Introduction
The tumor necrosis factor receptor-associated factor
(TRAF) family, consisting of seven members (TRAF1–7), is a group of
signaling adaptors which participate in various signaling pathways
through binding of the tumor necrosis factor receptor superfamily
(1–3). Two decades since the TRAF family was
first cloned, marked progress in the understanding of the functions
of the TRAF family has been acquired. TRAF family members have now
been suggested to be signal transducers, modulating immune
receptors, cytokine receptors, C-type lectin receptors and adaptive
immune receptors (3). Therefore,
the TRAF family has been demonstrated to contribute to the
pathology of a variety of human diseases including cancers,
autoimmune diseases and neurodegenerative diseases (4–6).
Considering the importance of the TRAF family, targeting these
molecules may lead to the development of therapeutic intervention
of TRAF-mediated human diseases.
Among the seven members, TRAF4 is a unique member of
the TRAF family, and is one of the conserved proteins during
evolution (7). TRAF4 is an
important gene in the regulation of an organism’s development and
has been reported to be involved in embryogenesis (8,9). In
addition, research has demonstrated that TRAF4 is required in
central nervous system myelin homeostasis (10). TRAF4 deficiency is embryonic lethal
and causes severe developmental abnormalities in the respiratory
system and axial skeleton in mouse models (11,12).
Moreover, dendritic cells from TRAF4-deficient mice showed reduced
cell migratory ability in vivo and in vitro (13). Interestingly, TRAF4 is the first
member of the TRAF family that has been found to be overexpressed
in cancers, and is now considered an oncogene (14,15).
TRAF4 was originally identified in a breast cancer cDNA library by
gene screening to play an important role in the initiation and
progression of primary breast cancers (16,17).
Studies have reported that TRAF4 has multiple subcellular
localizations including cytoplasmic, nuclear, and membrane and have
been described in the literature (16,18,19).
Particularly, the subcellular localization of TRAF4 in cancers is
mainly in the cytoplasm or nucleus (18). Moreover, breast cancer patients with
TRAF4 nuclear localization have poor survival due to its ability to
destabilize the p53 protein in the nucleus (20).
As a signal transducer, studies have revealed that
TRAF4 is engaged in several signaling pathways. Several studies
have shown that TRAF4 is recruited to different tumor necrosis
factor receptors and Toll-like receptors (21–23).
It has been reported that TRAF4 regulates the
glucocorticoid-induced NF-κB signaling pathway, implying its role
in modulating the suppressive functions of Treg cells (24). TRAF4 promotes MEKK4 oligomerization
to mediate Jun N-terminal kinase activation in embryos (25). In addition, TRAF4 is a negative
regulator of IL-17 signaling which is involved in regulating
Th17-mediated disease (26). In
spite of these findings, the underlying mechanism of TRAF4 in
tumorigenesis remains largely unknown.
In the present study, we aimed to explore the
biological functions of TRAF4 in breast cancer cells. By using
different breast cancer cells, we found that TRAF4 was particularly
overexpressed. Knockdown of TRAF4 significantly inhibited cell
proliferation and decreased cell migration and the invasion of
breast cancer cells. More importantly, we identified that TRAF4 was
capable of promoting Akt activation. Furthermore, a direct
interaction was observed between TRAF4 and Akt which was essential
for Akt membrane recruitment. Additionally, overexpression of
constitutively active Akt reversed the cell growth arrest in TRAF4
knockdown cells. Taken together, our data support the suggestion
that TRAF4 plays an important role in cancer cells through the
activation of Akt, and may be a potential candidate molecular
target for breast cancer prevention and therapy.
Materials and methods
Cell lines and cell culture
The human breast cancer cell lines MCF-7, T47D and
MDA-MB-468 were obtained from the American Type Culture Collection
(ATCC; Manassas, VA, USA). All cells were maintained according to
standard protocols. Briefly, cells were maintained in Dulbecco’s
modified Eagle’s medium (DMEM) supplemented with 10% fetal bovine
serum (FBS) to which 100 U/ml of penicillin G, 0.1 mg/ml
streptomycin sulfate and 0.25 μg/ml amphotericin B were added.
