Introduction
Acute myelogenous leukemia (AML) is a malignant
neoplastic disease caused by the abnormal proliferation of
hematopoietic stem cells. Although treatments for AML have made
considerable progress in recent years, more than 20% of patients
still cannot be completely cured; they eventually relapse and
succumb to the disease. It is known that cancerous cells undergo
indefinite cell proliferation caused by disorders of signal
transduction pathways (1). The
success of imatinib and other drugs in the treatment of chronic
myelogenous leukemia has demonstrated that targeted therapy of
leukemia is an important therapeutic development.
Oleanolic acid (OA) is a pentacyclic triterpenoid;
the molecular formula is C30H48O3.
OA was first separated from the leaves of a clover plant, Olea
europeae L, in 1908. Since the identification of OA as an
effective anti-hepatitis monomer, new biological properties have
been discovered, including blood sugar and blood lipid lowering
abilities, as well as anti-bacterial, anti-viral, anti-caries
(2), immunoregulatory (3), anti-oxidative, anti-mutation,
anti-cardiovascular and cerebrovascular disease (4), scar formation inhibitory (5) and antitumor effects. In recent years,
the antitumor effects of OA have attracted increased attention. OA
shows an inhibitory effect on solid tumors including skin cancer,
lung cancer, breast cancer and osteosarcomas. The mechanisms of the
antitumor effects of OA are complicated; it is involved in the
regulation of a variety of signal transduction pathways. ERK-P53
(6), mTOR (7), Bcl-2 (8,9), VEGF
(10), NO and TNF-α (11) can all participate in the anticancer
activity of OA. Our previous study found that inhibition of
proliferation and the promotion of apoptosis in OA-treated HL-60
cells were accompanied by increased caveolin-1 (CAV-1) expression
and inhibition of the PI3K/AKT signal transduction pathway.
The human cav-1 gene is located on chromosome
7q31.1. The molecular weight of CAV-1 is 22 kDa. CAV-1 binds to a
variety of signaling proteins and inhibits their activities;
therefore, it is considered a ‘broad spectrum’ kinase inhibitor
located at the center of various signaling pathways (12). Cav-1 is expressed in a variety of
tumor cells with significantly different expression patterns and
dual carcinogenic and tumor-suppressor functions. CAV-1 has a
variety of biological effects, including affecting tumor multidrug
resistance (13,14), regulating the cell cycle (15) and signal transduction, participating
in the regulation of aging (16,17),
enhancing cell autophagy function (18,19)
and participating in cell proliferation, apoptosis, migration and
differentiation processes. In leukemia patients, CAV-1 and MDR-1
expression levels are positively correlated (20), and external stimulating factors such
as HTLV can induce CAV-1 overexpression in T-cell lines (21). In untreated AML patients, CAV-1
expression levels were found to be significantly lower than levels
in the normal control group; the levels increased after remission
from induction chemotherapy and decreased again during relapse.
Thus, cav-1 has high research value for the treatment of
leukemia.
The goal of this study was to investigate the roles
of cav-1 in HL-60 cells and to explore the mechanism of OA effects.
The results showed that OA not only promoted CAV-1 expression in
HL-60 cells but also increased protein phosphorylation. To further
study whether OA affects HL-60 cells by activating downstream
signaling pathways via CAV-1, the lentiviral overexpression vector
pcDNA-CAV-1 and the shRNA expression vector PSIH1-H1-CAV-1 were
constructed to overexpress and silence cav-1 in HL-60 cells,
respectively. The cav-1-mediated cell proliferation and apoptosis
and the PI3K/AKT pathway were examined; OA efficacy was also
determined in HL-60 cells overexpressing cav-1. At same time, the
role of OA after cav-1 silencing was determined. The results showed
that OA, cav-1 overexpression or their combination in HL-60 cells
inhibited proliferation, promoted apoptosis, arrested the cell
cycle in the G1 phase, downregulated P110α, p-AKT (Thr308), p-AKT
(Ser473) and mTOR and upregulated p-CAV-1 levels. The effects of
the joint treatment were more significant. After cav-1 silencing,
OA showed no effect on HL-60 cell proliferation, apoptosis, cell
cycle and levels of P110α, p-AKT (Thr308), p-AKT (Ser473), mTOR and
p-CAV-1.
Materials and methods
Cell culture and detection of cav-1
expression in various cell lines
The human leukemia cell lines SUP-B15, HL-60, THP-1
and K562 were purchased from the Chinese Academy of Sciences Cell
Library. The cells were cultured in RPMI-1640 culture medium
containing 10% fetal bovine serum at 37°C and 5% CO2.
The cells were passaged every 2–3 days. The cells were collected at
the logarithmic growth phase. CLL primary tumor cells (CLP) and
white blood cells (WBCs) were collected through centrifugation
directly after separation. Expression of cav-1 in each cell line
was detected using qRT-PCR and western blot assays. Primers are
listed in Table I.
 | Table IPrimer sequences and annealing
temperatures for quantitative RT-PCR. |
Table I
Primer sequences and annealing
temperatures for quantitative RT-PCR.
| Gene name
(accession no.) | Primer name | Primer sequence
(5′-3′) | Temperature
(°C) | Product size
(bp) |
|---|
| β-actin
(NM_001101) | Forward primer |
CCTGTACGCCAACACAGTGC | 60 | 211 |
| Reverse primer |
ATACTCCTGCTTGCTGATCC | | |
| cav-1
(NM_001753.4) | Forward primer |
CGGGAACAGGGCAACATGTACA | 58 | 178 |
| Reverse primer |
TCCCTTCTGGTTCTGCAATCACAT | | |
Construction of recombinant lentiviral
expression vector pcDNA-CAV-1 and pSIH1-H1-CAV-1
First, total-RNA was extracted from freshly prepped
human leukocytes, and cav-1 cDNA with XbaI and BamHI
restriction sites at both ends was amplified using PCR. The
digested fragment was then inserted into pcDNA-EF1-GFP between the
corresponding restriction sites to construct the recombinant
plasmid pcDNA-CAV-1.
Second, three siRNA sequences targeting the cav-1
(NM_001753.4) CDS region were designed and synthesized by Shanghai
Invitrogen. The synthetic sequences were inserted into pSIH1-H1-GFP
shRNA vectors to construct the recombinant pSIH1-CAV-1-shRNA
plasmids (pSIH1-CAV-1-shRNA1, pSIH1-CAV-1-shRNA2,
pSIH1-CAV-1-shRNA3, pSIH1-CAV-1-Negative).
The positive clones of the above recombinant
plasmids pcDNA-CAV-1 and pSIH1-CAV-1-shRNA were screened and
sequenced, and the correct recombinant expression vector plasmid
DNA was purified to eliminate the endotoxin. The selected 293 cells
were transfected and divided into 7 groups: the blank control
(293), the pcDNA empty vector-transfected cells (293-P), the
pcDNA-CAV-1-transfected cells (293-PC) and four pcDNA-CAV-1 of
pSIH1-CAV-1-shRNA-transfected cells (293-PC-S1, 293-PC-S2,
293-PC-S3, 293-PC-N). Lipofectamine 2000 was used as the
transfection reagent. qRT-PCR and western blot assays were used to
detect cav-1 expression levels and to screen effective siRNA
sequences.
