Introduction
Gastric cancer is highly prevalent in developing
nations, particularly in China, yet there is a lack of effective
drugs for the treatment of advanced gastric cancer (1). Recently, tumor necrosis factor-related
apoptosis-inducing ligand (TRAIL) has received much attention due
to its high specificity and low toxicity in anticancer treatment
(2,3). However, various types of gastric
cancer cells are resistant to TRAIL (4,5) and
the mechanism of TRAIL resistance is not fully understood.
Survival pathway activation is known to be involved
in TRAIL resistance. For example, a previous study showed that
TRAIL-induced activation of epidermal growth factor receptor (EGFR)
resulted in TRAIL resistance in colorectal cancer cells (6), and we recently demonstrated that TRAIL
induced EGFR activation in TRAIL-resistant gastric cancer cells.
The plasma membrane lipid rafts regulate TRAIL-induced EGFR
activation (7). Lipid rafts are
composed of multiple molecules, but it is not clear which molecule
plays the most important role in the regulation of EGFR activation.
The scaffolding protein caveolin-1 is enriched in lipid rafts,
where it takes on a specialized role in regulating the activity of
cell signaling receptors and their downstream effectors, such as
EGFR (8). However, although
caveolin-1 phosphorylation was shown to be required for
stretch-induced EGFR/Akt activation in mesangial cells (9), its involvement in the regulation of
TRAIL-induced EGFR activation warrants further investigation.
EGFR is typically activated by its ligands, but it
can also be regulated by the non-receptor tyrosine kinase Src. In
non-small cell lung cancer cells, transfection with Src short
hairpin RNA diminished phosphorylation of EGFR and ErbB2 and
increased sensitivity to EGFR tyrosine kinase inhibitor (10). In mesangial cells, Src also promoted
the phosphorylation of caveolin-1 and vascular endothelial growth
factor (VEGF)-induced fibronectin upregulation (11). Moreover, Src activation increased
caveolin-1 phosphorylation and promoted nitric oxide
synthase/caveolin-1 binding in vascular endothelial cells (12). However, the mechanism by which Src
and caveolin-1 regulate TRAIL-induced EGFR activation is
unknown.
In the present study, we showed that Src interacted
with both EGFR and caveolin-1 to regulate EGFR activation, thus
antagonizing TRAIL-induced apoptosis in gastric cancer cells.
Materials and methods
Reagents and antibodies
Recombinant human TRAIL was purchased from
Cytolab/Peprotech Asia (USA). Src inhibitor PP2 was obtained from
Sigma (St. Louis, MO, USA). Anti-EGFR, anti-phospho-EGFR (Tyr1068),
anti-Src, anti-phospho-Src (Tyr416), anti-Akt and anti-phospho-Akt
(Ser473) antibodies were obtained from Cell Signaling Technology
(USA). Anti-ERK, anti-p-ERK 1/2 (Thr 202/Tyr 204), and anti-actin
antibodies were obtained from Santa Cruz Biotechnology (USA).
Anti-caveolin-1 and anti-phospho-caveolin-1 (Tyr14) antibodies were
obtained from BD Technology Co. (USA).
Cell cultures
SGC-7901 and MGC-803 cells were obtained from the
Type Culture Collection of the Chinese Academy of Sciences (China).
The cells were cultured in RPMI-1640 medium containing 10%
heat-inactivated fetal bovine serum at 37°C under an atmosphere of
95% air and 5% CO2.
Flow cytometric analysis
SGC-7901 and MGC-803 cells were incubated with the
indicated concentrations of agents for the indicated times. After
being fixed with ice-cold 70% ethanol overnight, the samples were
incubated with 20 μg/ml RNase A and 10 μg/ml propidium iodide for
30 min in the dark. Finally, the samples were evaluated by flow
cytometry, and the data were analyzed with CellQuest software
(Becton-Dickinson, San Jose, CA, USA).
Small interfering RNA transfections
Cavolin-1 small interfering RNA (siRNA) was obtained
from Shanghai GeneChem Co. Ltd. (China). Cavolin-1 siRNA was
synthesized: 5′-AACCAGAAGGGACACACAGUU-3′ (sense) and
5′-AACUGUGUGUCCCUUCUGGUU-3′ (antisense). SGC-7901 and MGC-803 cells
were transiently transfected with cavolin-1 siRNA using
Lipofectamine 2000 reagent (Invitrogen, USA), according to the
manufacturer’s instructions.
Isolation of lipid rafts
SGC-7901 and MGC-803 cells were solubilized in 150
μl of prechilled TXNE buffer (50 mM Tris-HCl pH 7.4, 150 mM NaCl, 5
mM EDTA and 0.1% Triton X-100) containing protease inhibitors
(chymostatin, leupeptin, antipain and pepstatin). Subsequently, the
cells were moved into 35% OptiPrep (Axis-Shield, Norway) in
polyallomer ultra tubes (Sorvall Instruments, USA) by adding 210 μl
of 60% OptiPrep/0.1% Triton X-100. The cell lysates were covered
with 3.5 ml 30% OptiPrep in TXNE buffer and 300 μl TXNE buffer.
After spinning in the ultracentrifuge (4 h, 200,000 × g, 4°C)
(Sorvall/Kendro, USA), six fractions were collected from the top.
The proteins in fractions 1–2 were taken as the lipid raft
fractions.
