Introduction
Gastric carcinoma (GC) remains the third leading
cause of cancer-related mortality in men worldwide (1). To date, there are few effective
clinical treatments for this highly malignant tumor, and
conventional adjuvant treatments have limited effects on the
survival of patients with advanced gastric cancer (2). Much research has been conducted to
identify efficient chemotherapeutic agents for the cure and
prevention of GC, and recently apoptosis has been shown to play a
significant role in the treatment of GC cells.
Arsenic trioxide (As2O3) is an
arsenic compound that has been used as a medicinal agent for more
than 2400 years (3). In the 1970s,
Chinese researchers were the first to discover its ability to cure
acute promyelocytic leukemia (APL) (4). Since then other research groups have
demonstrated worldwide that As2O3 also
inhibits the growth of various solid tumors, including esophageal
carcinoma (5,6), breast (3,7,8),
bladder (9), lung (10) and liver cancer (11), multiple myeloma (12), neuroblastoma (13), colon (14) and ovarian cancer (15). It has been shown that
As2O3 regulates proliferation, invasion,
differentiation, angiogenesis and apoptosis of cancer cells
(16). However, the precise
mechanism of As2O3-related apoptosis
induction of cancer cells is not fully understood. Recent
experiments confirm that As2O3 affects the
activities of protein kinase B (Akt), c-Jun N-terminal kinases
(JNK), nuclear factor κB (NF-κB), glutathione and calcium
signaling, reactive oxygen species (ROS), caspases, as well as pro-
and anti-apoptotic proteins (17–20).
It was noted that As2O3 can reduce the
activation of the Akt/mTOR pathway by reducing Akt, p70S6K and rpS6
phosphorylation in human leukemia cells (21). The PI3K/Akt/mTOR pathway is a
crucial regulatory cascade that is central to a variety of
physiological functions, including cell cycle regulation, survival,
protein synthesis, metabolism, motility, apoptosis, proliferation
and angiogenesis (22,23). The phosphoinositide 3-kinase (PI3K)
activates Akt, a serine/threonine kinase, which phosphorylates the
mammalian target of rapamycin (mTOR) repressor tuberous sclerosis
complex 2 (24), which in turn
activates mTOR downregulation of autophagy inducing
autophagy-related (Atg) proteins (25,26).
Recently it has been demonstrated that As2O3
suppresses PI3K/Akt activity and induces JNK activation thereby
enhancing chronic B-lymphocytic leukemia cell apoptosis (27).
The major apoptosis pathways are the extrinsic
pathways (death receptor) and intrinsic pathways (mitochondrial)
(28). The common effector for
extrinsic apoptotic pathway initiation is FASL, which regulates
apoptosis via binding to FASR, a member of the tumor necrosis
factor (TNF) receptor family of proteins. Deregulation of the FAS
pathway has been implicated in various malignancies and diseases
(29). After stimulation of the
death receptor pathway by the FAS ligand, conformational changes to
the FAS receptor lead to cleavage of pro-caspase 8 into its
activated form, which then cleaves other effector caspases
eventually leading to apoptosis. During the apoptotic process,
Bcl-2-associated × (Bax) inhibits the anti-apoptotic B-cell
lymphoma 2 (Bcl-2) protein (30)
and permeabilizes the mitochondrial outer membrane, leading to
cytochrome c release (31).
In the present study, we explored the effects of
As2O3 on expression levels of FAS, caspase-8
and Bax proteins in human gastric cancer SGC-7901 cells. Then we
further analyzed the role of As2O3 in the
Akt/mTOR pathway in As2O3-exposed cells.
Materials and methods
Materials
As2O3 solution was purchased
from YiDa Pharmacy. The stock solution of arsenic trioxide was 8 mM
and was stored at 4°C. RPMI-1640 medium and FBS were purchased from
Hyclone. Anti-Akt polyclonal antibodies (9272) were purchased from
Cell Signaling Technology. Anti-phospho-Akt (s473), anti-mTOR,
anti-phospho-mTOR (s2448), anti-Bax, anti-caspase-8 and anti-FAS
polyclonal antibodies were purchased from ImmunoWay. Anti-β-actin
monoclonal, goat anti-mouse and goat anti-rabbit secondary
antibodies conjugated to horseradish peroxidase were purchased from
Santa Cruz Biotechnology.
Cell culture and treatment
Human SGC-7901 gastric cancer cells were cultured in
RPMI-1640 medium containing 10% FBS in a 5% CO2
humidified atmosphere chamber at 37°C. For the experiments, FBS was
reduced to 2%, and exponentially growing cells were incubated for
the indicated time periods with different concentrations of
As2O3 (0, 2.5, 5, 7.5, 10, 12.5 and 15
μmol/l).
WST-1 proliferation assay
The effect of As2O3 on in
vitro growth inhibition of SGC-7901 cells was measured using
the WST-1 Cell Proliferation and Cytotoxicity Assay Kit (Beyotime
Institute of Biotechnology) according to the manufacturer’s
protocol. SGC-7901 cells were seeded in 96-well plates at a density
of 1.0×104 cells per well in 200 μl RPMI-1640 medium
containing 10% FBS for 24 h. Then the cells were exposed to
different concentrations of As2O3 (0, 2.5, 5,
7.5, 10, 12.5 and 15 μmol/l) in RPMI-1640 medium containing 2% FBS
for 24, 48 and 72 h. Finally 20 μl of WST-1 solution was added to
each well, and the cells were incubated for another 1 h. The
absorbance at 450 nm was measured using a microplate reader
(Finnpipette MK3 Multiskan). The amount of the formazan dye, which
is generated by activities of dehydrogenases in the cells, is
proportional to the number of living cells. Inhibitory rates of
cellular growth were calculated with the following formula:
Inhibitory rate (%) = (1 − A value of experimental group/A value in
the control group) × 100%. The 0 μmol/l group was used as the
control group. A graph with inhibitory cell growth rates (y-axis)
against the concentrations of As2O3 (x-axis)
was plotted.
