Introduction
Autophagy is a self-eating mechanism by which cells
degrade and renew cellular molecules and organelles. It can be
classified as macroautophagy, microautophagy and chaperone-mediated
autophagy. Macroautophagy (referred to as autophagy) involves the
formation of double membraned vesicles (autophagosomes) containing
materials to be degraded, such as unnecessary proteins and damaged
or aged organelles. Autophagosome then fuse with lysosomes, break
down the cargos and recycle the useful molecules. Autophagy at a
basal level is essential for cellular clearance of unwanted
materials. Dysregulated autophagy leads to dysfunction and even
death of cells, due to the cytotoxicity of the accumulated
materials. The role of autophagy in cancer biology depends on the
context. In normal cells, autophagy contributes to the prevention
of carcinogenesis by eliminating the molecules that may induce or
promote cancer formation. On the other hand, established cancer
cells utilize autophagy to survive some adverse conditions, such as
hypoxia, low nutrition and oxidative stress.
Oleanolic acid [(3β)-3-hydroxyolean-12-en-28-oic
acid], a natural pentacyclic triterpenoid that exists widely in
various type of plants, is believed to be one of the most important
compounds with bioactivity in vegetables and medicinal herbs
(1). Its bioactivity includes
anti-inflammation, antivirus, hepatoprotection and antitumor
(1). OA displayed no significant
cytotoxicity to normal cells (1).
Owing to these characteristics, researchers are making efforts to
improve its activity by developing its derivatives (2,3).
Considering that cancer is a severe risk to human health, the
antitumor activity of OA has attracted increasingly more attention
from researchers. As the major antitumor mechanism, OA induces
apoptotic death in a wide range of cancer cells (4,5). OA
stimulation leads to an increase in the ratio between the
proapoptotic protein Bax and the anti-apoptotic protein Bcl-2, in
the mitochondrial membrane, which promotes Bax to oligomerize. The
permeability of mitochondrial membrane was consequently elevated,
which facilitates the release of cytochrome c into
cytoplasm. The caspase family proteases were, therefore, cleaved
and activated.
However, some cells were found to be resistant to
OA-induced apoptosis. The mechanisms underlying the resistance
remain unknown. In the present study, we found that OA was able to
induce autophagy in cancer cells. Autophagy impaired the
pro-apoptotic effect of OA on cancer cells. Blocking autophagy
restored the sensitivity of cancer cells to OA-induced apoptotic
death.
Materials and methods
Cell lines
A549, MCF-7, U2OS, BXPC3, PANC-1 and PC-3 cells were
purchased from the American Type Culture Collection (ATCC;
Manassas, VA, USA) and cultured in the recommended media
supplemented with 10% fetal bovine serum (FBS; Invitrogen), 4 mM
glutamine, 100 U/ml penicillin. All the cells were cultured in a
humidified 5% CO2 atmosphere at 37°C.
Chemical reagents
Oleanolic acid (OA; O5504) and autophagy inhibitor
3-methyladenine (3-MA; M9281) were purchased from Sigma-Aldrich.
Atg7-targeting siRNAs (siATG7) (#6604) and control siRNA
(siControl) (#6568) were purchased from Cell Signaling Technology
(Beverly, MA, USA). Inhibitors that selectively blocked JNK
(SP600125, #8177) were purchased from Cell Signaling Technology.
mTOR activator MHY1485 (2 μM), Millipore, #5.00554.0001.
pGFP-LC3 transfection
Cancer cells were seeded on the top of coverslips in
a 24-well plate. The cells were transfected with the plasmid,
pEGFP-LC3, using Lipofectamine 2000 (Invitrogen) following the
manufacturer’s instruction. Forty-eight hours after transfection,
cells were treated with 100 μg/ml of OA for 12 h. The cells were
then fixed with 4% polyoxymethylene and observed under an Olympus
FluoView™ FV1000 confocal microscope (Olympus, Hamburg, Germany).
Cells with five GFP-LC3 puncta or more were counted per 100 random
cells in each group.
