Introduction
Osteosarcoma (OS) is the most common
non-haematological primary malignant bone tumour in children and
young adults (1). Although
neoadjuvant chemotherapy and improved surgical technology have
increased the survival rate to 65–75% (2), this combined treatment is still
unsuccessful in 30–40% of patients with localised tumours and in
80–85% of patients with metastatic disease at presentation
(3,4). Multidrug resistance (MDR), both
intrinsic and acquired, is still a major concern regarding the
clinical management of osteosarcoma patients and a key issue in the
failure of current treatment (5,6). The
development of an MDR phenotype can be mediated by several
mechanisms, including energy-dependent efflux of chemotherapeutic
drugs (7). The principal
transmembrane transporter responsible for this mechanism is
P-glycoprotein (P-gp), a drug efflux pump belonging to the
ATP-binding cassette (ABC) protein superfamily, encoded by the
multidrug resistance gene (MDR-1), which lowers intracellular drug
concentrations to sub-lethal levels (8,9).
Although P-gp appears to be involved in resistance or a poor
response to chemotherapy (10),
other undefined cellular factors also seem to participate in
modulating drug cytotoxicity in osteosarcoma cells. Thus, the
analysis of the molecular mechanisms underlying the resistance of
osteosarcoma cells to chemotherapy is essential for the development
of novel treatment strategies for this disease.
A growing body of evidence has demonstrated that the
microenvironment of the host has an important effect on the MDR of
tumours, including pH, temperature, partial pressure of oxygen
nutrition and extracellular matrix components (11–14).
In particular, there is a general consensus that hypoxia
dramatically decreases the chemosensitivity of tumour cells
promoting drug resistance to anticancer agents in a large variety
of neoplasias (15,16). A series of events occur during the
adaptation of a tumour to hypoxia through a group of
hypoxia-inducible factors (HIFs), of which HIF-1α appears to be the
most important in many different systems (17,18).
This is an heterodimeric protein that consists of a highly
regulated HIF-1α subunit and a constitutively expressed HIF-1β
subunit (19). HIF-1α is a
transcription factor which permits hypoxic tumour cells to
upregulate proteins that promote their survival and increase their
aggressiveness. When adequate oxygen (O2) is present,
the subunit HIF-1α becomes hydroxylated at several proline residues
and this leads to ubiquitination and proteasomal degradation.
However, when O2 is absent, this molecule survives and
translocates to the nucleus, where it forms a heterodimer with
HIF-1β (20). This dimer then binds
to highly conserved hypoxia-response elements (HREs) within
promoters of hypoxia-responsive genes inducing their transcription
(21). Genes containing functional
HREs encode proteins involved in angiogenesis (VEGF, endothelin-1),
maturation of red blood cells (erythropoietin, transferrin), energy
metabolism (glucose transporter 1 and 3), and cell proliferation
and viability (insulin-like growth factor 2, p21) (18). Notably, it has been well documented
that the MDR-1 gene, which encodes for P-glycoprotein, harbours
different HREs which are HIF-1α inducible (22). The drug resistance induced by
HIF-1α-mediated P-gp expression has been observed in a plethora of
tumour cells including glioma, breast carcinoma, gastric cancer and
colon cancer cells (23–26).
The aim of the present study was to investigate the
role of HIF-1α and its signalling pathways underlying drug
resistance in human osteosarcoma. For this purpose we generated and
characterised two drug-resistant osteosarcoma cell lines selected
for resistance to doxorubicin, a drug of choice in the treatment of
this tumour. Our data showed that the MDR phenotype in human
osteosarcoma cells was mediated by HIF-1α. A new potential model of
chemo-resistance in human osteosarcoma including target genes of
the non-canonical pathway of HIF-1α is proposed.
Materials and methods
Drug
Doxorubicin (DXR) was purchased from Sigma-Aldrich
(Milan, Italy). It was prepared as a 5 mg/ml fresh stock solution
in phosphate-buffered saline (PBS) and was stored at −20°C. The
drug was subsequently freshly diluted to the appropriate
concentration in the culture medium before each experiment.
Cell culture
The human osteosarcoma cell line MG-63 was obtained
from the American Type Culture Collection (ATCC; Manassas, VA, USA)
and grown in Iscove’s modified Dulbecco’s medium (IMDM), containing
10% heat inactivated fetal bovine serum (FBS) (Lonza) and
antibiotics (100 U/ml penicillin and 100 μg/ml streptomycin)
(Gibco). Each drug-resistant variant was continuously cultured in
the presence of the selective drug concentration. All cell lines
were maintained at 37°C in a humidified 5% CO2
atmosphere.
Isolation of DXR-resistant clones
DXR-resistant clones were established by continuous
exposure of the MG-63 cell line to increasing doses of DXR.
Initially, MG-63 cells were cultured in a medium containing 30
ng/ml DXR up to a concentration of 100 ng/ml. The DXR-resistant
cell lines were selected at 30 and 100 ng/ml and named MG-63DXR30
and MG-63DXR100, respectively. To maintain DXR resistance,
MG-63DXR30 and MG-63DXR100 cells were routinely cultured with the
appropriate concentration of DXR. From time to time, the
sensitivity of cells to DXR was evaluated by the analysis of their
resistance to cell death.