Immunoprecipitation and western blot
analysis
SGC-7901 and MGC-803 cells were solubilized in 1%
Triton lysis buffer (1% Triton X-100, 50 mM Tris-Cl pH 7.4, 150 mM
NaCl, 10 mM EDTA, 100 mM NaF, 1 mM Na3VO4, 1
mM PMSF, 2 μg/ml aprotinin). For immunoprecipitation, cell lysates
were mixed with Src or caveolin-1 antibody and 20 ml of protein
G-sepharose beads (Cell Signaling Technology) at 4°C overnight. The
immunoprecipitated proteins were eluted by heat treatment at 100°C
for 5 min with 2X sampling buffer. For the preparation of total
cell lysates, cell lysate proteins were separated by sodium dodecyl
sulfate-polyacrylamide gel electrophoresis and electrophoretically
transferred to a nitrocellulose membrane (Immoblin-P; Millipore,
Bedford, MA, USA). The membranes were blocked with 5% skim milk in
TBST buffer (10 mM Tris-Cl pH 7.4, 150 mM NaCl, 0.1% Tween-20) at
room temperature and incubated overnight at 4°C with the indicated
primary antibodies. After the appropriate secondary antibodies were
added at room temperature, the proteins were detected with enhanced
chemiluminescence reagent (SuperSignal Western Pico
Chemiluminescent Substrate; Pierce, USA) and visualized with the
Electrophoresis Gel Imaging Analysis System (DNR Bio-Imaging
Systems, Israel).
In situ proximity ligation assay
In situ proximity ligation assay (PLA) was
carried out to detect the interactions of Src, EGFR or caveolin-1.
We used Duolink in situ PLA (Olink Bioscience). In the
assay, oligonucleotide-conjugated ‘PLA probe’ antibodies are
directed against primary antibodies for Src, EGFR or caveolin-1.
Annealing of the ‘PLA probes’ occurs when Src, EGFR or caveolin-1
are in close proximity, which initiates the amplification of repeat
sequences recognized by the fluorescently labeled oligonucleotide
probe. SGC-7901 and MGC-803 cells were seeded on 8-well chamber
slides and stimulated as described above. Then cells were washed