Recombinant lentiviral infection of HL-60
cells
The third generation of the lentiviral vector was
used for viral packaging and production following the
manufacturer’s instructions. Briefly, 293TN cells were transfected
with a mixture of lentiviral vector (pcDNA-CAV-1 or
pSIH1-CAV-1-shRNA) and packaging plasmid, and the supernatant was
collected and filtered using a 0.45 μmol/l PVDF membrane. Viral
titers were determined through gradient centrifugation. After the
high-titer viral preparation met the requirements, the host HL-60
cells were infected at an optimal MOI. CAV-1 expression levels in
the infected HL-60 cells were detected through western blot assays.
The GFP reporter gene was present in both vectors. Transfection
efficiency was easily determined, as green fluorescence was a
necessary prerequisite for a high efficiency infection of HL-60
cells.
OA treatment and grouping
HL-60 cell suspensions in the logarithmic growth
phase before and after transfection were prepared and dispensed in
96-well plates, with a 100 μl cell suspension containing
1×104 cells per well. The HL-60 cells were divided into
10 groups: the control group (HL-60), the OA-treated HL-60 group
(HL-60+OA), the empty pcDNA-transfected cell group (HL-60-L), the
cav-1-overexpressing cell group (HL-60-LC), the OA-treated HL-60-L
group (HL-60-L+OA), the OA-treated HL-60-LC group (HL-60-LC+OA),
the pSIH1-CAV-1-shRNA3-transfected group (HL-60-LS3), the
pSIH1-CAV-1-negative-transfected cell group (HL-60-LN), the
OA-treated HL-60-LN group (HL-60-LN+OA) and the OA-treated
HL-60-LS3 group (HL-60-LS3+OA). The final OA concentration for cell
treatment was 80 μM/l.
Western blot analyses
The cells were collected and lysed for protein
extraction. Protein concentrations were determined using the BCA
technique, and the cell lysates were subjected to SDS-PAGE
electrophoresis. After separation, the proteins were transferred to
PVDF membranes; the membranes were blocked using TBST containing 5%
non-fat dry milk. Then, the anti-human antibodies anti-CAV-1,
P110α, p-AKT (Thr308), p-AKT (Ser473), p-CAV-1 (Tyr14) and mTOR
were added. Horseradish peroxidase-conjugated anti-rabbit (or
mouse) IgG (Cell Signaling Technology) was used for detection of
immunoreactive proteins by chemiluminescence (Pierce ECL western
blotting substrate).
Cell proliferation activity analysis
Dispensed cells of each group were placed into
96-well plates with 1×104 cells per well; 10 μl of CCK-8
reagent was added per well at 24, 48 and 72 h. After culturing the
treated plates in an incubator for 4 h, the supernatants were
transferred to another 96-well plate, and the absorbance at 450 nm
was measured using a microplate reader.
Flow cytometric analysis
The cells of each group were plated in 12-well
plates, with ~1×105 cells per well. The cells were
incubated to 75–80% confluency per well. Cell apoptosis was
detected using a double staining Annexin V:FITC Apoptosis Detection
Kit II. The cells were harvested, washed with ice-cold PBS twice
and fixed with 75% cold ethanol at 4°C overnight. Propidium iodide
(PI) staining of nuclei was used to monitor the phases of the cell
cycle. The fluorescence of DNA-bound PI in cells was measured with
a FACSCalibur flow cytometer (BD Biosciences).
Statistical analyses
The data are expressed as the mean ± standard
deviation (SD) and were compared by one-way analysis of variance
(one-way ANOVA), Dunnet t-test and least significant
difference-test (LSD). Statistical analyses were performed via SPSS
statistical software 16.0; P<0.05 or P<0.01 was considered to
indicate a statistically significant difference.
Results
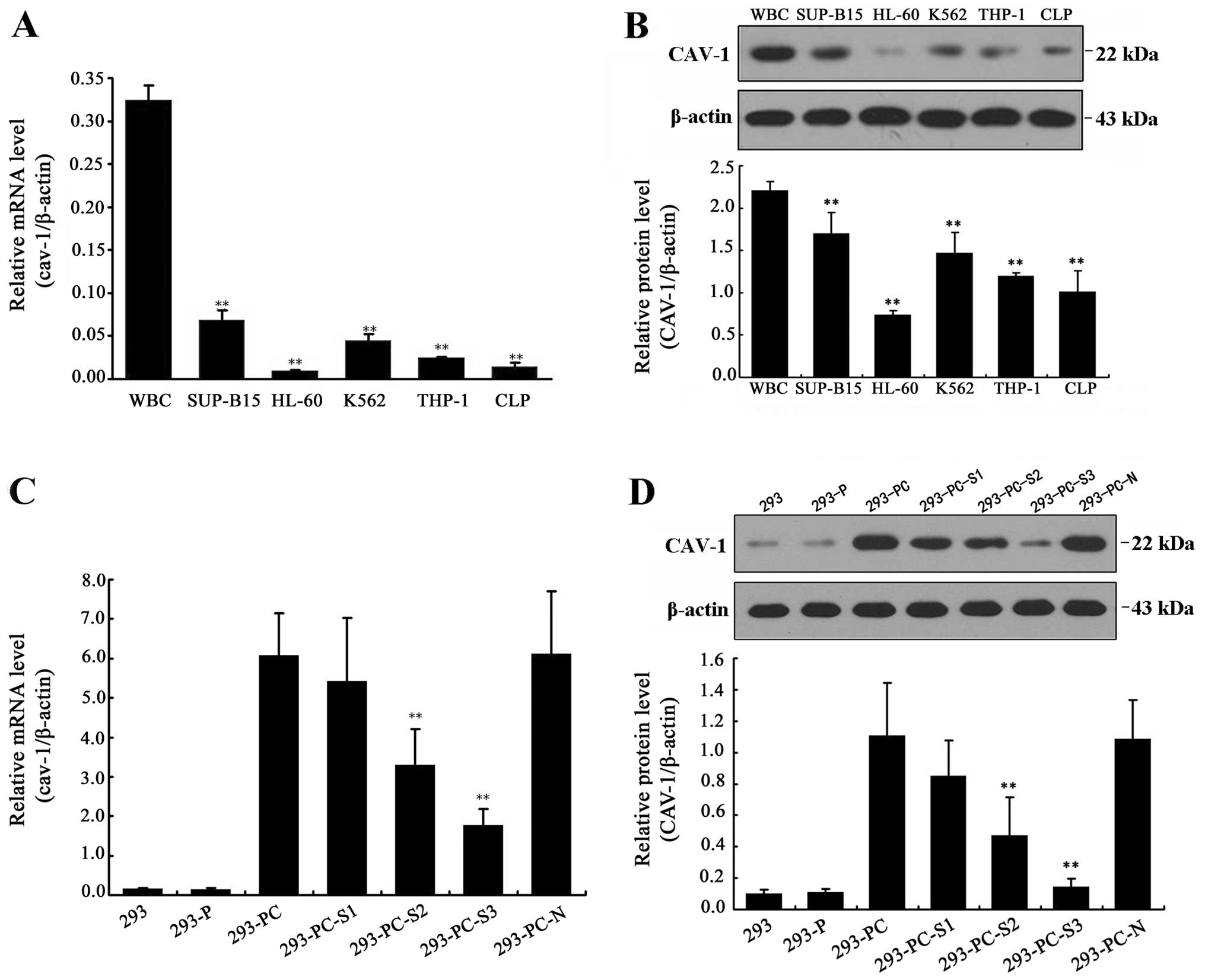
Cav-1 gene expression levels in various
leukemia cell lines
The qRT-PCR and western blotting results revealed
the cav-1 mRNA and protein expression levels in all tested cell
lines: WBC, SUP-B15, HL-60, K562, THP-1 and CLP. Compared with
WBCs, cav-1 expression levels were reduced in the other cell lines,
with the lowest cav-1 expression found in HL-60 cells (Fig. 1A and B). Thus, we chose the HL-60
cell line for subsequent experiments.