In vitro growth characteristics and
chemosensitivity
To determine the in vitro growth
characteristics of each cell line, cells were seeded in IMDM 10%
FBS in the absence of or with the appropriate concentration of DXR.
Cell density and viability were assessed by the erythrosine B
(Sigma-Aldrich) dye exclusion method. Doubling time was calculated
during the logarithmic phase of growth (from 48 to 96 h after
seeding). Drug sensitivity of each cell line was calculated from
the drug dose-response curve and expressed as IC50 (drug
concentration resulting in 50% inhibition of cell growth after 72 h
of in vitro treatment). The degree of DXR resistance was
expressed as the ratio of the IC50 value of the
resistant variant to that of the parental cell line.
Immunoblot analysis
Whole-cell extracts from the parental MG-63 cells
and the MG-63DXR30 and MG-63DXR100 resistant variants were analysed
by SDS-polyacrylamide gel electrophoresis (PAGE). The protein
concentration was quantified by bicinchoninic acid (BCA) (Pierce)
protein assay. Primary antibodies used were MDR-1/P-gp, HIF-1α,
c-Myc, p21 (Santa Cruz Biotechnology, Santa Cruz, CA, USA) and
β-actin (Sigma-Aldrich). Immunological complexes were visualised by
an ECL detection system (Amersham Biosciences, GE Healthcare) and
analysed using Quantity One software (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA).
Statistical analysis
The IC50 value was determined by the
linear regression method. All experiments were performed in
triplicate, unless otherwise indicated, and data values were
presented as means ± SE. Data were analysed using the Student’s
t-test; P-values of ≤0.05 were considered to indicate statistical
significance (P<0.001, P<0.01, P<0.05 are indicated).
Results
Selection and establishment of
DXR-resistant MG-63 sublines
DXR-resistant variants of the MG-63 human
osteosarcoma cell line were obtained by initially exposing the
parental cell line to 30 ng/ml DXR. Selection was then continued by
stepwise increased DXR concentrations up to 100 ng/ml.
Establishment of adequate in vitro growth at each new DXR
concentration required ~12–18 weeks (corresponding to between 8–20
in vitro passages). All the experiments were performed on
cell lines maintained in culture for at least 4–6 months after
selection with DXR. The established resistant cell lines were
selected at 30 and 100 ng/ml and named MG-63DXR30 and MG-63DXR100,
respectively.
In vitro growth characteristics
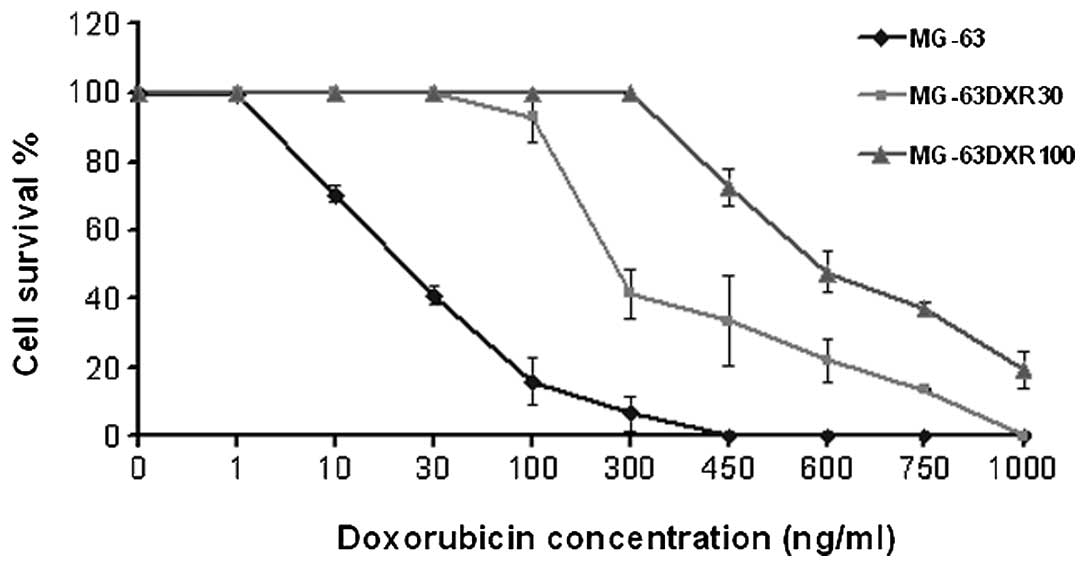
The IC50 values and the increased
resistance to DXR in the selected variants were drawn from the DXR
dose-response curve of each cell line (Fig. 1). The increase in DXR resistance
compared with that of the parental cell line ranged from 10-fold
for the MG-63DXR30 variant, to 28-fold for the MG-63DXR100 variant
(Table I). The analysis of in
vitro growth characteristics of the DXR-resistant variants
showed that the doubling time of the MG-63DXR30 cells was similar
to that of the MG-63 parental cell line whereas it was
significantly longer in the MG-63DXR100 cells (Table I).