Immunoblotting assay
Proteins of human tissues or cultured cells were
harvested with M-PER Mammalian Protein Extraction reagent (Thermo
Fisher Scientific, Rockford, IL, USA), separated using
polyacrylamide gel electrophoresis and transferred onto 0.45-μm
nitrocellulose membranes. The membranes were blocked with 5%
fat-free dry milk in phosphate-buffered saline (PBS) and incubated
with primary antibodies at 4°C. Overnight, the membrane was
incubated with corresponding secondary antibodies and visualized
with SuperSignal West Dura Extended Duration Substrate (Thermo
Fisher Scientific).
The involved primary antibodies in the present study
included: LC3B, #3868; Beclin-1, #3495; cleaved PARP, CST, #5625;
cleaved caspase-3 CST, #9664; cleaved caspase-9, CST, #9501;
cytochrome c, GST, #11940; phospho-JNK (Thr183/Tyr185),
#4668; JNK, #9258; phospho-mTOR (Ser2448), #5536; mTOR, #2983. The
intensity of blots in each figure was determined with ImageJ
software.
Apoptotic rate determination by flow
cytometry
The rates of apoptotic cells were evaluated by flow
cytometric analysis. Briefly, cells were fixed in cold 70% ethanol
for 1 h, and were then incubated with 10 mg/ml RNase A at 37°C for
1 h. The cells were stained with propidium iodide (PI; 200 mg/ml),
and immediately subjected to flow cytometric analysis (BD FACSAria
II; BD Biosciences). Cells (1×105) were counted for each
sample.
Cell viability assay
Cancer cells (1×104) were seeded in
96-well plates. Fifty milliliters of 3-(4,
5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) (1
mg/ml) was used at the indicated time-points. After 4 h, MTT was
removed and 150 ml of dimethyl sulfoxide (DMSO) was then added. The
absorbance at 570 nm was evaluated on a model 550 microplate reader
(Bio-Rad Laboratories, Hercules, CA, USA) with a reference
wavelength of 655 nm.
Statistical analysis
The experiments, with the exception of the
immunoblot assays, were performed at least three times. All values
are reported as means ± SD, and compared at a given time-point by
unpaired, two-tailed t-test. Differences were considered
statistically significant when *P<0.05 and
**P<0.01.
Results
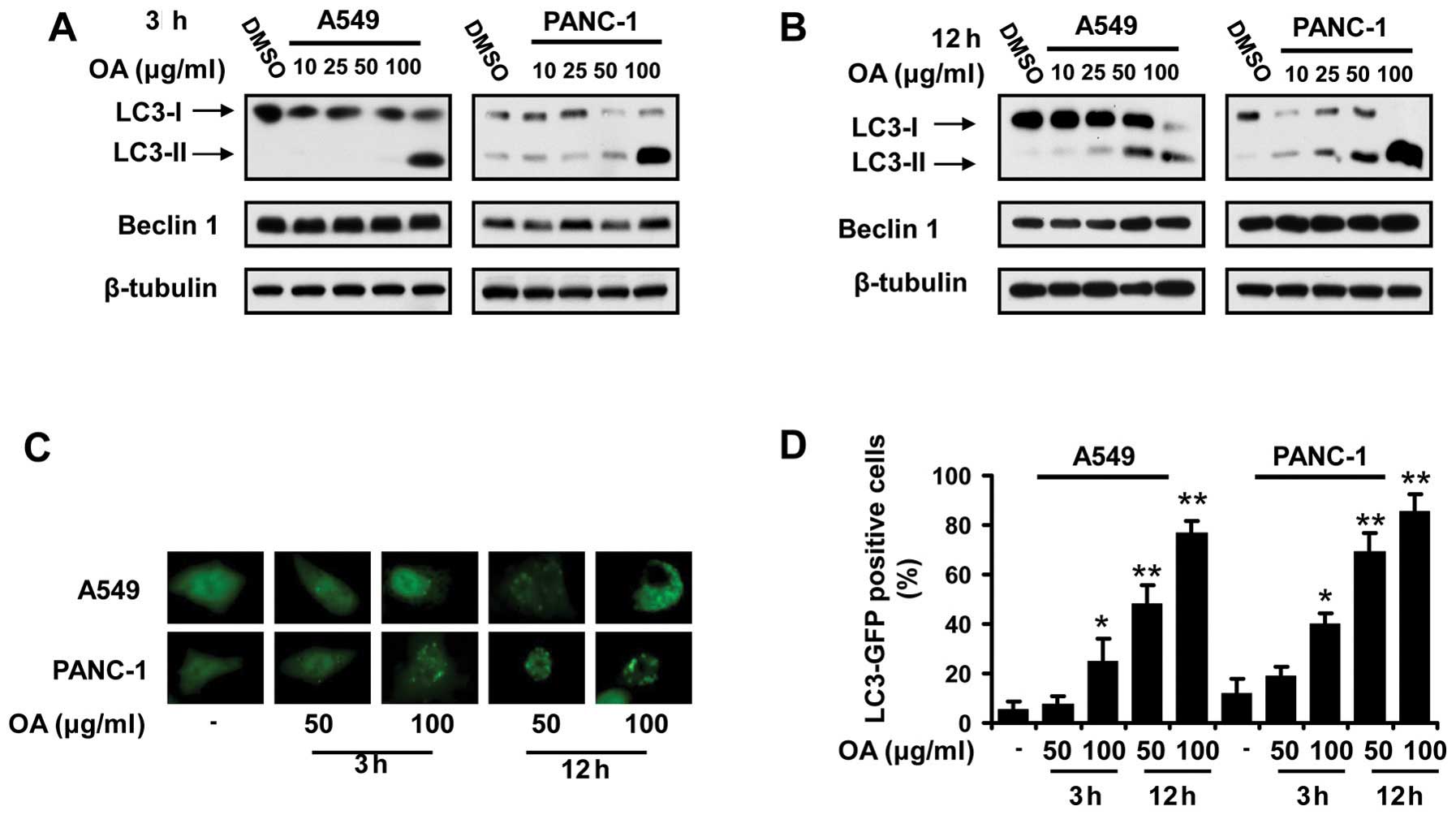