Construction of the recombinant vectors
pcDNA-CAV-1 and pSIH1-H1-shRNA-CAV-1
The positive pcDNA-EF1-GFP clones containing cav-1
and the PSIH1-shRNA plasmid containing cav-1 siRNA were screened
and sequenced. The results showed that the sequences were identical
to that of the recombinant plasmid. The two vectors were
successfully transfected into 293 cells using Lipo 2000™ and were
effectively expressed. qRT-PCR results showed that the mRNA
expression levels were significantly higher in the 293-PC cells
than in the control cells. The three designed siRNA sequences were
screened on the basis of cav-1 overexpression in the stably
transfected 293 cell line. The screening results indicated that the
293-PC-S3 cells showed the strongest inhibition of cav-1 mRNA and
protein expression levels among the tested cells (Fig. 1C and D). Therefore, we chose
siRNA-3, which showed the highest cav-1 gene silencing efficiency,
for subsequent experiments.
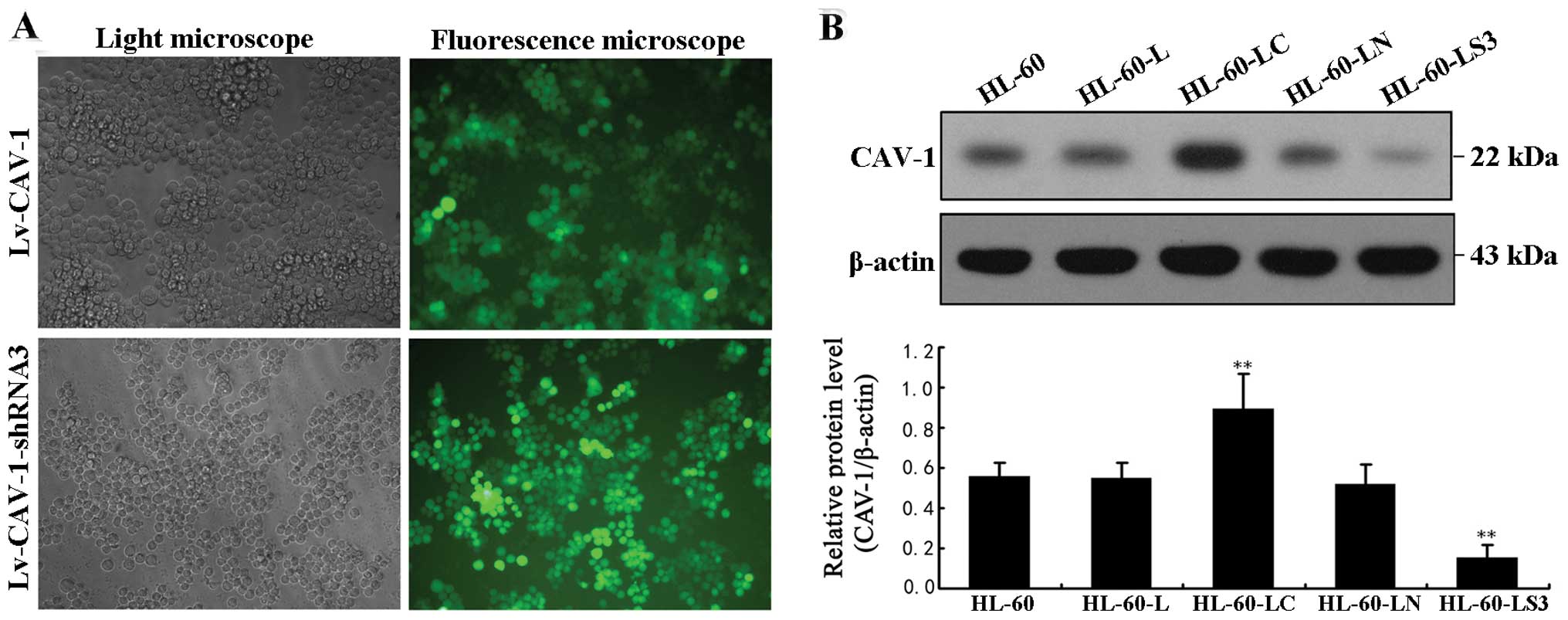
Lv-CAV-1 and Lv-CAV-1-shRNA3 successfully
infected HL-60 cells
HL-60 cells were observed using green fluorescence
microscopy 72 h after transfection. The transfection efficiencies
of Lv-CAV-1 and Lv-CAV-1-shRNA3 were 85 and 90%, respectively
(Fig. 2A). Western blotting results
96 h after transfection (Fig. 2B)
showed that CAV-1 protein levels were significantly increased in
the HL-60-LC cells and significantly reduced in the HL-60-LS3
cells.
