Introduction
Endometrial cancer has become the most common female
genital tract tumor in some western countries (1,2) and
accounts for 70–80% of all primary uterine malignancies in the
United States (3,4). The incidence of endometrial cancer is
continuously rising worldwide. There are mainly 2 types of
endometrial cancers based on their histopathological
characteristics (5); type I
endometrial cancers, which account for 80% of endometrial cancers,
are the endometrioid endometrial adenocarcinomas. They are
generally estrogen-related, low grade and with a relatively high
survival rate (~85% at 5 years). Surgery remains the cornerstone
treatment for patients with type I endometrial cancer (6). Type II endometrial cancers develop
mainly in elderly and post-menopausal women. They are usually
hormone-independent and belong to high grade endometrioid
adenocarcinomas, papillary serous and clear cell carcinomas and
carcinosarcomas. Compared with type I endometrial cancer, the
prognosis of type II endometrial cancer is poorer due to its high
invasiveness and metastasis. In addition, 50% of all relapses occur
in patients with type II endometrial cancer (7). The molecular mechanism underlying the
initiation and progression of type II endometrial cancer remains
unclear.
MicroRNAs (miRNAs) are a class of non-protein-coding
small RNAs containing ~18–24 nucleotides. MiRNAs have been shown to
regulate gene expression by complementarily binding to the 3′
untranslated region (UTR) of target mRNAs, leading to the
degradation of target genes or the reduction of gene translation
(8). Numerous studies have shown
that miRNAs are involved in the regulation of several key
biological processes, including proliferation, apoptosis,
differentiation and metabolism. Emerging evidence suggests that
miRNA dysregulation is associated with the initiation and
progression of multiple human cancers, including lymphoma, gastric,
prostate and pancreatic cancer (9,10).
Depending on the downstream targets, miRNAs can function either as
oncogenes or as tumor suppressors (11,12).
MicroRNA-449a (MiR-449a) has been demonstrated to
inhibit cancer cell proliferation in multiple cancers, such as
breast, prostate, bladder, liver, gastric and lung cancer, by
targeting a broad spectrum of molecules involved in cell cycle
regulation and apoptosis (13–18).
MiR-449a expression in these tumor tissues is reduced or lost
(13–18). However, the role of miR-449a in
endometrial cancer has not been clarified. In this study, we
examined the expression of miR-449 in normal endometrium, type I
and type II endometrial cancer tissues and further investigated the
effects of exogenous overexpression of miR-449a on the behavior of
endometrial cancer cells.
Materials and methods
Patient tissue specimen
This study was approved by the Institutional Ethics
Committee of the First Affiliated Hospital of Wenzhou Medical
University. All patients signed the informed consent form. The
patients with endometrial cancers underwent abdominal hysterectomy
in our department. Surgical tissue specimens from 2 patients with
type I endometrial cancer, 2 patients with type II endometrial
cancer and 2 patients undergoing uterine fibroids hysterectomy were
obtained. The diagnosis for endometrial cancer was confirmed by
histopathological examination. The benign endometrial tissue
specimens were confirmed to be free of malignant lesions. The
endometrium was dissected. The cancerous lesions from endometrium
were removed and used for the determination of miR-449 levels.
Small pieces of the lesions were sectioned and stained with H&E
to further confirm that >90% of the cells in the lesions were
cancer cells.
Immunohistochemical staining for estrogen
and progesterone receptor
Endometrial tissues were sectioned and fixed in 4%
PFA. After blocking overnight, tissue sections were then incubated
with primary antibodies to estrogen receptor (ER; 1:100, Abcam,
USA) or progesterone receptor (PR; 1:100, Abcam, USA) overnight,
then washed and incubated with secondary antibody. Tissue sections
stained with secondary antibody only were used as the negative
control. The stained tissue section was observed under a microscope
at a magnification of ×100.
Determination of miR-449 level in tissue
specimens
Total RNA was extracted from tissues by the TRIzol
method and 100 ng total RNA was used for reverse transcription
(RT). The primer sequences for reverse transcription of miR-449a
and miR-449b are listed in Table I.
The total volume of the RT reaction was 20 μl. The RT conditions
were: incubation at 16°C for 10 min, 37°C for 30 min and 65°C for 5
min. The cDNA products were used as the templates for quantitative
PCR. The primer sequences for PCR are shown in Table I. U6 was used as the housekeeping
gene. The forward primer for U6 was: 5′-CTCGCTTCGGCAGCACA-3′; the
reverse primer for U6 was: 5′-AACGCTTCACGAATTTGCGT-3′. The volume
of PCR reaction was 20 μl. The PCR conditions were: incubation at
95°C for 10 min, 95°C for 15 sec, 60°C for 1 min, 40 cycles. The
PCR instrument was ABI 7900 HT Fast Real-Time PCR system. Cycle
threshold of each sample was determined.
 | Table IRT and PCR primer sequences for
miR-449a and miR-449b. |
Table I
RT and PCR primer sequences for
miR-449a and miR-449b.
| miR-449a | miR-449b |
|---|
| Mature sequence |
TGGCAGTGTATTGTTAGCTGGT |
AGGCAGTGTATTGTTAGCTGGC |
| Stem-loop |
GTCGTATCCAGTGCAGGGTCCG |
GTCGTATCCAGTGCAGGGTCCG |
| RT primer |
AGGTATTCGCACTGGATACGACACCAGC |
AGGTATTCGCACTGGATACGACGCCAGC |
| Universal sense
primer | GTGCAGGGTCCGAGGT | GTGCAGGGTCCGAGGT |
| Specific antisense
primer |
CGCTGGCAGTGTATTGTTAGCT |
TGGGAGGCAGTGTATTGTTAGC |
Cell culture
The endometrial cancer cell line (HEC-1B) was
purchased from Shanghai Institute of Cell Biology (Shanghai,
China). The cells were maintained in Dulbecco’s modified Eagle’s
medium (DMEM; Gibco, USA) supplemented with 10% fetal bovine serum
(FBS; Gibco) in a humidified incubator at 37°C with 5%
CO2.
miRNA transfection
The sense sequence of hsa-miR-449a mimics was:
5′-UGGCAGUGUAUUGUUAGCUGGU3′ and the antisense sequence was:
5′-CAGCUAACAAUACACUGC CAUU-3′. The sequences for miRNA negative
control (NC) were: 5′-UUCUCCGAACGUGUCACGUTT-3′ for sense and
5′-ACGUGACACGUUCGGAGAATT-3′ for antisense. All the RNA
oligonucleotides were purchased from GenePharma (Shanghai, China).