OA induces autophagy in a wide range of
cancer cells in a dose- and time-dependent manner
Autophagy has been reported to be initiated in
neoplastic cells by natural and synthetic compounds (6,7).
First, we investigated if OA induced autophagy in cancer cells by
immunoblot analysis of light chain 3 (LC3) isoforms. OA was found
to increase the expression level of LC3-II in various cancer cell
lines. This effect occurred in a dose- and time-dependent manner
(Fig. 1A and 1B). Autophagic event
was further confirmed by observing the fluorescence in the cancer
cells transfected with pGFP-LC3, a plasmid expressing a fusion
protein that tracks the location of LC3. The GFP-LC3 puncta were
frequently seen in the cancer cells treated with OA, whereas the
control cells had a relatively homogeneous LC3 expression pattern
(Fig. 1C).
Blocking autophagy potentiates the
apoptotic death of cancer cells treated with OA
Considering the role of autophagy in apoptotic
pathways in cancer cells, we aimed to determine the effect of
autophagic events in OA-induced apoptosis in cancer cells. A small
molecule inhibitor of autophagy, 3-MA, was used to abolish the
occurrence of autophagy induced by OA. Cell viability was shown to
be further reduced when OA and 3-MA was added to the media
(Fig. 2A). Immunoblot assays
revealed that 3-MA suppression of autophagy was able to enhance the
cleavage and activation and caspase family proteases as well as
PARP, which are all the biomarkers of apoptosis induction (Fig. 2B). Consistently, higher
sub-G0/G1 population was detected in cancer
cells co-treated with OA and 3-MA, in comparison with the cells
treated with OA alone (Fig. 2C). A
siRNA silencing Atg7 expression (siATG7) was also employed to
suppress autophagic event in OA-incubated cancer cells. Similar to
the results obtained from 3-MA-based assays, siATG7 also
synergistically decreased the survival of cancer cells, potentiated
the activation of apoptotic pathways, and heightened the percentage
of sub-G0/G1 population with OA, in cancer
cells (Fig. 2D–F). These data
supported the hypothesis that autophagy induced by OA exerted a
suppressive effect on apoptotic pathways in cancer cells.