 | Table IIC50 values, the increase
in the doxorubicin resistance and the doubling time of MG-63 cells
and its doxorubicin-resistant variants. |
Table I
IC50 values, the increase
in the doxorubicin resistance and the doubling time of MG-63 cells
and its doxorubicin-resistant variants.
| Cell line | IC50
valuesa (ng/ml) | Increase in drug
resistanceb | Doubling time
(h) |
|---|
| MG-63 | 20±3.6 | - | 23.1 |
| MG-63DXR30 | 200±14.2 | 10-fold | 29 |
| MG-63DXR100 | 559±29 | 28-fold | 34.5 |
Analysis of protein expression
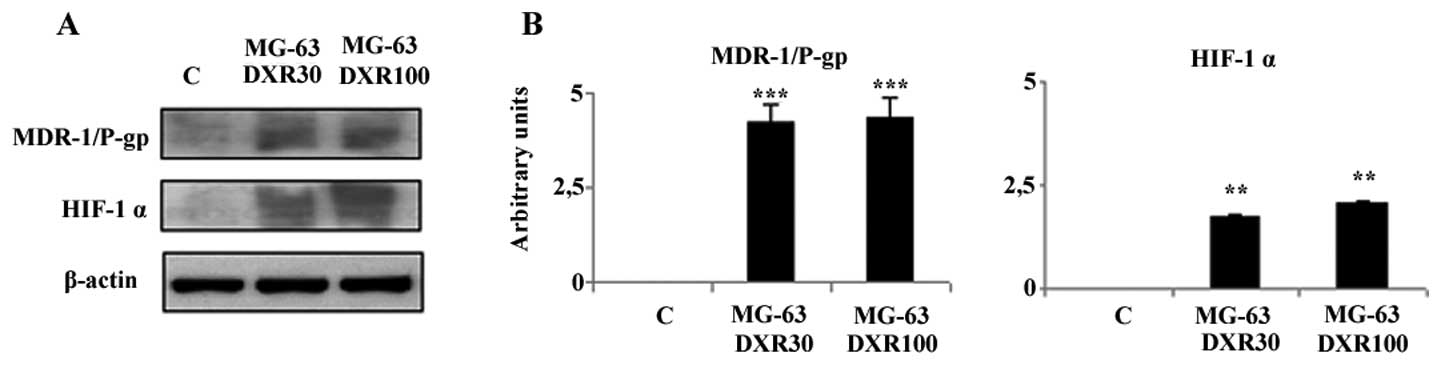
MDR-1/P-gp and HIF-1α are activated in DXR-resistant
MG-63 cells. To evaluate whether the drug-resistance acquired by
MG-63 cells is associated with MDR-1/P-gp activation, we examined
its expression by western blot analysis. As shown in Fig. 2, a marked induction of this protein
was evident in the DXR-resistant cell lines, whereas in the
parental MG-63 cell line, MDR-1/P-gp was undetectable. In order to
understand the mechanism of drug-resistance induced by DXR in this
cell line we investigated whether the drug exposure leads to the
activation of HIF-1α, known to enhance MDR-1 gene transcription. As
shown in Fig. 2, a strong induction
of HIF-1α was noted both in the MG-63DXR30 and MG-63DXR100 cells
when compared to that in the parental cell line that did not
express this protein. Therefore, the continuous exposure to DXR
seems to significantly activate HIF-1α and consequently MDR-1/P-gp,
the responsible factor of drug-resistance development.