HEC-1B cells were seeded into 6-well plates at a density of
3×105 cells/well or into 96-well plates at a density of
2.5×103 cells/well 24 h before transfection. After
reaching 30–50% confluence, the cells were transfected with miRNA
duplexes using Lipofectamine 2000 (Invitrogen, Shanghai) at a final
concentration of 50 nM. The hsa-miR-449a mimics labeled with 6-FAM
at the 5′-end were used to determine the transfection
efficiency.
Cell proliferation assay
HEC-1B cells were seeded into a 96-well plate at a
density of 2.5×103 cells/well and transfected the next
day. At 48 h after transfection, cell proliferation was analyzed
using the Cell Counting Kit-8 (CCK-8) assay (Beyotime, China).
Briefly, 10 μl CCK-8 solution was added to each well and incubated
at 37°C for 1–2 h. The absorbance at 450 nm was measured using a
microplate reader.
Colony formation assay
HEC-1B cells were harvested 24 h after transfection,
re-suspended in DMEM supplemented with 10% FBS and seeded in 6-well
plates at a density of 800 cells/well. The cells were cultured
under standard culture conditions for 10 days. The cells were then
fixed with methanol for 15 min and stained with crystal violet for
10–30 min.
Determination of apoptosis by flow
cytometry
HEC-1B cells were harvested 48 h after transfection,
washed twice with cold phosphate-buffered saline (PBS) and
re-suspended in 100 μl Annexin-binding buffer at the concentration
of 2×105–1×106cells/ml. Annexin V and PI
double-staining was performed according to the manufacturer’s
protocol. The Annexin V-FITC apoptosis detection kit was purchased
from Invitrogen. Cell apoptosis was analyzed within 1 h after
staining using the flow cytometer system with FACSDiva software
(FACSCalibur).
Transwell invasion assay
HEC-1B cells were harvested 24 h after transfection
and re-suspended in serum-free DMEM medium at the concentration of
1.5×105/ml. A total of 200 μl cell suspension was added
into the Matrigel-coated Transwell chambers (BD Biosciences, USA)
and 500 μl DMEM supplemented with 10% FBS was added into the lower
chamber of each well. The number of penetrating cells was
determined after 24 h incubation.
RNA extraction and real-time quantitative
PCR (qPCR)
Total RNA was extracted from HEC-1B cells 48 h after
transfection using TRIzol reagent (Invitrogen, Shanghai) according
to the manufacturer’s instructions. Briefly, 500 ng of total RNA
containing miRNA was polyadenylated by poly(A) polymerase and then
reverse transcribed to cDNA using miRNA First-Strand cDNA Synthesis
and qRT-PCR kits (Invitrogen). The cDNA was then used as the
template for SYBR real-time PCR using SYBR-Green PCR Master mix
(Toyobo, Osaka, Japan). The hsa-miR-449a specific qPCR reverse
primer was provided by the cDNA Synthesis and qRT-PCR kits. The
specific forward primer was designed for qPCR according to the
manufacturer’s protocol. Human U6 was used as the housekeeping
control. For CDC25A mRNA qPCR, cDNA was synthesized by using qPCR
RT kit (Toyobo) and human GAPDH was used as housekeeping gene for
normalization. The sequence of the forward primer for hsa-miR-449a
was: 5′-TGG CAG TGT ATT GTT AGC TGG T-3′, and for U6 was: 5′-CGC
AAG GAT GAC ACG CAA ATT C-3′. The sequence of the forward primer
for CDC25A mRNA was: 5′-TAC CTA CTG ATG GCA AGC GTG T-3′, and the
sequence for the reverse primer was: 5′-GAC TGG CAT TTC ATA AAG AAC
TCC T-3′. The sequence of GAPDH forward primer was: 5′-AGA AGG CTG
GGG CTC ATT TG-3′, and the sequence of GAPDH reverse primer was:
5′-AGG GGC CAT CCA CAG TCT TC-3′. The qPCR was performed on the
Applied Biosystems 7500 detection system. The 10 μl PCR system
contained 1 μl template (diluted 1:10), 5 μl SYBR-Green I Master
Mix, 1 μl specific forward primer of miR-449a/CDC25A mRNA (1 μM), 1
μl reverse primer (1 μM), 1 μl plus solution and 1 μl RNase and
DNase free water. The PCR reaction conditions were: incubation at
95°C for 3 min, 95°C for 15 sec, 60°C for 1 min and 40 cycles.
Cycle threshold (Ct) values for all samples were determined. The
ΔΔCt method was used to compare the expression of miR-449a and
CDC25A mRNA in each group.
Western blot analysis
Forty-eight hours after transfection, total protein
was extracted using a cell lysing buffer containing 1 mol/l
Tris-HCl (PH 6.8), glycerin, SDS, DTT (Sangon Biotech, China) and
ddH2O. Fifty-microgram of protein was loaded in the gel.
After transfer, the blot was probed with CDC25A (1:500) and GAPDH
(1:1000) antibody (Abcam, USA) and visualized by the secondary
antibodies.
