Introduction
Gastric cancer is one of the most common
malignancies worldwide, with an estimated 934,000 cases reported
globally in 2002, and is the second most common cause of death from
cancer (1). The prognosis of
gastric cancer is poor with an estimated relative 5-year survival
rate of <20%. Currently, one of the urgent tasks is to discover
the early molecular mechanisms involved in the development of
gastric cancer as well as reliable biomarkers and possible
therapeutic targets (2).
Epidemiological studies have shown that nonsteroid
anti-inflammatory drugs (NSAIDs), whose target is the COX enzyme,
are associated with a reduced risk of gastric cancer. Of the two
cloned COX genes, COX-2 expression is elevated in gastric
adenocarcinomas (GACs), and correlates with clinicopathological
parameters, including depth of invasion and lymph node metastasis,
suggesting the promotive role of COX-2 in the aggressive behavior
of GAC (3). COX-2 is significantly
correlated with vascular endothelial growth factor (VEGF),
platelet-derived growth factor (PDGF) and Bcl-2, as independent
prognostic factors for overall survival in gastric cancer patients,
suggesting that COX-2 contributes to gastric cancer development by
promoting angiogenesis and inhibiting apoptosis (4). COX-2 expression is also upregulated in
early gastric cancer, and bile acids induce its expression in
gastric cell lines, indicating a role of bile reflux in gastric
carcinogenesis (5). Therefore,
COX-2 is an independent prognostic factor in gastric cancer, and
some strategies to target COX-2 may provide insight into the
effective treatment of GAC (6).
Proliferating cell nuclear antigen (PCNA) is a
36-kDa nuclear protein associated with the cell cycle. The abnormal
expression of PCNA protein is related with the oncogenesis of
gastric carcinoma (7), and is
positively associated with the differentiation status and Lauren’s
classification, suggesting the connection of PCNA with tumor
malignancy (8). PCNA is
significantly correlated with the depth of wall invasion and local
lymph node involvement in metastatic lymph nodes of GAC (9). Moreover, the extract of curcumae was
found to downregulate the expression levels of COX-2 and PCNA in
the gastric mucosa of rats during carcinogenesis, and reduce the
incidence of gastric cancer, suggesting PCNA may be a potential
therapeutic target for gastric cancer (10).
In the present study, we assessed the expression of
COX-2 and PCNA proteins in human GAC using immunohistochemical
(IHC) assay through a tissue microarray procedure. We also used the
loss-of-function approach and pretreatment with a COX-2 inhibitor
to investigate the effects of targeted knockdown of COX-2 on the
proliferative activities, migratory and invasive potential in human
GAC cells. We hypothesized that targeting COX-2 signaling may be an
effective strategy to improve the treatment of GAC.
Materials and methods
Materials
The human GAC SGC-7901 and MKN-45 cell lines used in
the experiments were from the Institute of Biochemistry and Cell
Biology (Shanghai, China). The adenovirus-mediated COX-2 siRNA
vector and the negative control vector were from Genechem
(Shanghai, China): COX-2 siRNA sense, 5′-AAC UGC UCA ACA CCG GAA
Udtdt-3′ and antisense, 5′-AUU CCG GUG UUG AGC AGU Udtdt-3′;
negative control vector (nonsense siRNA) sense, 5′-UUC UCC GAA CGU
GUC ACG Utt-3′ and antisense, 5′-ACG UGA CAC GUU CGG AGA Att-3′.
The primers for COX-2 and PCNA were synthesized by ABI (Framingham,
USA). The tissue microarray of human GAC was purchased from a
branch of Biomax (Xi’an, China). All antibodies were obtained from
Santa Cruz Biotechnology (Dallas, TX, USA).
Drugs and reagents
Celecoxib was purchased from LKT Laboratories (St.
Paul, MN, USA). Dulbecco’s modified Eagle’s medium (DMEM) and fetal
bovine serum (FBS) were from Thermo Fisher Scientific Inc.
(Waltham, MA, USA); TRIzol reagent and Lipofectamine 2000 were from
Invitrogen (Carlsbad, CA, USA); M-MLV Reverse Transcriptase was
from Promega (Madison, WI, USA); SYBR-Green Master Mix was from
Takara (Otsu, Japan). ECL-Plus kit was from GE Healthcare
(Piscataway, NJ, USA).
Clinical samples and data
The tissue microarray was prepared for IHC. Human
GAC tissues and the ANCTs were obtained from biopsies in a total of
45 consecutive cases of GAC admitted to our hospital from January
2006 to December 2010. The study was approved by the Medical Ethics
Committee of Shanghai Jiao Tong University, and written informed
consent was obtained from the patients or their parents before
sample collection. Two pathologists respectively reviewed all of
the cases.
Tissue microarray
The Advanced Tissue Arrayer (ATA-100, Chemicon
International, Tamecula, CA, USA) was used to create holes in a
recipient paraffin block and to acquire cylindrical core tissue
biopsies with a diameter of 1 mm from the specific areas of the
‘donor’ block. The tissue core biopsies were transferred to the
recipient paraffin block at defined array positions. The tissue
microarrays contained tissue samples from 45 formalin-fixed
paraffin-embedded cancer specimens with known diagnosis, and
corresponding ANCTs from these patients. The block was incubated in
an oven at 45°C for 20 min to allow complete embedding of the
grafted tissue cylinders in the paraffin of the recipient block,
and then stored at 4°C until microtome sectioning.
Immunohistochemical staining
Anti-COX-2 and -PCNA antibodies were used for
immunohistochemical (IHC) detection of the expression of COX-2 and
PCNA proteins in the tissue microarrays. Tissue microarray sections
were processed for IHC analysis of COX-2 and PCNA proteins as
follows. Immunohistochemical examinations were carried out on 3-mm
sections. For anti-COX-2 and PCNA immunohistochemistry, unmasking
was performed with 10 mM sodium citrate buffer, pH 6.0, at 90°C for
30 min. For anti-COX-2 and -PCNA immunohistochemistry, antigen
unmasking was not necessary. Sections were incubated in 0.03%
hydrogen peroxide for 10 min at room temperature to remove
endogenous peroxidase activity, and then in blocking serum [0.04%
bovine serum albumin (A2153; Sigma-Aldrich, Shanghai, China) and
0.5% normal goat serum (X0907; Dako Corp., Carpinteria, CA, USA) in
PBS] for 30 min at room temperature. Anti-COX-2 and -PCNA
antibodies were used at a dilution of 1:200. The antibody was
incubated overnight at 4°C. Sections were then washed three times
for 5 min in PBS. Non-specific staining was blocked with 0.5%
casein and 5% normal serum for 30 min at room temperature. Finally,
staining was developed using diaminobenzidine substrate, and
sections were counterstained with hematoxylin. Normal serum or PBS
was used to replace anti-COX-2 and -PCNA antibodies in the negative
controls.
