Introduction
Central nervous system leukemia (CNSL) is a
relatively common and severe form of leukemia relapse in clinical
practice. The incidence of CNSL in children with acute
non-lymphocytic leukemia (ANLL) is 6–29% (1,2). In
the last two decades, considerable progress has been achieved in
determining the molecular mechanisms of leukemia, and
nonobese-severe immunodeficiency (NOD/SCID) mice have been widely
used in hematology research. However, studies on extramedullary
infiltration in leukemia are limited, especially studies on this
mechanism in CNSL. Furthermore, reports on CNSL are rare, and there
is a lack of a convenient and practical ANLL-CNSL model.
Acute monocytic leukemia (FAB classification of M5b)
SHI-1 cells have a characteristic t(6;11) (q27;q23) chromosomal
translocation and express the MLL/AF6 fusion gene product; they
have a defect in the p53 gene and their tumorigenicity is high when
inoculated subcutaneously in nude mice (3,4). In
the present study, we constructed a CNSL model in NOD/SCID mice
through intracerebroventricular injection of SHI-1 cells into the
lateral ventricle, and in this way established an efficient and
reproducible model of CNSL that could be used for further study on
the mechanism of extramedullary infiltration of leukemia cells and
on possible therapies for CNSL.
Materials and methods
Five to six-week-old NOD/SCID mice were purchased
from the Experimental Animal Center of Zhongshan University and
adapted to the environment in a specific pathogen-free (SPF)
laminar flow cabinet for 3 days. They had free access to food and
water except on the day the intraventricular injection was
performed. Food, bedding and other substances the mice came into
direct contact with underwent irradiation sterilization or
high-pressure treatment before use.
Each mouse in the 2 CNSL groups underwent
inoculation of 2×104 SHI-1 cells in the right lateral
ventricle under sterile conditions. Mice in the control group had
intraventricular inoculation of saline under the same conditions.
There were 21 NOD/SCID mice, divided into 3 groups, an experimental
group, a survival group and a control group. In the experimental
group (n=10), 2 mice were sacrificed at day 3 after lateral
ventricle injection in order to examine the spread of the SHI-1
cells and 2 mice were sacrificed for the same purpose each week
after that until all mice died. Timely treatment was given to
moribund mice. In the survival group (n=8), the times after the
lateral ventricle injection of leukemic cells that death occurred
were recorded. In the control group (n=3), 10 μl saline was
injected into the lateral ventricle, and the health and survival of
the mice were monitored.
Cells from the human monocytic leukemia cell line
SHI-1 (a gift from Dr Suning Chen at the Jiangsu Institute of
Hematology, Suzhou, China) were cultured in Iscove’s modified
Dulbecco’s medium (IMDM) (Gibco, Carlsbad, CA, USA) and 10% fetal
bovine serum (FBS; Hangzhou Evergreen Biological Co., Ltd.,
Hangzhou, China) at 37°C in an incubator with 5% CO2 and
saturated humidity. Cells in the logarithmic growth phase were
collected and inoculated into the right ventricle of the mice.
For lateral ventricle injection of SHI-1 cells, mice
were deeply anesthetized with 5% chloral hydrate (50 μl/10 g), and
fixed on a mouse brain stereotaxic apparatus to expose the anterior
fontanelle. Right ventricle positioning was adopted and a
microsyringe was used to aspirate 10 μl cell suspension and insert
it into the right lateral ventricle perpendicularly at a location 2
mm posterior to the anterior fontanelle and 1.0 mm lateral to the
sagittal suture. The depth of the needle insertion was 2 mm below
the skull surface. The cell suspension was injected at a constant
flow rate of 1 μl/min, and a total of 10 μl of cell suspension
(2×104 cells) was injected. The duration of the
injection was 10 min. The needle was then left at the injection
site for 5 min before withdrawal. The needle was rotated while
retreating, and the wound was disinfected and the skin sutured
after the needle was withdrawn.
Histopathological examination was performed on
animals that died in the survival group and in the experimental
group, on the 2 mice that were anesthetized and sacrificed each
week after inoculation until all mice in this group had died. The
following procedures were performed on animals when death or
near-death occurred. Various tissues and organs including heart,
liver, spleen, lung, kidney and cervical lymph nodes were collected
and cryopreserved in liquid nitrogen. The femur was rinsed with
IMDM medium several times until the bone mass was transparent. The
flushing fluid was collected and centrifuged, and red blood cells
were removed using erythrocyte lysis buffer. After centrifugation,
cells were fully dissolved in TRIzol medium and stored in a
refrigerator at −80°C. Nested PCR was used to detect the MLL/AF6
fusion gene in tissues and bone marrow cells (5).