Statistical analysis
Statistical analysis of all data was performed using
the software SPSS 17.0 (SPSS, Inc.). The differences between groups
were analyzed using the Student’s t-test when two groups were
analyzed or assessed by one-way analysis of variance (ANOVA) when
three or more groups were compared. All test results were two-sided
and the significance level was set at P<0.05. The data are
presented as the mean ± standard error (SE).
Results
The levels of miR-449a and miR-449b are
markedly reduced in type II endometrial cancer tissues
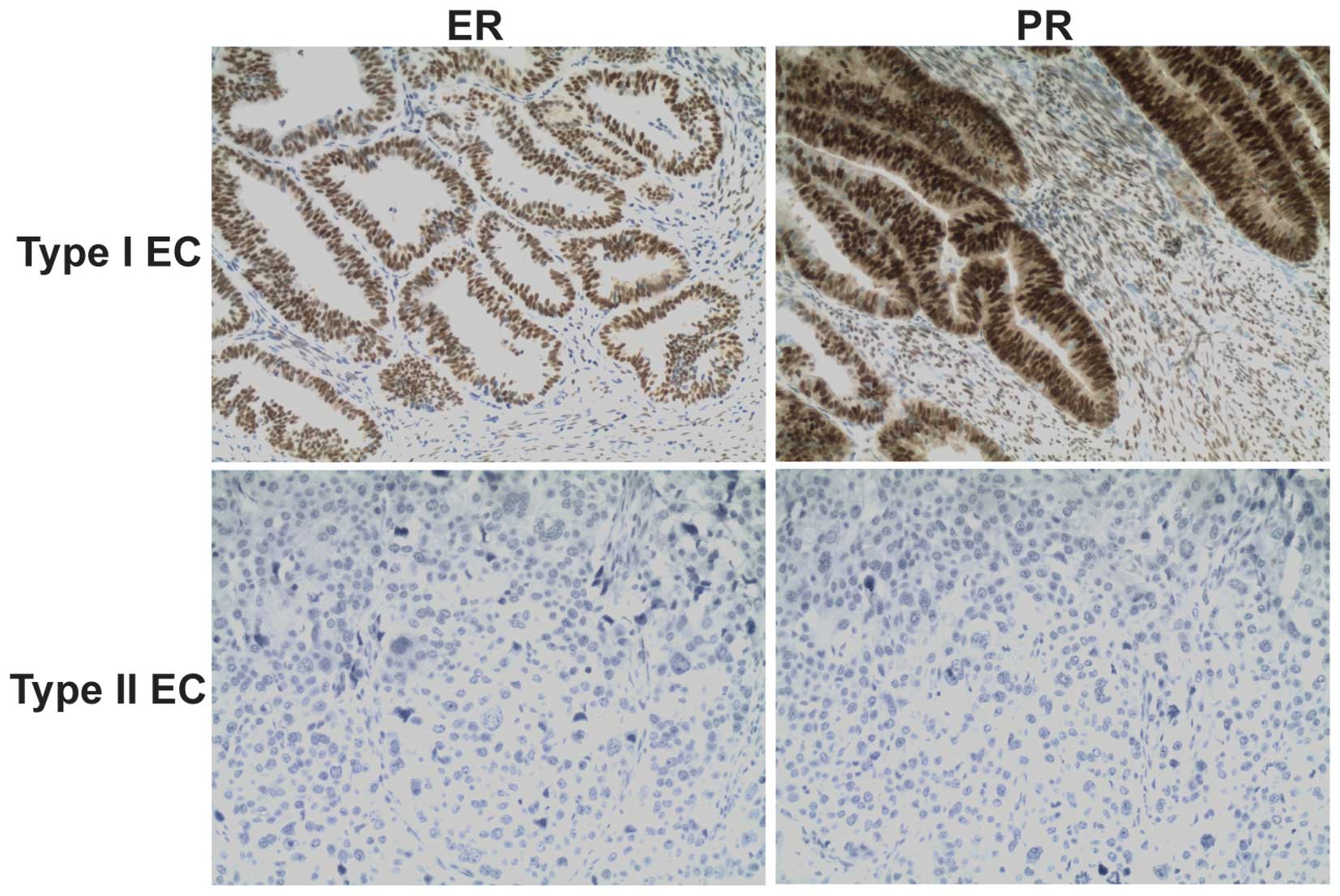
Immunohistochemical staining for ER and PR showed
that the type I endometrial cancer tissues were ER+ and
PR+ and the type II endometrial cancer tissues were
ER− and PR− (Fig.
1). Our RT-PCR results demonstrated that miR-449a and miR-449b
levels in type II endometrial cancer tissues were markedly reduced
by 288 and 424-fold, respectively, compared with benign endometrial
tissues and type I endometrial cancer tissues (Table II). Type I endometrial cancer
tissues had similar levels of miR-449a and miR-449b as the benign
tissues.
| Table IICycle threshold of RT-PCR for miR-449a
and miR-449b amplification. |
Table II
Cycle threshold of RT-PCR for miR-449a
and miR-449b amplification.