Analysis of nuclear morphology by DAPI
staining
Apoptosis was assessed based on changes in the
nuclear morphology by staining the cells with the fluorescent DNA
dye 4′,6-diamidino-2′-phenylindole dihydrochloride (DAPI) (Roche).
Briefly, cells were treated with As2O3 (0 and
10 μmol/l) in RPMI-1640 medium containing 2% FBS for 48 h. Then the
cells were washed with PBS and incubated with 1 μg/ml DAPI in
methanol for 30 min at 37°C in darkness. Slides were viewed using a
fluorescence microscope with ultraviolet (UV) excitation at 300–500
nm. Cells were evaluated as normal or apoptotic depending on
morphological characteristics. Normal nuclei (smooth nuclei) and
apoptotic nuclei (condensed or fragmented chromatin) were
observed.
Analysis of apoptosis
Cells were treated with different concentrations of
As2O3 (0, 7.5, 10, 12.5 and 15 μmol/l) in 2%
FBS and RPMI-1640 for 48 h, collected and then stained using the
Annexin V-FITC Apoptosis Detection Kit I (BD Biosciences) for flow
cytometric analyses. The 0 μmol/l group served as the control.
Protein extraction and western blot
analysis
Cells were treated with different concentrations of
As2O3 (0, 5, 7.5, 10, 12.5 and 15 μmol/l) in
2% FBS and RPMI-1640 medium for 48 h. Both adherent and floating
cells were harvested and lysed with RIPA lysis buffer and
phenylmethanesulfonyl fluoride (Beyotime Institute of
Biotechnology), incubated at 4°C for 40 min and centrifuged for 10
min at 12,000 rpm. Total protein in the cell lysate was measured
with an enhanced BCA protein assay kit (Beyotime Institute of
Biotechnology). For western blot analysis, equal amounts of protein
were separated by SDS-PAGE and then transferred onto PVDF membranes
(Millipore). The membranes were blocked for 1.5 h in a non-fat
dried milk solution containing 1% Tween-20. The membranes were then
incubated with primary antibodies for β-actin (1:800), Akt (1:800),
p-Akt (1:800), mTOR (1:800), p-mTOR (1:800), Bax (1:1000),
caspase-8 (1:1000) and FAS (1:1000) overnight at 4°C, followed by
incubation with anti-mouse or anti-rabbit (1:5000) secondary
antibodies for 1 h. Finally, protein bands were detected using a
chemiluminescent substrate (HRP) kit (Beyotime Institute of
Biotechnology). The β-actin level was used as an internal
standard.
Statistical analyses
All experiments were performed at least three times.
Data for each series of experiments (performed in triplicates) are
expressed as the mean values ± standard deviation of the mean (SD).
Statistical significance of differences between groups was analyzed
using ANOVA analysis. P<0.05 was considered to indicate a
statistically significant difference.
Results
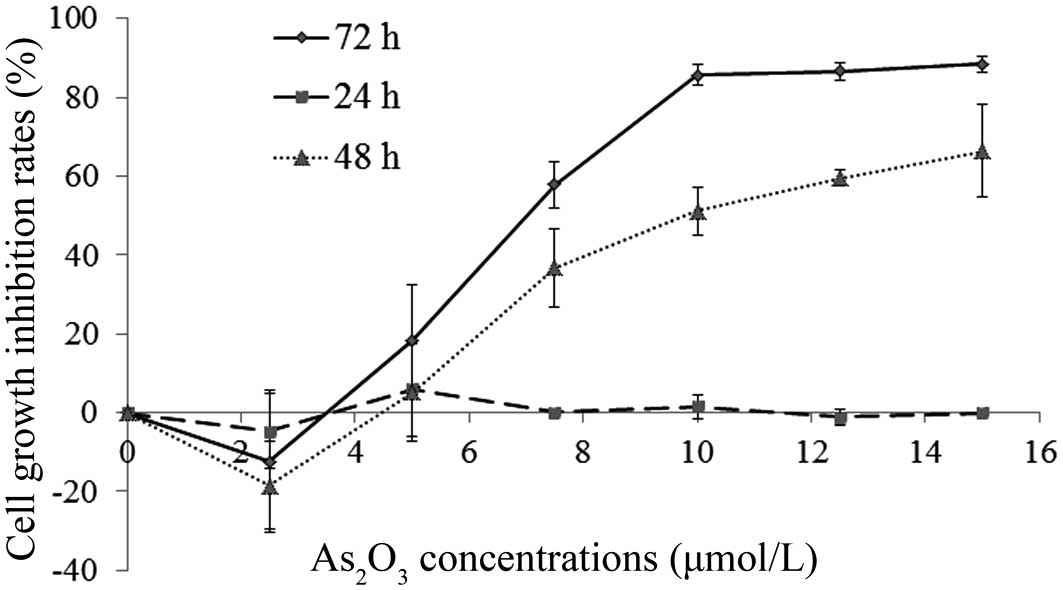
Extended incubation with
As2O3 leads to cell growth inhibition
Human SGC-7901 gastric cancer cells were incubated
with different concentrations of As2O3 (0,
2.5, 5, 7.5, 10, 12.5 and 15 μmol/l) for 24, 48 and 72 h and the
cell growth inhibition was recorded using the WST-1 assay. When the
As2O3 solution concentrations were >5
μmol/l, the cell growth was significantly reduced after 48 and 72-h
incubation periods, whereas after a 24-h incubation none of the
As2O3 concentrations had an effect on cell
growth. Concentrations <5 μmol/l led to reduced growth
inhibition (Fig. 1).