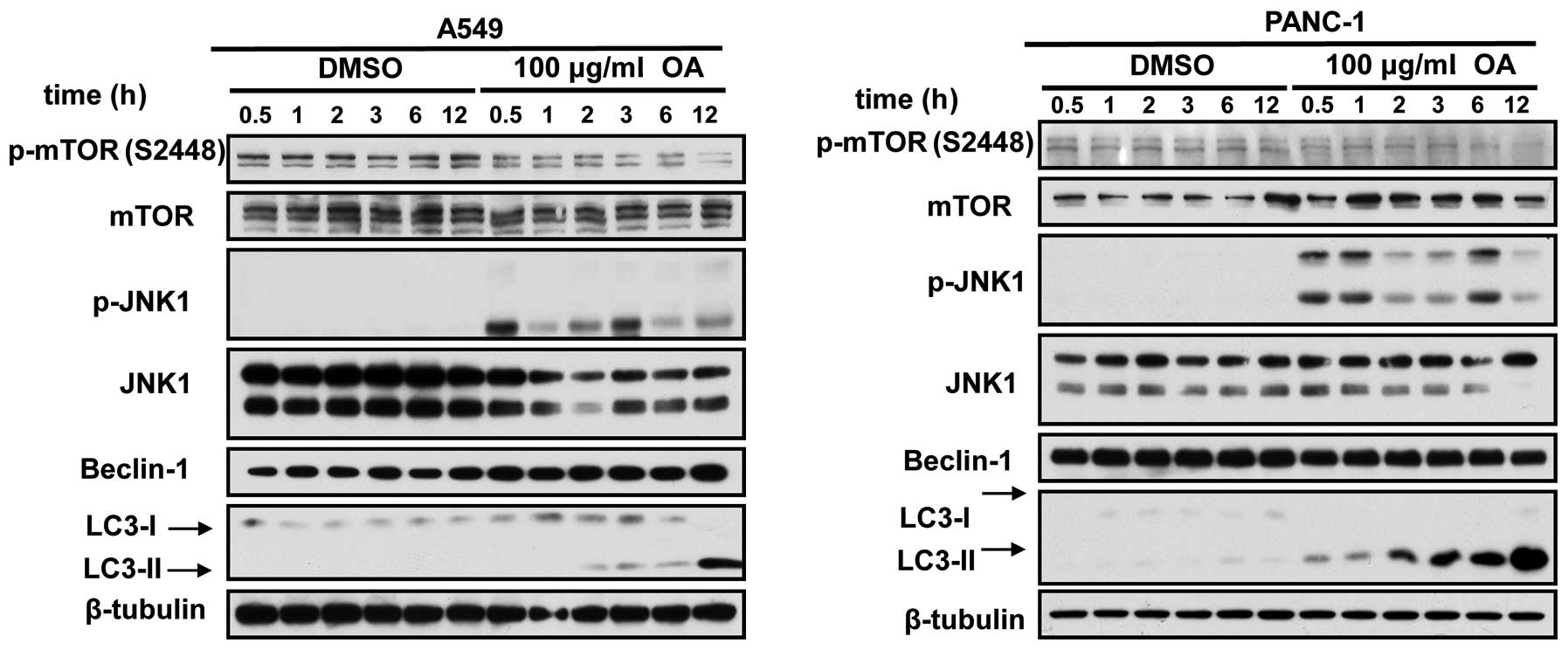
OA induces JNK activation and mTOR
suppression in cancer cells
To study the mechanisms by which OA induced
autophagy, we focused on c-Jun N-terminal kinase 1 (JNK) and mTOR
signaling cascades. JNK is closely associated with the initiation
of autophagic events in a variety of cell types. Activated JNK-1
phosphorylates N-terminal region of Bcl-2 and triggers its
dissociation from Beclin 1, the most potent inducer of autophagic
pathway (8). The released Beclin 1
subsequently induced the assembly of autophagosomes. OA was found
to induce the JNK activation in a range of cancer cells, in a
time-dependent manner (Fig. 3).
mTOR has been well documented as the major suppressor of cellular
autophagy. Immunoblot assays also indicated that mTOR
phosphorylation was diminished in cancer cells under the treatment
of OA (Fig. 3).