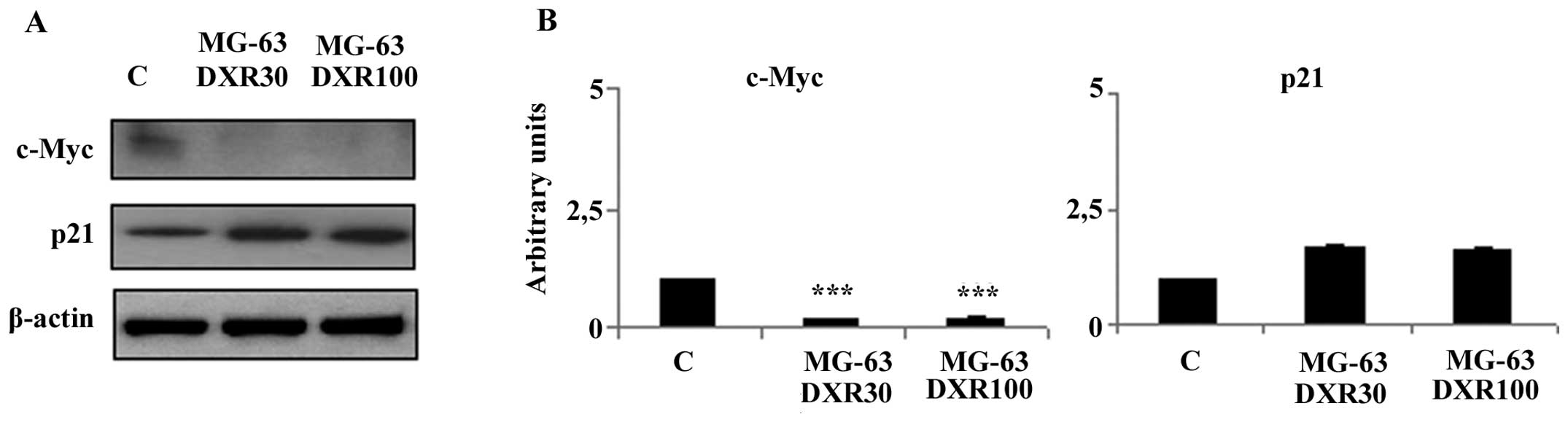
HIF-1α inhibits DXR-mediated
apoptosis
It is known that HIF-1α activity leads to the
upregulation or downregulation of target genes that are involved in
many aspects of cancer progression, cell proliferation and
survival. To better understand the role of HIF-1α in DXR resistance
acquired by MG-63 cells, we analysed the expression of several
proteins encoded by HIF-1α target genes involved in the apoptotic
pathways induced by DXR. Using western blot analysis we observed a
significant decrease in the c-Myc expression levels in both
DXR-resistant variants when compared to this level in the parental
cell line (Fig. 3). Moreover, the
downregulation of this protein was accompanied by an increased p21
protein expression (Fig. 3).
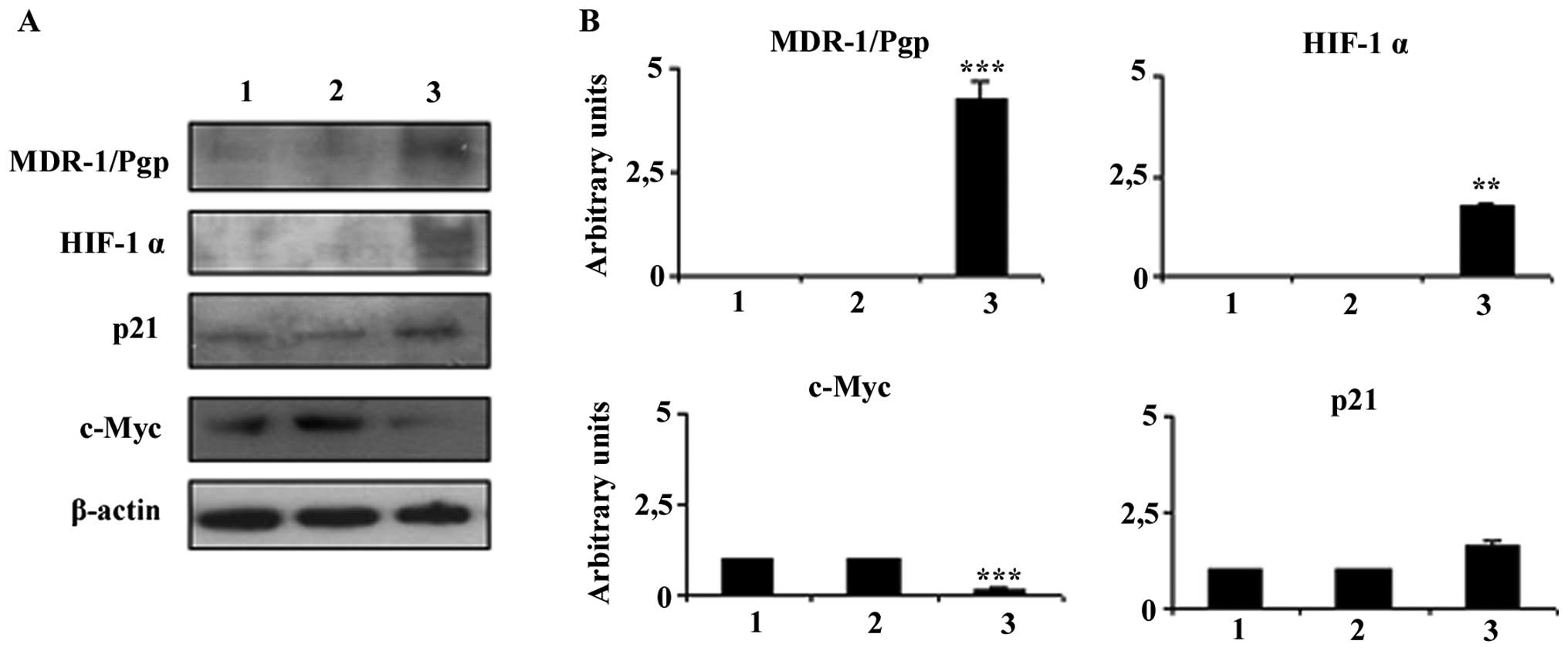
Stability of in vitro DXR resistance
The MG-63 DXR-resistant phenotypes were stable up to
2 months (8–10 in vitro passages) of continuous culture in
drug-free medium. After this period, the level of DXR-resistance
was significantly decreased with an IC50 value
comparable to the MG-63 parental cell line (data not shown). In
parallel, the expression of the markers identified and linked to
DXR resistance in the MG-63 cells gradually returned to
parental-like levels (Fig. 4).