As2O3 leads to
apoptosis of SGC-7901 cells
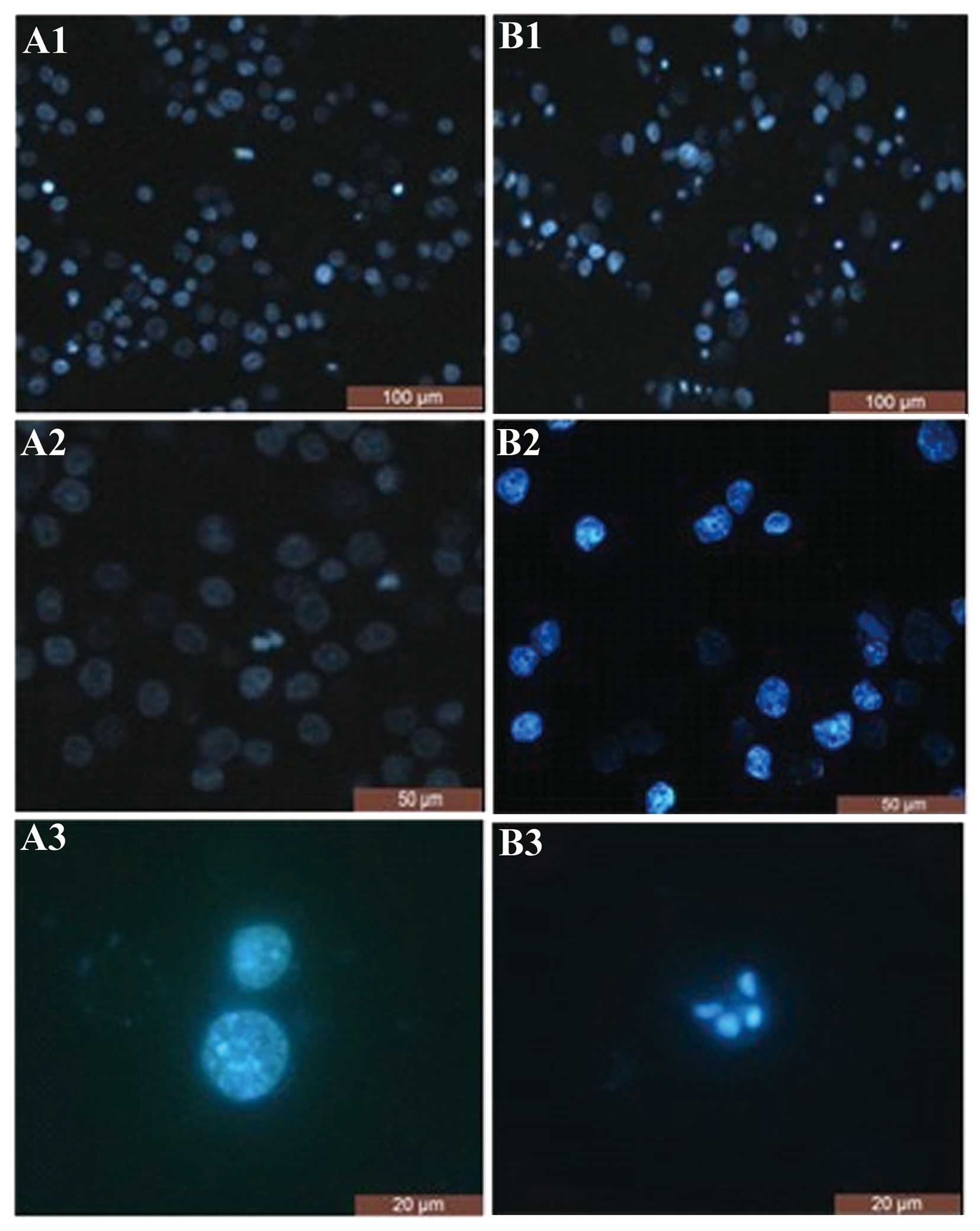
Analysis of nuclear morphology as
assessed by DAPI staining
The SGC-7901 cells were treated with 10 μmol/l
As2O3 for 48 h, and apoptosis was visualized
by DAPI staining using fluorescence microscopy. Compared with the
control, a large number of cells displayed morphological changes
exhibiting the typical characteristics of apoptotic cell death,
including cell shrinkage, chromatin condensation, chromatin
crescent formation/margination, DNA fragmentation and apoptotic
body formation (Fig. 2).