| Sample type | miR-449a | miR-449b | U6 |
|---|
| Benign | 21.97±0.59 | 21.80±0.53 | 18.73±0.50 |
| Type I endometrial
cancer | 22.17±0.90 | 22.15±0.70 | 19.20±0.36 |
| Type II endometrial
cancer | 30.39±0.03 | 30.78±0.30 | 18.98±0.77 |
Overexpression of miR-449a in HEC-1B
cells inhibits cell proliferation and invasion
To further investigate the role of miR-449a in
regulating the behavior of endometrial cancer cells, we
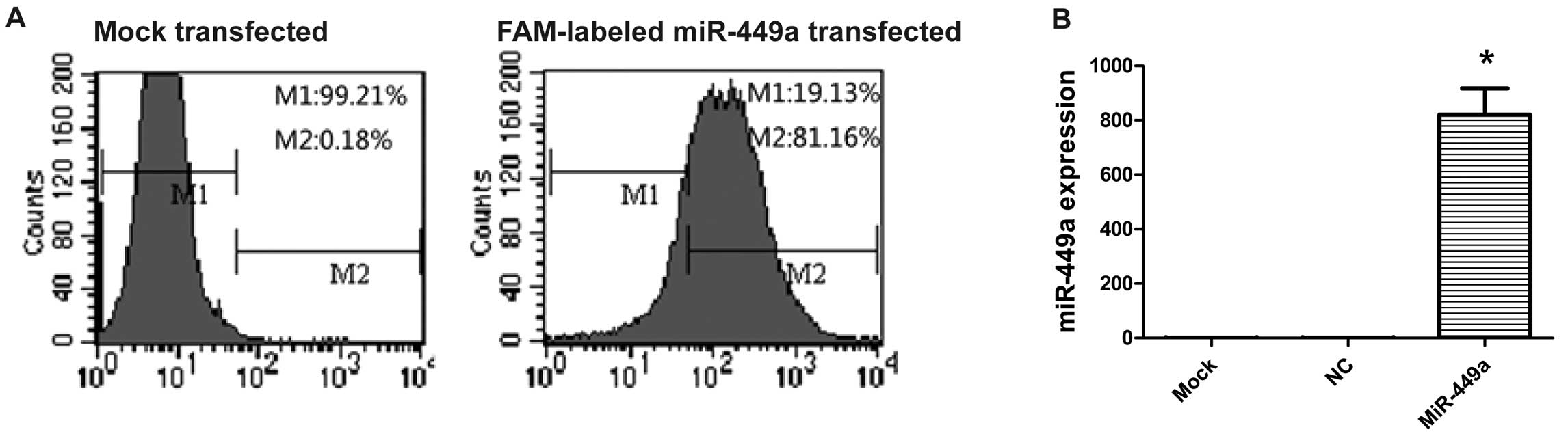
overexpressed miR-449a in HEC-1B cells. We first determined the
miR-449a transfection efficiency. Flow cytometry showed 88.7% of
cells carrying FAM-labeled miR-449a 24 h after transfection
(Fig. 2A). The expression of
miR-449a was further confirmed by RT-PCR. The level of miR-449a
increased significantly in cells transfected with miR-449a compared