The cranial and spinal skin and subcutaneous tissue
were collected and fixed in 10% neutral formalin for 24–48 h. After
decalcification in decalcification solution containing 10%
hydrochloric acid for 24 h, slices were prepared. Five coronal
sections (6) were selected for each
head sample. The first section was perpendicular above the
bilateral eyes. The second section was perpendicular and 0.6 cm
from the first section. The third section was oblique and cut from
the posterior fontanel toward the lower left, which was 90 degrees
from the horizontal line. The fourth section was perpendicular and
0.2 cm outside the third section, and the fifth section was
perpendicular 0.2 cm outside the fourth section. All specimens were
routinely embedded in paraffin and cut into slices that were 2 μm
thick. H&E staining was performed. Immunohistochemical staining
was performed to detect expression of human leukocyte common
antigen (LCA) in order to assess leukemic cell infiltration.
According to the degree of leukemia cell
infiltration into the brain (7),
the CNSL was classified into three grades: Grade I, small amount of
leukemic cell infiltration was present in the subdural space, the
pia mater and arachnoid. Grade II, an extensive and large amount of
leukemic cell infiltration was present in the subdural space, the
pia mater, and arachnoid, but there was no brain parenchyma
infiltration. Grade III, a large amount of leukemic cell
infiltration was present in the subdural space, the pia mater and
arachnoid and was accompanied by brain parenchyma infiltration.
Data analysis
Continuous data of WBC counts and survival time are
presented as mean ± SD and median and range, respectively.
Categorical data for CNSL grading are presented as counts and
percentages. Survival curves were constructed by the Kaplan-Meier
method and compared by the log-rank test. Differences in peripheral
WBC counts between the survival and control groups were detected by
independent t-test each week from the 1st to the 5th week. The
association between CNSL grading and survival time (≤20 vs. >20)
was evaluated by Fisher’s exact test. Statistical analyses were
performed with SAS software version 9.2 (SAS Institute Inc., Cary,
NC, USA). A two-tailed P<0.05 indicated a statistically
significant result.
Results
Survival time
Three groups of NOD/SCID mice were included in the
present study: an experimental group (n=10), a survival group (n=8)
and a control group (n=3). Mice in the experimental group were
sacrificed at the following time-points: one mouse on day 3 and 2
mice on days 7, 14 and 21. Three mice in the experimental group
died naturally, one mouse on days 24, 26 and 31.
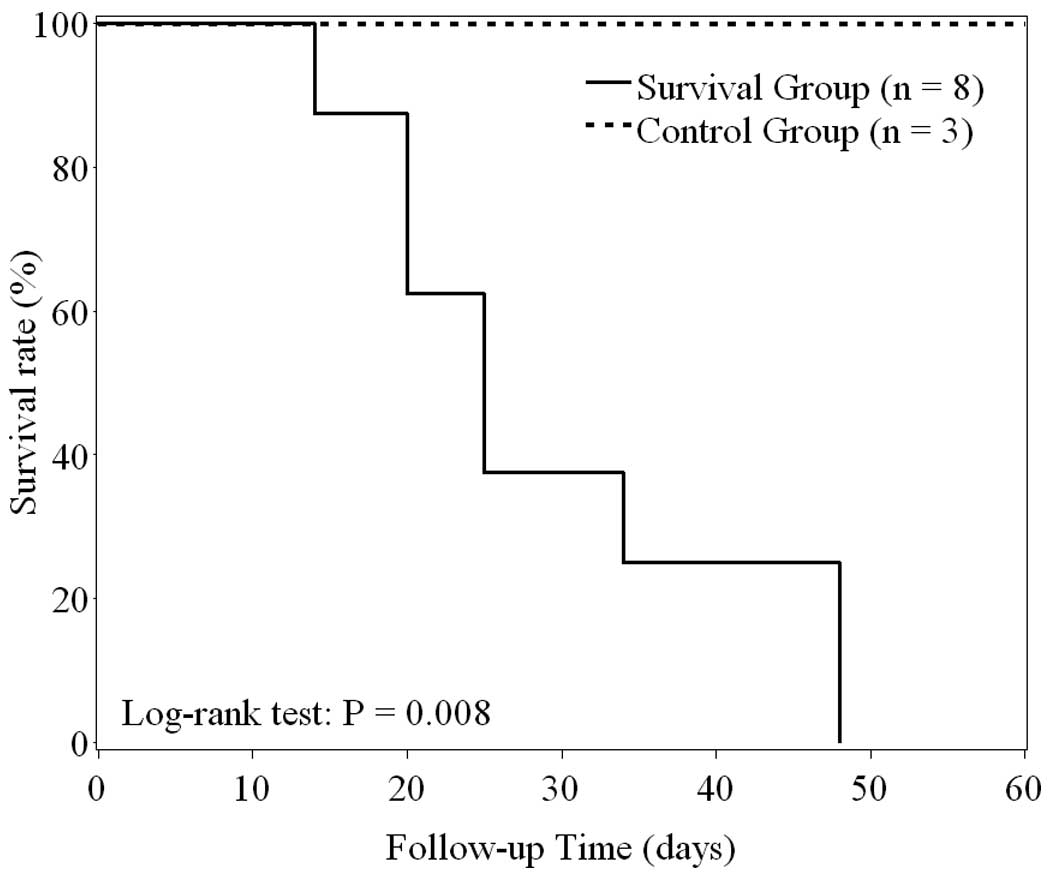
In the survival group, the median survival time was
25 days (range, 14–48 days). In the control group, all mice
survived more than 60 days. The Kaplan-Meier survival curves
(Fig. 1) indicated that survival
time was significantly shorter in the survival than in the control
group (log-rank test, P=0.008). The Kaplan-Meier estimate for the
30-day survival rate for mice in the survival group was 37.5% (95%
confidence interval, 8.7–67.4%).
