. 116:235–246. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yin F, Liu X, Li D, Wang Q, Zhang W and Li
L: Tumor suppressor genes associated with drug resistance in
ovarian cancer (Review). Oncol Rep. 30:3–10. 2013.PubMed/NCBI
|
|
11
|
Wu H, Cao Y, Weng D, Xing H, Song X, Zhou
J, Xu G, Lu Y, Wang S and Ma D: Effect of tumor suppressor gene
PTEN on the resistance to cisplatin in human ovarian cancer cell
lines and related mechanisms. Cancer Lett. 271:260–271. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Swisher EM, Sakai W, Karlan BY, Wurz K,
Urban N and Taniguchi T: Secondary BRCA1 mutations in
BRCA1-mutated ovarian carcinomas with platinum resistance.
Cancer Res. 68:2581–2586. 2008.PubMed/NCBI
|
|
13
|
Haber D and Harlow E: Tumour-suppressor
genes: evolving definitions in the genomic age. Nat Genet.
16:320–322. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jones PA and Laird PW: Cancer epigenetics
comes of age. Nat Genet. 21:163–167. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Acquati F, Morelli C, Cinquetti R, Bianchi
MG, Porrini D, Varesco L, Gismondi V, Rocchetti R, Talevi S,
Possati L, Magnanini C, Tibiletti MG, Bernasconi B, Daidone MG,
Shridhar V, Smith DI, Negrini M, Barbanti-Brodano G and Taramelli
R: Cloning and characterization of a senescence inducing and class
II tumor suppressor gene in ovarian carcinoma at chromosome region
6q27. Oncogene. 20:980–988. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Trubia M, Sessa L and Taramelli R:
Mammalian Rh/T2/S-glycoprotein ribonuclease family genes: cloning
of a human member located in a region of chromosome 6 (6q27)
frequently deleted in human malignancies. Genomics. 42:342–344.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
McClure BA, Haring V, Ebert PR, Anderson
MA, Simpson RJ, Sakiyama F and Clarke AE: Style
self-incompatibility gene products of Nicotlana alata are
ribonucleases. Nature. 342:955–957. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Steinemann D, Gesk S, Zhang Y, Harder L,
Pilarsky C, Hinzmann B, Martin-Subero JI, Calasanz MJ, Mungall A,
Rosenthal A, Siebert R and Schlegelberger B: Identification of
candidate tumor-suppressor genes in 6q27 by combined deletion
mapping and electronic expression profiling in lymphoid neoplasms.
Genes Chromosomes Cancer. 37:421–426. 2003. View Article : Google Scholar
|
|
19
|
Kim TY, Zhong S, Fields CR, Kim JH and
Robertson KD: Epigenomic profiling reveals novel and frequent
targets of aberrant DNA methylation-mediated silencing in malignant
glioma. Cancer Res. 66:7490–7501. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Monti L, Rodolfo M, Lo Russo G, Noonan D,
Acquati F and Taramelli R: RNASET2 as a tumor antagonizing
gene in a melanoma cancer model. Oncol Res. 17:69–74. 2008.
|
|
21
|
Acquati F, Possati L, Ferrante L,
Campomenosi P, Talevi S, Bardelli S, Margiotta C, Russo A,
Bortoletto E, Rocchetti R, Calza R, Cinquetti R, Monti L, Salis S,
Barbanti-Brodano G and Taramelli R: Tumor and metastasis
suppression by the human RNASET2 gene. Int J Oncol. 26:1159–1168.
2005.PubMed/NCBI
|
|
22
|
Acquati F, Bertilaccio S, Grimaldi A,
Monti L, Cinquetti R, Bonetti P, Lualdi M, Vidalino L, Fabbri M,
Sacco MG, van Rooijen N, Campomenosi P, Vigetti D, Passi A, Riva C,
Capella C, Sanvito F, Doglioni C, Gribaldo L, Macchi P, Sica A,
Noonan DM, Ghia P and Taramelli R: Microenvironmental control of
malignancy exerted by RNASET2, a widely conserved extracellular
RNase. Proc Natl Acad Sci USA. 108:1104–1109. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang X, Xiao Z, Chen Z, Li C, Li J,
Yanhui Y, Yang F, Yang Y and Oyang Y: Comparative proteomics
analysis of the proteins associated with laryngeal
carcinoma-related gene 1. Laryngoscope. 116:224–230. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li Y and Chen Z: Molecular cloning and
characterization of LCRG1 a novel gene localized to the
tumor suppressor locus D17S800-D17S930. Cancer Lett. 209:75–85.