Quantification of protein expression
The expression of COX-2 and PCNA was
semi-quantitatively estimated as total immunostaining scores, which
were calculated as the product of a proportion score and an
intensity score. The proportion and intensity of the staining was
evaluated independently by two observers. The proportion score
reflected the fraction of positive staining cells (score 0, <5%;
score 1, 5–10%; score 2, >10–50%; score 3, >50–75%; score 4,
>75%), and the intensity score represented the staining
intensity (score 0, no staining; score 1, weak positive; score 2,
moderate positive; score 3, strong positive). Finally, a total
expression score was given ranging from 0 to 12. ‘−’, total score
of 0 to <2; ‘+’, total score ≥2 to <5; ‘++’, total score of
≥5 to <8; ‘+++’, total score of ≥8 to 12. Based on the analysis
in advance, COX-2 and PCNA were regarded as negative expression in
GAC tissues if the score was <2, and positive expression if the
score was ≥2.
Cell culture and infection
GAC cells were cultured in DMEM supplemented with
10% heat-inactivated FBS, 100 U/ml of penicillin and 100 μg/ml of
streptomycin. They were all placed in a humidified atmosphere
containing 5% CO2 at 37°C. On the day of transduction,
GAC SGC-7901 and MKN-45 cells were replated at 5×104
cells/well in 24-well plates containing serum-free growth medium
with polybrene (5 mg/ml). When the cells reached 50% confluency,
they were infected with the recombinant adenovirus or the control
virus at the optimal MOI (multiplicity of infection) of 50, and
cultured at 37°C in 5% CO2 for 4 h. Then supernatant was
discarded, and serum-containing growth medium was added. At 4 days
post-transduction, transduction efficiency was measured by the
frequency of green fluorescent protein (GFP)-positive cells.
Positive stable transfectants were selected and expanded for
further study. The clones in which the COX-2 siRNA virus vector was
transfected were named the siCOX-2 group, and the negative control
vector-transfected cells were named the NC group.
Quantitative real-time PCR
To quantitatively determine the mRNA expression
levels of COX-2 and PCNA in the SGC-7901 and MKN-45 cell lines,
real-time PCR was used. Total RNA of each clone was extracted with
TRIzol according to the manufacturer’s protocol.
Reverse-transcription was carried out using M-MLV, and cDNA
amplification was carried out using the SYBR-Green Master Mix kit
according to the manufacturer’s protocol. Target genes were
amplified using specific oligonucleotide primers, and the human
β-actin gene was used as an endogenous control. The PCR primer
sequences were: COX-2, 5′-GAAGTACCAAGCTGTGCTTGAATAA-3′ and
5′-GGCTTGATTCCAATGCACCTA-3′; PCNA, 5′-CCATCCTCAAGAAGGTGTTGG-3′ and
5′-GTGTCCCATATCCGCAATTTTAT-3′; β-actin,
5′-ATGGGTCAGAAGGATTCCTATG-3′ and 5′-CAGCTCGTAGCTCTTCTCCA-3′. Data
were analyzed using the comparative Ct method (2−ΔΔCt).
Three separate experiments were performed for each clone.
Western blot assay
GAC cells were harvested and extracted using lysis
buffer (Tris-HCl, SDS, mercaptoethanol, glycerol). Cell extracts
were boiled for 5 min in loading buffer and then equal amounts of
cell extracts were separated on 15% SDS-PAGE gels. Separated
protein bands were transferred onto polyvinylidene fluoride (PVDF)
membranes, and the membranes were blocked in 5% skim milk powder.
The primary antibodies against COX-2 and PCNA were diluted
according to the instructions for use of the antibodies and
incubated overnight at 4°C. Then, horseradish peroxidase-linked
secondary antibodies were added at a dilution ratio of 1:1000, and
incubated at room temperature for 2 h. The membranes were washed
with PBS for three times, and the immunoreactive bands were
visualized using ECL Plus kit according to the manufacturer’s
instructions. The relative protein level in the different groups
was normalized to the GAPDH concentration. Three separate
experiments were performed for each clone.
Fluorescence microscopy
Twenty-four hours after infection, cells were plated
on glass coverslips, and 48 h post-infection the coverslips were
washed extensively in phosphate-buffered saline (PBS) and fixed
with 4% paraformaldehyde in PBS. After additional washing, the
cells were permeabilized with 1% Triton X-100 in PBS for 10 min.
The coverslips were then washed and blocked with 1% BSA for 30 min.
Cells were incubated in the appropriate primary antibody (COX-2 or
PCNA) overnight at 4°C. Samples were then washed and incubated with
species-specific secondary rhodamine-labeled antibodies (TRITC) in
PBS (1:100 dilution) for 60 min. Nuclei were stained with DAPI at
RT for 10 min and coverslips were mounted with Antifade solution
prior to imaging on a confocal microscope.
Cell proliferation assay
Cell proliferation was analyzed with the MTT assay.
Briefly, cells infected with the COX-2 siRNA virus or pretreated
with celecoxib were incubated in 96-well plates at a density of
1×105 cells/well with DMEM supplemented with 10% FBS.
Cells were treated with 20 μl MTT dye at 0, 24, 48 and 72 h and
then incubated with 150 μl of DMSO for 5 min. The color reaction
was measured at 570 nm with an enzyme immunoassay analyzer
(Bio-Rad, Hercules, CA, USA). The proliferation activity was
calculated for each clone.
Wound-healing assay
GAC cells were plated in each well of a 6-well
culture plate and allowed to grow to 90% confluency. Treatment with
COX-2 siRNA was then performed. On the next day, a wound was
created using a micropipette tip. The migration of cells towards
the wound was monitored daily, and images were captured at time
intervals of 24 h.
Transwell invasion assay
Transwell filters were coated with Matrigel (3.9
μg/μl, 60–80 μl) on the upper surface of a polycarbonic membrane
(diameter, 6.5 mm; pore size, 8 μm). After incubation at 37°C for
30 min, the Matrigel solidified and served as the extracellular
matrix for analysis of tumor cell invasion. Harvested cells
(1×105) in 100 μl of serum-free DMEM were added into the
upper compartment of the chamber. A total of 200 μl conditioned
medium derived from NIH3T3 cells was used as a source of
chemoattractant, and was placed in the bottom compartment of the
chamber. After 24 h of incubation at 37°C with 5% CO2,
the medium was removed from the upper chamber. The non-invaded
cells on the upper side of the chamber were scraped off with a
cotton swab. The cells that had migrated from the Matrigel into the
pores of the inserted filter were fixed with 100% methanol, stained
with hematoxylin, and mounted and dried at 80°C for 30 min. The
number of cells invading through the Matrigel were counted in three
randomly selected visual fields from the central and peripheral
portion of the filter using an inverted microscope (magnification,
×200). Each assay was repeated three times.