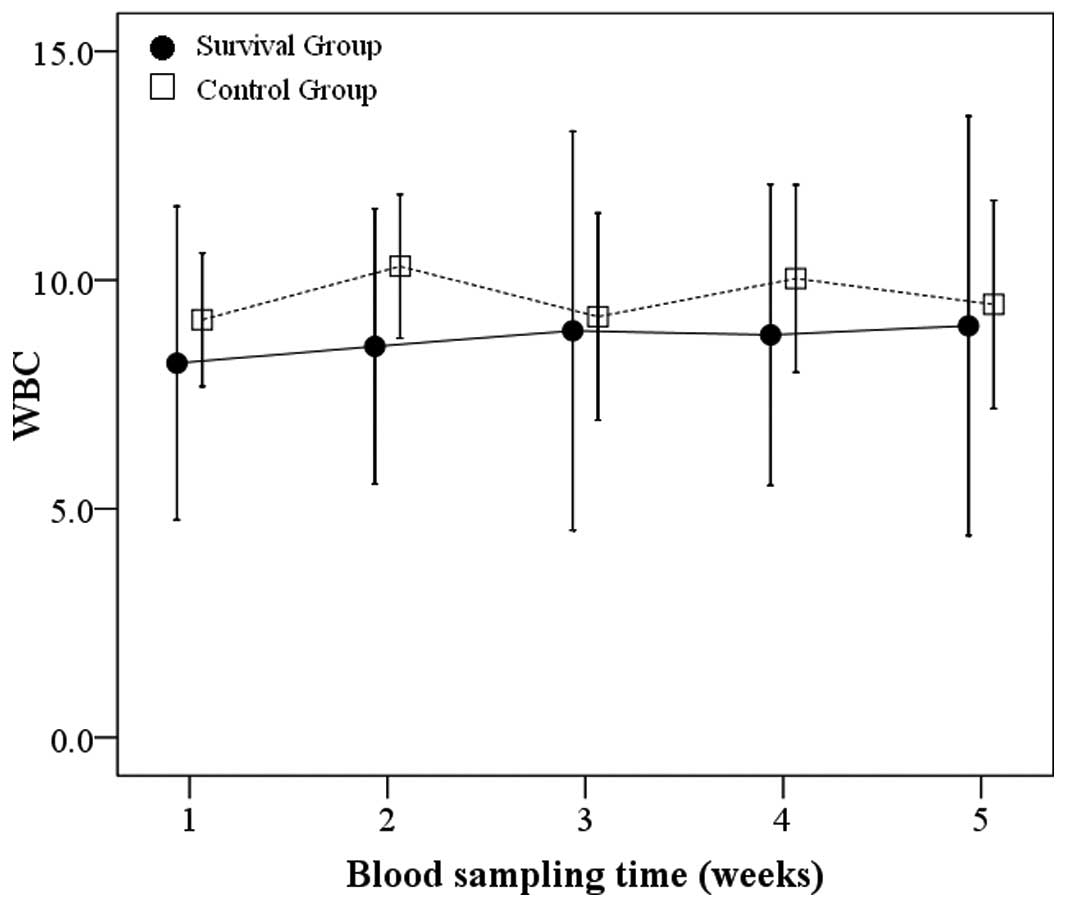
Changes in peripheral WBC counts over
time
Blood specimens were obtained from tail vein and WBC
counts were measured on the first day of each week for five weeks
for mice in both the survival and control groups. At each
corresponding time-point, the control group showed slightly higher
average WBC counts than the survival group (Fig. 2), but this difference did not reach
statistical significance (all P>0.05). No human leukemia cells
were observed in Wright-stained peripheral blood smears. Mice that
had not been inoculated with SHI-1 cells were generally in good
condition, and no deaths occurred in this group during the 60 days
after their saline inoculation.