with NC or mock-transfected cells (Fig.
2B). The endogenous miR-449a level in HEC-1B cells was very
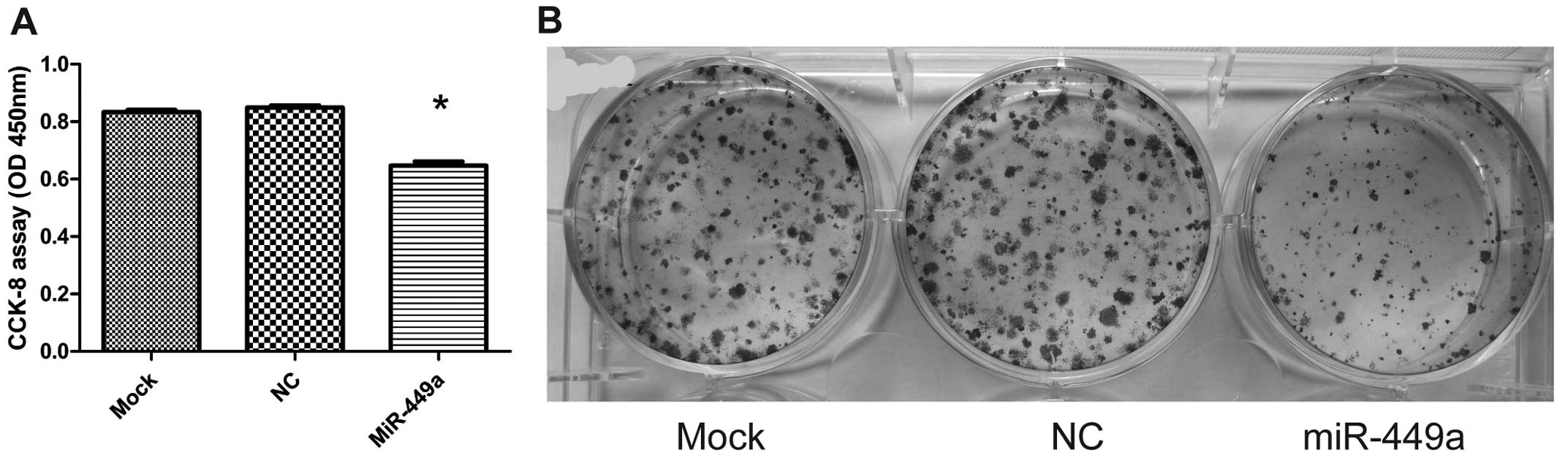
low. Transfection of miR-449a reduced cell proliferation
significantly (0.834±0.016 or 0.850±0.015 for mock or NC
transfection vs. 0.648±0.028 for miR-449a transfection, P<0.01;
Fig. 3A). To further confirm the
inhibitory effect of miR-449a on HEC-1B cell proliferation, we
performed colony formation assay. Consistently, our results showed
that overexpression of miR-449a suppressed clonogenicity of HEC-1B
cells (Fig. 3B). In addition to
inhibiting cell proliferation, overexpression of miR-449a also
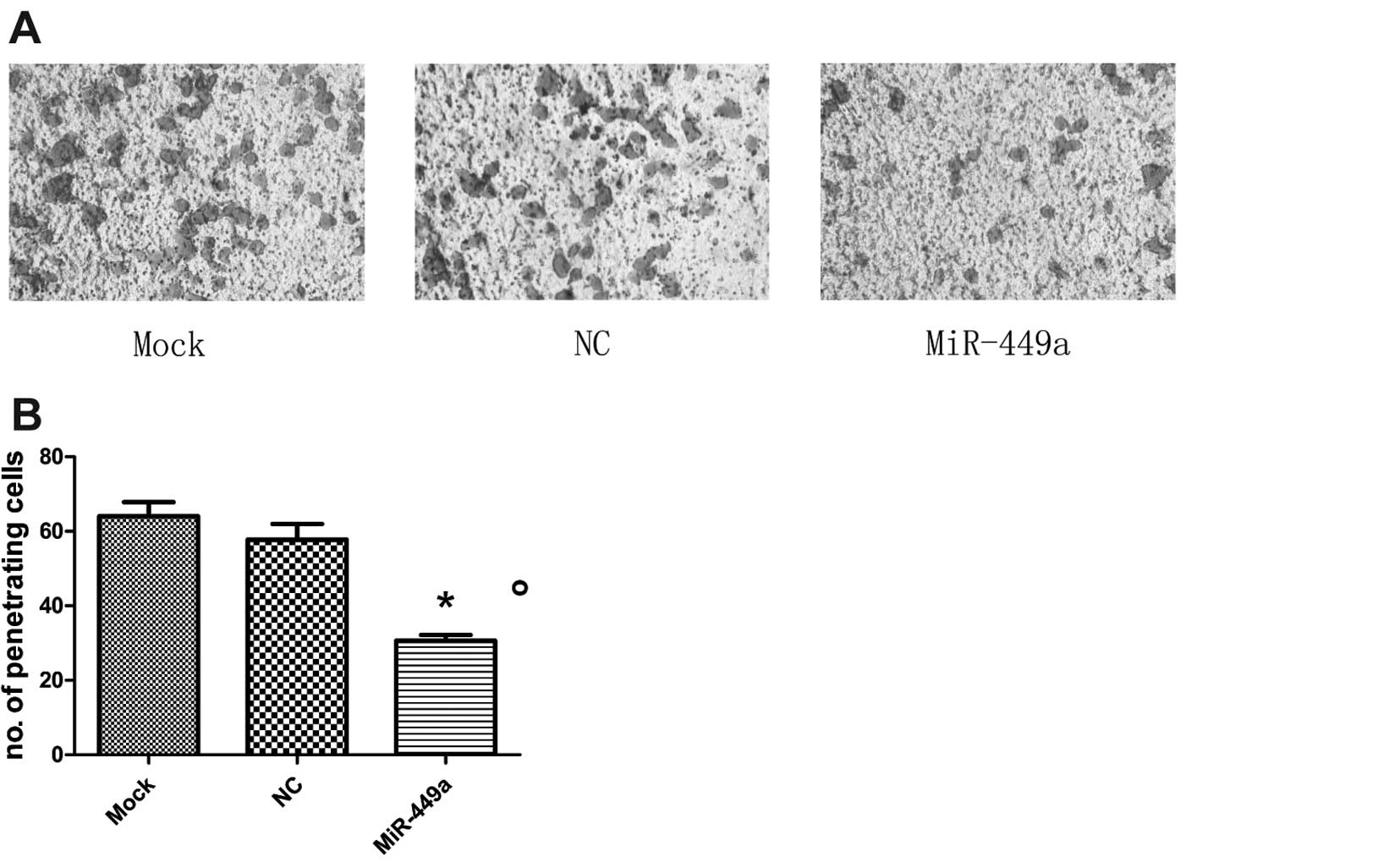
significantly reduced HEC-1B cell invasion. The Transwell invasion
assay revealed that the invasion of miR-449a-transfected cells was