Human mammary epithelial cells (HMECs) and 293T cells were cultured
in DMEM containing 10% FBS and 1% antibiotics. All cells were
cultured at 37°C with 5% CO2 in an incubator (Life
Technologies, Baltimore, MD, USA).
Plasmids and small interference RNA
(siRNA) transfection
Cells were seeded in a 6-well culture plate
(2×105 cells/well) under standard conditions. When the
cells reached 80% confluency, cell transfection was performed as
per the standard protocols of the manufacturer. Briefly, plasmid
DNA [pCDNA3.0-TRAF4, Myc-Akt1 or constitutively active (CA)-Akt1;
Addgene] or siRNA (TRAF siRNA, sc-36713; Santa Cruz Biotechnology,
Santa Cruz, CA, USA) was diluted in 500 μl of DMEM with 5 μl
Lipofectamine (Invitrogen, Carlsbad, CA, USA), before being mixed
and incubated at room temperature for 15 min. The mixtures were
then added to the cells to a final volume of 3 ml medium and
incubated for 36 to 48 h, before proteins were extracted for
further analysis.
Bromodeoxyuridine (BrdU) assay
The BrdU cell proliferation assay kit (Millipore,
Billerica, MA, USA) was used for cell proliferation analysis as per
the manufacturer’s instructions. Briefly, cells in 96-well plates
were transfected with plasmids or siRNA for 24 h, then BrdU
solution (10 μl/well) was added and incubated for 2 h. The old
medium was discarded, the Fixing/Denaturing solution (100 μl/well)
was added and incubated at room temperature for 15 min. After that,
the supernatants were removed, and the prepared detection antibody
solution (100 μl/well) was added for incubation at room temperature
for 1 h. Subsequently, plates were washed three times with wash
buffer, and the prepared horseradish peroxidase (HRP)-conjugated
secondary antibody solution (100 μl/well) was added and incubated
for 30 min at room temperature. Then, plates were washed, and
tetramethylbenzidine (TMB) substrate (100 μl) was added for
incubation at room temperature for 30 min. The amount of BrdU
incorporation into the cells was measured at 450 nm by a microplate
reader (Bio-Rad, Hercules, CA, USA). Experiments were performed in
quintuplicate and repeated three times.
Cell invasion and migration assays
Cells (2×105) were suspended in a volume
of 50 μl serum-free medium which wrer then plated in the upper
chamber of chemotaxis chambers (Neuro Probe, Gaithersburg, MD,
USA). Complete medium (75 μl) was added to the lower chamber and
incubated at 37°C for 48 h; then, the inserts were removed and
submerged in PBS to remove the unattached cells, before being fixed
and stained by Diff Quick (IHC World, Bethesda, MD, USA). After
that, membranes were cut and mounted on slides. Images were
captured of the migrated cells (x20) on the underside of the
membrane, and 10 visual fields in each membrane were randomly
selected for cell number counting. For the invasion assays, cells
(5×105) in 60 μl serum-free medium were plated in the
top compartment of Matrigel-coated invasion chambers (8-μm pore
membrane). Fibroblast conditioned medium (0.75 ml) was added to the
bottom chambers, and cultures were incubated at 37°C for 48 h. The
membranes were fixed and stained using Diff Quick. Images were
captured of the invaded cells (x20), and 10 visual fields in each
membrane were randomly selected for cell number counting. Each
assay was repeated 3 times independently.