Discussion
Neoadjuvant chemotherapy with subsequent surgical
resection has produced great advancement in osteosarcoma therapy in
the last 20 years, with a more favourable prognosis and a nearly
70% long-term survival rate. However, in certain cases, relapse
occurs in initially responsive patients due to the emergence of
tumour resistance to anticancer agents (27,28).
This phenomenon may be caused by multiple factors including hypoxia
(15). In the present study, using
human doxorubicin-resistant osteosarcoma cells under normoxic
conditions, we demonstrated that the development of drug resistance
described in this tumour may be related to the activation of
HIF-1α, the main factor responsible for the adaptation of a tumour
to O2 deficiency. The critical mechanism of drug
resistance, both intrinsic and acquired, involves the ABC
(ATP-binding cassette) protein transporters P-gp, MRP1 and ABCG2,
which pump drug molecules out of cells (7). We first observed that selected
doxorubicin-resistant MG-63 cells presented a strong induction of
the MDR-1/P-gp protein, thus demonstrating that the resistance
developed by these human osteosarcoma cells as a consequence of
doxorubicin treatment was mediated by MDR-1 gene activation. This
result is in agreement with data indicating that P-gp can detect
and bind a large variety of anticancer drugs and other hydrophobic
compounds, including anthracyclines such as daunorubin, epirubicin,
mitoxantrone and doxorubicin (29).
This drug binding activity results in the activation of one of the
P-gp ATP binding domains and subsequent hydrolysis of ATP, leading
to a major change in the shape of P-gp, which causes extrusion of
the drug from the cancer cell (30).
Subsequently, we also observed that
doxorubicin-resistant MG-63 osteosarcoma cells obtained and
maintained in normoxic conditions showed a significant induction of
HIF-1α expression, which may be caused by its decreased degradation
in response to hypoxic stress induced by doxorubicin. Indeed, there
are many toxic mechanisms caused by this drug in mammalian tissues,
with an increase in free radical formation and in oxidative stress
as the most likely (31).
Doxorubicin undergoes chemical reduction to a semiquinone radical,
which in turn reduces oxygen to a superoxide that may contribute to
cytotoxicity (32).
Previous reports showed that an increase in
MDR-1/P-gp expression associated with the activation of HIF-1α in
tumour cells may be due to the presence of a functional HIF-1α
binding site (HRE) within the MDR-1 gene promoter (16,22–24).
On the basis of these observations, we suggest that doxorubicin, by
producing hypoxic stress in MG-63 cells, activates HIF-1α which in
turn induces MDR-1 gene transcription, and consequently the
expression of MDR-1/P-gp, responsible for the doxorubicin
resistance developed by this cell line.
Previous reports have demonstrated that HIF-1α not
only controls P-gp expression, thus, limiting the drug accumulation
within cells, but that it is also able to modify the cellular
response to the chemotherapeutic agent, for example by altering
drug-induced apoptosis, thus nullifying its healthy effect
(33). On the basis of this
finding, we analysed the expression of molecules involved in the
apoptotic pathways mediated by doxorubicin, and observed a
significant downregulation of c-Myc in parallel with an increase in
p21 protein expression in both doxorubicin-resistant clones. c-Myc
is a transcription factor with a dual capacity, as it is involved
both in cellular death and in cell proliferation, and is known to
be required for the induction of apoptosis by doxorubicin (34). The c-Myc downregulation in
doxorubicin-resistant MG-63 cells together with MDR-1/P-gp
overexpression suggest an inverse correlation between the two
proteins in human osteosarcoma, as already described for N-Myc and
MDR-1 in neuroblastoma (35) and
for c-Myc and MDR-1 in rhabdomyosarcoma (36). Importantly, HIF-1α has recently been
found to repress the activity of various molecules, including
c-Myc, by an innovative HRE-independent mechanism that does not
require DNA binding activity (37,38).
Moreover, Hayashi et al (39) demonstrated a non-canonical mechanism
of action of HIF-1α that counteracts c-Myc effects on gene
expression. In the present study, the HIF-1α-c-Myc pathway was
shown to play a role in mediating drug resistance of human
osteosarcoma cells. Therefore, our results are consistent with the
study of Hayashi et al, which identified the escape from
apoptosis driven by the HIF-1α-c-Myc pathway among the mechanisms
of malignant progression in tumourigenesis.
In addition, several regulators of the cell cycle,
notably p15, p18, p27, p57 and p21, are targets for repression by
c-Myc (40,41) and the increase in p21 expression
levels that we observed in the doxorubicin-resistant MG-63 cells
may be explained by the lack of c-Myc inhibitory activity on its
promoter. Although p21 was initially thought to be an inhibitor of
cell cycle progression, other studies have suggested that this
protein might also be a positive modulator of cell survival and
cell cycle progression and may protect various types of cells from
death following anticancer treatments (42,43).