markedly reduced compared to the mock or NC-transfected-cells
(P<0.01; Fig. 4A and B).
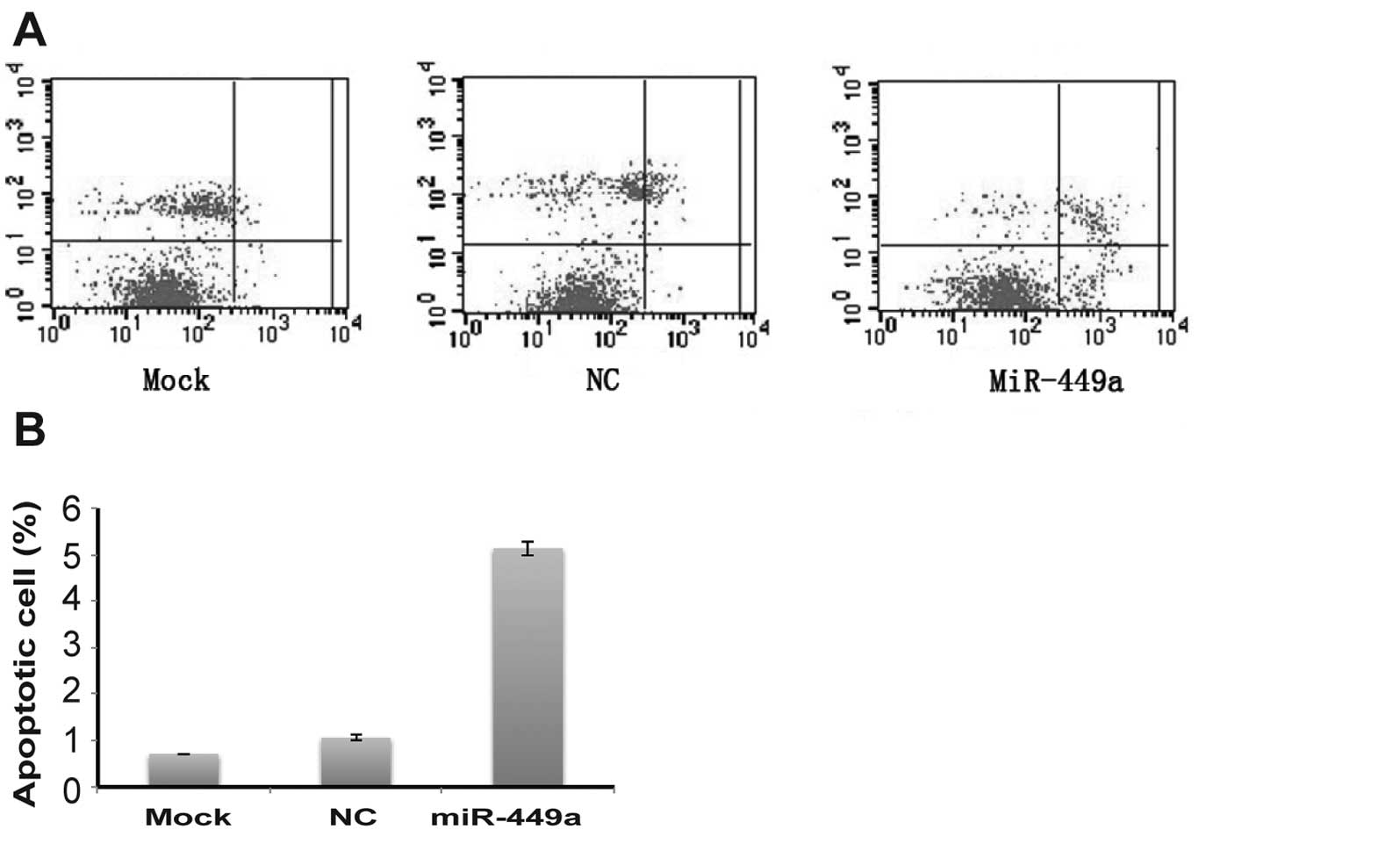
Overexpression of miR-449a induces
apoptosis and suppresses CDC25A expression in HEC-1B cells
We also examined whether overexpression of miR-449a
affected apoptosis in HEC-1B cells. Our results showed that the
population of apoptotic cells in miR-449a-transfected HEC-1B cells
was markedly increased compared with that in mock or NC-transfected
cells (P<0.01; Fig. 5A and B).
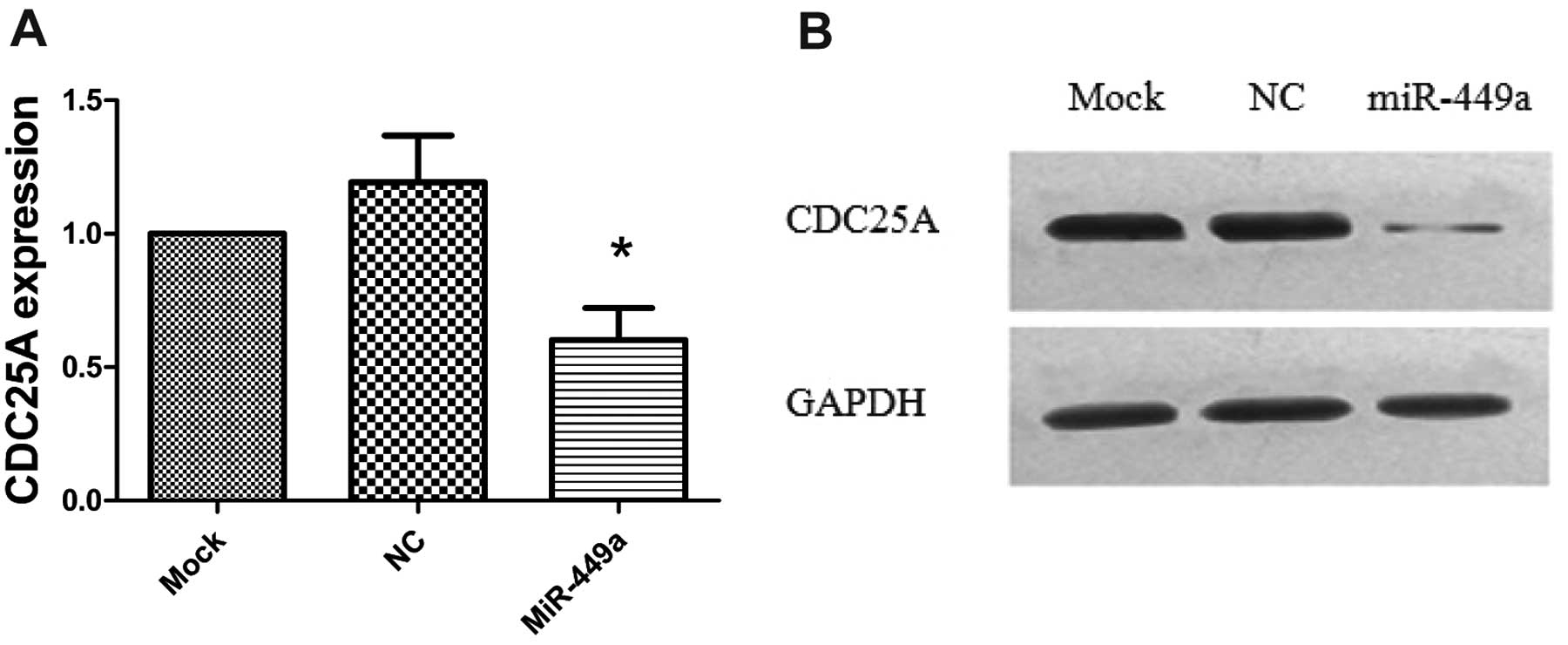
CDC25A is a member of the CDC25 family of phosphatases and is known
to play a critical role in regulating cell cycle. It has been
demonstrated that CDC25A is one of the targets of miR-449a in
breast and bladder cancer (13,15).