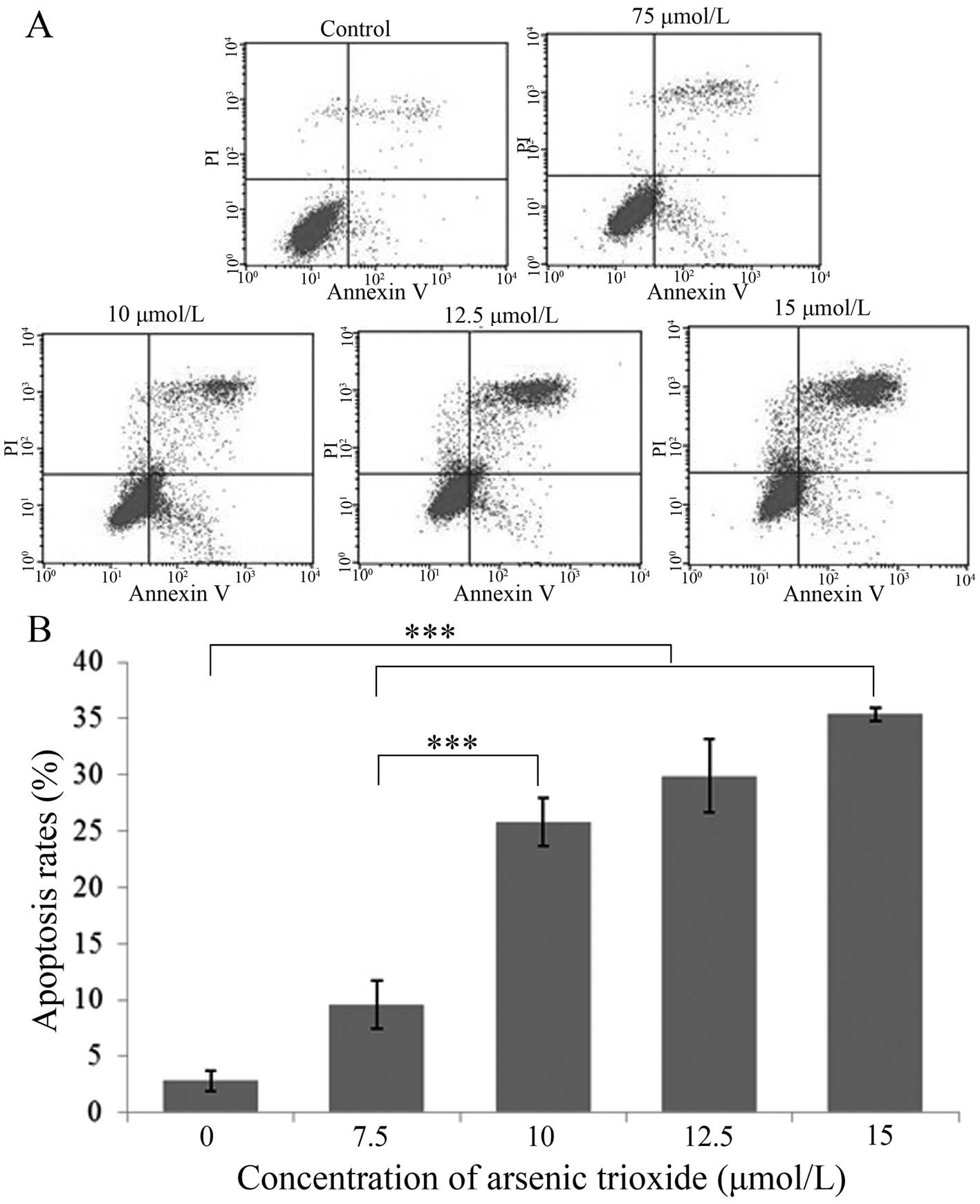
Analysis of apoptosis by flow
cytometry
With increasing concentrations (0, 7.5, 10, 12.5 and
15 μmol/l) of As2O3 in the growth media, the
apoptosis rates increased after 48 h from 2.83±0.88, 9.85±2.18,
25.81±2.17 and 29.92±3.30 to 35.40±0.58%, which indicated that
As2O3 induced the apoptosis of human gastric
cancer SGC-7901 cells in a dose-dependent manner (Fig. 3).