In particular, this pro-survival activity has been observed in
breast adenocarcinoma cells treated with chemotherapeutic agents
such as taxol, where high levels of p21 appeared to be correlated
with enhanced survival and chemoresistance (44). Furthermore, high levels of p21 have
also been associated with drug resistance in acute myelogenous
leukaemia, head and neck carcinomas, and colon carcinoma (45–47).
Moreover, in many late-stage glioblastomas, known to be
extraordinarily resistant to chemotherapy and radiation (48,49),
resistance to apoptosis has been linked to elevated p21 levels
(50,51). Altogether, these data support a
positive role for p21 in tumour cell survival; this protein may be
induced by chemotherapeutic agents promoting its expression, as
observed in our doxorubicin-resistant MG-63 cells.
Lastly, we evaluated whether or not the
doxorubicin-resistant MG-63 cells maintained their acquired
resistance with time. After 2 months of continuous culture in
drug-free medium these cells lost their resistance and became
sensitive to the doxorubicin cytotoxic activity similar to the
MG-63 parental cell line. Likewise, we observed that the HIF-1α,
MDR-1/P-gp, c-Myc and p21 expression returned to parental cell-like
levels. The cells no longer exposed to the drug appeared to
gradually return to a basal condition, where HIF-1α is degraded by
the ubiquitine-proteasome system, thus confirming the role of
doxorubicin in the induction of its expression in tumour cells.
In conclusion, we suggest a key role for HIF-1α in
MDR development in human osteosarcoma MG-63 cell lines. On the one
hand, it facilitates the outward transport of intracellular
doxorubicin by expressing P-glycoprotein; on the other hand, it
contrasts the apoptotic drug effect by downregulating c-Myc and
consequently inducing p21 overexpression. This may be considered a
new model of chemo-resistance in human osteosarcoma involving two
different pathways of HIF-1α and may provide an experimental basis
for clinical applications in the field of MDR in human
osteosarcoma.
Acknowledgements
The present study was supported by grants from the
Italian Association for the Cancer Research (grant no. 11426 to
Professor N.B.) and the Italian Ministry of Health, Financial
Support for Scientific Research ‘5 per mille’ 2010.
References
|
1
|
Heare T, Hensley MA and Dell’Orfano S:
Bone tumors: osteosarcoma and Ewing’s sarcoma. Curr Opin Pediatr.
21:365–372. 2009.
|
|
2
|
Mankin HJ, Hornicek FJ, Rosenberg AE,
Harmon DC and Gebhardt MC: Survival data for 648 patients with
osteosarcoma treated at one institution. Clin Orthop Relat Res.
429:286–291. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bacci G, Briccoli A, Rocca M, Ferrari S,
Donati D, Longhi A, Bertoni F, Bacchini P, Giacomini S, Forni C,
Manfrini M and Galletti S: Neoadjuvant chemotherapy for
osteosarcoma of the extremities with metastases at presentation:
recent experience at the Rizzoli Institute in 57 patients treated
with cisplatin, doxorubicin, and a high dose of methotrexate and
ifosfamide. Ann Oncol. 14:1126–1134. 2003. View Article : Google Scholar
|
|
4
|
Lewis IJ, Nooij MA, Whelan J, Sydes MR,
Grimer R, Hogendoorn PC, Memon MA, Weeden S, Uscinska BM, van
Glabbeke M, Kirkpatrick A, Hauben EI and Craft AW: Improvement in
histologic response but not survival in osteosarcoma patients
treated with intensified chemotherapy: a randomized phase III trial
of the European Osteosarcoma Intergroup. J Natl Cancer Inst.
99:112–128. 2007. View Article : Google Scholar
|
|
5
|
Baldini N, Scotlandi K, Barbanti-Bròdano
G, Manara MC, Maurici D, Bacci G, Bertoni F, Picci P, Sottili S,
Campanacci M and Serra M: Expression of P-glycoprotein in
high-grade osteosarcomas in relation to clinical outcome. N Engl J
Med. 333:1380–1385. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bielack SS, Carrle D, Hardes J, Schuck and
Paulussen M: Bone tumors in adolescents and young adults. Curr
Treat Options Oncol. 9:67–80. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gottesman MM, Fojo T and Bates SE:
Multidrug resistance in cancer: role of ATP-dependent transporters.
Nat Rev Cancer. 2:48–58. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Borst P and Elferink RO: Mammalian ABC
transporters in health and disease. Annu Rev Biochem. 71:537–592.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ambudkar SV, Kimchi-Sarfaty C, Sauna ZE
and Gottesman MM: P-glycoprotein: from genomics to mechanism.