CNSL grading
CNSL grading for mice in the experimental and
survival groups is shown in Table
I. The proportion of mice classified as CNSL grade III in the
experimental and survival groups was 50% (5/10) and 87.5% (7/8),
respectively. No significant association was found between CNSL
grading and survival time in either group (all P>0.05). With an
increase in survival time, CNSL symptoms were aggravated and the
degree of brain tissue infiltration worsened. The maximum survival
time was 48 days.
 | Table IThe distribution of CNSL grades in
NOD/SCID mice in experimental and survival groups, according to
survival time. |
Table I
The distribution of CNSL grades in
NOD/SCID mice in experimental and survival groups, according to
survival time.
| CNSL grading |
|---|
|
|
|---|
| I
n (%) | II
n (%) | III
n (%) |
|---|
| Experimental group
(n=10) | 2 (20) | 3 (30) | 5 (50) |
| Survival time ≤20
days | 2 (40) | 2 (40) | 1 (20) |
| Survival time >20
days | 0 (0) | 1 (20) | 4 (80) |
| Survival group
(n=8) | 0 (0) | 1 (12.5) | 7 (87.5) |
| Survival time ≤20
days | 0 (0) | 0 (0) | 3 (100) |
| Survival time >20
days | 0 (0) | 1 (20) | 4 (80) |
Autopsy of the mice did not show any tumor masses in
the abdominal cavity, head and face or torso. CNSL cell
infiltration was clearly visible and bone mass destruction in the
skull and vertebrae and a large amount of leukemia cell
infiltration into the vertebral bone marrow cavity were present.
The leukemia cells were able to break through the bone mass and
infiltrate outside or infiltrate the spinal parenchyma inward.
Pathological examination of the head tissue showed a small amount
of leukemic cell infiltration in the intracranial arachnoid and the
pia mater as early as week 1. By day 21, the degree of tumor cell
infiltration in brain tissue was severe, and a large amount of
leukemia cell infiltration in the subdural space, arachnoid and the
pia mater was accompanied by cerebral parenchymal infiltration.
Some lumpy masses formed, resulting in compression of the
surrounding tissue.
In the medullary cavity of the skull, a considerable
amount of leukemia cell infiltration and bone mass destruction were
seen, but no lumps had formed on the surface of the skull or in the
surrounding soft tissue. Intracranial infiltration occurred mostly
in the subdural space, arachnoid and pia mater, and was distributed
along the Virchow-Robin space of the blood vessels on the surface
of the pia mater. Leukemia cells had attached to the cerebrum,
cerebellum and on the surface of the fissures extensively. A large
number of leukemia cells were seen to infiltrate from the surface
of the cortex to the deep parenchyma in grade III mice. The normal
brain tissue structure was damaged, and focal perivascular
distribution of leukemia cells was seen in deeper brain tissue.
Infiltration of brain parenchyma was accompanied by leukemia cell
infiltration in the arachnoid, pia mater and surrounding skull and