Protein extraction and western blot
analysis
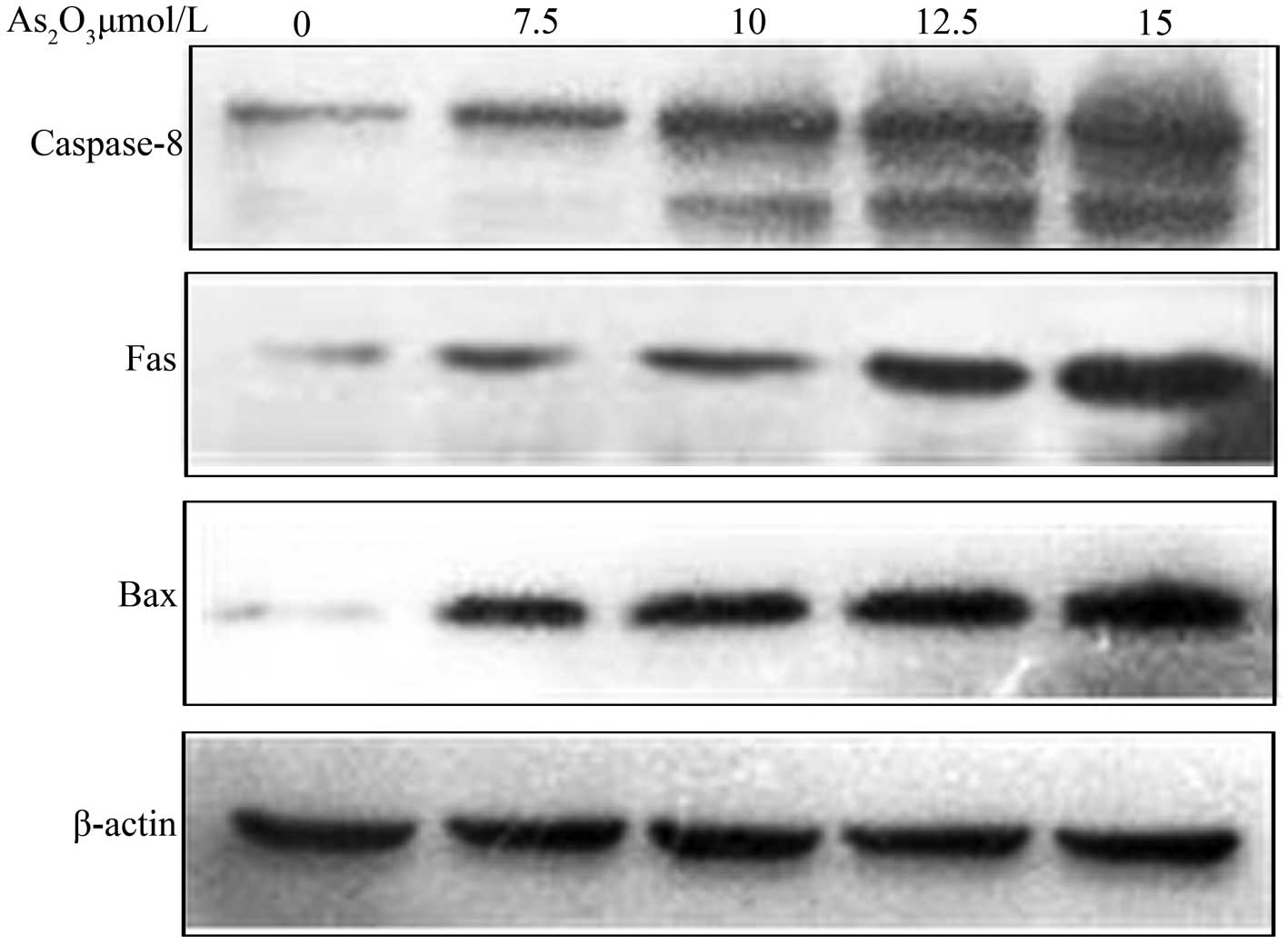
As2O3 induces
Bax, Fas and caspase-8 activation
SGC-7901 cells were incubated with different
As2O3 concentrations (0, 7.5, 10, 12.5 and 15
μmol/l) for 48 h, and then Bax, Fas and caspase-8 protein
expression levels were analyzed via western blotting. As shown in
Fig. 4, expresssion of Bax, Fas and
caspase-8 protein was increased with increasing
As2O3 concentrations.
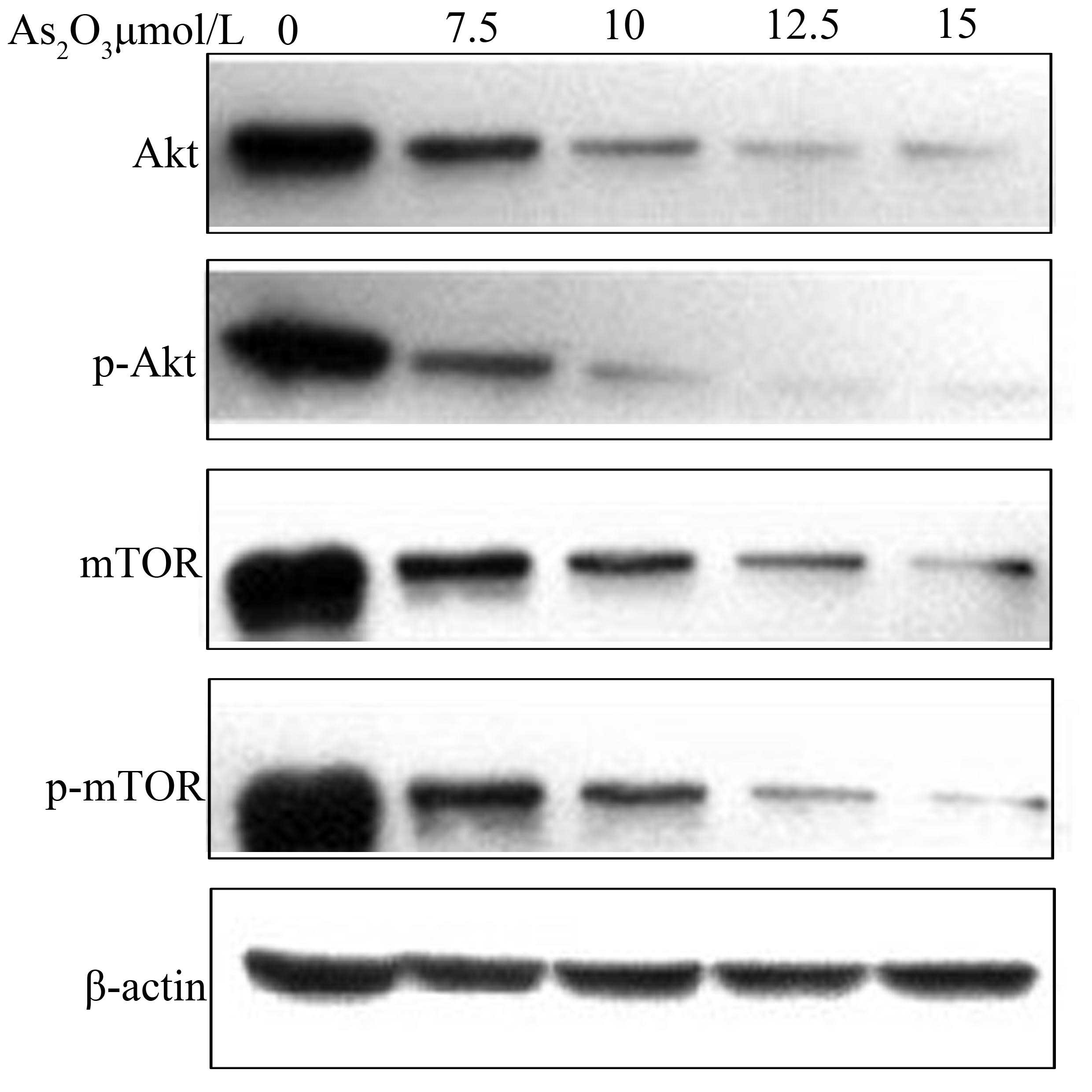
As2O3 suppresses
Akt, p-Akt, mTOR and p-mTOR activation
SGC-7901 cells were incubated with different
As2O3 concentrations (0, 7.5, 10, 12.5 and 15
μmol/l) for 48 h and Akt, p-Akt, mTOR and p-mTOR protein expression
levels were analyzed via western blotting. As shown in Fig. 5, Akt, p-Akt, mTOR and p-mTOR protein
expression levels decreased with increasing
As2O3 concentrations.
Discussion
In the present study, we demonstrated that
As2O3 induced the apoptosis of human gastric
cancer SGC-7901 cells in a dose- and time-dependent manner, which
is in agreement with previous findings of the
As2O3-triggered apoptosis of lung cancer
cells (32).
Further analyses revealed that the apoptotic
proteins Bax, Fas and caspase-8 were upregulated and the
anti-apoptotic proteins Akt, p-Akt, mTOR as well as phosphorylated
mTOR (p-mTOR) were downregulated. The PI3K (phosphatidylinositol 3
kinase) pathway is a signal transduction cascade, which is at the
center of many physiological functions including cell cycle
regulation, cell survival, protein synthesis, metabolism as well as
blood vessel formation. There are two key elements (Akt and mTOR)
in the PI3K transduction pathway. Akt (serine/threonine kinase) is
the regulator of the PI3K transduction pathways by regulating a
variety of downstream effectors. A variety of growth factors,
cytokines and hormones lead to the phosphorylation of Akt, which in
turn activates downstream effectors including mTOR directly or
indirectly by preventing the combination of mTORC1 and mTORC2
thereby promoting protein synthesis and cell growth (33,34).
Akt also inactivates cell cycle inhibitors (p21 and p27) and
promotes cell cycle proteins (c-Myc and cyclinD1) to maintain cell
survival (35,36). Another study found that Akt
suppressed the apoptosis inhibition genes (BIM and BAD) and reduced
the expression of the tumor-suppressor protein (p53) restricting