Subcutaneous tumor model and gene
therapy
Six-week-old female immune-deficient nude mice
(BALB/c-nu) were bred at the laboratory animal facility (Institute
of Chinese Academy of Sciences, Shanghai), and were housed
individually in microisolator ventilated cages with free access to
water and food. All experimental procedures were performed
according to the Regulations and Internal Biosafety and Bioethics
Guidelines of Shanghai Jiao Tong University and the Shanghai
Municipal Science and Technology Commission. Three mice were
injected subcutaneously with 1×108 GAC cells (MKN-45) in
50 μl of PBS pre-mixed with an equal volume of Matrigel matrix
(Becton Dickinson). Mice were monitored daily and developed
subcutaneous tumors. When the tumor size reached ~5 mm in length,
the tumors were surgically removed, cut into 1–2 mm3
pieces, and re-seeded individually into other mice. When the tumor
size reached ~5 mm in length, the mice were randomly assigned as
the NC group and the siCOX-2 group. In the treatment group, 15 μl
of siCOX-2 was injected into subcutaneous tumors using a multi-site
injection format. Injections were repeated every other day after
initial treatment. The tumor volume every three days was measured
with a caliper, using the formula: Volume = (length ×
width)2/2.
Statistical analysis
SPSS 20.0 was used for the statistical analysis.
Kruskal-Wallis H and Chi-square tests were used to analyze the
expression rate in all groups. One-way analysis of variance (ANOVA)
was used to analyze the differences between groups. The LSD method
of multiple comparisons was used when the probability for ANOVA was
statistically significant. Statistical significance was set at
P<0.05.
Results
Expression of COX-2 and PCNA in GAC
tissues
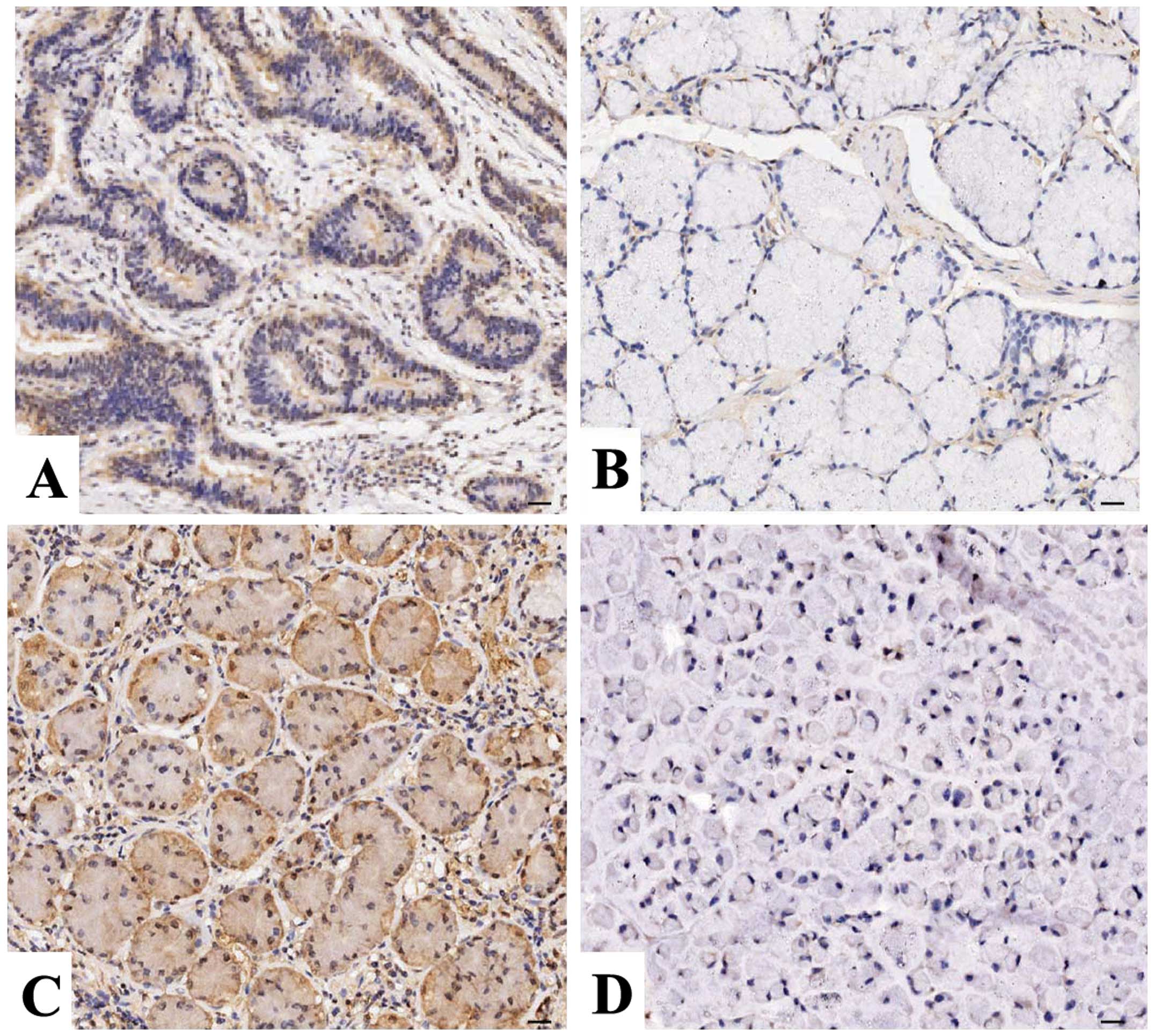
The expression of COX-2 and PCNA protein was
evaluated using IHC staining in GAC tissues. As shown in Fig. 1, different levels of positive
expression of COX-2 and PCNA protein were examined in the GAC
tissues. Positive COX-2 immunostaining was localized in the
cytoplasm, while PCNA was localized in the nucleus in the GAC
tissue cells. According to COX-2 and PCNA immunoreactive intensity,
the positive expression of COX-2 and PCNA was significantly
increased in the GAC tissues compared with these levels in the
ANCTs (P=0.011; P=0.047) (Table I).