Thus, we tested if CDC25A was also affected by miR-449a in HEC-1B
cells. Indeed, the RT-PCR and western blotting assay showed that
both mRNA and protein levels were significantly downregulated in
miR-449a-transfected cells compared to NC or mock-transfected cells
(P<0.05; Fig. 6A and B).
Discussion
MiRNAs are considered new candidate therapeutic
agents for endometrial cancer owing to their involvement in cancer
initiation and progression (19).
Boren et al demonstrated that the miRNA expression profile
in endometrial cancer patients exhibits unique characteristics
compared with that of healthy individuals (20). They found that 12 miRNAs are
associated with endometrial cancer development, including 5
upregulated (let-7i, miR-221, miR-193, miR-152 and miR-30c) and 7
downregulated miRNAs (miR-185, miR-106a, miR-181a, miR-210,
miR-103, miR-107 and let-7c) (20).
In addition, miR-152 and miR-34b have also been demonstrated to
play important roles in the development of endometrial carcinoma
(21,22). In our study, we found that the
expression of miR-449a and miR-449b was markedly reduced in type II
endometrial cancer tissues but not in type I endometrial cancer
tissues compared with normal endometrium, suggesting that miR-449
may be involved in the development of type II endometrial cancer.
This result also indicates that different molecular mechanisms
drive the initiation and progression of type I and type II
endometrial cancer, resulting in distinct clinical characteristics
of the two types of endometrial cancer. We also found that
overexpression of miR-449a in endometrial cancer cells
significantly reduced cell proliferation and invasion and induced
apoptosis, suggesting that miR-449a may act as a tumor suppressor
in endometrial cancer.
Our findings are consistent with studies on miR-449a
in other types of malignancy, such as breast, prostate, bladder,
gastric, liver and lung cancer (13–18).
Yang et al reported that miR-449a/b expression leads to pRB
dephosphorylation and cell cycle arrest at G1 phase in breast
cancer cells by inhibiting oncogenic CDK6 and CDC25A (13). Noonan et al found that the
expression of miR-449 in prostate cancer tissue is downregulated
compared to patient-matched control tissue and overexpression of
miR-449a in prostate cancer PC-3 cells results in cell cycle
arrest, apoptosis and a senescence-like phenotype (14). Downregulation of miR-449a is also
detected in human bladder cancer tissue (15). Exogenous miR-449a can significantly
suppress the growth of bladder cancer cell T24 xenografts (15). In their study on gastric cancer, Bou
Kheir et al found that miR-449 expression is reduced in
mouse gastric cancer tissue and overexpression of miR-449 in
gastric cancer cell line markedly inhibits cell growth and induces
apoptosis. They further identified GMNN, MET, CCNE2, SIRT1 and CDK6
as the targets of miR-449 (17).
Similarly, overexpression of miR-449 also inhibits hepatocellular
carcinoma cell proliferation and induces apoptosis (16). In lung cancer, Ren et al
showed that the level of miR-449a is significantly reduced in human
lung cancer tissues and the low level of miR-449a correlates with
cancer recurrence and survival of lung cancer patients (18). Consistent with other types of
cancer, transient introduction of miR-449a causes cell cycle arrest
and senescence in lung cancer cell line A549 and 95D (18).
The molecular mechanism underlying the
miR-449a-mediated inhibition of cancer growth remains inconclusive.
Several molecules appear to be the key targets of miR-449a. Our
results demonstrated that CDC25A is a target of miR-449a in
endometrial cancer. Similarly, CDC25A has also been shown to be a
target of miR-449a in breast and bladder cancer cells (13,15).
CDC25 is thought to be oncogenic and its expression is upregulated
in various types of tumor tissues, including breast, ovarian,
prostate, lung and endometrial cancer (23). CDC25 regulates cell cycle through
the ATR/ATM-Chk1/Chk2-CDC25A-CDK2 signaling pathway (24). MiR-449a may inhibit cancer growth by
blocking the expression and activity of the oncogene CDC25A. The
loss of miR-449a leads to the upregulation of CDC25A, consequently
resulting in malignant transformation. In addition to CDC25A, CD6,
MET, p53 and p21 are also regulated by miR-449a (13–18).
In conclusion, we demonstrated that the levels of
miR-449a and miR-449b in type II endometrial cancer tissues were
markedly reduced compared to normal endometrium or type I
endometrial cancer tissues and overexpression of miR-449a in
endometrial cancer cells markedly reduced cell proliferation and
invasion, induced apoptosis and suppressed CDC25A expression. Our
findings suggest that miR-449a may act as a tumor suppressor in
endometrial cancer and miR-449a may be a potential therapeutic
agent for endometrial cancer.
Acknowledgements
This study was supported by Zhejiang Provincial
Natural Science Foundation of China (Y2090699).
References
|
1
|
Cho H, Kim YT and Kim JH: Accuracy of
preoperative tests in clinical stage I endometrial cancer: the
importance of lymphadenectomy. Acta Obstet Gynecol Scand.