programmed cell death and promoting cell survival (37). Our results showed that
As2O3 concentrations <5 μmol/l promoted
cell growth and during the 24-h incubation cell growth was not
inhibited by any As2O3 concentration
(Fig. 1). In previous studies, the
apoptotic effect of As2O3 was attributed to
reactive oxygen species development (38,39),
and As2O3 has also been shown to inhibit
mitochondrial respiration, thereby enhancing ROS occurrence
(40), which has been used to
sensitize tumor cells for radiation therapy (41). Autophagy constitutes a stress
adaptation that avoids cell death, and cells can compensate
oxidative stress damages to a certain extent through autophagy,
which was demonstrated by different cell reactions upon low and
high dosage exposures to safingol, which is a ROS inducer (42). Autophagy following
As2O3 exposure has also been reported
(43). We suggest that at low doses
of As2O3 up to 5 μmol/l, autophagy is the
main mechanism triggered in SGC-7901 cells and apoptosis is blocked
(44) leading to somewhat reduced
growth inhibition. Moreover, in short periods (24 h), the ROS
development is under the threshold for inducing apoptosis, probably
also due to oxygen radical squelching mechanisms (45). This is supported by the finding that
apoptotic effects of As2O3 are most
pronounced in tumor cells with low GSH levels, and ascorbic acid
could further enhance its capacity for apoptosis induction
(45).
A drawback of our study was that apoptotic
mechanisms are complex and this study is a preliminary study of the
Akt/mTOR anti-apoptotic pathway, while other anti-apoptosis
pathways need further investigation. In addition, the effective
dose of As2O3 was >5 μmol/l, which is
higher than the allowed clinical therapeutic dose of 1–2 μmol/l,
thus further long-term and sensitizing agent evaluations are
warranted (45).
In conclusion, our in vitro results showed
that As2O3 can induce apoptosis in human
gastric cancer SGC-7901 cells. As2O3
treatment led to enhanced expression of the apoptotic proteins Bax,
Fas and caspase-8, and reduced the expression of the anti-apoptotic
proteins Akt and mTOR as well as their phosphorylated forms p-Akt
and p-mTOR in a time- and dose- dependent manner. Since the
effective dose of As2O3 was higher than the
therapeutic limit and growth inhibition rate reductions were
incubation time-dependent, further research is necessary to
establish As2O3 for the treatment of gastric
cancers.