skull destruction. At other places, such as the nasopharynx and
retrobulbar area, a considerable amount of leukemia cell
infiltration was found to form tumor-like tissues (Fig. 3).
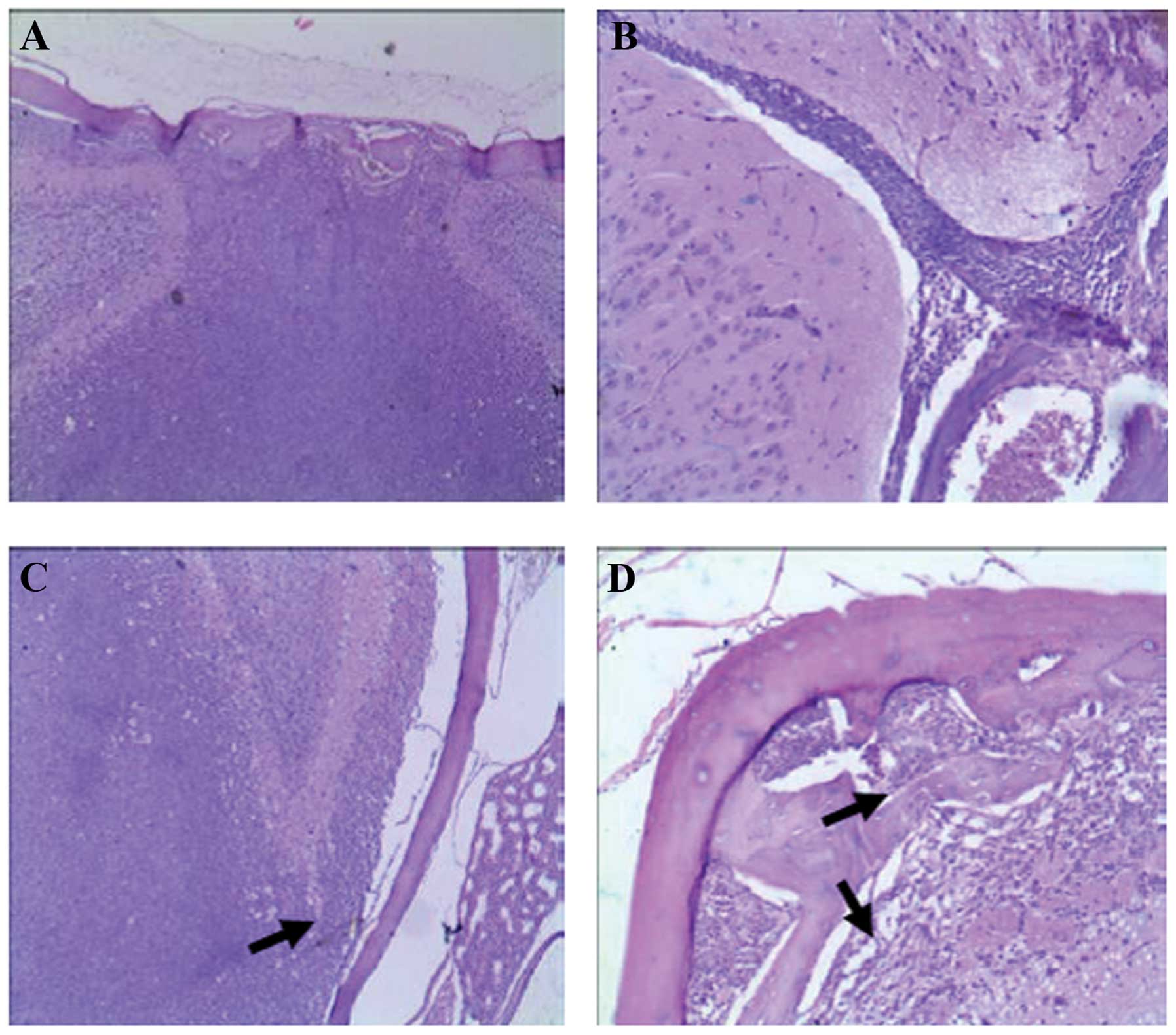 | Figure 3Acute leukemia cell infiltration in
the central nervous system. (A and D) A relatively large defect is
seen on the inner side of the skull (arrow), and a large number of
SHI-1 cells infiltrated the brain parenchyma; they invaded the
subdural space, pia mater, and arachnoid from the skull bone
marrow, and caused multiple defects on the inner side of the skull
(black arrow). (B and H) SHI-1 cells extended deeply along the
interhemispheric fissure (black arrow), and local infiltration was
seen in the pia mater (white arrow). (C) A considerable amount of
SHI-1 cell infiltration was present in the brain parenchyma. Cell
infiltration was observed in the pia mater, but no bone mass
destruction of the skull was found. (E and G) SHI-1 cells showed
invasive growth in the brain, infiltrating the parenchyma along the
Virchow-Robin space (black arrow). Multiple perivascular
infiltration foci were seen in the deep brain parenchyma. (F) A
considerable amount of SHI-1 cell infiltration was seen; the tumor
cells entered the sinuses, causing a nasal infiltration defect
(black arrow) and forming a local mass (red arrow). (I) A large
number of SHI-1 cells infiltrated the spinal parenchyma, showing
diffuse infiltration (black arrow). (J) SHI-1 cells infiltrated the
spinal parenchyma (black arrow) and spinal pia mater, invading the
vertebrae and resulting in local bone mass destruction (black
arrow). (K) SHI-1 cells extended deeply along the fissures (black
arrow) and infiltrated local spinal pia mater (black arrow). (L)
SHI-1 cells showed invasive growth in the spinal parenchyma (black
arrow) and focal perivascular infiltration (white arrow). (M and N)
Immunohistochemical staining showed that the cells infiltrating the
brain parenchyma (M) and vertebrae (N) were human LCA-positive
cells, indicating that the infiltration was caused by human
leukemia SHI-1 cells (black arrow). (A–L) H&E staining and (M
and N) immunohistochemical staining of human LCA. Original
magnification, A, C, F, H, I and J, ×100; E, L and N, ×400; K, D,
G, B and M, ×200. |
Expressions of S fusion gene MLL/AF6 in
various tissues of NOD/SCID mice
To further study the times of infiltration of
various organs, two mice were sacrificed each week after SHI-1 cell
inoculation. RT-PCR was used to detect SHI-1 cell-specific fusion
gene MLL/AF6 in various organs. No SHI-1 cell fusion gene was
amplified in any organ in weeks 1, 2 and 4. The SHI-1 cell fusion
gene was amplified in the lung, bone marrow and cervical lymph
nodes in one mouse in week 3, and the product was 280 bp.
Fig. 4 shows PCR
amplification results of MLL/AF6 of leukemia cells in NOD/SCID
mice. The MLL/AF6 fusion gene was amplified in specimens 11, 13 and
14, in lung, bone marrow and lymph node, respectively, of the same
mouse.