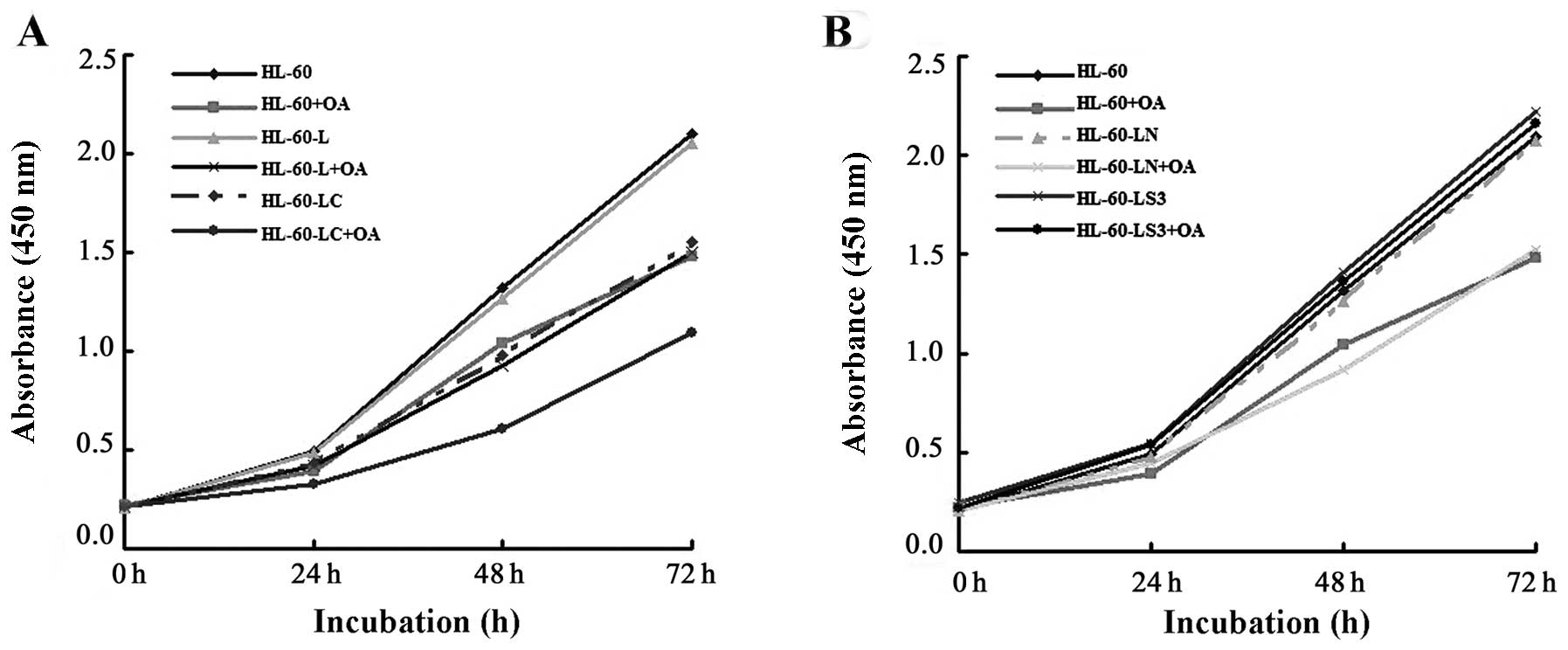
Cell proliferation
HL-60 cell activity was significantly decreased 72 h
after the addition of OA, with absorbance values of 2.10±0.22 vs.
1.48±0.12 (P<0.01). The activities of the HL-60-LC cells were
decreased at 48 and 72 h, with absorbance values of 1.32±0.12 vs.
0.98±0.14 (P<0.05) and 2.10±0.22 vs. 1.55±0.09 (P<0.01),
respectively. The proliferation of HL-60-LC+OA cells decreased in
the first 24 h, with absorbance value of 0.49±0.10 vs. 0.33±0.02
(P<0.05). Cell proliferation inhibition was more significant in
the HL-60-LC+OA cells than in the HL-60+OA and HL-60-LC cells.
These results indicated that both CAV-1 and OA inhibited HL-60 cell
proliferation, and the inhibitory effect was more significant in
the HL-60-LC+OA cells (Fig.
3A).
At 72 h after the addition of CCK-8, cell
proliferation inhibition was lower in the HL-60-LS3+OA cells than
in the HL-60+OA cells, with absorbance values of 2.16±0.27 vs.
1.48±0.12 (P<0.01). However, there was no difference in
proliferation between the HL-60-LS3+OA and HL-60 cells. These
results indicated that the lentiviral-mediated gene silencing of
cav-1 hampered OA-induced inhibition of HL-60 cell proliferation
(Fig. 3B).
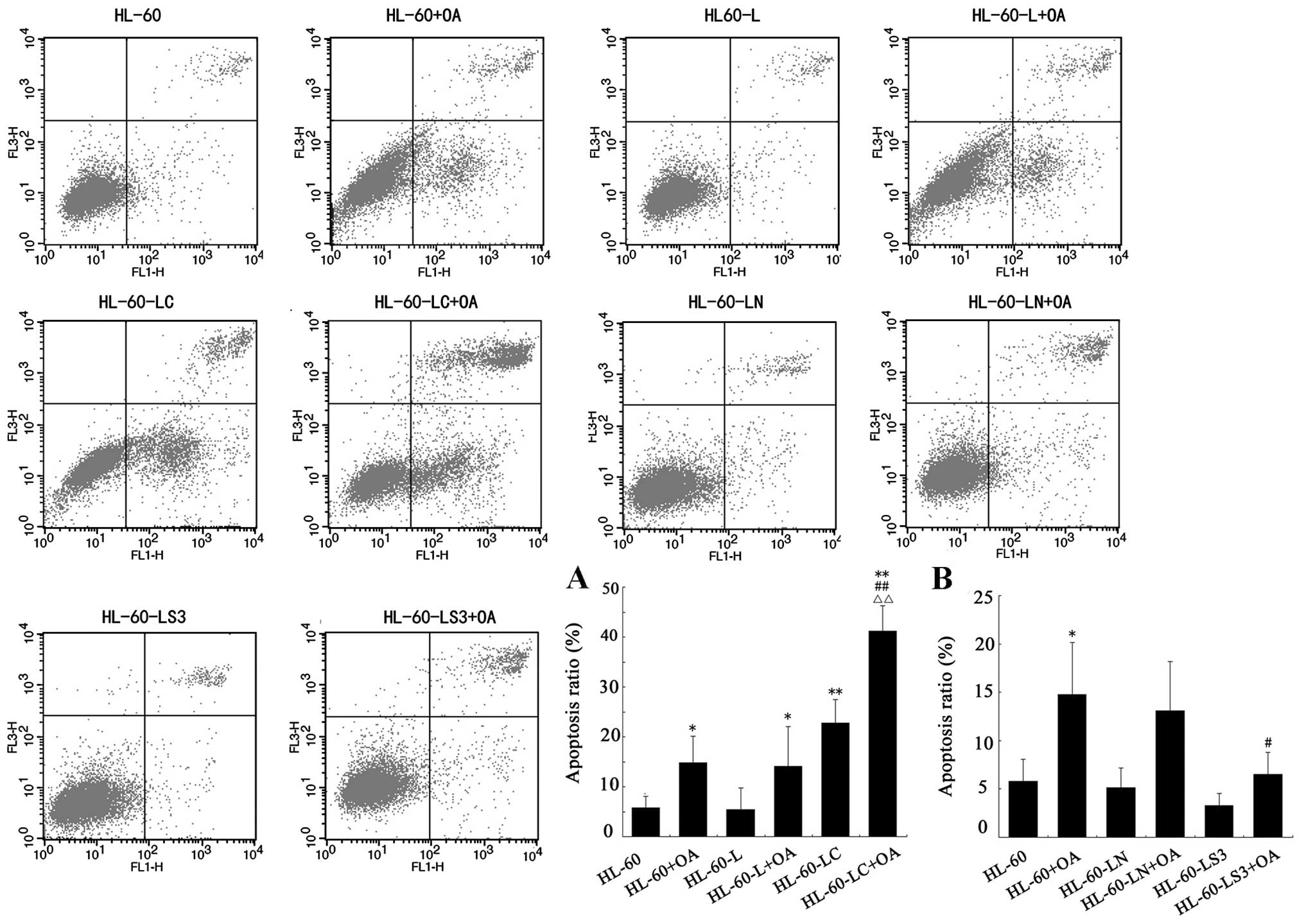
Detection of apoptosis
At 24 h after each treatment, the apoptosis levels
of each group were detected using flow cytometry (Fig. 4). The apoptotic rates of the HL-60,
HL-60+OA, HL-60-LC, HL-60-LC+OA and HL-60-LS3+OA groups were
5.79±2.30, 14.78±5.39, 22.8±4.64, 41.19±5.07 and 6.55±2.23%,
respectively. The results showed that both OA treatment (P<0.05)
and cav-1 overexpression (P<0.01) promoted HL-60 cell apoptosis,
and the apoptosis levels of the HL-60-LC+OA cells were
significantly higher than that of the HL-60+OA (P<0.01) or
HL-60-LC cells (P<0.01). These data suggest that OA and CAV-1
promote apoptosis of HL-60 cells, and the joint treatment had a
more significant pro-apoptotic effect (Fig. 4A).