Acknowledgements
We thank Xuguang Zhang for the help and support.
This work was supported by grants from the Heilongjiang Province
Natural Science Fund Project (D200862).
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar
|
|
2
|
Lim L, Michael M, Mann GB and Leong T:
Adjuvant therapy in gastric cancer. J Clin Oncol. 23:6220–6232.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang Y, Zhang Y, Yang L, et al: Arsenic
trioxide induces the apoptosis of human breast cancer MCF-7 cells
through activation of caspase-3 and inhibition of HERG channels.
Exp Ther Med. 2:481–486. 2011.PubMed/NCBI
|
|
4
|
Zhou J: Arsenic trioxide: an ancient drug
revived. Chin Med J. 125:3556–3560. 2012.PubMed/NCBI
|
|
5
|
Shen ZY, Shen J, Cai WJ, Hong C and Zheng
MH: The alteration of mitochondria is an early event of arsenic
trioxide induced apoptosis in esophageal carcinoma cells. Int J Mol
Med. 5:155–158. 2000.PubMed/NCBI
|
|
6
|
Shen ZY, Zhang Y, Chen JY, et al:
Intratumoral injection of arsenic to enhance antitumor efficacy in
human esophageal carcinoma cell xenografts. Oncol Rep. 11:155–159.
2004.PubMed/NCBI
|
|
7
|
Chow SK, Chan JY and Fung KP: Inhibition
of cell proliferation and the action mechanisms of arsenic trioxide
(As2O3) on human breast cancer cells. J Cell
Biochem. 93:173–187. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ye J, Li A, Liu Q, Wang X and Zhou J:
Inhibition of mitogen-activated protein kinase enhances apoptosis
induced by arsenic trioxide in human breast cancer MCF-7 cells.
Clin Exp Pharmacol Physiol. 32:1042–1048. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jutooru I, Chadalapaka G, Sreevalsan S, et
al: Arsenic trioxide downregulates specificity protein (Sp)
transcription factors and inhibits bladder cancer cell and tumor
growth. Exp Cell Res. 316:2174–2188. 2010. View Article : Google Scholar
|
|
10
|
Chien CW, Yao JH, Chang SY, Lee PC and Lee
TC: Enhanced suppression of tumor growth by concomitant treatment
of human lung cancer cells with suberoylanilide hydroxamic acid and
arsenic trioxide. Toxicol Appl Pharmacol. 257:59–66. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li H, Gong J, Jiang X and Shao H: Arsenic
trioxide treatment of rabbit liver VX-2 carcinoma via hepatic
arterial cannulation-induced apoptosis and decreased levels of
survivin in the tumor tissue. Croat Med J. 54:12–16. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Matulis SM, Morales AA, Yehiayan L, Lee
KP, Cai Y and Boise LH: Alterations in glutathione levels and
apoptotic regulators are associated with acquisition of arsenic
trioxide resistance in multiple myeloma. PloS One. 7:e526622012.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ora I, Bondesson L, Jonsson C, et al:
Arsenic trioxide inhibits neuroblastoma growth in vivo and promotes
apoptotic cell death in vitro. Biochem Biophys Res Commun.
277:179–185. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nakagawa Y, Akao Y, Morikawa H, et al:
Arsenic trioxide-induced apoptosis through oxidative stress in
cells of colon cancer cell lines. Life Sci. 70:2253–2269. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bornstein J, Sagi S, Haj A, Harroch J and
Fares F: Arsenic trioxide inhibits the growth of human ovarian
carcinoma cell line. Gynecol Oncol. 99:726–729. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu Y, Zhang W, Zhang X, Qi Y, Huang D and
Zhang Y: Arsenic trioxide inhibits invasion/migration in SGC-7901
cells by activating the reactive oxygen species-dependent
cyclooxygenase-2/matrix metalloproteinase-2 pathway. Exp Biol Med.
236:592–597. 2011. View Article : Google Scholar
|
|
17
|
Bowling BD, Doudican N, Manga P and Orlow
SJ: Inhibition of mitochondrial protein translation sensitizes
melanoma cells to arsenic trioxide cytotoxicity via a reactive
oxygen species dependent mechanism. Cancer Chemother Pharmacol.
63:37–43. 2008. View Article : Google Scholar
|
|
18
|
Florea AM and Büsselberg D: Anti-cancer
drugs interfere with intracellular calcium signaling.
Neurotoxicology. 30:803–810. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gao F, Yi J, Shi GY, Li H, Shi XG and Tang
XM: The sensitivity of digestive tract tumor cells to
As2O3 is associated with the inherent
cellular level of reactive oxygen species. World J Gastroenterol.
8:36–39. 2002.PubMed/NCBI
|
|
20
|
Izdebska M, Grzanka A, Szczepanski MA and
Litwiniec A: Selected mechanisms of the therapeutic effect of
arsenic trioxide in cancer treatment. Postepy Hig Med Dosw.