2004.
|
|
25
|
Guan R, Wen XY, Wu J, Duan R, Cao H, Lam
S, Hou D, Wang Y, Hu J and Chen Z: Knockdown of ZNF403 inhibits
cell proliferation and induces G2/M arrest by modulating cell-cycle
mediators. Mol Cell Biochem. 365:211–222. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tang Z: Verification of differential
expression gene and its epigenic and genic study between
carbo-resistance cell line and its parental cell line of ovarian
carcinoma (unpublished PhD dissertation). Guangxi Medical
University. 2010.
|
|
27
|
Dweep H, Sticht C, Pandey P and Gretz N:
miRWalk - database: prediction of possible miRNA binding sites by
‘walking’ the genes of three genomes. J Biomed Inform. 44:839–847.
2011.
|
|
28
|
Huang da W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2008.PubMed/NCBI
|
|
29
|
Huang da W, Sherman BT and Lempicki RA:
Bioinformatics enrichment tools: paths toward the comprehensive
functional analysis of large gene lists. Nucleic Acids Res.
37:1–13. 2009.PubMed/NCBI
|
|
30
|
Zhang P, Gao W, Li H, Reed E and Chen F:
Inducible degradation of checkpoint kinase 2 links to
cisplatin-induced resistance in ovarian cancer cells. Biochem
Biophys Res Commun. 328:567–572. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Jansen AP, Camalier CE, Stark C and
Colburn NH: Characterization of programmed cell death 4 in multiple
human cancers reveals a novel enhancer of drug sensitivity. Mol
Cancer Ther. 3:103–110. 2004.PubMed/NCBI
|
|
32
|
Shiota M, Izumi H, Tanimoto A, Takahashi
M, Miyamoto N, Kashiwagi E, Kidani A, Hirano G, Masubuchi D,
Fukunaka Y, Yasuniwa Y, Naito S, Nishizawa S, Sasaguri Y and Kohno
K: Programmed cell death protein 4 down-regulates Y-box binding
protein-1 expression via a direct interaction with Twist1 to
suppress cancer cell growth. Cancer Res. 69:3148–3156. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang X, Wang X, Song X, Liu C, Shi Y,
Wang Y, Afonja O, Ma C, Chen YH and Zhang L: Programmed cell death
4 enhances chemosensitivity of ovarian cancer cells by activating
death receptor pathway in vitro and in vivo. Cancer Sci.
101:2163–2170. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chetram MA and Hinton CV: PTEN regulation
of ERK1/2 signaling in cancer. J Recept Signal Transduct Res.
32:190–195. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Steelman LS, Chappell WH, Abrams SL, Kempf
RC, Long J, Laidler P, Mijatovic S, Maksimovic-Ivanic D, Stivala F,
Mazzarino MC, Donia M, Fagone P, Malaponte G, Nicoletti F, Libra M,
Milella M, Tafuri A, Bonati A, Bäsecke J, Cocco L, Evangelisti C,
Martelli AM, Montalto G, Cervello M and McCubrey JA: Roles of the
Raf/MEK/ERK and PI3K/PTEN/Akt/mTOR pathways in controlling growth
and sensitivity to therapy-implications for cancer and aging.
Aging. 3:192–222. 2011.PubMed/NCBI
|
|
36
|
Chalhoub N and Baker SJ: PTEN and the
PI3-kinase pathway in cancer. Annu Rev Pathol. 4:127–150. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Okumura N, Yoshida H, Kitagishi Y,
Nishimura Y and Matsuda S: Alternative splicings on p53, BRCA1 and
PTEN genes involved in breast cancer. Biochem Biophys Res Commun.
413:395–399. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Selvendiran K, Tong L, Vishwanath S,
Bratasz A, Trigg NJ, Kutala VK, Hideg K and Kuppusamy P: EF24
induces G2/M arrest and apoptosis in cisplatin-resistant human
ovarian cancer cells by increasing PTEN expression. J Biol Chem.