with ice-cold phosphate-buffered saline (PBS) and fixed with 4%
paraformaldehyde PBS for 30 min at room temperature. The cells were
permeabilized with 0.2% Triton X-100 PBS for 10 min at room
temperature. After that, the cells were assayed with Duolink in
situ PLA. Anti-Src Ab and anti-EGFR Ab, or anti-Src Ab and
anti-caveolin-1 Ab or anti-EGFR Ab and anti-caveolin-1 Ab were used
as primary antibodies. PLA probe anti-mouse Minus and PLA probe
anti-rabbit Plus were used as 2 PLA probes. For detection, Duolink
detection kit 563 was used. ProLong Gold Antifade Reagent with
4′,6-diamidino-2-phenylindole (DAPI; Molecular Probes) was used as
the mounting medium. The specimens were observed using a
fluorescence microscope (FV1000S-SIM/IX81; Olympus, Japan).
Statistical analysis
Data were confirmed in three independent
experiments, and are expressed as the means ± standard deviation
(SD). Differences between groups were compared using the Student’s
t-test. SPSS 18.0 computer software was used for statistical
analysis, and P<0.05 was considered to indicate a statistically
significant result.
Results
TRAIL induces EGFR and Src translocation
into lipid rafts and activates EGFR, Src and caveolin-1 in gastric
cancer cells
Since caveolin-1 is an essential structural
constituent of lipid rafts in the cell membrane (13), we isolated membrane lipid rafts by
ultracentrifugation and located them using caveolin-1 (fractions 1
and 2; Fig. 1A). TRAIL was used at
100 ng/ml in all experiments (14),
and was shown to induce EGFR and Src translocation into
caveolin-1-enriched lipid raft fractions (1 and 2) in SGC-7901 and
MGC-803 cells (Fig. 1A). TRAIL also
induced the activation of Src, caveolin-1, EGFR and downstream
Akt/ERK in a temporal manner (Fig.
1B). These data suggest that Src and caveolin-1 are involved in
EGFR activation in TRAIL-resistant gastric cancer cells.
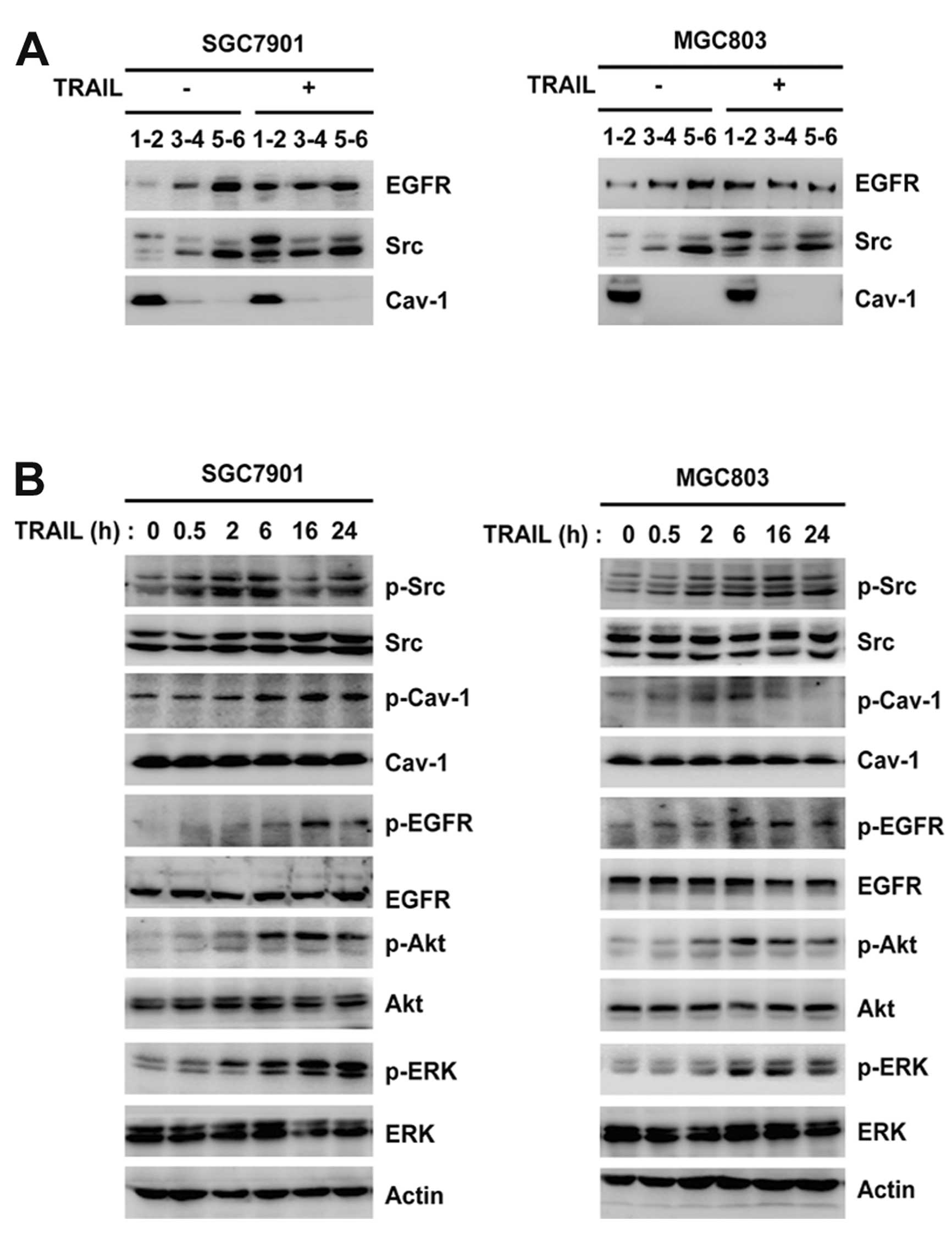 | Figure 1TRAIL induces the translocation of