The rate of apoptosis of the HL-60-LS3+OA cells did
not significantly differ from that of the HL-60 cells; however, the
rate of 8.23±3.16% (P<0.05) was lower than that of the HL-60+OA
cells, indicating that silencing of the cav-1 gene hindered OA
promotion of HL-60 cell apoptosis (Fig.
4B).
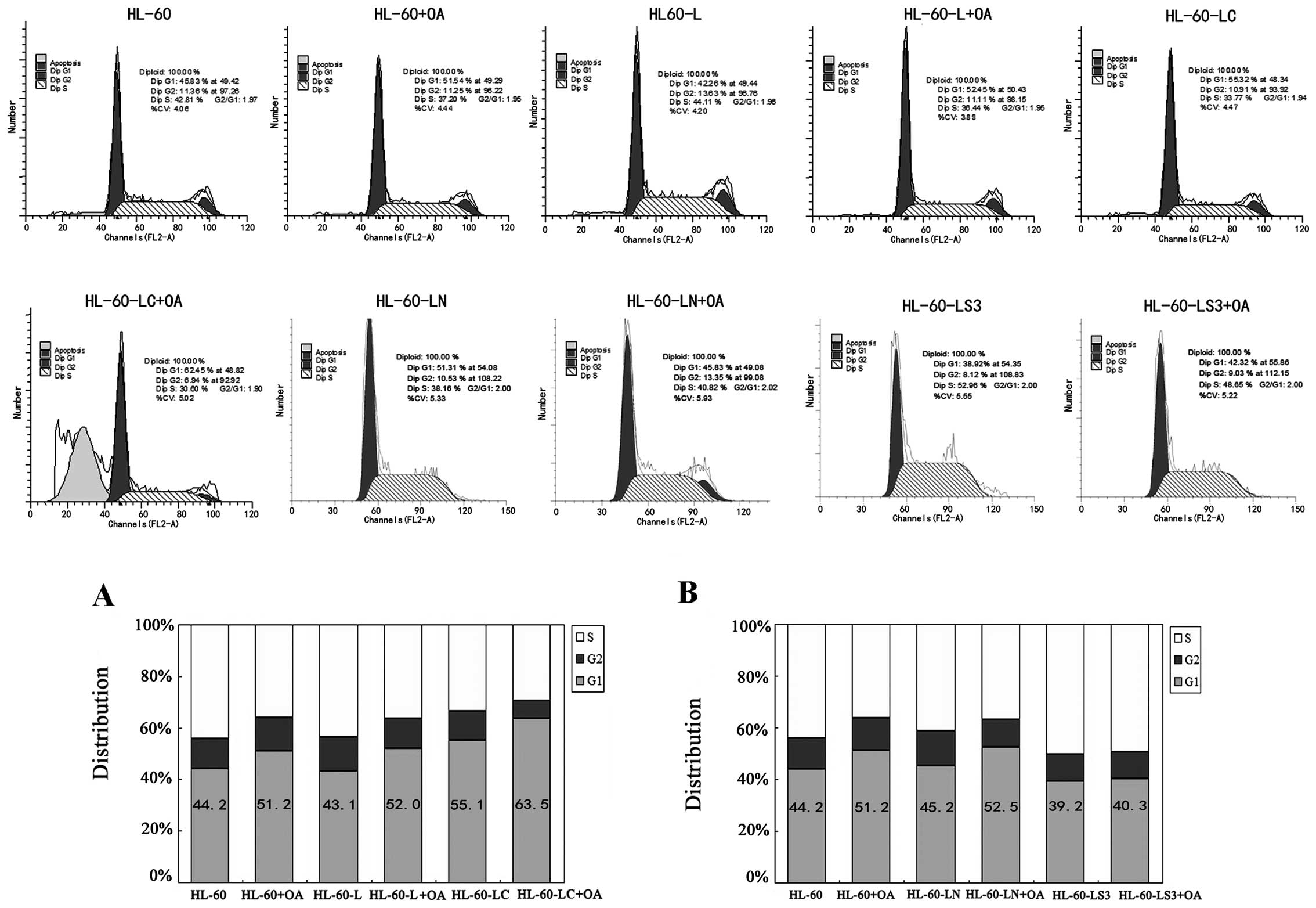
Detection of cell cycle progression
We performed cell cycle analyses to further examine
the effects of OA administration, cav-1 overexpression and
cav-1-siRNA on the HL-60 cell cycle (Fig. 5). The percentage of HL-60+OA cells
and HL-60-LC cells in the G1 phase increased by 7.0±1.1 (P<0.05)
and 10.9±1.9% (P<0.01), respectively. The percentage of
HL-60-LC+OA cells in the G1 phase increased by 12.3±0.6 (P<0.01)
and 8.4±1.4% (P<0.01) compared with the HL-60+OA and HL-60-LC
cells, respectively. Both OA and CAV-1 arrested the HL-60 cells at
the G1 phase of the cell cycle, and the cell cycle arrest was more
significant in the CAV-1+OA cells (Fig.
5A).
The percentage of HL-60-LS3+OA cells in the G1 phase
did not differ from that of the HL-60 group but was decreased by
10.9±3.4% (P<0.01) compared with that of the HL-60+OA group,
indicating that cav-1 gene silencing hampered the OA effect on the
G1 arrest of HL-60 cells (Fig.
5B).