282:28609–28618. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lee S, Choi EJ, Jin C and Kim DH:
Activation of PI3K/Akt pathway by PTEN reduction and PIK3CA
mRNA amplification contributes to cisplatin resistance in an
ovarian cancer cell line. Gynecol Oncol. 97:26–34. 2005.PubMed/NCBI
|
|
40
|
Yuan M, Tomlinson V, Lara R, Holliday D,
Chelala C, Harada T, Gangeswaran R, Manson-Bishop C, Smith P,
Danovi SA, Pardo O, Crook T, Mein CA, Lemoine NR, Jones LJ and Basu
S: Yes-associated protein (YAP) functions as a tumor suppressor in
breast. Cell Death Differ. 15:1752–1759. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Li J, Zou C, Bai Y, Wazer DE, Band V and
Gao Q: DSS1 is required for the stability of BRCA2. Oncogene.
25:1186–1194. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Sakai W, Swisher EM, Jacquemont C,
Chandramohan KV, Couch FJ, Langdon SP, Wurz K, Higgins J, Villegas
E and Taniguchi T: Functional restoration of BRCA2 protein by
secondary BRCA2 mutations in BRCA2-mutated ovarian
carcinoma. Cancer Res. 69:6381–6386. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Vitari AC, Leong KG, Newton K, Yee C,
O’Rourke K, Liu J, Phu L, Vij R, Ferrando R, Couto SS, Mohan S,
Pandita A, Hongo JA, Arnott D, Wertz IE, Gao WQ, French DM and
Dixit VM: COP1 is a tumour suppressor that causes degradation of
ETS transcription factors. Nature. 474:403–406. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Migliorini D, Bogaerts S, Defever D, Vyas
R, Denecker G, Radaelli E, Zwolinska A, Depaepe V, Hochepied T,
Skarnes WC and Marine JC: Cop1 constitutively regulates c-Jun
protein stability and functions as a tumor suppressor in mice. J
Clin Invest. 121:1329–1343. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Li YF, Wang DD, Zhao BW, Wang W, Huang CY,
Chen YM, Zheng Y, Keshari RP, Xia JC and Zhou ZW: High level of
COP1 expression is associated with poor prognosis in primary
gastric cancer. Int J Biol Sci. 8:1168–1177. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Liang J, Song W, Tromp G, Kolattukudy PE
and Fu M: Genome-wide survey and expression profiling of CCCH-zinc
finger family reveals a functional module in macrophage activation.
PLoS One. 3:e28802008. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Shih JY and Yang PC: The EMT regulator
slug and lung carcinogenesis. Carcinogenesis. 32:1299–1304. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Paige AJ, Taylor KJ, Taylor C, Hillier SG,
Farrington S, Scott D, Porteous DJ, Smyth JF, Gabra H and Watson
JE: WWOX: a candidate tumor suppressor gene involved in
multiple tumor types. Proc Natl Acad Sci USA. 98:11417–11422. 2001.
View Article : Google Scholar
|
|
49
|
Chiang MF, Yeh ST, Liao HF, Chang NS and
Chen YJ: Overexpression of WW domain-containing oxidoreductase WOX1
preferentially induces apoptosis in human glioblastoma cells
harboring mutant p53. Biomed Pharmacother. 66:433–438. 2012.
View Article : Google Scholar
|
|
50
|
Liu YY, Li L, Li DR, Zhang W and Wang Q:
Suppression of WWOX gene by RNA interference reverses platinum
resistance acquired in SKOV3/SB cells. Zhonghua Fu Chan Ke Za Zhi.
43:854–858. 2008.(In Chinese).
|
|
51
|
de Leeuw N1, Dijkhuizen T, Hehir-Kwa JY,
Carter NP, Feuk L, Firth HV, Kuhn RM, Ledbetter DH, Martin CL, van
Ravenswaaij-Arts CM, Scherer SW, Shams S, Van Vooren S, Sijmons R,
Swertz M and Hastings R: Diagnostic interpretation of array data
using public databases and internet sources. Hum Mutat. 33:930–940.