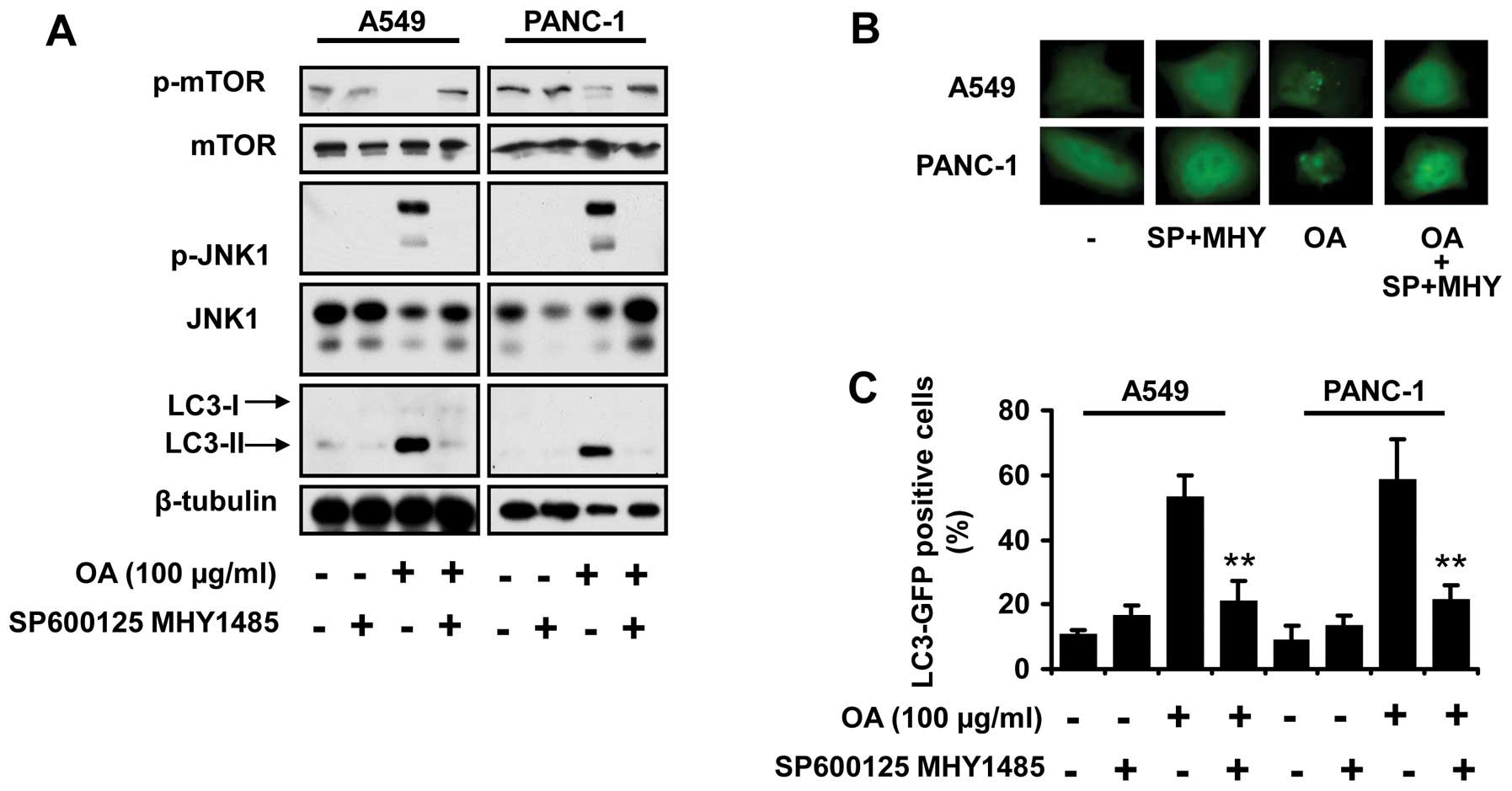
The alteration in JNK and mTOR activity
mediates OA-induced autophagy
Having established that JNK activation and mTOR
suppression was caused by OA treatment in cancer cells, we
subsequently investigated whether the changes in their activity
accounted for the occurrence of autophagic events. SP600125 and
MHY1485 were used to inhibit JNK activation and reactivate mTOR,
respectively. OA-related autophagy was abrogated by these two
compounds, indicated by the reduction in LC3-II expression
(Fig. 4A), as well as the amount of
LC3 puncta (Fig. 4B and C).