EGFR and Src into lipid rafts and the activation of EGFR, Src and
caveolin-1 in gastric cancer cells. (A) SGC-7901 and MGC-803 cells
were treated with 100 ng/ml TRAIL for 6 h, and were then lysed and
fractionated by ultracentrifuge. Locations of lipid rafts
(fractions 1–2) were determined using caveolin-1. The distribution
of EGFR and Src was analyzed by western blotting. (B) SGC-7901 and
MGC-803 cells were incubated with 100 ng/ml TRAIL for 0.5, 2, 6, 16
and 24 h. The phosphorylation of Src, caveolin-1, EGFR, Akt and ERK
was analyzed by western blotting. TRAIL, tumor necrosis
factor-related apoptosis-inducing ligand; EGFR, epidermal growth
factor receptor. |
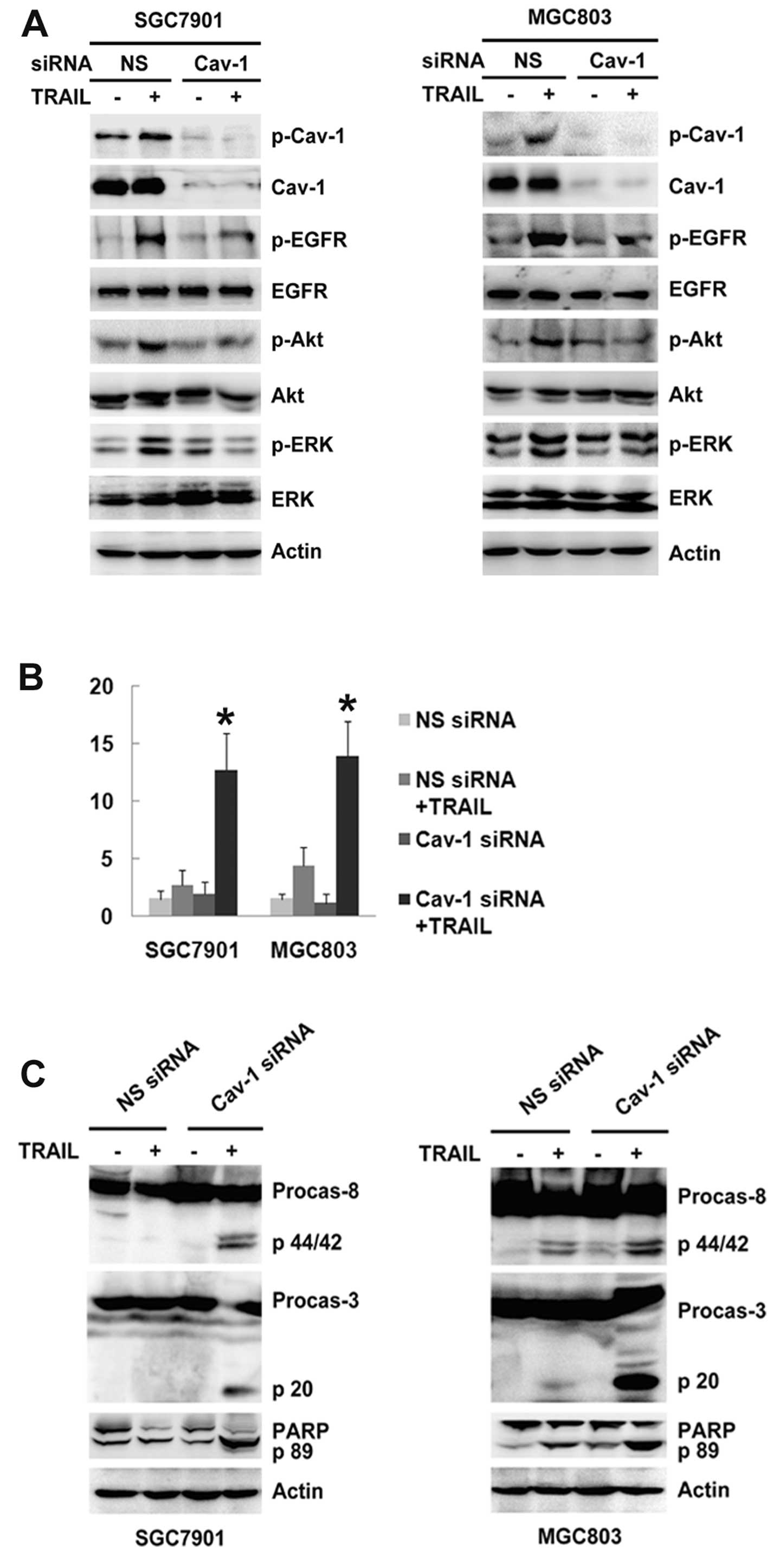
Knockdown of caveolin-1 enhances TRAIL
sensitivity via inhibition of EGFR activation in gastric cancer
cells
To further delineate the role of caveolin-1 in TRAIL
resistance, SGC-7901 and MGC-803 cells were transiently transfected
with caveolin-1 siRNA for 48 h prior to TRAIL treatment for 6 h. As
shown in Fig. 2A, knockdown of
caveolin-1 partially prevented EGFR and downstream Akt/ERK
activation in the SGC-7901 and MGC-803 cells. Caveolin-1 siRNA had
no significant effect on apoptosis (Fig. 2B). However, preincubation with
caveolin-1 siRNA for 48 h followed by TRAIL treatment for 24 h
resulted in a significant increase in apoptosis compared with TRAIL
treatment alone in the SGC-7901 and MGC-803 cells (12.70±3.24 vs.
2.70±1.34% and 13.90±3.07 vs. 4.40±1.57%, respectively, P<0.05;
Fig. 2B). Caspase-3 and -8
activation and poly(ADP-ribose) polymerase (PARP) cleavage were
also detected (Fig. 2C). These
results suggest that caveolin-1 promotes EGFR activation and
antagonizes TRAIL-induced apoptosis in gastric cancer cells.
Inhibition of Src activation increases
TRAIL sensitivity by the inhibition of caveolin-1 and EGFR
activation in gastric cancer cells
To investigate the effect of Src on
caveolin-1-regulated EGFR activation, SGC-7901 and MGC-803 cells