2012.PubMed/NCBI
|
|
52
|
Moitra K, Im K, Limpert K, Borsa A,
Sawitzke J, Robey R, Yuhki N, Savan R, Huang da W, Lempicki RA,
Bates S and Dean M: Differential gene and microRNA expression
between etoposide resistant and etoposide sensitive MCF7 breast
cancer cell lines. PLoS One. 7:e452682012. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Boren T, Xiong Y, Hakam A, Wenham R, Apte
S, Chan G, Kamath SG, Chen DT, Dressman H and Lancaster JM:
MicroRNAs and their target messenger RNAs associated with ovarian
cancer response to chemotherapy. Gynecol Oncol. 113:249–255. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Ko MA, Zehong G, Virtanen C, Guindi M,
Waddell TK, Keshavjee S and Darling GE: MicroRNA expression
profiling of esophageal cancer before and after induction
chemoradiotherapy. Ann Thorac Surg. 94:1094–1103. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Eitan R, Kushnir M, Lithwick-Yanai G,
David MB, Hoshen M, Glezerman M, Hod M, Sabah G, Rosenwald S and
Levavi H: Tumor microRNA expression patterns associated with
resistance to platinum based chemotherapy and survival in ovarian
cancer patients. Gynecol Oncol. 114:253–259. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Jung EJ, Santarpia L, Kim J, Esteva FJ,
Moretti E, Buzdar AU, Di Leo A, Le XF, Bast RC Jr, Park ST, Pusztai
L and Calin GA: Plasma microRNA 210 levels correlate with
sensitivity to trastuzumab and tumor presence in breast cancer
patients. Cancer. 118:2603–2614. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Guo L, Liu Y, Bai Y, Sun Y, Xiao F and Guo
Y: Gene expression profiling of drug-resistant small cell lung
cancer cells by combining microRNA and cDNA expression analysis.
Eur J Cancer. 46:1692–1702. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Wu XM, Shao XQ, Meng XX, Zhang XN, Zhu L,
Liu SX, Lin J and Xiao HS: Genome-wide analysis of microRNA and
mRNA expression signatures in hydroxycamptothecin-resistant gastric
cancer cells. Acta Pharmacol Sin. 32:259–269. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Sun JY, Huang Y, Li JP, Zhang X, Wang L,
Meng YL, Yan B, Bian YQ, Zhao J, Wang WZ, Yang AG and Zhang R:
MicroRNA-320a suppresses human colon cancer cell proliferation by
directly targeting β-catenin. Biochem Biophys Res Commun.
420:787–792. 2012.PubMed/NCBI
|
|
60
|
Zhang Y, He X, Liu Y, Ye Y, Zhang H, He P,
Zhang Q, Dong L, Liu Y and Dong J: microRNA-320a inhibits tumor
invasion by targeting neuropilin 1 and is associated with liver
metastasis in colorectal cancer. Oncol Rep. 27:685–694. 2012.
|
|
61
|
Zhang H, Li W, Nan F, Ren F, Wang H, Xu Y
and Zhang F: MicroRNA expression profile of colon cancer stem-like
cells in HT29 adenocarcinoma cell line. Biochem Biophys Res Commun.
404:273–278. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Yang N, Kaur S, Volinia S, Greshock J,
Lassus H, Hasegawa K, Liang S, Leminen A, Deng S, Smith L,
Johnstone CN, Chen XM, Liu CG, Huang Q, Katsaros D, Calin GA, Weber
BL, Bützow R, Croce CM, Coukos G and Zhang L: MicroRNA microarray
identifies Let-7i as a novel biomarker and therapeutic
target in human epithelial ovarian cancer. Cancer Res.
68:10307–10314. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Hisaoka M, Matsuyama A and Nakamoto M:
Aberrant calreticulin expression is involved in the
dedifferentiation of dedifferentiated liposarcoma. Am J Pathol.
180:2076–2083. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Li M, Balch C, Montgomery JS, Jeong M,
Chung JH, Yan P, Huang TH, Kim S and Nephew KP: Integrated analysis
of DNA methylation and gene expression reveals specific signaling
pathways associated with platinum resistance in ovarian cancer. BMC
Med Genomics. 2:342009. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Wang Y, Niu XL, Qu Y, Wu J, Zhu YQ, Sun WJ
and Li LZ: Autocrine production of interleukin-6 confers cisplatin
and paclitaxel resistance in ovarian cancer cells. Cancer Lett.