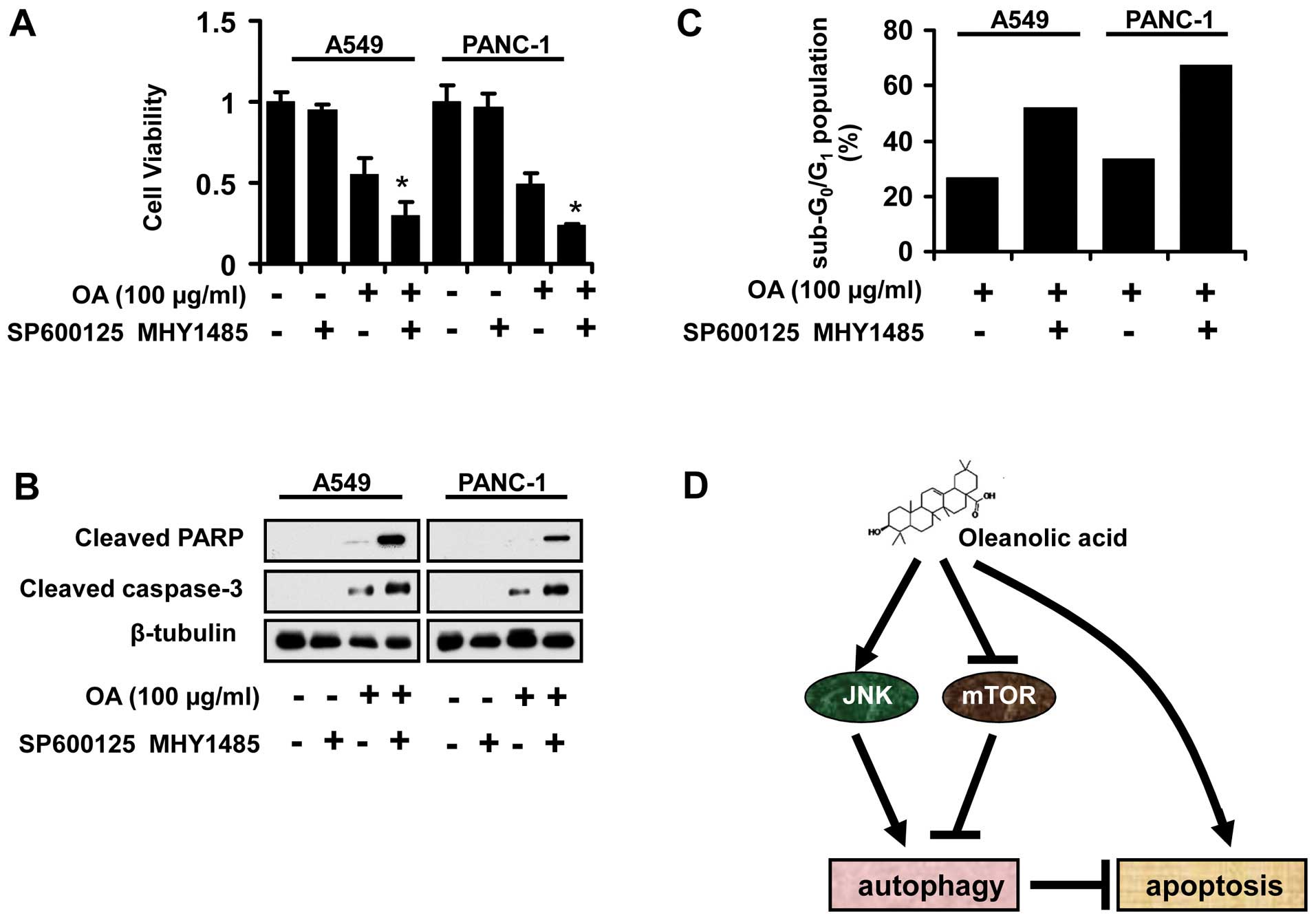
JNK suppression and mTOR activation
enhance the apoptosis induced by OA
To further confirm the requirement of JNK
suppression and mTOR activation in the effect of OA on cancer
cells, we employed SP600125 and MHY1485 to change the active status
of these two pathways. The data revealed that SP600125 and MHY1485
potentiated the antitumor activity of OA on A549 and PANC-1 cells
(Fig. 5A). Furthermore, the
activation of apoptosis-related proteins was also detected by
immunoblot assays. SP600125 and MHY1485 led to increased expression
of cleaved caspase-3 and PARP in cancer cells co-treated with OA
(Fig. 5B). Also, the percentage of
sub-G0/G1 was higher in cancer cells when the
JNK and mTOR pathways were suppressed and activated, respectively
(Fig. 5C).