were pretreated with 10 μM PP2 (a specific Src inhibitor) for 1 h
before TRAIL treatment for 6 h. As shown in Fig. 3A, this not only inhibited
TRAIL-induced phosphorylation of caveolin-1, but also prevented
EGFR and downstream Akt/ERK activation. However, knockdown of
caveolin-1 did not influence Src activation (Fig. 3B). PP2 alone had little effect on
cell apoptosis after TRAIL treatment for 24 h (Fig. 3C). However, compared with TRAIL
alone, preincubation with PP2 significantly increased the rate of
cell apoptosis induced by TRAIL in the SGC-7901 and MGC-803 cells
(17.32±3.43 vs. 4.72±2.15% and 18.74±3.14 vs. 6.10±2.83%,
respectively, P<0.05; Fig. 3C).
Meanwhile, cleavage of caspase-8 and -3, and PARP was detected in
cells treated with TRAIL and PP2 (Fig.
3D). These results suggest that Src promotes
caveolin-1-regulated EGFR activation and antagonizes TRAIL-induced
apoptosis in gastric cancer cells.
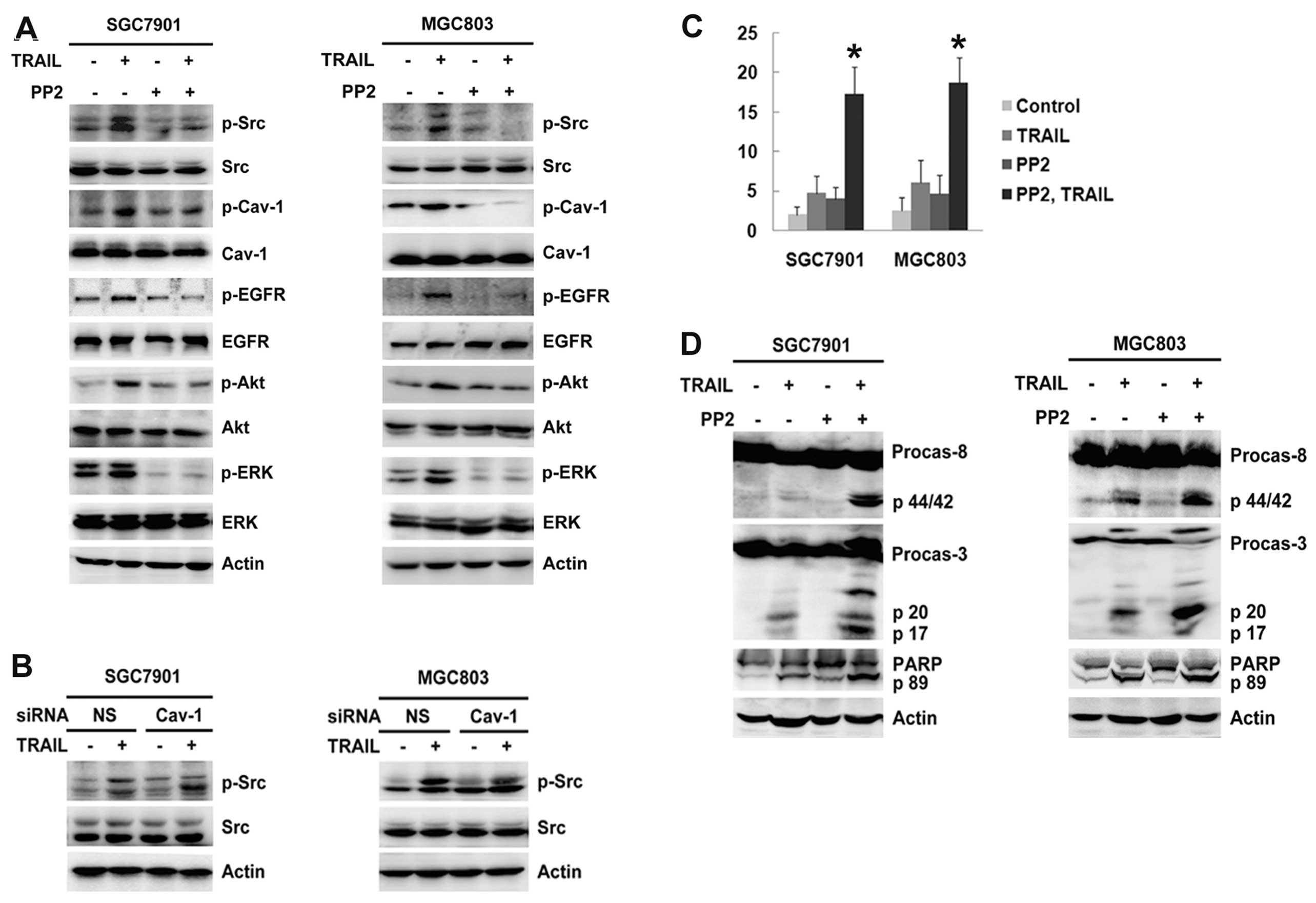 | Figure 3Inhibition of Src activation increases
TRAIL sensitivity via the inhibition of caveolin-1 and EGFR
activation in gastric cancer cells. (A) SGC-7901 and MGC-803 cells
were preincubated with 10 μM Src inhibitor PP2 for 1 h, and then
treated with 100 ng/ml TRAIL for 6 h. The phosphorylation of Src,
caveolin-1, EGFR, Akt and ERK was analyzed by western blotting. (B)
SGC-7901 and MGC-803 cells were transiently transfected with
caveolin-1 siRNA for 48 h, followed by 100 ng/ml TRAIL for 6 h. The
phosphorylation of Src was analyzed by western blotting. (C)
SGC-7901 and MGC-803 cells were incubated with 10 μM PP2 for 1 h,
and then treated with 100 ng/ml TRAIL for 24 h. Cell apoptosis was
quantified using flow cytometry. *P<0.05, incubation
with PP2 and TRAIL vs. with TRAIL alone. (D) SGC-7901 and MGC-803
cells were treated under similar conditions to those in A. The
expression of caspase-3, and -8, and PARP was analyzed by western