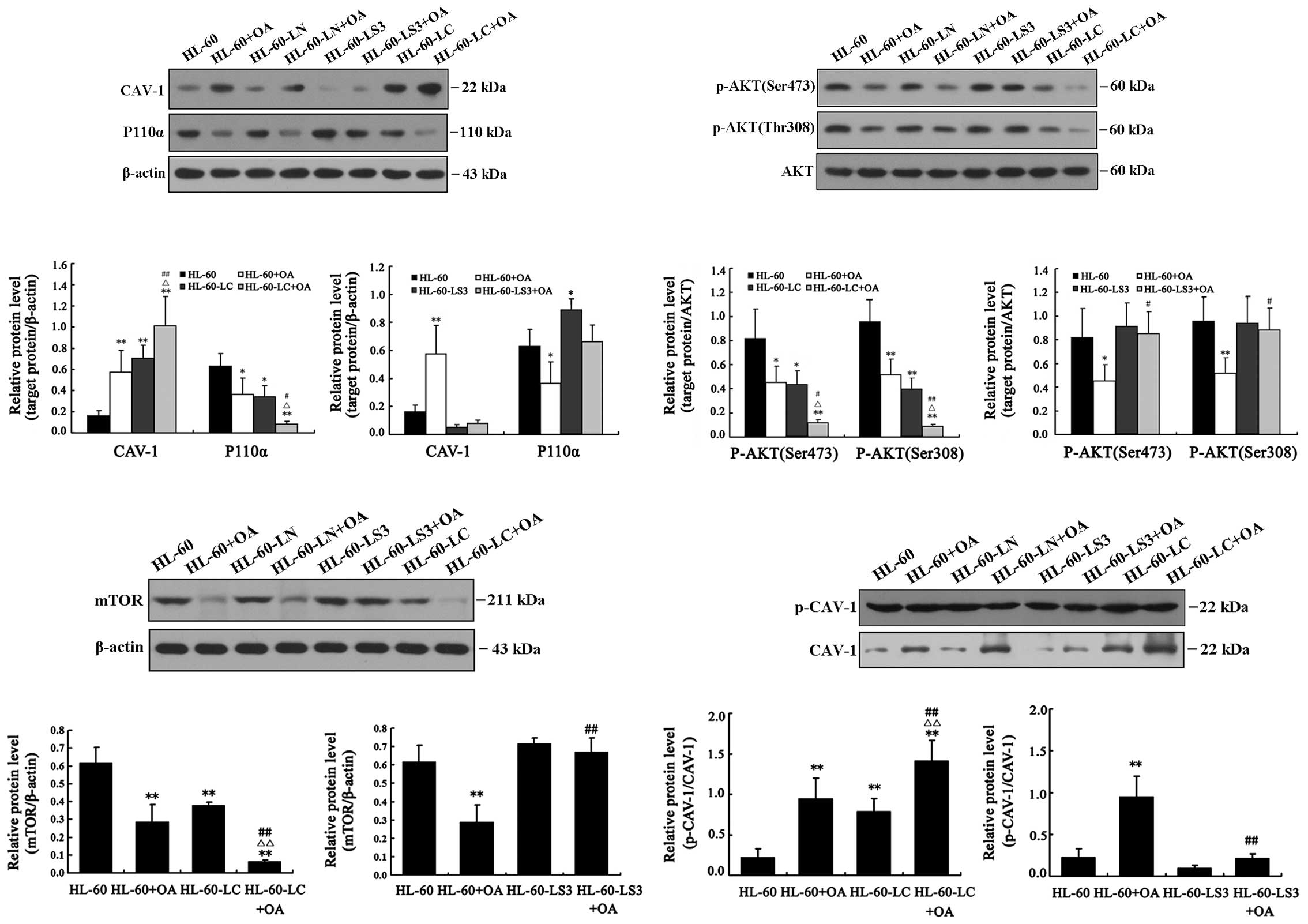
Western blot analysis results
The PI3K/AKT/mTOR signaling pathway is a key pathway
of tumorigenesis and tumor development. Therefore, we detected not
only CAV-1 protein levels and its phosphorylation but also the
levels of P110α, AKT, p-AKT (Ser473), p-AKT (Thr308) and mTOR to
determine the relationships between OA, CAV-1 and the PI3K/AKT/mTOR
signaling pathways.
Lentivirus-mediated overexpression of
cav-1 and OA treatment increased the expression of CAV-1 and CAV-1
(Tyr14) phosphorylation levels and cav-1 silencing inhibited the OA
effect on CAV-1 phosphorylation
Both OA and Lv-CAV-1 upregulated CAV-1 expression
levels, increased CAV-1α phosphorylation at Tyr14 and decreased AKT
phosphorylation at Ser473 and Thr308. The decreased CAV-1
expression in HL-60-LS3 cells indicated the effect of cav-1
silencing. CAV-1 expression in HL-60-LS3+OA cells was significantly
lower than in HL-60+OA cells (P<0.01) and showed no differences
compared with HL-60 cells, indicating that cav-1 siRNA blocked
OA-induced overexpression of CAV-1 (Fig. 6).
Cav-1 overexpression and OA treatment
suppressed the PI3K/AKT/mTOR signaling pathway; OA did not suppress
the pathway after cav-1 silencing
The expression levels of P110α, p-AKT (Ser473),
p-AKT (Thr308) and mTOR were decreased in the HL-60+OA, HL-60-LC
and HL-60-LC+OA cells. The smallest decrease was observed in the
HL-60-LC+OA cells. P110α expression levels were increased in the
HL-60-LS3 cells (P<0.05). The levels of P110α, p-AKT (Ser473),
p-AKT (Thr308) and mTOR were all lower in the HL-60+OA cells
compared with the HL-60-LS3+OA cells. Compared with the HL-60
cells, the levels of P110α, p-AKT (Ser473), p-AKT (Thr308) and mTOR
did not change in the HL-60-LS3+OA cells (Fig. 6). These results indicated that
silencing cav-1 upregulated P110α expression and blocked the
inhibitory effect of OA on the PI3K/AKT/mTOR signaling pathway. It
is likely that OA regulated cav-1 expression to alter the
downstream PI3K/AKT/mTOR signaling pathways, and CAV-1 is likely a
key factor in this regulatory cascade.
Discussion
Chemotherapy and bone marrow transplantation have
traditionally been the main treatments for leukemia. Hematopoietic
stem cell transplantation is difficult to widely implement due to
matching difficulties, while chemotherapy is toxic to the normal
cells of the body, making many patients abandon treatment due to
intolerance to side effects. Oleanolic acid is widely found in
plants such as privet fruit and the mileensis whole plant. It was
found in 1992 that as an active ingredient in the methanol extract
of clove, OA induced differentiation of murine leukemia cells and
inhibited HL-60 cell proliferation (22). In vitro experiments also
showed that OA inhibited tumor growth and reduced radiation-induced
damage to hematopoietic tissues (23). OA has been unexpectedly linked to
CAV-1 and the PI3K pathway, and this study was designed to further
explore the mechanism of OA in leukemia and to determine whether
CAV-1 plays a key role.
The cav-1 gene plays a role as a tumor
suppressor in HL-60 cells
cav-1 plays an important role in tumorigenesis and
the development of leukemia. To study the differences in cav-1 gene
expression between leukemia tumor cells and normal human
leukocytes, we chose the ALL cell line SUP-B15, the ANLL cell lines
HL-60 and THP-1, the CML cell line K562 and CLL primary tumor cells
(named CLP) as the experimental subjects and found low cav-1 mRNA
and protein expression in all cell lines. The HL-60 cell line with
the lowest cav-1 expression was selected for lentivirus-mediated
cav-1 gene transfection. The results showed that overexpression of
cav-1 inhibited HL-60 cell proliferation, promoted apoptosis,
provoked cell cycle arrest in the G1 phase and suppressed
PI3K/AKT/mTOR pathway activity. Thus, cav-1 plays a role as a tumor
suppressor in HL-60 cells.
The CAV-1 and PI3K/AKT/mTOR pathways are
involved in HL-60 cell proliferation inhibition, promotion of
apoptosis and cell cycle arrest in the G1 phase by OA
The nature of carcinogenesis is the deregulation of
signal transduction pathways leading to unlimited cell
proliferation. Searching for targets has been a focus of anticancer
drug discovery in recent years. The study of CAV-1 and
PI3K/AKT/mTOR signaling pathways has recently attracted attention
and has been mainly limited to mouse embryonic stem cells.