Oncogene. 22:7468–7485. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Baldini N, Scotlandi K, Serra M, Picci P,
Bacci G, Sottili S and Campanacci M: P-glycoprotein expression in
osteosarcoma: a basis for risk-adapted adjuvant chemotherapy. J
Orthop Res. 17:629–632. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Harguindey S, Orive G, Luis Pedraz J,
Paradiso A and Reshkin SJ: The role of pH dynamics and the
Na+/H+antiporter in the etiopathogenesis and
treatment of cancer. Two faces of the same coin - one single
nature. Biochim Biophys Acta. 1756:1–24. 2005.PubMed/NCBI
|
|
12
|
Shicang Y, Guijun H, Guisheng Q, Yuying L,
Guoming W and Ruiling G: Efficacy of chemotherapeutic agents under
hypoxic conditions in pulmonary adenocarcinoma multidrug resistant
cell line. J Chemother. 19:203–211. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen KG and Sikic BI: Molecular pathways:
regulation and therapeutic implications of multidrug resistance.
Clin Cancer Res. 18:1863–1869. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cheng GM and To KK: Adverse cell culture
conditions mimicking the tumor microenvironment upregulate ABCG2 to
mediate multidrug resistance and a more malignant phenotype. ISRN
Oncol. 2012:7460252012. View Article : Google Scholar
|
|
15
|
Rohwer N and Cramer T: Hypoxia-mediated
drug resistance: novel insights on the functional interaction of
HIFs and cell death pathways. Drug Resist Updat. 14:191–201. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Semenza GL: Hypoxia-inducible factors:
mediators of cancer progression and targets for cancer therapy.
Trends Pharmacol Sci. 33:207–214. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
El Naggar A, Clarkson P, Zhang F, Mathers
J, Tognon C and Sorensen PH: Expression and stability of hypoxia
inducible factor 1α in osteosarcoma. Pediatr Blood Cancer.
59:1215–1222. 2012.
|
|
18
|
Ke Q and Costa M: Hypoxia-inducible
factor-1 (HIF-1). Mol Pharmacol. 70:1469–1480. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang GL, Jiang BH, Rue EA and Semenza GL:
Hypoxia-inducible factor 1 is a basic-helix-loop-helix-PAS
heterodimer regulated by cellular O2 tension. Proc Natl
Acad Sci USA. 92:5510–5514. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kallio PJ, Pongratz I, Gradin K, McGuire J
and Poellinger L: Activation of hypoxia-inducible factor 1α:
posttranscriptional regulation and conformational change by
recruitment of the Arnt transcription factor. Proc Natl Acad Sci
USA. 94:5667–5672. 1997.
|
|
21
|
Semenza GL: Hypoxia-inducible factor 1:
master regulator of O2 homeostasis. Curr Opin Genet Dev.
8:588–594. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Comerford KM, Wallace TJ, Karhausen J,
Louis NA, Montalto MC and Colgan SP: Hypoxia-inducible
factor-1-dependent regulation of the multidrug resistance
(MDR1) gene. Cancer Res. 62:3387–3394. 2002.PubMed/NCBI
|
|
23
|
Chen L, Feng P, Li S, Long D, Cheng J, Lu
Y and Zhou D: Effect of hypoxia-inducible factor-1α silencing on
the sensitivity of human brain glioma cells to doxorubicin and
etoposide. Neurochem Res. 34:984–990. 2009.
|
|
24
|
Li J, Shi M, Cao Y, Yuan W, Pang T, Li B,
Sun Z, Chen L and Zhao RC: Knockdown of hypoxia-inducible factor-1α
in breast carcinoma MCF-7 cells results in reduced tumor growth and
increased sensitivity to methotrexate. Biochem Biophys Res Commun.
342:1341–1351. 2006.
|
|
25
|
Liu L, Ning X, Sun L, Zhang H, Shi Y, Guo
C, Han S, Liu J, Sun S, Han Z, Wu K and Fan D: Hypoxia-inducible
factor-1 α contributes to hypoxia-induced chemoresistance in
gastric cancer. Cancer Sci. 99:121–128. 2008.
|
|
26
|
Ding Z, Yang L, Xie X, Xie F, Pan F, Li J,
He J and Liang H: Expression and significance of hypoxia-inducible
factor-1 alpha and MDR1/P-glycoprotein in human colon carcinoma
tissue and cells. J Cancer Res Clin Oncol. 136:1697–1707. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chi SN, Conklin LS, Qin J, Meyers PA,
Huvos AG, Healey JH and Gorlick R: The patterns of relapse in
osteosarcoma: the Memorial Sloan-Kettering experience. Pediatr
Blood Cancer. 42:46–51. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chou AJ and Gorlick R: Chemotherapy
resistance in osteosarcoma: current challenges and future
directions. Expert Rev Anticancer Ther. 6:1075–1085. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Saraswathy M and Gong S: Different
strategies to overcome multidrug resistance in cancer. Biotechnol
Adv. 31:1397–1407. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gillet JP, Efferth T and Remacle J:
Chemotherapy-induced resistance by ATP-binding cassette transporter
genes. Biochim Biophys Acta. 1775:237–262. 2007.PubMed/NCBI
|
|
31
|
Yokochi T and Robertson KD: Doxorubicin
inhibits DNMT1, resulting in conditional apoptosis. Mol Pharmacol.