62:463–467. 2008.(In Polish).
|
|
21
|
Calvino E, Estan MC, Simon GP, et al:
Increased apoptotic efficacy of lonidamine plus arsenic trioxide
combination in human leukemia cells. Reactive oxygen species
generation and defensive protein kinase (MEK/ERK, Akt/mTOR)
modulation. Biochem Pharmacol. 82:1619–1629. 2011. View Article : Google Scholar
|
|
22
|
Carracedo A and Pandolfi PP: The PTEN-PI3K
pathway: of feedbacks and cross-talks. Oncogene. 27:5527–5541.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Markman B, Dienstmann R and Tabernero J:
Targeting the PI3K/Akt/mTOR pathway - beyond rapalogs. Oncotarget.
1:530–543. 2010.PubMed/NCBI
|
|
24
|
Hay N: The Akt-mTOR tango and its
relevance to cancer. Cancer Cell. 8:179–183. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
He C and Klionsky DJ: Regulation
mechanisms and signaling pathways of autophagy. Annu Rev Genet.
43:67–93. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Meijer AJ and Codogno P: Regulation and
role of autophagy in mammalian cells. Int J Biochem Cell Biol.
36:2445–2462. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Redondo-Munoz J, Escobar-Diaz E, Hernandez
Del Cerro M, et al: Induction of B-chronic lymphocytic leukemia
cell apoptosis by arsenic trioxide involves suppression of the
phosphoinositide 3-kinase/Akt survival pathway via c-jun-NH2
terminal kinase activation and PTEN upregulation. Clin Cancer Res.
16:4382–4391. 2010. View Article : Google Scholar
|
|
28
|
Ghobrial IM, Witzig TE and Adjei AA:
Targeting apoptosis pathways in cancer therapy. CA Cancer J Clin.
55:178–194. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Komarov AP, Rokhlin OW, Yu CA and Gudkov
AV: Functional genetic screening reveals the role of mitochondrial
cytochrome b as a mediator of FAS-induced apoptosis. Proc Natl Acad
Sci USA. 105:14453–14458. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Oltvai ZN, Milliman CL and Korsmeyer SJ:
Bcl-2 heterodimerizes in vivo with a conserved homolog, Bax, that
accelerates programmed cell death. Cell. 74:609–619. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Suen DF, Norris KL and Youle RJ:
Mitochondrial dynamics and apoptosis. Genes Dev. 22:1577–1590.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Han B, Zhou G, Zhang Q, et al: Effect of
arsenic trioxide (ATO) on human lung carcinoma PG cell line: ATO
induced apoptosis of PG cells and decreased expression of Bcl-2,
Pgp. J Exp Ther Oncol. 4:335–342. 2004.PubMed/NCBI
|
|
33
|
Guertin DA and Sabatini DM: Defining the
role of mTOR in cancer. Cancer Cell. 12:9–22. 2007. View Article : Google Scholar
|
|
34
|
Sarbassov DD, Guertin DA, Ali SM and
Sabatini DM: Phosphorylation and regulation of Akt/PKB by the
rictor-mTOR complex. Science. 307:1098–1101. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Brunet A, Bonni A, Zigmond MJ, et al: Akt
promotes cell survival by phosphorylating and inhibiting a Forkhead
transcription factor. Cell. 96:857–868. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Diehl JA, Cheng M, Roussel MF and Sherr
CJ: Glycogen synthase kinase-3beta regulates cyclin D1 proteolysis
and subcellular localization. Genes Dev. 12:3499–3511. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Engelman JA, Luo J and Cantley LC: The
evolution of phosphatidylinositol 3-kinases as regulators of growth
and metabolism. Nat Rev Genet. 7:606–619. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chen YC, Lin-Shiau SY and Lin JK:
Involvement of reactive oxygen species and caspase 3 activation in
arsenite-induced apoptosis. J Cell Physiol. 177:324–333. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Woo SH, Park IC, Park MJ, et al: Arsenic
trioxide induces apoptosis through a reactive oxygen
species-dependent pathway and loss of mitochondrial membrane
potential in HeLa cells. Int J Oncol. 21:57–63. 2002.
|
|
40
|
Pelicano H, Feng L, Zhou Y, et al:
Inhibition of mitochondrial respiration: a novel strategy to
enhance drug-induced apoptosis in human leukemia cells by a
reactive oxygen species-mediated mechanism. J Biol Chem.
278:37832–37839. 2003. View Article : Google Scholar
|
|
41
|
Diepart C, Karroum O, Magat J, et al:
Arsenic trioxide treatment decreases the oxygen consumption rate of
tumor cells and radiosensitizes solid tumors. Cancer Res.
72:482–490. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ling LU, Tan KB, Lin H and Chiu GN: The
role of reactive oxygen species and autophagy in safingol-induced
cell death. Cell Death Dis. 2:e1292011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhang G, Liu J, Zhang Y, et al:
Cbl-b-dependent degradation of FLIP(L) is involved in ATO-induced
autophagy in leukemic K562 and gastric cancer cells. FEBS Lett.
586:3104–3110. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Cheng Y, Qiu F, Ye YC, et al: Autophagy
inhibits reactive oxygen species-mediated apoptosis via activating
p38-nuclear factor-kappa B survival pathways in oridonin-treated
murine fibrosarcoma L929 cells. FEBS J. 276:1291–1306. 2009.
View Article : Google Scholar
|
|
45
|
Dai J, Weinberg RS, Waxman S and Jing Y:
Malignant cells can be sensitized to undergo growth inhibition and
apoptosis by arsenic trioxide through modulation of the glutathione
redox system. Blood. 93:268–277. 1999.PubMed/NCBI
|