Estradiol 17β stimulates the proliferation of mouse ES cells partly
by upregulating CAV-1 through the Src, PI3K and ERK1/2 pathways
(24); the epidermal growth factor
can induce DNA synthesis and cell migration, in which CAV-1 plays a
potential role through the Src, FAK and PI3K/AKT signaling pathways
(25). Silencing the cav-1 gene can
also activate the estrogen receptor 36-dependent PI3K/AKT pathway
(26). CAV-1 participates in
fibronectin’s induction of mouse embryonic stem cells through the
FAK, RhoA, PI3K/AKT and ERK1/2 pathways (27). The relationship between CAV-1 and
PI3K/AKT/mTOR in leukemia has not yet been reported.
This experiment verified that OA not only
upregulated CAV-1 expression but also enhanced CAV-1
phosphorylation. At the same time, OA treatment inhibited the
PI3K/AKT pathway, leading to a series of changes in the activities
of P110α, AKT in the pathway and downstream mTOR. The cav-1
overexpression in HL-60 cells via transfection caused changes in
cell proliferation, apoptosis, cell cycle and the PI3K pathway,
consistent with OA treatment. These results suggest that OA likely
increases cav-1 expression by increasing its transcription or
translation efficiency or acting on its promoter, thereby
inhibiting the PI3K/AKT/mTOR signaling pathway to trigger a series
of biological reactions.
OA improves CAV-1 protein phosphorylation
in HL-60 cells
Using real-time quantitative RT-PCR, Tsuji et
al demonstrated that in adult T-cell leukemia cell lines, CpG
site methylation of the caveolin promoter was irrelevant to
caveolin expression levels, and that other mechanisms may be
involved in the regulation of caveolin expression (28). The changes of cav-1 epigenetic
modification in AML will be the focus of future studies. We
considered the change in CAV-1 phosphorylation while detecting the
CAV-1 protein expression level. CAV-1α has 31 more amino acid
residues than CAV-1β. It is widely believed that CAV-1α
phosphorylation at Tyr14, which is lacking in the β isoform, is the
key for tumorigenesis and development; this phosphorylation site
could be the key factor causing the two isoforms to function
differently. Therefore, in the present study, we detected p-CAV-1
(Tyr14) of the α isoform, and the results confirmed that OA
increased not only CAV-1 protein expression levels but also its
phosphorylation at Tyr14. p-CAV-1 may be the active form of CAV-1
that regulates cell proliferation, apoptosis and cell cycle
progression by regulating the PI3K/AKT pathway.
CAV-1 increases HL-60 cell sensitivity to
OA
After OA treatment in cav-1-overexpressing cell
lines, HL-60 cell proliferation activity decreased, the apoptotic
rate increased, the percentage of cells in the G1 phase increased,
and the PI3K/AKT/mTOR pathway was significantly inhibited. The
combination of CAV-1 and OA was significantly more effective than
any single factor, suggesting that CAV-1 can improve HL-60 cell
sensitivity to OA. Gene therapy supplemented by clinical medicine
will be a novel treatment for AML drug resistance.
CAV-1 is a key factor in OA function
After cav-1 gene silencing by RNA interference, OA
treatment could no longer regulate HL-60 cell proliferation,
apoptosis, cell cycle and the PI3K pathway, i.e., CAV-1 shRNA
hindered OA functioning in HL-60 cells. CAV-1 plays a very
important role as a key factor for OA function in HL-60 cells.
Cav-1 acts as an oncogene according to some studies.
For instance, the CAV-1 expression level is increased in renal
cancer cells, and the survival rate in patients with CAV-1 is
significantly lower than that of CAV-1-negative patients (29). CAV-1 showed an effect on anoikis
resistance in non-small cell lung cancer, and CAV-1-positive
patients have a poor prognosis (30). There have also been studies
suggesting that cav-1 acts as a tumor-suppressor gene. For example,
mutations at codon 132 and 141 of the cav-1 gene have been found in
breast cancer (31). CAV-1 can play
a regulatory role in tumors through a variety of pathways, such as
PI3K/AKT (32), MEK/ERK, RAS and
MAPK. Cav-1 gene deletion mutation, a high density of methylation
at CpG loci in the promoter region (33), abnormal Y14 phosphorylation of the
CAV-1α isoform (34), abnormal
activation or upregulated expression of CAV-1 by upstream
transcription factors and signaling proteins can cause cancer. The
results of this experiment found lower cav-1 expression levels in
AML tumor cells than in normal white blood cells, suggesting cav-1
to be a possible tumor-suppressor gene. OA can block cell cycle
progression through ERK-P53, promote apoptosis through the
mitochondrial pathway, inhibit the mTOR signaling pathway (7) or downregulate Bcl-2 to upregulate Bax
and Bad to achieve antitumor effects (8); VEGF is also involved in the OA
inhibitory effects on proliferation, invasion and tube formation by
endothelial cells (10). The
present study suggests that the cav-1 gene is a new target for the
treatment of AML through cav-1 overexpression in HL-60 cells and
provides a potential new strategy for research and development of
new drugs for AML treatment and drug resistance. As a natural
medicinal extract, OA can specifically increase CAV-1 protein
expression levels in this target and inhibit the PI3K/AKT pathway.
OA effectiveness was lost after cav-1 silencing, indicating that
CAV-1 plays a key role in how OA affects HL-60 cells suggesting
that OA can be used in AML therapy targeting CAV-1. However, how OA
promotes CAV-1 expression, whether CAV-1 protein directly or
indirectly inhibits the PI3K pathway, and where the anchor point is
located require further research.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (no. 81070428), the Natural Science
Foundation of Heilongjiang Province (D201024), the Heilongjiang
Provincial Health Department Scientific Research Project (no.
2010-484), the Heilongjiang Provincial Health Department Scientific
Research Project (no. 2013-176), the Jiamusi University Key
Scientific Research Project of Heilongjiang Provincial Health
Department Scientific Research Projects (no. 2010-228),
Heilongjiang Province Postdoctoral Scientific Research Startup
Funding (no. LBH-Q12005) and Mudanjiang City, Heilongjiang Province
Science and Technology Program (no. Z2013s048).
References
|
1
|
Huang WL: Signal Transduction. Beijing
People’s Medical Publishing House Press; Beijing: pp. 151–199.
2005
|
|
2
|
Zhou L, Ding Y, Chen W, Zhang P, Chen Y
and Lv X: The in vitro study of ursolic acid and oleanolic acid
inhibiting cariogenic microorganisms as well as biofilm. Oral Dis.
19:494–500. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang J, Shan A, Liu T, Zhang C and Zhang
Z: In vitro immunomodulatory effects of an oleanolic acid-enriched
extract of Ligustrum lucidum fruit (Ligustrum lucidum
supercritical CO2 extract) on piglet immunocytes. Int
Immunopharmacol. 4:758–763. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mapanga RF, Rajamani U, Dlamini N, et al:
Oleanolic acid: a novel cardioprotective agent that blunts
hyperglycemia-induced contractile dysfunction. Plos One.