66:1415–1420. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wardman P: Electron transfer and oxidative
stress as key factors in the design of drugs selectively active in
hypoxia. Curr Med Chem. 8:739–761. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Weidemann A and Johnson RS: Biology of
HIF-1α. Cell Death Differ. 15:621–627. 2008.
|
|
34
|
Grassilli E, Ballabeni A, Maellaro E, Del
Bello B and Helin K: Loss of MYC confers resistance to
doxorubicin-induced apoptosis by preventing the activation of
multiple serine protease- and caspase-mediated pathways. J Biol
Chem. 279:21318–21326. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Nakagawara A, Kadomatsu K, Sato S, Kohno
K, Takano H, Akazawa K, Nose Y and Kuwano M: Inverse correlation
between expression of multidrug resistance gene and N-myc
oncogene in human neuroblastomas. Cancer Res. 50:3043–3047.
1990.PubMed/NCBI
|
|
36
|
Prados J, Melguizo C, Fernandez A, Aranega
AE, Alvarez L and Aranega A: Inverse expression of mdr1 and
c-myc genes in rhabdomyosarcoma cell line resistant to
actinomycin d. J Pathol. 180:85–89. 1996.
|
|
37
|
Dang CV, Kim JW, Gao P and Yustein J: The
interplay between MYC and HIF in cancer. Nat Rev Cancer. 8:51–56.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yoo YG, Christensen J and Huang LE: HIF-1α
confers aggressive malignant traits on human tumor cells
independent of its canonical transcriptional function. Cancer Res.
71:1244–1252. 2011.
|
|
39
|
Hayashi M, Yoo YG, Christensen J and Huang
LE: Requirement of evading apoptosis for HIF-1α-induced malignant
progression in mouse cells. Cell Cycle. 10:2364–2372.
2011.PubMed/NCBI
|
|
40
|
Wanzel M, Herald S and Eilers M:
Transcriptional repression by Myc. Trends Cell Biol. 13:146–150.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Herkert B and Eilers M: Transcriptional
repression: the dark side of myc. Genes Cancer. 1:580–586. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Gorospe M, Wang X and Holbrook NJ:
Functional role of p21 during the cellular response to stress. Gene
Expr. 7:377–385. 1999.PubMed/NCBI
|
|
43
|
Li Y, Dowbenko D and Lasky LA: AKT/PKB
phosphorylation of p21Cip/WAF1 enhances protein
stability of p21Cip/WAF1 and promotes cell survival. J
Biol Chem. 277:11352–11361. 2002.PubMed/NCBI
|
|
44
|
Barboule N, Chadebech P, Baldin V, Vidal S
and Valette A: Involvement of p21 in mitotic exit after paclitaxel
treatment in MCF-7 breast adenocarcinoma cell line. Oncogene.
15:2867–2875. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhang W, Kornblau SM, Kobayashi T, Gambel
A, Claxton D and Deisseroth AB: High levels of constitutive
WAF1/Cip1 protein are associated with chemoresistance in acute
myelogenous leukaemia. Clin Cancer Res. 1:1051–1057.
1995.PubMed/NCBI
|
|
46
|
Erber R, Klein W, Andl T, Enders C, Born
Al, Conradt C, Bartek J and Bosch FX: Aberrant
p21CIP1/WAF1 protein accumulation in head-and-neck
cancer. Int J Cancer. 74:383–389. 1997.
|
|
47
|
Wang Y, Blandino G and Givol D: Induced
p21waf expression in H1299 cell line promotes cell
senescence and protects against cytotoxic effect of radiation and
doxorubicin. Oncogene. 18:2643–2649. 1999.
|
|
48
|
Haas-Kogan D, Shalev N, Wong M, Mills G,
Yount G and Stokoe D: Protein kinase B (PKB/Akt) activity is
elevated in glioblastoma cells due to mutation of the tumor
suppressor PTEN/MMAC. Curr Biol. 8:1195–1198. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wang CC, Liao YP, Mischel PS, Iwamoto KS,
Cacalano NA and McBride WH: HDJ-2 as a target for
radiosensitization of glioblastoma multiforme cells by the
farnesyltransferase inhibitor R115777 and the role of p53/p21
pathway. Cancer Res. 66:6756–6762. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Glaser T, Wagenknecht B and Weller M:
Identification of p21 as a target of cycloheximide-mediated
facilitation of CD95-mediated apoptosis in human malignant glioma
cells. Oncogene. 20:4757–4767. 2001. View Article : Google Scholar
|
|
51
|
Happold C, Roth P, Wick W, Schmidt N,
Florea AM, Silginer M, Reifenberger G and Weller M: Distinct
molecular mechanisms of acquired resistance to temozolomide in
glioblastoma cells. J Neurochem. 122:444–455. 2012. View Article : Google Scholar : PubMed/NCBI
|