295:110–123. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Wang Y, Qu Y, Niu XL, Sun WJ, Zhang XL and
Li LZ: Autocrine production of interleukin-8 confers cisplatin and
paclitaxel resistance in ovarian cancer cells. Cytokine.
56:365–375. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Lian L, Qu LJ, Sun HY, Chen YM, Lamont SJ,
Liu CJ and Yang N: Gene expression analysis of host spleen
responses to Marek’s disease virus infection at late tumor
transformation phase. Poult Sci. 91:2130–2138. 2012.
|
|
68
|
Chang MY, Yu YP, Tsai JR, Sheu CC, Chong
IW and Lin SR: Combined oligonucleotide microarray-bioinformatics
and constructed membrane arrays to analyze the biological pathways
in the carcinogenesis of human lung adenocarcinoma. Oncol Rep.
18:569–579. 2007.
|
|
69
|
Morales M, Planet E, Arnal-Estape A,
Pavlovic M, Tarragona M and Gomis RR: Tumor-stroma interactions a
trademark for metastasis. Breast. 20(Suppl 3): S50–S55. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Che CL, Zhang YM, Zhang HH, Sang YL, Lu B,
Dong FS, Zhang LJ and Lv FZ: DNA microarray reveals different
pathways responding to paclitaxel and docetaxel in non-small cell
lung cancer cell line. Int J Clin Exp Pathol. 6:1538–1548.
2013.PubMed/NCBI
|
|
71
|
Wang JM, Wu JT, Sun DK, Zhang P and Wang
L: Pathway crosstalk analysis based on protein-protein network
analysis in prostate cancer. Eur Rev Med Pharmacol Sci.
16:1235–1242. 2012.PubMed/NCBI
|
|
72
|
Wilson C, Purcell C, Seaton A, Oladipo O,
Maxwell PJ, O’Sullivan JM, Wilson RH, Johnston PG and Waugh DJ:
Chemotherapy-induced CXC-chemokine/CXC-chemokine receptor signaling
in metastatic prostate cancer cells confers resistance to
oxaliplatin through potentiation of nuclear factor-κB transcription
and evasion of apoptosis. J Pharmacol Exp Ther. 327:746–759.
2008.PubMed/NCBI
|
|
73
|
He D, Xu Q, Yan M, Zhang P, Zhou X, Zhang
Z, Duan W, Zhong L, Ye D and Chen W: The NF-kappaB inhibitor,
celastrol, could enhance the anti-cancer effect of gambogic acid on
oral squamous cell carcinoma. BMC Cancer. 9:3432009. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Choi BH, Kim CG, Lim Y, Shin SY and Lee
YH: Curcumin down-regulates the multidrug-resistance mdr1b
gene by inhibiting the PI3K/Akt/NFκB pathway. Cancer Lett.
259:111–118. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Godwin P, Baird AM, Heavey S, Barr MP,
O’Byrne KJ and Gately K: Targeting nuclear factor-kappa B to
overcome resistance to chemotherapy. Front Oncol. 3:1202013.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Yang G, Xiao X, Rosen DG, Cheng X, Wu X,
Chang B, Liu G, Xue F, Mercado-Uribe I, Chiao P, Du X and Liu J:
The biphasic role of NF-κB in progression and chemoresistance of
ovarian cancer. Clin Cancer Res. 17:2181–2194. 2011.
|
|
77
|
Lee SC, Xu X, Lim YW, Iau P, Sukri N, Lim
SE, Yap HL, Yeo WL, Tan P, Tan SH, McLeod H and Goh BC:
Chemotherapy-induced tumor gene expression changes in human breast
cancers. Pharmacogenet Genomics. 19:181–192. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Arafa el-SA, Zhu Q, Barakat BM, Wani G,
Zhao Q, El-Mahdy MA and Wani AA: Tangeretin sensitizes
cisplatin-resistant human ovarian cancer cells through
downregulation of phosphoinositide 3-kinase/Akt signaling pathway.
Cancer Res. 69:8910–8917. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Li QQ, Yunmbam MK, Zhong X, Yu JJ,
Mimnaugh EG, Neckers L and Reed E: Lactacystin enhances cisplatin
sensitivity in resistant human ovarian cancer cell lines via
inhibition of DNA repair and ERCC-1 expression. Cell Mol Biol.