7:e473222012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhang H, Zhang Y, Jiang YP, et al:
Curative effects of oleanolic acid on formed hypertrophic scars in
the rabbit ear model. Evid Based Complement Alternat Med.
2012:8375812012.PubMed/NCBI
|
|
6
|
Wang X, Bai H, Zhang X, et al:
ERK-p53-mediated cell cycle arrest and mitochondrial-dependent
apoptosis. Carcinogenesis. 34:1323–1330. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhou R, Zhang Z, Zhao L, et al: Inhibition
of mTOR signaling by oleanolic acid contributes to its anti-tumor
activity in osteosarcoma cells. J Orthop Res. 29:846–852. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pratheeshkumar P and Kuttan G: Oleanolic
acid induces apoptosis by modulating p53, Bax, Bcl-2 and caspase-3
gene expression and regulates the activation of transcription
factors and cytokine profile in B16F. J Environ Pathol Toxicol
Oncol. 30:21–31. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Feng L, Au-Yeung W, Xu YH, Wang SS, Zhu Q
and Xiang P: Oleanolic acid from Prunella Vulgaris L. induces
SPC-A-1 cell line apoptosis via regulation of Bax, Bad and Bcl-2
expression. Asian Pac J Cancer Prev. 12:403–408. 2011.
|
|
10
|
Wei JT, Liu M, Liu HZ, Zhao J, Xiao L, Han
LJ and Lin XK: Oleanolic acid inhibits proliferation of HUVECs, and
inhibits migration and tube formation via VEGF pathway. Yao Xue Xue
Bao. 47:1457–1462. 2012.(In Chinese).
|
|
11
|
Choi CY, You HJ and Jeong HG: Nitric oxide
and tumor necrosis factor-alpha production by oleanolic acid via
nuclear factor-kappaB activation in macrophages. Biochem Biophys
Res Commun. 288:49–55. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Schwencke C, Braun-Dullaeus RC, Wunderlich
C and Strasser RH: Caveolae and caveolin in transmembrane
signaling: Implications for human disease. Cardiovascular Res.
70:42–49. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yi JS, Mun DG, Lee H, et al: PTRF/cavin-1
is essential for multidrug resistance in cancer cells. J Proteome
Res. 12:605–614. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lavie Y, Fiucci G and Liscovitch M:
Up-regulation of caveolae and caveolar constituents in
multidrug-resistant cancer cells. J Biol Chem. 273:32380–32383.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fang K, Fu W, Beardsley AR, Sun X, Lisanti
MP and Liu J: Overexpression of caveolin-1 inhibits endothelial
cell proliferation by arresting the cell cycle at G0/G1 phase. Cell
Cycle. 6:199–204. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zou H, Stoppani E, Volonte D and Galbiati
F: Caveolin-1, cellular senescence and age-related diseases. Mech
Ageing Dev. 132:533–542. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bai L, Deng X, Li J, et al: Regulation of
cellular senescence by the essential caveolar component
PTRF/Cavin-1. Cell Res. 21:1088–1101. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Martinez-Outschoorn UE, Whitaker-Menezes
D, Lin Z, et al: Cytokine production and inflammation drive
autophagy in the tumor microenvironment: role of stromal caveolin-1
as a key regulator. Cell Cycle. 10:1784–1793. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu Y, Wang Y, Shi D and Zou W: Autophagy
and Caveolin-1 in cancer: a review. Chen Wu Gong Chen Xue Bao.
28:912–917. 2012.(In Chinese).
|
|
20
|
Pang A, Au WY and Kwong YL: Caveolin-1
gene is coordinately regulated with the multidrug resistance 1 gene
in normal and leukemic bone marrow. Leuk Res. 28:973–977. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sawada S, Ishikawa C, Tanji H, et al:
Overexpression of caveolin-1 in adult T-cell leukemia. Blood.
115:2220–2230. 2010. View Article : Google Scholar
|
|
22
|
Umehara K, Takagi R, Kuroyanagi M, Ueno A,
Taki T and Chen YJ: Studies on differentiation-inducing activities
of triterpenes. Chem Pharm Bull (Tokyo). 40:401–405. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hsu HY, Yang JJ and Lin CC: Effects of
oleanolic acid and ursolic acid on inhibiting tumor growth and
enhancing the recovery of hematopoietic system postirradiation in
mice. Cancer Lett. 111:7–13. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Park JH, Lee MY and Han HJ: A potential
role for CAV1 in estradiol-17 beta-induced proliferation of mouse
embryonic stem cells: involvement of Src, PI3K/Akt, and MAPKs
pathways. Int J Biochem Cell Biol. 41:659–665. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Park JH and Han HJ: CAV1 plays important
role in EGF-induced migration and proliferation of mouse embryonic
stem cells: involvement of PI3K/Akt and ERK. Am J Physiol Cell
Physiol. 297:C935–C944. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Feng S, Wang Y, Wang X, et al: CAV1 gene
silencing promotes the activation of PI3K/AKT dependent on
ERalpha36 and the transformation of MCF10ACE. Sci China Life Sci.
53:598–605. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Park JH, Ryu JM and Han HJ: Involvement of
CAV1 in fibronectin-induced mouse embryonic stem cell
proliferation: role of FAK, RhoA, PI3K/Akt, and ERK 1/2 pathways. J
Cell Physiol. 226:267–275. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tsuji Y, Nakagawa T, Hatanaka M, Takeuchi
T, Matsumoto E, Takenaka H and Shimizu A: Quantification of
caveolin isoforms using quantitative real-time RT-PCR, and analysis
of promoter CpG methylation of caveolin-1alpha in human T cell
leukemia cell lines. Int J Mol Med. 18:489–495. 2006.PubMed/NCBI
|
|
29
|
Joo HJ, Oh DK, Kim YS, Lee KB and Kim SJ:
Increased expression of CAV1 and microvessel density correlates
with metastasis and poor progrosis in clear cell renal cell
carcinoma. BJU Int. 93:291–296. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chunhacha P and Chanvorachote P: Roles of
Caveolin-1 on anoikis resistance in non small cell lung cancer. Int
J Physiol Pathophysiol Pharmacol. 4:149–155. 2012.PubMed/NCBI
|
|
31
|
Lee MY, Lee SH, Park JH and Han HJ:
Interaction of galectin-1 with caveolae induces mouse embryonic
stem cell proliferation through the Src, ERas, Akt and mTOR
signaling pathways. Cell Mol Life Sci. 66:1467–1478. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ryu JM and Han HJ: L-threonine regulates
G1/S phase transition of mouse embryonic stem cells via PI3K/Akt,
MAPKs, and mTORC pathways. J Biol Chem. 286:23667–23678. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Rao X, Evans J, Chae H, et al: CpG island
shore methylation regulates caveolin-1 expression in breast cancer.
Oncogene. 32:4519–4528. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Felicetti F, Parolini I, Bottero L, et al:
Caveolin-1 tumor-promoting role in human melanoma. Int J Cancer.
125:1514–1522. 2009. View Article : Google Scholar : PubMed/NCBI
|