Spearman rank correlation analysis also indicated a positive
correlation between COX-2 and PCNA expression in the GAC
tissues.
 | Table IExpression of COX-2 and PCNA protein
in the human GAC tissues. |
Table I
Expression of COX-2 and PCNA protein
in the human GAC tissues.
| | | Score | | | |
|---|
| | |
| | | |
|---|
| Target | Group | Total | − | + | ++ | +++ | Positive rate
(%) | χ2 | P-value |
|---|
| COX-2 | GAC | 45 | 9 | 18 | 11 | 7 | 80.0 | 6.409 | 0.011 |
| ANCT | 45 | 21 | 13 | 8 | 3 | 53.3 | | |
| PCNA | GAC | 45 | 14 | 17 | 9 | 5 | 68.9 | | |
| ANCT | 45 | 23 | 14 | 5 | 3 | 48.9 | 3.942 | 0.047 |
Association between COX-2 expression and
clinicopathological parameters
The relationship between COX-2 expression and
various clinical and pathological parameters of the GAC patients
was analyzed. As indicated in Table
II, no significant correlation was found between COX-2
expression and age, gender, tumor size or pathological TNM stage.
The cases were divided into two groups: those with and those
without lymph node metastasis. The rate of COX-2 expression was
higher in 55.6% (25/45) of the GAC patients with lymph node
metastasis than that in 24.4% (11/45) of the GAC patients without
lymph node metastasis (P=0.011).
| Table IICorrelation of COX-2 expression with
clinicopathologic parameters of the GAC patients. |
Table II
Correlation of COX-2 expression with
clinicopathologic parameters of the GAC patients.
| | COX-2 | | |
|---|
| |
| | |
|---|
| Variables | Cases (n) | − | + | χ2 | P-value |
|---|
| Total | 45 | 9 | 36 | | |
| Age (years) | | | | | |
| <60 | 24 | 6 | 18 | 0.786 | 0.375 |
| ≥60 | 21 | 3 | 18 | | |
| Gender | | | | | |
| Male | 32 | 5 | 27 | 1.296 | 0.255 |
| Female | 13 | 4 | 9 | | |
| TNM stage | | | | | |
| I+II | 31 | 7 | 24 | 0.406 | 0.524 |
| III+IV | 14 | 2 | 12 | | |
| Tumor size | | | | | |
| T1+T2 | 27 | 6 | 21 | 0.204 | 0.652 |
| T3+T4 | 18 | 3 | 15 | | |
| Lymph node
metastases | | | | | |
| No | 18 | 7 | 11 | 6.541 | 0.011 |
| Yes | 27 | 2 | 25 | | |
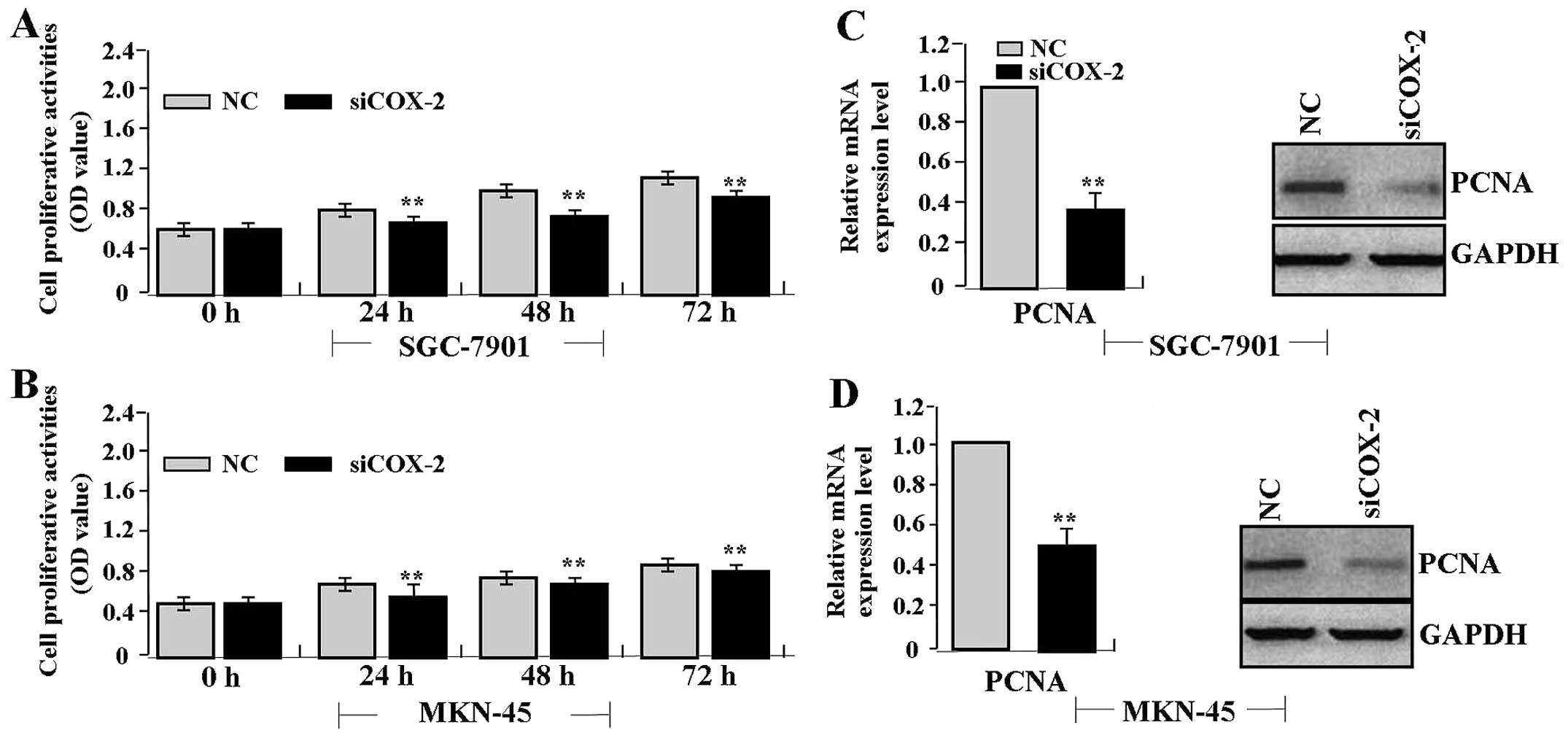
Effect of COX-2 siRNA on PCNA
expression
To examine the effect of siRNA-mediated COX-2
knockdown on PCNA expression in gastric cancer cells, SGC-7901 and
MKN-45 cells were infected with adenovirus-mediated COX-2 siRNA.
Quantitative real-time PCR was performed at 48 h recovery to
measure the mRNA expression levels. As shown in Fig. 2A, the mRNA expression levels of
COX-2 and PCNA were much lower in the siCOX-2 group than levels in
the NC group (each P<0.01). As for their protein expression
indicated by fluorescence microscopy (Fig. 2B), their expression levels were
decreased in the siCOX-2 group compared with the NC group.