blotting. TRAIL, tumor necrosis factor-related apoptosis-inducing
ligand; EGFR, epidermal growth factor receptor; siRNA, small
interfering RNA; PARP, poly(ADP-ribose) polymerase. |
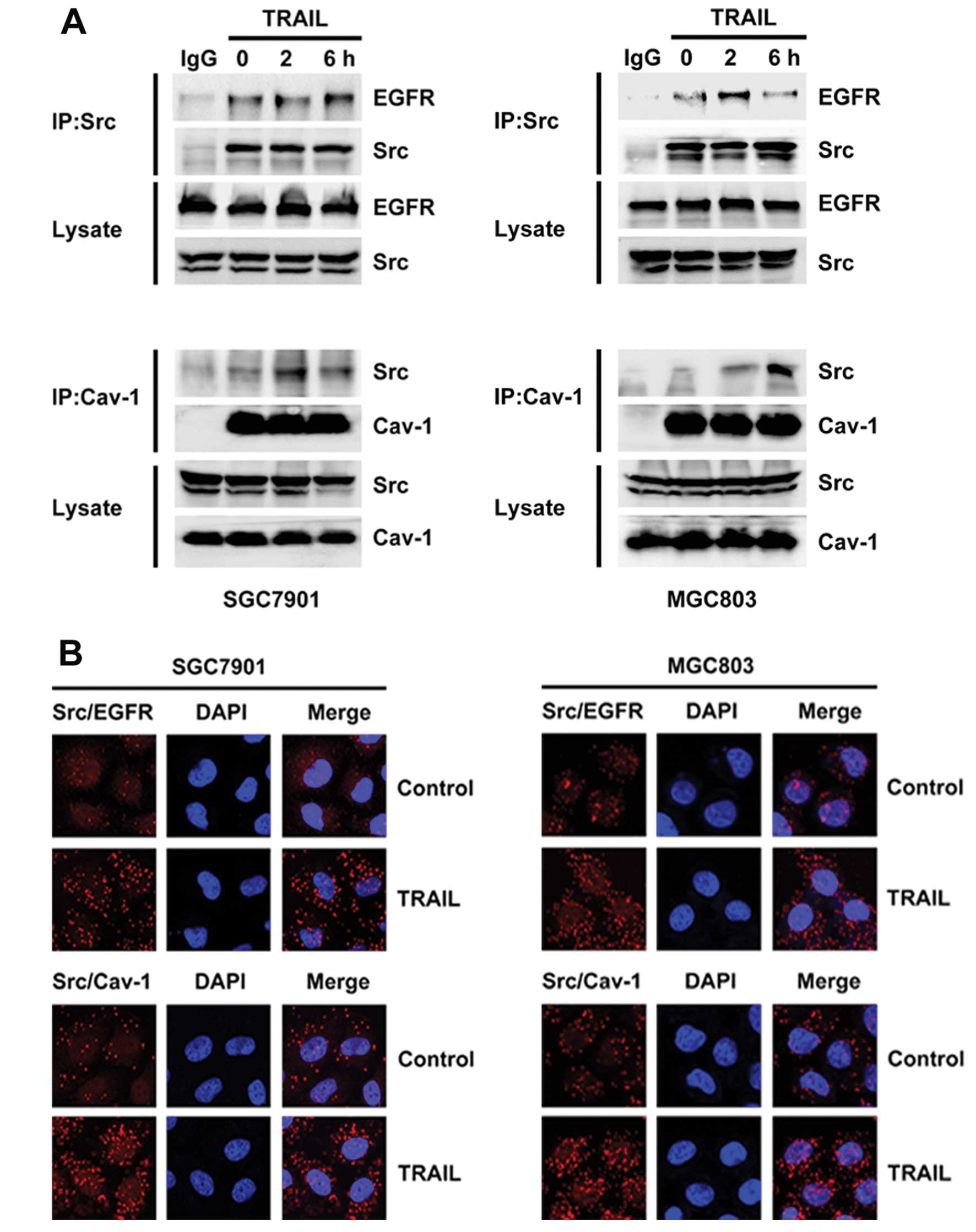
TRAIL induces the interaction of Src with
both EGFR and caveolin-1 in gastric cancer cells
To identify the regulatory mechanism of Src and
caveolin-1 in EGFR activation, we investigated the interaction of
Src with EGFR and caveolin-1. The Src-EGFR and Src-caveolin-1
interactions were detected by immunoprecipitation (Fig. 4A). In the untreated SGC-7901 and
MGC-803 cells, Src interacted weakly with both EGFR and caveolin-1
(Fig. 4A). However, the interaction
of Src with both EGFR and caveolin-1 was strengthened after TRAIL
treatment (Fig. 4A). The Src-EGFR
and Src-caveolin-1 interactions were also detected by the proximity
ligation assay Duolink (Fig. 4B).
This technique confirmed the promotion of these interactions by
TRAIL (Fig. 4B), although no
interaction between EGFR and caveolin-1 was detected (data not
shown). These data indicate that Src is an important mediator
between EGFR and caveolin-1 in TRAIL-induced EGFR activation.
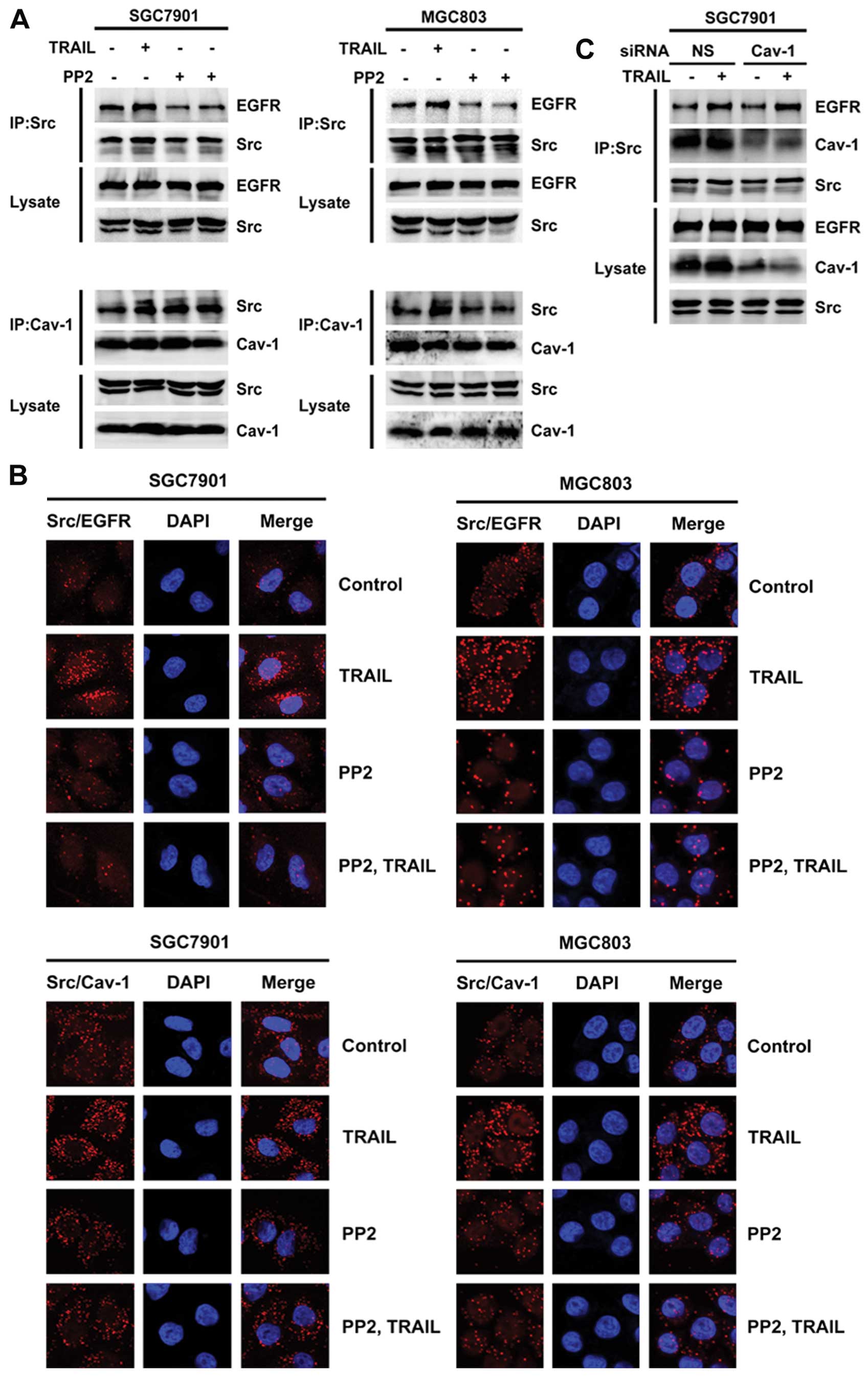
Inhibition of Src activation prevents the
interaction of Src with both EGFR and caveolin-1 in gastric cancer
cells
As shown in Fig. 5A,
the Src inhibitor PP2 alone did not influence the interaction of
Src with either EGFR or caveolin-1. However, compared with TRAIL
alone, a combination of PP2 and TRAIL partially reversed the
interaction of Src with both EGFR and caveolin-1. As before, the
Src-EGFR and Src-caveolin-1 interactions were also detected by
Duolink (Fig. 5B), and PP2 was
shown to partially inhibit these TRAIL-induced interactions
(Fig. 5B). However, knockdown of
caveolin-1 did not influence the interaction of Src and EGFR
(Fig. 5C). These results indicate
that the TRAIL-induced Src activation triggered the interaction of
Src with both EGFR and caveolin-1.