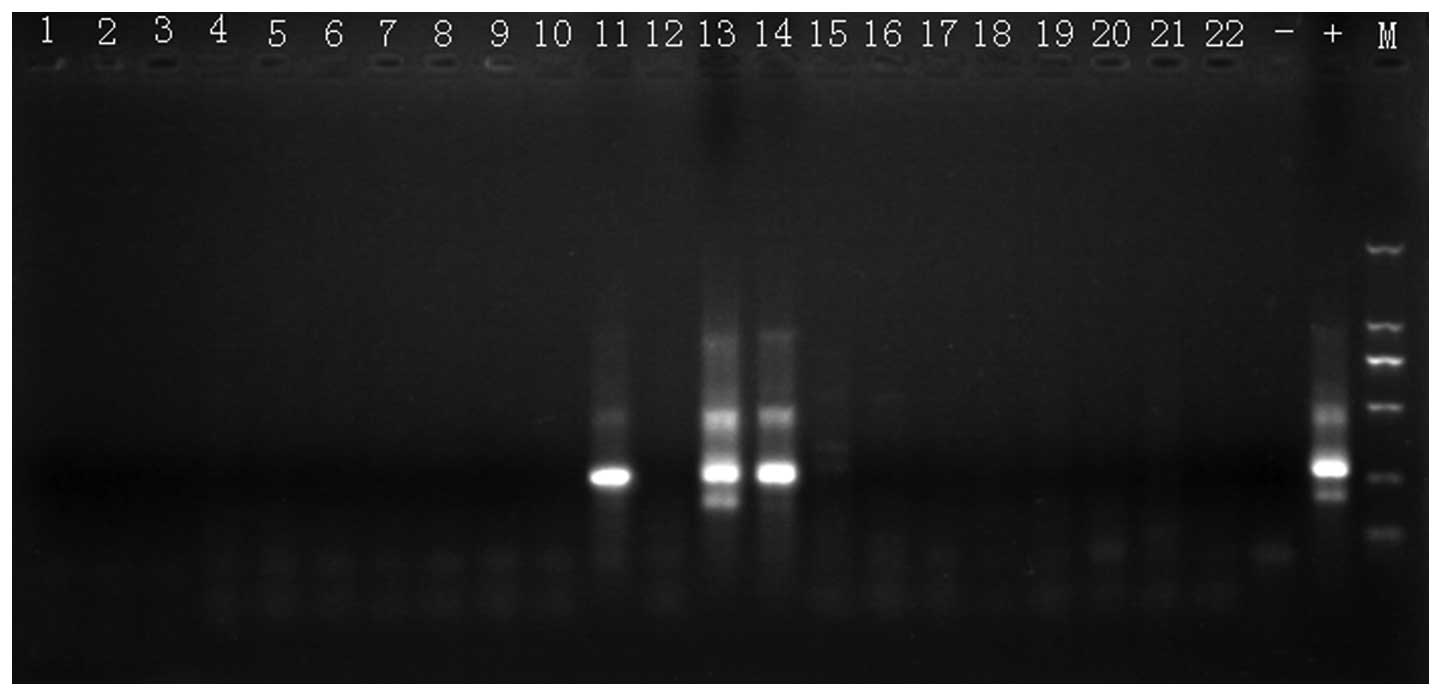 | Figure 4Exemplary PCR amplification of
NOD/SCID mouse leukemia cells MLL/AF6. Specimens were loaded in
order as lanes 1–7, 8–14 and 15–21 of three mice as heart, liver,
kidney, lung, spleen, bone marrow cells and lymph node,
respectively. Lane 22 is a heart sample from the fourth mouse.
Lanes 23 and 24 are negative control and positive control. The last
lane is molecular weight marker with sizes from top to bottom:
2,000, 1,000, 750, 500, 250 and 100 bp. Positive amplified human
MLL/AF6 fusion genes were observed in mouse lung (lane 11), bone
marrow cells (lane 13) and lymph nodes (lane 14). |
Discussion
In the present study, intracerebroventricular direct
injection of leukemic cells in mice was used to construct a model
of CNSL. As time after inoculation increased, various symptoms of
CNSL appeared, tumor cells gradually spread in the brain and spinal
cord and in one mouse, cells spread outside the central nervous
system into lymph node, lung and bone marrow. The median survival
time was 25 days.
Other mouse models of leukemia have used i.p., i.v.
and s.c. injections to introduce leukemic cells. Kamel-Reid et
al (8), in an early study using
i.p. injection of ALL cells into irradiated SCID mice, reported ALL
cells in bone marrow at 4 weeks, but did not see ALL cells in brain
or any animal deaths until 12 weeks after injection. Gunther et
al (9) using i.v. injection of
ALL cells into SCID mice, reported that ALL cells were seen in
brain and spinal cord by days 21–23, that all mice developed CNSL,
but that leukemic cells were not detected deep within the brain
parenchyma. In a study examining the characteristics of SHI-1
cells, (3) s.c. injection in mice
produced tumor masses after 9–19 days. In a study in which SHI-1
cells were injected i.v. into BALB/c nude mice that had been
subjected to splenectomy, cytoxan treatment and irradiation, cells
were found in the brain in the 3rd week, and median survival was 41
days.