Effect of COX-2 siRNA on cell
proliferation
To determine the effect of COX-2 siRNA on the
proliferative activities of gastric cancer cells, we investigated
the growth of SGC-7901 and MKN-45 cells by MTT assay. We found that
knockdown of COX-2 significantly diminished the proliferative
activities of gastric cancer cells in a time-dependent manner
compared with the NC group (each P<0.01) (Fig. 3A and B). In addition, to confirm the
effect of COX-2 siRNA on endogenous expression of PCNA, the
expression of PCNA was examined by real-time PCR and western blot
assays, indicating that, the amount of PCNA was significantly
decreased in the siCOX-2 group compared to the NC group (each
P<0.01) (Fig. 3C and D).
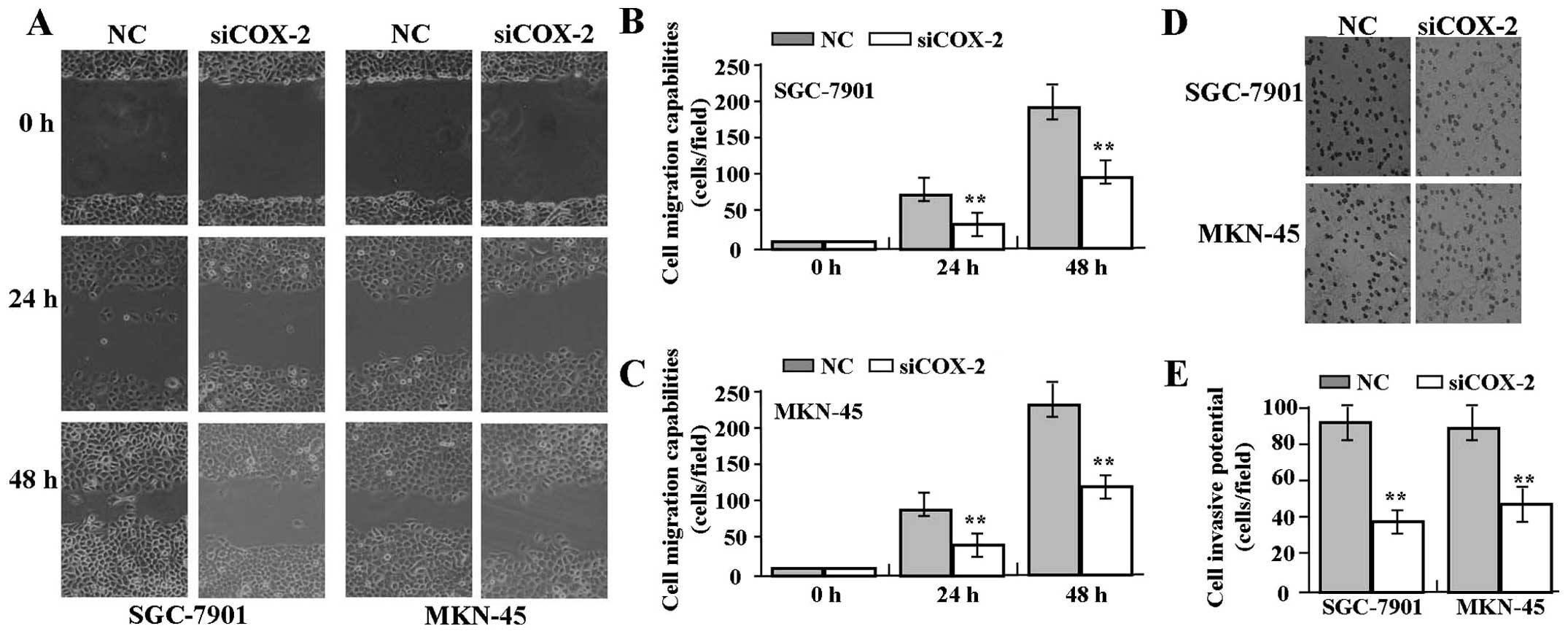
Effect of COX-2 siRNA on cell migration
and invasion
To determine the effect of COX-2 siRNA on cell
migration and invasion, wound-healing and Transwell assays were
performed. The results indicated that the migratory capability of
GAC cells in the siCOX-2 group was markedly decreased compared to
that in the NC group (Fig. 4A–C).
The invasive potential in the Transwell assay was determined on the
basis of the ability of cells to invade a matrix barrier containing
laminin and type IV collagen, the major components of the basement
membrane. Representative micrographs of Transwell filters are shown
in Fig. 4D. The invasive potential
of GAC cells was distinctly weakened in the siCOX-2 group compared
to that in the NC group (each P<0.01) (Fig. 4E).
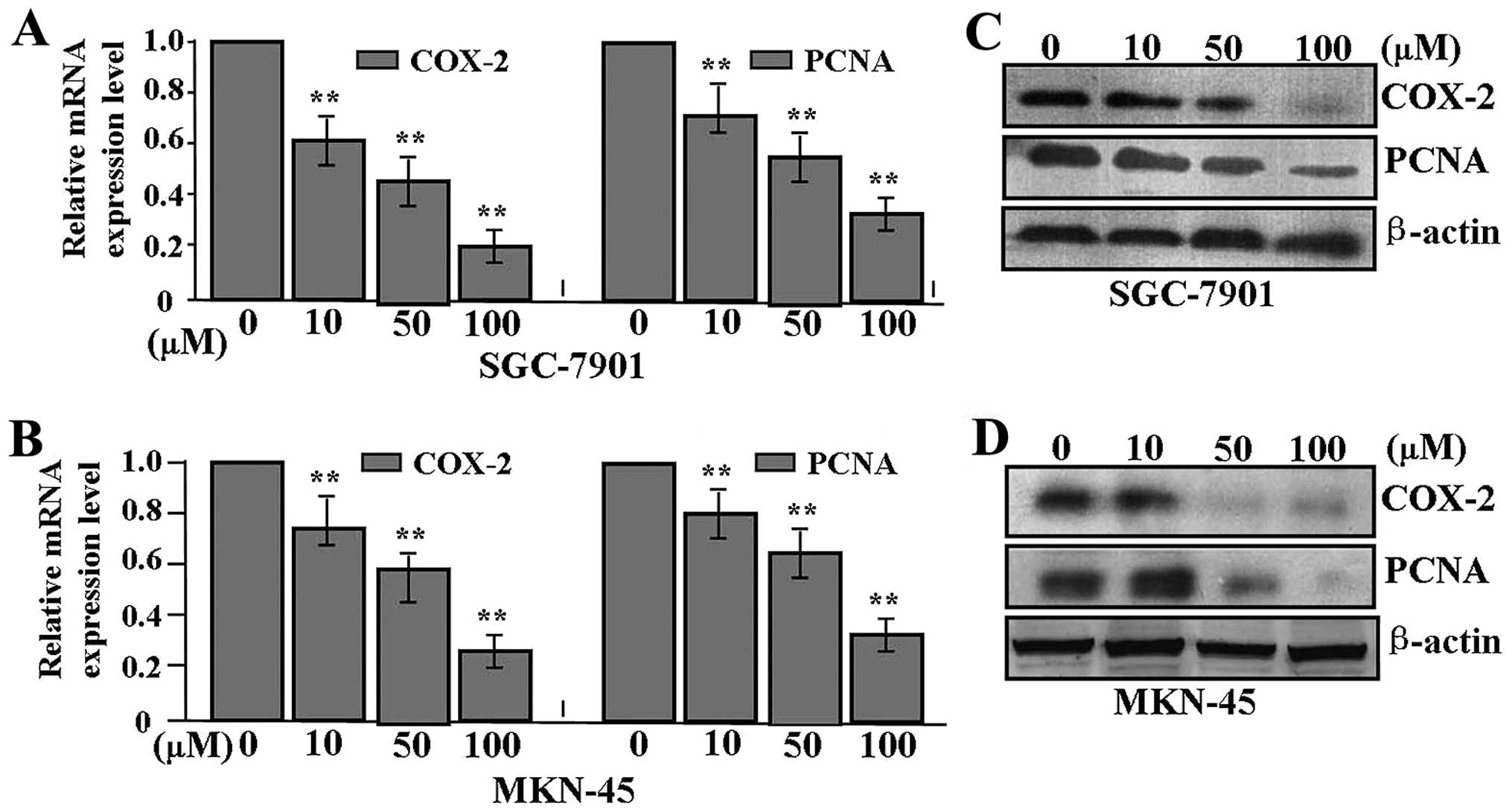
Effect of celecoxib on the expression of
COX-2 and PCNA
To examine the effect of celecoxib on the expression
of COX-2 and PCNA in GAC cells, SGC-7901 and MKN-45 cells were
pretreated with different concentrations of celecoxib. Quantitative
real-time PCR was performed at 48 h recovery to measure the mRNA