Discussion
Caveolin-1-enriched lipid rafts provide an important
platform for triggering receptor tyrosine kinase activation
(15). For example, stretch-induced
EGFR transactivation in mesangial cells requires caveolin-1
phosphorylation (9), while
caveolin-1 also drives the internalization and nuclear transport of
EGFR in the regulation of DNA repair in irradiated cells (16). However, it has not been determined
whether caveolin-1 regulates TRAIL-induced EGFR activation. In the
present study, TRAIL was shown to induce EGFR translocation into
lipid rafts, leading to the activation of EGFR and caveolin-1 in
gastric cancer SGC-7901 and MGC-803 cells. Knockdown of caveolin-1
partially inhibited EGFR activation and enhanced TRAIL sensitivity,
indicating that caveolin-1 promotes TRAIL-induced EGFR activation
and antagonizes TRAIL-triggered apoptosis.
EGFR activation is also regulated by the
non-receptor tyrosine kinase Src, and the Src family kinase
regulates caveolin-1 activation. Src activation was shown to be
essential for ATP-dependent activation of the sheddase ADAM17,
which is responsible for EGFR activation in airway epithelial cells
(17), while a blockade of Src
signaling significantly increased TRAIL-induced cell death in
lapatinib (a dual EGFR and ErbB-2 tyrosine kinase
inhibitor)-resistant breast cancer cells (18). It was also reported that
Src-dependent caveolin-1 phosphorylation was a critical event in
transforming growth factor (TGF)-β1-initiated SMAD2/3 activation
(19), and Src-mediated shedding of
TGF-α was found to be critical for TRAIL-induced EGFR activation in
colorectal cancer cells (6). In the
present study, TRAIL activated Src and induced its translocation
into lipid rafts. PP2 inhibition of Src activation reduced the
activation of caveolin-1 and EGFR, and increased TRAIL sensitivity.
However, knockdown of caveolin-1 did not influence Src activation.
Thus, it is conceivable that Src promotes caveolin-1-regulated EGFR
activation and antagonizes TRAIL-induced gastric cancer cell
apoptosis.
To identify the regulatory mechanism of Src and
caveolin-1 involved in EGFR activation, the interaction of these
molecules was investigated. Irradiation was shown to promote the
stability of Src in complex with phosphorylated EGFR in bronchial
carcinoma cells (20), while the
phosphorylation of caveolin-1 by Src and subsequent binding of the
Src SH2 domain to caveolin-1 led to the accumulation of activated
Src in focal adhesions of COS-7 and REF52 cells (21). Moreover, in the angiotensin
II-induced epithelial-to-mesenchymal transition, angiotensin II
caused EGFR to associate with phosphorylated caveolin-1 and
increased its phosphorylation in the caveolin-1-enriched fraction
(22). However, the role of Src and
caveolin-1 in the regulation of TRAIL-induced EGFR activation is
not clear. In the present study, TRAIL was found to induce the
interaction of Src with both EGFR and caveolin-1, but no
interaction between EGFR and caveolin-1 was detected. The
inhibition of Src activation partially reversed the interaction of
Src with both EGFR and caveolin-1, while, conversely, knockdown of
caveolin-1 had no effect on the Src-EGFR interaction. Thus, TRAIL
induced EGFR activation at least partially by the interaction of
both Src-EGFR and Src-caveolin-1.
In summary, we showed that Src activated EGFR
through the interaction of both Src-EGFR and Src-caveolin-1, and
then antagonized TRAIL-induced apoptosis in gastric cancer cells.
It can therefore be hypothesized that the Src-caveolin-1-EGFR
activation pathway is involved in the TRAIL resistance mechanism of
certain cell types.
Acknowledgements
This study was supported by Chinese National
Foundation of National Sciences grants (nos. 81201802, 81172369,
81172198, 81372546 and 81372485), the Specialized Research Fund for
the Doctoral Program of Higher Education (nos. 20102104120008 and
20112104110005), the National Science and Technology Major Project
(no. 2013ZX09303002) and the Science and Technology Plan Project of
Liaoning Province (no. 2011404013-1). The authors thank Jian Gao
and Lu Yao (Experiment Technology Center of China Medical
University) for kindly providing technical support.
Abbreviations:
|
EGFR
|
epidermal growth factor receptor
|
|
PARP
|
poly(ADP-ribose) polymerase
|
|
TGF
|
transforming growth factor
|
|
TRAIL
|
TNF-related apoptosis-inducing
ligand
|
|
VEGF
|
vascular endothelial growth factor
|
References
|
1
|
Cervantes A, Roda D, Tarazona N, Roselló S
and Pérez-Fidalgo JA: Current questions for the treatment of
advanced gastric cancer. Cancer Treat Rev. 39:60–67. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Schaefer U, Voloshanenko O, Willen D and
Walczak H: TRAIL: a multifunctional cytokine. Front Biosci.