The human monocytic leukemia cell line SHI-1
originated from mononuclear cells in the bone marrow of an AML-M5b
leukemia patient during relapse and is highly invasive (3). Western blot analysis is used to detect
EMMPRIN expression in a variety of leukemia cell lines, and all
types of leukemia cells have high expressions of EMMPRIN, while
SHI-1 has a fixed MLL/AF6 fusion gene. This individual
characteristic of the SHI-1 cell line is convenient for
experimental tracing. We injected human leukemia cells directly
into the lateral ventricle of NOD/SCID mice via a microsyringe to
induce direct exogenous CNSL and to establish a CNSL model. Our use
of intracerebroventricular injection not only excludes an influence
of leukemic cell infiltration in sites outside the bone marrow and
central nervous system, but also prevents the harmful effects of
the large doses of immunosuppressive agents or preoperative
radiation that are required for successful modeling through
peripheral injection. Therefore, our model simulates the
extramedullary infiltration characteristics of CNSL directly.
In our mouse model, a large amount of cell
infiltration was seen in brain tissue, and various degrees of
infiltration were also observed in the spinal cord. In addition to
non-specific manifestations including hair thinning, torpor and
decreased movement, arched back and reduced dietary intake, obvious
lower extremity weakness and paralysis occurred in some mice.
However, no convulsions or paralysis of both lower extremities was
observed, although convulsions and bilateral lower extremity
paralysis have been found in other reports (7,9). Our
results indicate that the CNSL model established via lateral
ventricle injection of SHI-1 has a more severe and direct impact on
brain tissue than models established using peripheral injection.
However, it has less or lagging impact on the spinal cord and the
spine, which results in more pronounced symptoms of brain
damage.
CNSL is a type of childhood leukemia that is
difficult to treat and has a high mortality rate. It can occur in
the early or late stage during chemotherapy (1). Currently, the chemotherapy regimens
advocate early prevention of CNSL; therefore, CNSL often occurs
after complete remission in clinical practice and may lead to or be
accompanied by bone marrow relapse, eventually leading to
chemotherapy failure. The pathogenesis of CNSL has not been fully
determined, and it is not fully understood through which pathway
tumor cells enter the central nervous system and what factors are
involved in tumor cell infiltration into the central nervous
system. In terms of the origin of leukemia cells in the central
nervous system, there are exogenous and endogenous theories. Most
investigators believe in the exogenous theory, namely, that
leukemia cells in the central nervous system have come directly
from the blood circulation (10,11).
However, some investigators believe that in CNSL, leukemia cells
are likely to originate in the nervous system itself, since a very
small number of hematopoietic stem cells in the choroid or meninges
might transform into leukemia cells under the action of certain
inducers, and from these locations proliferate and infiltrate and
form CNSL (12). Price and Johnson
(10) conducted a pathological
study on the brain in leukemia patients and considered that
leukemia cells enter the brain from the peripheral blood
circulation and colonize at the arachnoid, damaging the pia mater
and then invading the brain parenchyma. They also thought that the
incidence of CNSL might not be related to the blood-brain barrier.
Cavallo et al (13) found
that leukemic cells infiltrating the skull and the vertebral bone
marrow could directly invade the dura, and then invade the
arachnoid along the veins between the dura and the arachnoid,
finally causing parenchymal infiltration (9).
In the present study, when various tissues and
organs of mice were collected for RT-PCR assay, positive results
were found in the lymph nodes, lung and bone marrow in one mouse.
We applied nested RT-PCR in the present study, and the results had
a very high specificity. Our results showed the rationality and
possibility of the endogenous doctrine. If leukemic cells spread
from the brain tissue to peripheral organs and lymph tissue, the
following two situations would occur: i) the cells would infiltrate
from the pia mater, pass through the arachnoid and reach the dura,
infiltrate local skull tissue and then spread to the whole body;
ii) the cells would move from the cerebrospinal fluid and pass
through the blood-brain barrier, spread to the blood vessels or
enter the lymphatic vessels and lymph nodes, and then spread to the
whole body. However, the second situation only occurred in 1 of the
19 mice. Tumor cell infiltration occurred at multiple places in
that mouse 3 weeks after lateral ventricle injection, indicating
that as time increased, tumor cells proliferated and spread
continuously, and that leukemia cells are able to spread from the
brain tissue to the periphery. This result is consistent with the
sequence of disease progression in clinical practice from CNSL
relapse to bone marrow relapse to systemic recurrence. In one
mouse, local tissue infiltration occurred and entered the forehead,
causing formation of a local tumor mass. This is also consistent
with the characteristics of acute myeloid leukemia in that it can
easily form local infiltration and chloroma. The results of this
study indicate that both of the above-mentioned methods for
transfer from inside to outside may occur. Thomas (14) studied human and mouse CNSL and found
that the two leukemias have many common characteristics. Leukemia
cell stasis, leukemia nodule formation and dura mater and arachnoid
hemorrhage in the brain were found in human CNSL. These were also
observed in our animal model in the present study.