expression levels. As shown in Fig. 5A
and B, the mRNA expression levels of COX-2 and PCNA were
significantly downregulated in a dose-dependent manner in the
celecoxib-treated group compared with the untreated group (each
P<0.01). As for the protein expression indicated by western blot
assay (Fig. 5C and D), their
expression levels were also reduced in the siCOX-2 group when
compared with these levels in the NC group.
Effect of celecoxib on cell
proliferation
In order to evaluate the effect of celecoxib on cell
proliferation, we investigated the proliferative activities of GAC
cells by MTT assay. Celecoxib significantly diminished the
proliferative activities of GAC cells in a dose- and time-dependent
manner in comparison with the NC group (Table III), suggesting that inhibition of
COX-2 by celecoxib inhibited the proliferation of GAC cells.
| Table IIIEffect of celecoxib on the
proliferative activities of GAC cells (OD values). |
Table III
Effect of celecoxib on the
proliferative activities of GAC cells (OD values).
| 24 h | 48 h | 72 h |
|---|
|
|
|
|
|---|
| Group | SGC-7901 | MKN-45 | SGC-7901 | MKN-45 | SGC-7901 | MKN-45 |
|---|
| CON | 0.49±0.03 | 0.51±0.04 | 0.51±0.10 | 0.58±0.08 | 0.68±0.13 | 0.71±0.09 |
| 10 μM | 0.46±0.03 | 0.48±0.02 | 0.38±0.03a | 0.42±0.03a | 0.42±0.04a | 0.52±0.05a |
| 50 μM | 0.38±0.04a | 0.40±0.03a | 0.34±0.04a | 0.37±0.04a | 0.18±0.02a | 0.26±0.03a |
| 100 μM | 0.19±0.05a | 0.23±0.03a | 0.12±0.01a | 0.21±0.02a | 0.09±0.02a | 0.18±0.04a |
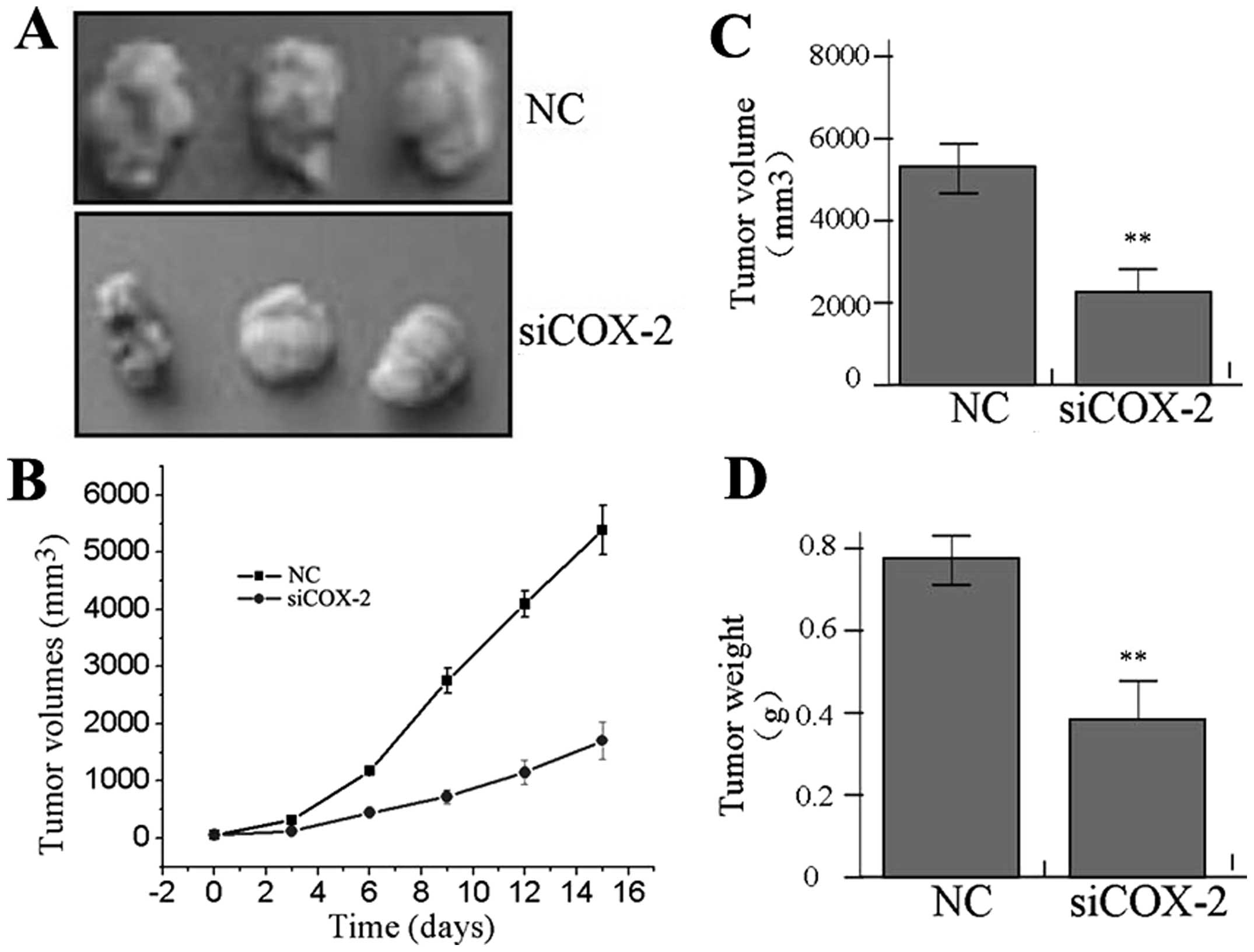
Effect of COX-2 knockdown on xenograft
tumor growth
Our in vitro experiments demonstrated the
suppressive effect of COX-2 inhibition on tumor growth. Therefore,
we further investigated the effect of COX-2 siRNA on xenograft
tumor growth in vivo. The mean volume of tumors in the
experimental mice before treatment was 83.22±21.45 mm3.