89:175–181. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Nguyen ML, Lafargue CJ, Pua TL and
Tedjarati SS: Grade 1 endometrioid endometrial carcinoma presenting
with pelvic bone metastasis: a case report and review of the
literature. Case Rep Obstet Gynecol. 2013:8072052013.PubMed/NCBI
|
|
3
|
Banno K, Nogami Y, Kisu I, Yanokura M,
Umene K, Masuda K, Kobayashi Y, Yamagami W, Susumu N and Aoki D:
Candidate biomarkers for genetic and clinicopathological diagnosis
of endometrial cancer. Int J Mol Sci. 14:12123–12137. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jemal A, Siegel R, Xu J and Ward E: Cancer
statistics, 2010. CA Cancer J Clin. 60:277–300. 2010. View Article : Google Scholar
|
|
5
|
Jiang F, Liu T, He Y, Yan Q, Chen X, Wang
H and Wan X: MiR-125b promotes proliferation and migration of type
II endometrial carcinoma cells through targeting TP53INP1 tumor
suppressor in vitro and in vivo. BMC Cancer. 11:4252011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wild PJ, Ikenberg K, Fuchs TJ, Rechsteiner
M, Georgiev S, Fankhauser N, Noske A, Roessle M, Caduff R, Dellas
A, Fink D, Moch H, Krek W and Frew IJ: p53 suppresses type II
endometrial carcinomas in mice and governs endometrial tumour
aggressiveness in humans. EMBO Mol Med. 4:808–824. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gehrig PA and Bae-Jump VL: Promising novel
therapies for the treatment of endometrial cancer. Gynecol Oncol.
116:187–194. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen F and Hu SJ: Effect of microRNA-34a
in cell cycle, differentiation, and apoptosis: a review. J Biochem
Mol Toxicol. 26:79–86. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Iorio MV and Croce CM: MicroRNAs in
cancer: small molecules with a huge impact. J Clin Oncol.
27:5848–5856. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Calin GA and Croce CM: Chromosomal
rearrangements and microRNAs: a new cancer link with clinical
implications. J Clin Invest. 117:2059–2066. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
He L, He X, Lim LP, de Stanchina E, Xuan
Z, Liang Y, Xue W, Zender L, Magnus J, Ridzon D, Jackson AL,
Linsley PS, Chen C, Lowe SW, Cleary MA and Hannon GJ: A microRNA
component of the p53 tumour suppressor network. Nature.
447:1130–1134. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Olive V, Jiang I and He L: mir-17-92, a
cluster of miRNAs in the midst of the cancer network. Int J Biochem
Cell Biol. 42:1348–1354. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yang X, Feng M, Jiang X, Wu Z, Li Z, Aau M
and Yu Q: miR-449a and miR-449b are direct transcriptional targets
of E2F1 and negatively regulate pRb-E2F1 activity through a
feedback loop by targeting CDK6 and CDC25A. Genes Dev.
23:2388–2393. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Noonan EJ, Place RF, Pookot D, Basak S,
Whitson JM, Hirata H, Giardina C and Dahiya R: miR-449a targets
HDAC-1 and induces growth arrest in prostate cancer. Oncogene.
28:1714–1724. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen H, Lin YW, Mao YQ, Wu J, Liu YF,
Zheng XY and Xie LP: MicroRNA-449a acts as a tumor suppressor in
human bladder cancer through the regulation of pocket proteins.
Cancer Lett. 320:40–47. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Buurman R, Gürlevik E, Schäffer V, Eilers
M, Sandbothe M, Kreipe H, Wilkens L, Schlegelberger B, Kühnel F and
Skawran B: Histone deacetylases activate hepatocyte growth factor
signaling by repressing microRNA-449 in hepatocellular carcinoma
cells. Gastroenterology. 143:811–820. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bou Kheir T, Futoma-Kazmierczak E,
Jacobsen A, Krogh A, Bardram L, Hother C, Grønbæk K, Federspiel B,
Lund AH and Friis-Hansen L: miR-449 inhibits cell proliferation and
is down-regulated in gastric cancer. Mol Cancer.
10:292011.PubMed/NCBI
|
|
18
|
Ren XS, Yin MH, Zhang X, Wang Z, Feng SP,
Wang GX, Luo YJ, Liang PZ, Yang XQ, He JX and Zhang BL:
Tumor-suppressive microRNA-449a induces growth arrest and
senescence by targeting E2F3 in human lung cancer cells. Cancer
Lett. 344:195–203. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Umene K, Banno K, Kisu I, Yanokura M,
Nogami Y, Tsuji K, Masuda K, Ueki A, Kobayashi Y, Yamagami W,
Tominaga E, Susumu N and Aoki D: New candidate therapeutic agents
for endometrial cancer: Potential for clinical practice (Review).
Oncol Rep. 29:855–860. 2013.PubMed/NCBI
|
|
20
|
Boren T, Xiong Y, Hakam A, Wenham R, Apte
S, Wei Z, Kamath S, Chen DT, Dressman H and Lancaster JM: MicroRNAs
and their target messenger RNAs associated with endometrial
carcinogenesis. Gynecol Oncol. 110:206–215. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tsuruta T, Kozaki K, Uesugi A, Furuta M,
Hirasawa A, Imoto I, Susumu N, Aoki D and Inazawa J: miR-152 is a
tumor suppressor microRNA that is silenced by DNA hypermethylation
in endometrial cancer. Cancer Res. 71:6450–6462. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hiroki E, Suzuki F, Akahira J, Nagase S,
Ito K, Sugawara J, Miki Y, Suzuki T, Sasano H and Yaegashi N:
MicroRNA-34b functions as a potential tumor suppressor in
endometrial serous adenocarcinoma. Int J Cancer. 131:E395–E404.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Meng F, Henson R, Lang M, Wehbe H,
Maheshwari S, Mendell JT, Jiang J, Schmittgen TD and Patel T:
Involvement of human micro-RNA in growth and response to
chemotherapy in human cholangiocarcinoma cell lines.
Gastroenterology. 130:2113–2129. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu T, Yu X, Li G, Yuan R, Wang Q, Tang P,
Wu L, Liu X, Peng X and Shao J: Rock2 regulates Cdc25A through
ubiquitin proteasome system in hepatocellular carcinoma cells. Exp
Cell Res. 318:1994–2003. 2012. View Article : Google Scholar : PubMed/NCBI
|