12:3813–3824. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Morizot A, Mérino D, Lalaoui N, et al:
Chemotherapy overcomes TRAIL-R4-mediated TRAIL resistance at the
DISC level. Cell Death Differ. 18:700–711. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Xu L, Qu X, Luo Y, et al: Epirubicin
enhances TRAIL-induced apoptosis in gastric cancer cells by
promoting death receptor clustering in lipid rafts. Mol Med Rep.
4:407–411. 2011.
|
|
5
|
Cho IR, Koh SS, Min HJ, et al: Reovirus
infection induces apoptosis of TRAIL-resistant gastric cancer cells
by down-regulation of Akt activation. Int J Oncol. 36:1023–1030.
2010.PubMed/NCBI
|
|
6
|
Van Schaeybroeck S, Kelly DM, Kyula J,
Stokesberry S, Fennell DA, Johnston PG and Longley DB: Src and
ADAM-17-mediated shedding of transforming growth factor-α is a
mechanism of acute resistance to TRAIL. Cancer Res. 68:8312–8321.
2008.PubMed/NCBI
|
|
7
|
Xu L, Zhang Y, Liu J, et al:
TRAIL-activated EGFR by Cbl-b-regulated EGFR redistribution in
lipid rafts antagonises TRAIL-induced apoptosis in gastric cancer
cells. Eur J Cancer. 48:3288–3299. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang S, Yu S, Shi W, Ge L, Yu X, Fan J and
Zhang J: Curcumin inhibits the migration and invasion of mouse
hepatoma Hca-F cells through down-regulating caveolin-1 expression
and epidermal growth factor receptor signaling. IUBMB Life.
63:775–782. 2011.
|
|
9
|
Zhang B, Peng F, Wu D, Ingram AJ, Gao B
and Krepinsky JC: Caveolin-1 phosphorylation is required for
stretch-induced EGFR and Akt activation in mesangial cells. Cell
Signal. 19:1690–1700. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang J, Kalyankrishna S, Wislez M, et al:
SRC-family kinases are activated in non-small cell lung cancer and
promote the survival of epidermal growth factor receptor-dependent
cell lines. Am J Pathol. 170:366–376. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wu T, Zhang B, Ye F and Xiao Z: A
potential role for caveolin-1 in VEGF-induced fibronectin
upregulation in mesangial cells: involvement of VEGFR2 and Src. Am
J Physiol Renal Physiol. 304:F820–F830. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen Z, Bakhshi FR, Shajahan AN, et al:
Nitric oxide-dependent Src activation and resultant caveolin-1
phosphorylation promote eNOS/caveolin-1 binding and eNOS
inhibition. Mol Biol Cell. 23:1388–1398. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang Y, Roche O, Xu C, et al: Hypoxia
promotes ligand-independent EGF receptor signaling via
hypoxia-inducible factor-mediated upregulation of caveolin-1. Proc
Natl Acad Sci USA. 109:4892–4897. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xu L, Qu X, Zhang Y, et al: Oxaliplatin
enhances TRAIL-induced apoptosis in gastric cancer cells by
CBL-regulated death receptor redistribution in lipid rafts. FEBS
Lett. 583:943–948. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Irwin ME, Mueller KL, Bohin N, Ge Y and
Boerner JL: Lipid raft localization of EGFR alters the response of
cancer cells to the EGFR tyrosine kinase inhibitor gefitinib. J
Cell Physiol. 226:2316–2328. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Dittmann K, Mayer C, Kehlbach R and
Rodemann HP: Radiation-induced caveolin-1 associated EGFR
internalization is linked with nuclear EGFR transport and
activation of DNA-PK. Mol Cancer. 7:692008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sham D, Wesley UV, Hristova M and van der
Vliet A: ATP-mediated transactivation of the epidermal growth
factor receptor in airway epithelial cells involves DUOX1-dependent
oxidation of Src and ADAM17. PLoS One. 8:e543912013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
De Luca A, D’Alessio A, Gallo M, Maiello
MR, Bode AM and Normanno N: Src and CXCR4 are involved in the
invasiveness of breast cancer cells with acquired resistance to
lapatinib. Cell Cycle. 13:148–156. 2014.PubMed/NCBI
|
|
19
|
Samarakoon R, Chitnis SS, Higgins SP,
Higgins CE, Krepinsky JC and Higgins PJ: Redox-induced Src
kinase and caveolin-1 signaling in TGF-β1-initiated SMAD2/3
activation and PAI-1 expression. PLoS One. 6:e228962011.PubMed/NCBI
|
|
20
|
Dittmann K, Mayer C, Kehlbach R, Rothmund
MC and Peter Rodemann H: Radiation-induced lipid peroxidation
activates src kinase and triggers nuclear EGFR transport. Radiother
Oncol. 92:379–382. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gottlieb-Abraham E, Shvartsman DE,
Donaldson JC, Ehrlich M, Gutman O, Martin GS and Henis YI:
Src-mediated caveolin-1 phosphorylation affects the targeting of
active Src to specific membrane sites. Mol Biol Cell. 24:3881–3895.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen J, Chen JK and Harris RC: Angiotensin
II induces epithelial-to-mesenchymal transition in renal epithelial
cells through reactive oxygen species/Src/caveolin-mediated
activation of an epidermal growth factor receptor-extracellular
signal-regulated kinase signaling pathway. Mol Cell Biol.
32:981–991. 2012. View Article : Google Scholar
|