During the entire tumor growth period, the tumor growth activity
was measured. The tumors treated with siCOX-2 grew substantially
slower compared to the NC group (Fig.
6A and B). When the tumors were harvested, the average weight
and volume of the tumors in the siCOX-2 group were significantly
reduced than those of the NC group (Fig. 6C and D), suggesting that COX-2
knockdown suppressed the growth of the gastric cancer cells.
Discussion
Gastric cancer is a common cancer in China. The
mechanisms of gastric carcinogenesis are not completely known, but
the molecular biology of cancer suggests that the initiation and
progression of gastric cancer are a consequence of a cumulative
series of multiple gene alterations. Studies indicate that COX-2,
as an early alteration in cancer, may be involved in gastric
carcinogenesis, and may correlate with the depth of cancer
(11). COX-2 is preferentially
upregulated in Barrett’s esophagus and intestinal-type gastric
cancer, and may play an important role in the development of
inflammation-related GAC (12). In
addition, COX-2 was found to be overexpressed in GAC and to be
associated with the high abundance of vascular endothelial growth
factor-C (VEGF-C) and lymphatic metastasis (13), suggesting that COX-2 is an
independent prognostic factor for human GAC (14).
However, Yamac et al (15) showed that COX-2 expression is
inversely correlated with tumor size, TNM stage, and lymph node
status of gastric carcinoma. To clarify the role of COX-2 in GAC,
in the present study, positive expression of COX-2 was found in the
cellular cytoplasm of cancer tissues with a higher strong
reactivity rate, compared with the ANCTs, increasing with tumor
malignancy, suggesting that the cytoplasmic accumulation of COX-2
might be involved in the development of GAC. In addition, our
findings showed that COX-2 expression was positively associated
with lymph node metastasis of GAC patients, and provide the basis
for further cell functional experiments. The highly invasive GAC
cell lines (SGC-7901 and MKN-45) with high COX-2 expression were
chosen for the functional study. Using a loss-of-function
experiment, we found that knockdown of COX-2 by siRNA inhibited the
proliferation, migration and invasion of GAC cells in vitro
and in vivo. Studies have previously demonstrated that COX-2
is overexpressed in GAC, and targeting COX-2 exerts inhibitory
effects on malignant tumor cell proliferation, invasion and
metastasis, suggesting that COX-2 may be an important therapeutic
target for malignant tumors (16,17).
Since upregulation of COX-2 has been reported in
esophageal and gastric cancers of different stages, COX-2 selective
inhibitors have been considered as an excellent alternative to
treat gastrointestinal tumors (18). Hu et al (19) examined the chemopreventive effect of
a COX-2 inhibitor on stomach carcinogenesis, and found that
treatment with celecoxib reduced gastric cancer incidence and
growth in rats. A combination of rofecoxib with octreotide
significantly enhanced the anti-proliferative effect in GAC
(20), while combining S-1 and
COX-2 inhibitor administration achieved a synergistic inhibitory
effect on gastric cancer metastasis (21). Moreover, COX-2 is associated with an
increased risk of gastric cancer, particularly interacting with
H. pylori infection (22)
and can be activated by H. pylori infection in gastric
cancer cells in vitro and in vivo (23). Our present study indicated that
celecoxib as a COX-2 inhibitor suppressed the expression of COX-2
and the proliferation of GAC cells, suggesting that celecoxib may
exert an antitumor effect dependent on the COX-2 pathway in gastric
cancer. These findings indirectly provide experimental evidence
that celecoxib may inhibit the H. pylori-induced development
of gastric cancer through blockade of the COX-2 pathway, which has
also been suggested in another study (24).
The PCNA labeling index not only represents the
tumor proliferative activity, but is also correlated with lymph
node invasion and metastasis in gastrointestinal carcinoma
(25). It is also related to
decreased apoptosis and increased proliferation in gastric
carcinoma cells, and may be a prognostic factor in advanced gastric
carcinoma (26). However, PCNA is
not associated with cell proliferation in some forms of neoplasia,
including breast and gastric cancer and in cell lines (27). Of note, in the present study, our
findings showed that PCNA expression was significantly higher in
the cellular nucleus of GAC tissues than that in the ANCTs,
suggesting that nuclear accumulation of PCNA may be associated with
the development of GAC. Spearman rank correlation analysis also
indicated a positive correlation between COX-2 and PCNA expression
in GAC tissues. According to another study (25), the anticancer effects of celecoxib
on gastric cancer were found to be mediated by cell cycle arrest
and apoptosis, and not by COX-2 suppression alone. Furthermore, we
chose the poorly differentiated GAC cell lines (SGC-7901 and
MKN-45) with high COX-2 and PCNA expression for the functional
study. Our findings demonstrated that COX-2 inhibition by siRNA or
celecoxib suppressed the expression of PCNA, and the proliferation
and invasion of GAC cells, suggesting that celecoxib may exert
antitumor effects on gastric cancer via specific blockade of the
COX-2/PCNA signaling pathway.
In conclusion, our findings showed that COX-2 and
PCNA are highly expressed in GAC, and elevated expression of COX-2
correlates with lymph node metastasis of GAC patients. COX-2
inhibition by siRNA or celecoxib suppressed the proliferation,
migration and invasion of GAC cells. These data offer a strong
pre-clinical rationale to target COX-2 signaling as a therapeutic
strategy to improve the treatment of gastric adenocarcinoma.