Discussion
Although OA treatment has been demonstrated to
trigger the onset of many cellular processes, such as apoptosis
(5), cell cycle arrest (9) and impaired motility (10), it remains unknown if OA can induce
autophagic events in cancer cells. Autophagy is a biological
process widely employed by a wide range of cells to defend
themselves against adverse stimuli, including the treatment of
antitumor compounds. In fact, it has been reported that autophagy
can be induced by many bioactive natural compounds in cancer cells
(6). However, the role of induced
autophagy in the antitumor potency of natural compounds appears to
vary between different compounds. Curcumin (11), α-eleostearic acid (12), evodiamine (13) and resveratrol (14), have been shown to result in cell
death by inducing autophagy. In contrast, cancer cells employ
autophagy as a protective mechanism against the antitumor activity
of isobavachalcone (15) and
quercetin (16). In the present
study, we experimentally indicated that OA was able to induce
autophagy in cancer cells, which can enhance the resistance of
cancer cells to apoptosis. To the best of our knowledge, this is
the first study to provide evidence that autophagy can be triggered
by OA stimulation. Furthermore, OA-induced autophagy may account
for the resistance against apoptosis of some types of cancer
cells.
The pathways that initiate or maintain the
activation of autophagy have been well studied. Among them, JNK and
mTOR pathways are the most important (17). It has also been well documented that
these two pathways mediated the onset of autophagy in cancer cells
under the treatment of some compounds (11,18).
Previously, OA or its derivative was demonstrated to activate the
JNK pathway both in normal cells (19) and malignant cells (20), as well as the suppression of mTOR
pathway (21). The outcome of these
alterations was mainly limited to apoptosis and cell cycle arrest.
It remains unexplored if other effects result from JNK activation
and mTOR suppression by OA. The present study revealed, for the
first time, that OA’s effect on JNK and mTOR activation lead to
autophagy in cancer cells.
In conclusion, OA induced protective autophagy in
cancer cells by JNK activation and mTOR inhibition, which enhanced
the resistance of cancer cells to apoptotic events. Our findings
contribute to a better understanding of OA’s action mode.
Furthermore, we provided evidence that suppression of autophagy is
an effective strategy to enhance antitumor activity of OA.
Acknowledgements
The present study was supported by the National
Innovative Drug Development Projects (2014ZX-09102043-001). The
study was also supported in part by the National Natural Science
Foundation of China (81302906, 81273550 and 41306157).
References
|
1
|
Pollier J and Goossens A: Oleanolic acid.
Phytochemistry. 77:10–15. 2012. View Article : Google Scholar
|
|
2
|
Liby KT, Yore MM and Sporn MB:
Triterpenoids and rexinoids as multifunctional agents for the
prevention and treatment of cancer. Nat Rev Cancer. 7:357–369.
2007. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Petronelli A, Pannitteri G and Testa U:
Triterpenoids as new promising anticancer drugs. Anticancer Drugs.
20:880–892. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chakravarti B, Maurya R, Siddiqui JA, et
al: In vitro anti-breast cancer activity of ethanolic extract of
Wrightia tomentosa: role of pro-apoptotic effects of
oleanolic acid and urosolic acid. J Ethnopharmacol. 142:72–79.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wei J, Liu M, Liu H, et al: Oleanolic acid
arrests cell cycle and induces apoptosis via ROS-mediated
mitochondrial depolarization and lysosomal membrane
permeabilization in human pancreatic cancer cells. J Appl Toxicol.