47:OL61–OL72. 2001.PubMed/NCBI
|
|
80
|
Menendez JA, Vellon L, Mehmi I, Teng PK,
Griggs DW and Lupu R: A novel CYR61-triggered
‘CYR61-αvβ3 integrin loop’
regulates breast cancer cell survival and chemosensitivity through
activation of ERK1/ERK2 MAPK signaling pathway. Oncogene.
24:761–779. 2005.PubMed/NCBI
|
|
81
|
Galan-Moya EM, de la Cruz-Morcillo MA,
Llanos Valero M, Callejas-Valera JL, Melgar-Rojas P, Hernadez Losa
J, Salcedo M, Fernández-Aramburo A, Ramon y Cajal S and
Sánchez-Prieto R: Balance between MKK6 and MKK3 mediates p38 MAPK
associated resistance to cisplatin in NSCLC. PLoS One.
6:e284062011. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Zhang J, Wang Y, Zhen P, Luo X, Zhang C,
Zhou L, Lu Y, Yang Y, Zhang W and Wan J: Genome-wide analysis of
miRNA signature differentially expressed in doxorubicin-resistant
and parental human hepatocellular carcinoma cell lines. PLoS One.
8:e541112013. View Article : Google Scholar
|
|
83
|
Lange TS, Stuckey AR, Robison K, Kim KK,
Singh RK, Raker CA and Brard L: Effect of a vitamin D3
derivative (B3CD) with postulated anti-cancer activity in an
ovarian cancer animal model. Invest New Drugs. 28:543–553.
2010.PubMed/NCBI
|
|
84
|
Zhang P, Ling G, Pan X, Sun J, Zhang T, Pu
X, Yin S and He Z: Novel nanostructured lipid-dextran sulfate
hybrid carriers overcome tumor multidrug resistance of mitoxantrone
hydrochloride. Nanomedicine. 8:185–193. 2012. View Article : Google Scholar
|
|
85
|
Calzolari A, Papucci A, Baroni G, Ficarra
G, Porfirio B, Chiarelli I and Di Lollo S: Epstein-Barr virus
infection and P53 expression in HIV-related oral large B cell
lymphoma. Head Neck. 21:454–460. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Longley DB and Johnston PG: Molecular
mechanisms of drug resistance. J Pathol. 205:275–292. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Yin F, Liu X, Li D, Wang Q, Zhang W and Li
L: Bioinformatic analysis of chemokine (C-C motif) ligand 21 and
SPARC-like protein 1 revealing their associations with drug
resistance in ovarian cancer. Int J Oncol. 42:1305–1316.
2013.PubMed/NCBI
|
|
88
|
Kong KA, Yoon H and Kim MH: Akt1 as a
putative regulator of Hox genes. Gene. 513:287–291. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Sharan R, Ulitsky I and Shamir R:
Network-based prediction of protein function. Mol Syst Biol.
3:882007. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Mostafavi S, Ray D, Warde-Farley D,
Grouios C and Morris Q: GeneMANIA: a real-time multiple association
network integration algorithm for predicting gene function. Genome
Biol. 9(Suppl 1): S42008. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Jenssen TK, Laegreid A, Komorowski J and
Hovig E: A literature network of human genes for high-throughput
analysis of gene expression. Nat Genet. 28:21–28. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Behm-Ansmant I, Rehwinkel J and Izaurralde
E: MicroRNAs silence gene expression by repressing protein
expression and/or by promoting mRNA decay. Cold Spring Harb Symp
Quant Biol. 71:523–530. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Bartel DP: MicroRNAs: genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Kloosterman WP and Plasterk RH: The
diverse functions of microRNAs in animal development and disease.
Dev Cell. 11:441–450. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Croce CM and Calin GA: miRNAs, cancer, and
stem cell division. Cell. 122:6–7. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Tili E, Michaille JJ, Gandhi V, Plunkett
W, Sampath D and Calin GA: miRNAs and their potential for use
against cancer and other diseases. Future Oncol. 3:521–537. 2007.
View Article : Google Scholar : PubMed/NCBI
|