References
|
1
|
Parkin DM, Bray F, Ferlay J, et al: Global
cancer statistics, 2002. CA Cancer J Clin. 55:74–108. 2005.
View Article : Google Scholar
|
|
2
|
Tong QS, Zheng LD, Wang L, et al:
Downregulation of XIAP expression induces apoptosis and enhances
chemotherapeutic sensitivity in human gastric cancer cells. Cancer
Gene Ther. 12:509–514. 2005.PubMed/NCBI
|
|
3
|
Saukkonen K, Rintahaka J, Sivula A, et al:
Cyclooxygenase-2 and gastric carcinogenesis. APMIS. 111:915–925.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tatsuguchi A, Matsui K, Shinji Y, et al:
Cyclooxygenase-2 expression correlates with angiogenesis and
apoptosis in gastric cancer tissue. Hum Pathol. 35:488–495. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yasuda H, Yamada M, Endo Y, et al:
Elevated cyclooxygenase-2 expression in patients with early gastric
cancer in the gastric pylorus. J Gastroenterol. 40:690–697. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mrena J, Wiksten JP, Thiel A, et al:
Cyclooxygenase-2 is an independent prognostic factor in gastric
cancer and its expression is regulated by the messenger RNA
stability factor HuR. Clin Cancer Res. 11:7362–7368. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ji J, Zhao P and Huang B: Study of gastric
carcinoma and PCNA and c-met gene abnormality. Wei Sheng Yan Jiu.
37:479–482. 2008.(In Chinese).
|
|
8
|
Czyzewska J, Guzińska-Ustymowicz K, Lebelt
A, et al: Evaluation of proliferating markers Ki-67, PCNA in
gastric cancers. Rocz Akad Med Bialymst. 49:64–66. 2004.PubMed/NCBI
|
|
9
|
Czyzewska J, Guzińska-Ustymowicz K,
Pryczynicz A, et al: Immunohistochemical evaluation of Ki-67, PCNA
and MCM2 proteins proliferation index (PI) in advanced gastric
cancer. Folia Histochem Cytobiol. 47:289–296. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lu B, Yu L, Xu L, et al: The effects of
radix curcumae extract on expressions of VEGF, COX-2 and PCNA in
gastric mucosa of rats fed with MNNG. Curr Pharm Biotechnol.
11:313–317. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Forones NM, Kawamura KY, Segreto HR, et
al: Expression of COX-2 in stomach carcinogenesis. J Gastrointest
Cancer. 39:4–10. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sonoda R, Naomoto Y, Shirakawa Y, et al:
Preferential up-regulation of heparanase and cyclooxygenase-2 in
carcinogenesis of Barrett’s oesophagus and intestinal-type gastric
carcinoma. Histopathology. 57:90–100. 2010.PubMed/NCBI
|
|
13
|
Liu J, Yu HG, Yu JP, et al: Overexpression
of cyclooxygenase-2 in gastric cancer correlates with the high
abundance of vascular endothelial growth factor-C and lymphatic
metastasis. Med Oncol. 22:389–397. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gou HF, Chen XC, Zhu J, et al: Expressions
of COX-2 and VEGF-C in gastric cancer: correlations with
lymphangiogenesis and prognostic implications. J Exp Clin Cancer
Res. 30:142011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yamac D, Ayyildiz T, Coşkun U, et al:
Cyclooxygenase-2 expression and its association with angiogenesis,
Helicobacter pylori, and clinicopathologic characteristics
of gastric carcinoma. Pathol Res Pract. 204:527–536. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang J, Zhang QY, Fu YC, et al:
Expression of p-Akt and COX-2 in gastric adenocarcinomas and
adenovirus mediated Akt1 and COX-2 ShRNA suppresses SGC-7901
gastric adenocarcinoma and U251 glioma cell growth in vitro and in
vivo. Technol Cancer Res Treat. 8:467–478. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chan MW, Wong CY, Cheng AS, et al:
Targeted inhibition of COX-2 expression by RNA interference
suppresses tumor growth and potentiates chemosensitivity to
cisplatin in human gastric cancer cells. Oncol Rep. 18:1557–1562.
2007.PubMed/NCBI
|
|
18
|
Jiménez P, García A, Santander S and
Piazuelo E: Prevention of cancer in the upper gastrointestinal
tract with COX-inhibition. Still an option? Curr Pharm Des.
13:2261–2273. 2007.PubMed/NCBI
|
|
19
|
Hu PJ, Yu J, Zeng ZR, et al:
Chemoprevention of gastric cancer by celecoxib in rats. Gut.
53:195–200. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tang C, Liu C, Zhou X, et al: Enhanced
inhibitive effects of combination of rofecoxib and octreotide on
the growth of human gastric cancer. Int J Cancer. 112:470–474.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tendo M, Yashiro M, Nakazawa K, et al: A
synergic inhibitory-effect of combination with selective
cyclooxygenase-2 inhibitor and S-1 on the peritoneal metastasis for
scirrhous gastric cancer cells. Cancer Lett. 244:247–251. 2006.
View Article : Google Scholar
|
|
22
|
Zhang X, Zhong R, Zhang Z, et al:
Interaction of cyclooxygenase-2 promoter polymorphisms with
Helicobacter pylori infection and risk of gastric cancer.
Mol Carcinog. 50:876–883. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sierra JC, Hobbs S, Chaturvedi R, et al:
Induction of COX-2 expression by Helicobacter pylori is
mediated by activation of epidermal growth factor receptor in
gastric epithelial cells. Am J Physiol Gastrointest Liver Physiol.
305:G196–G203. 2013.PubMed/NCBI
|
|
24
|
Lan C, Yang L, Fan L, et al: Celecoxib
inhibits Helicobacter pylori-induced invasion of gastric
cancer cells through an adenine nucleotide translocator-dependent
mechanism. Anticancer Agents Med Chem. 13:1267–1272.
2013.PubMed/NCBI
|
|
25
|
Kunimoto Y, Nakamura T, Ohno M, et al:
Relationship between immunohistochemical evaluation of thymidylate
synthase and proliferating cell nuclear antigen labeling index in
gastrointestinal carcinoma. Oncol Rep. 12:1163–1167. 2004.
|
|
26
|
Tao K, Chen D, Tian Y, et al: The
relationship between apoptosis and the expression of proliferating
cell nuclear antigen and the clinical stages in gastric carcinoma.
J Tongji Med Univ. 20:222–224. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hall PA, Levison DA, Woods AL, et al:
Proliferating cell nuclear antigen (PCNA) immunolocalization in
paraffin sections: an index of cell proliferation with evidence of
deregulated expression in some neoplasms. J Pathol. 162:285–294.
1990. View Article : Google Scholar : PubMed/NCBI
|