The model described in the present study is a stable
and repeatable CNSL model that is constructed in NOD/SCID mice
using the human acute monocytic leukemia cell line SHI-1. It
partially reproduces the progression and infiltration seen in acute
monocytic leukemia in human brain tissue, and may provide a
relatively ideal experimental animal model for studies on CNSL and
on the mechanism of extramedullary infiltration of leukemic cells,
treatment of leukemia, molecular mechanisms of leukemia and
targeted gene therapy for leukemia.
In conclusion, intracerebroventricular injection of
acute monocytic leukemia cell line SHI-1 cells in the lateral
ventricle can successfully establish a NOD/SCID mouse model of
central nervous system leukemia.
Acknowledgements
This study was supported by the Medical Scientific
and Technology Research Foundation of Guangdong Province, China
(B2012106).
References
|
1
|
Johnston DL, Alonzo TA, Gerbing RB, Lange
BJ and Woods WG: Risk factors and therapy for isolated central
nervous system relapse of pediatric acute myeloid leukemia. J Clin
Oncol. 23:9172–9178. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kobayashi R, Tawa A, Hanada R, Horibe K,
Tsuchida M and Tsukimoto I; Japanese Childhood AML Cooperative
Study Group. Extramedullary infiltration at diagnosis and prognosis
in children with acute myelogenous leukemia. Pediatr Blood Cancer.
48:393–398. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chen S, Xue Y, Zhang X, Wu Y, Pan J, Wang
Y and Ceng J: A new human acute monocytic leukemia cell line SHI-1
with t(6;11)(q27;q23), p53 gene alterations and high tumorigenicity
in nude mice. Haematologica. 90:766–775. 2005.PubMed/NCBI
|
|
4
|
Chen SN, Xue YQ, Zhang XG, Wu YF, Pan JL,
Wang Y and Cen JN: Establishment and characterization of a human
acute monocytic leukemic cell line, SHI-1, carrying t(6;11)(q27;23)
and p53 gene alteration. Zhonghua Xue Ye Xue Za Zhi. 26:94–99.
2005.(In Chinese).
|
|
5
|
Mitterbauer G, Zimmer C, Pirc-Danoewinata
H, et al: Monitoring of minimal residual disease in patients with
MLL-AF6-positive acute myeloid leukaemia by reverse transcriptase
polymerase chain reaction. Br J Haematol. 109:622–628. 2000.
View Article : Google Scholar
|
|
6
|
Zhang H, Fu T, Xing D, et al:
Establishment of L615 murine model of meningeal leukemia and
preliminary exploration on pathogenesis. Chin J Hematol. 4:335–339.
1983.
|
|
7
|
Li ZJ, Chen ZX, Lu J, Cen JN, He J and Guo
LC: Growth and infiltration of human monocytic leukemia cell in
nude mice: a model for central nervous system leukemia. Zhonghua
Xue Ye Xue Za Zhi. 374–378. 2006.(In Chinese).
|
|
8
|
Kamel-Reid S, Letarte M, Sirard C, et al:
A model of human acute lymphoblastic leukemia in immune-deficient
SCID mice. Science. 246:1597–1600. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gunther R, Chelstrom LM, Tuel-Ahlgren L,
Simon J, Myers DE and Uckum FM: Biotherapy for xenografted human
central nervous system leukemia in mice with severe combined
immunodeficiency using B43 (anti-CD19)-pokeweed antiviral protein
immunotoxin. Blood. 85:2537–2545. 1995.
|
|
10
|
Price RA and Johnson WW: The central
nervous system in childhood leukemia. I The arachnoid. Cancer.
31:520–533. 1973. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li Z, Chen Z, Lu J, et al: Establishment
of a nude mice model of human monocytic leukemia with CNS and
multiorgan extramedullary infiltration. Eur J Haematol. 77:128–133.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sullivan MP: Leukemia infiltration of
meninges spinal nerve roots. Pediatrics. 32:63–72. 1963.PubMed/NCBI
|
|
13
|
Cavallo F, Forni M, Riccardi C, et al:
Growth and spread of human malignant T lymphoblasts in
immunosuppressed nude mice: a model for meningeal leukemia. Blood.
80:1279–1283. 1992.PubMed/NCBI
|
|
14
|
Thomas LB: Pathology of leukemia in the
brain and meninges: postmortem studies of patients with acute
leukemia and of mice given inoculations of L1210 leukemia. Cancer
Res. 25:1555–1571. 1965.PubMed/NCBI
|