33:756–765. 2012. View
Article : Google Scholar
|
|
6
|
Zhang X, Chen LX, Ouyang L, Cheng Y and
Liu B: Plant natural compounds: targeting pathways of autophagy as
anti-cancer therapeutic agents. Cell Prolif. 45:466–476. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Livesey KM, Tang D, Zeh HJ and Lotze MT:
Autophagy inhibition in combination cancer treatment. Curr Opin
Investig Drugs. 10:1269–1279. 2009.PubMed/NCBI
|
|
8
|
Wei Y, Pattingre S, Sinha S, Bassik M and
Levine B: JNK1-mediated phosphorylation of Bcl-2 regulates
starvation-induced autophagy. Mol Cell. 30:678–688. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang X, Bai H, Zhang X, et al: Inhibitory
effect of oleanolic acid on hepatocellular carcinoma via
ERK-p53-mediated cell cycle arrest and mitochondrial-dependent
apoptosis. Carcinogenesis. 34:1323–1330. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Guo G, Yao W, Zhang Q and Bo Y: Oleanolic
acid suppresses migration and invasion of malignant glioma cells by
inactivating MAPK/ERK signaling pathway. PLoS One. 8:e720792013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Aoki H, Takada Y, Kondo S, Sawaya R,
Aggarwal BB and Kondo Y: Evidence that curcumin suppresses the
growth of malignant gliomas in vitro and in vivo through induction
of autophagy: role of Akt and extracellular signal-regulated kinase
signaling pathways. Mol Pharmacol. 72:29–39. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Eom JM, Seo MJ, Baek JY, et al:
Alpha-eleostearic acid induces autophagy-dependent cell death
through targeting AKT/mTOR and ERK1/2 signal together with the
generation of reactive oxygen species. Biochem Biophys Res Commun.
391:903–908. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rasul A, Yu B, Zhong L, Khan M, Yang H and
Ma T: Cytotoxic effect of evodiamine in SGC-7901 human gastric
adenocarcinoma cells via simultaneous induction of apoptosis and
autophagy. Oncol Rep. 27:1481–1487. 2012.PubMed/NCBI
|
|
14
|
Filippi-Chiela EC, Villodre ES, Zamin LL
and Lenz G: Autophagy interplay with apoptosis and cell cycle
regulation in the growth inhibiting effect of resveratrol in glioma
cells. PLoS One. 6:e208492011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhao S, Ma CM, Liu CX, et al: Autophagy
inhibition enhances isobavachalcone-induced cell death in multiple
myeloma cells. Int J Mol Med. 30:939–944. 2012.PubMed/NCBI
|
|
16
|
Wang K, Liu R, Li J, et al: Quercetin
induces protective autophagy in gastric cancer cells: involvement
of Akt-mTOR- and hypoxia-induced factor 1α-mediated signaling.
Autophagy. 7:966–978. 2011.PubMed/NCBI
|
|
17
|
Mehrpour M, Esclatine A, Beau I and
Codogno P: Overview of macroautophagy regulation in mammalian
cells. Cell Res. 20:748–762. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xavier CP, Lima CF, Pedro DF, Wilson JM,
Kristiansen K and Pereira-Wilson C: Ursolic acid induces cell death
and modulates autophagy through JNK pathway in apoptosis-resistant
colorectal cancer cells. J Nutr Biochem. 24:706–712. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang X, Ye XL, Liu R, et al: Antioxidant
activities of oleanolic acid in vitro: possible role of Nrf2 and
MAP kinases. Chem Biol Interact. 184:328–337. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zou W, Yue P, Khuri FR and Sun SY:
Coupling of endoplasmic reticulum stress to CDDO-Me-induced
up-regulation of death receptor 5 via a CHOP-dependent mechanism
involving JNK activation. Cancer Res. 68:7484–7492. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhou R, Zhang Z, Zhao L, et al: Inhibition
of mTOR signaling by oleanolic acid contributes to its anti-tumor
activity in osteosarcoma cells. J Orthop Res. 29:846–852. 2011.
View Article : Google Scholar : PubMed/NCBI
|