Membrane fractionation
Cells were starved for 24 h in DMEM containing 0.1%
FBS in 6-well plates. Epidermal growth factor (EGF) (50 ng/ml) was
added and incubated for 30 min. After that, the membrane and
cytosolic fractions were extracted using the Membrane Protein
Extraction Kit (Sangon, Shanghai, China) according to the
manufacturer’s instructions. Briefly, cells were washed with wash
buffer at least three times. Then, 1 ml of extract buffer
containing DTT (1 μg/ml) was added and homogenized under an
ice-cold condition followed by centrifugation (14,000 rpm) at 4°C
for 10 min. The supernatants were collected, bathed at 37°C for 10
min, then centrifuged (13,000 rpm) at room temperature for 5 min.
The samples were divided into 2 layers. The bottom layer,
containing cytoplasmic proteins, was collected and stored for
further analysis. The bottom layer, containing membrane proteins,
was dissolved in 500 μl of ice-cold sterile water for 5 min at 4°C,
followed by a water bath at 37°C for 5 min. After centrifugation
(13,000 rpm) at room temperature for 5 min, the bottom layer was
collected and dissolved in ice-cold sterile water again following
the above steps. Finally, the membrane extracts were collected, and
the protein concentration was measured using the BCA kit (Pierce,
Rockford, IL, USA). For SDS-PAGE analysis, a total of 100 μl
membrane protein was mixed with 0.9 ml acetone, incubated in an
ice-cold condition for 20 min followed by centrifugation (10,000
rpm) for 20 min. The supernatants were removed, and 100 μl of
loading buffer and 2 μl of β-mercaptoethanol were added to dissolve
the sediments for sodium dodecyl sulfate polyacrylamide gel
electrophoresis (SDS-PAGE) analysis.
Co-immunoprecipitation
The cells were lysed and centrifuged, and extracts
were then collected. Protein A-Sepharose beads (Amersham
Biosciences AB, Uppsala, Sweden) mixed with a mouse monoclonal
anti-Flag (Sigma, St. Louis, MO, USA), or mouse IgG as a control,
were incubated at 4°C in 500 μl of lysis buffer for 1 h. Cell
extracts were added to the prepared antibody-bead mixture and
incubated at 4°C for 2 h. Then, the bead complexes were collected
and washed for a total of three times. Then, the protein complexes
were eluted from the beads using glycine buffer (pH 2.5), and
separated by SDS-PAGE and examined by western blotting.
Western blot analysis
Proteins were extracted from cells, and
concentrations were measured using the BCA kit. A total of 20 μg
protein was separated by 12% SDS-PAGE electrophoresis followed by
electro-blotting onto a nitrocellulose membrane (Amersham, Little
Chalfont, UK). Then, the membrane was incubated in Tris-buffered
saline (TBS) containing 2% non-fat dry milk to block non-specific
binding at room temperature for 1 h. The membrane was then washed
with TBS and incubated with primary antibodies [antibodies against
TRAF4, GAPDH, myc-tag, α-tubulin and E-cadherin (all from Santa
Cruz Biotechnology), phosphorylated Akt (Ser473), total Akt and
Akt1 (all from Cell Signaling Technology, Danvers, MA, USA) diluted
(1:1,000)] in the blocking buffer overnight at 4°C. Subsequently,
the membrane was washed with TBS. After that, the membrane was
incubated in horseradish peroxidase (HRP)-conjugated secondary
antibodies (Boster Corporation, Wuhan, China) diluted in blocking
buffer for 1 h. Finally, the membrane was washed followed by the
addition of 4-chloro-1-naphthol (4-CN; 1 ml) with TBS (9 ml)
containing 6 μl of H2O2, which was used for
protein visualization.
Statistical analysis
Data are expressed as means ± standard deviation
(SD). The statistical significance of differences between two
groups was determined by the Student’s t-test, and among multiple
groups was determined by one-way ANOVA. P<0.05 was considered to
indicate a statistically significant difference. All statistical
analyses were performed using SPSS 11.5 Software (SPSS Inc.,
Chicago, IL, USA).
Results
TRAF4 is specifically overexpressed in
human breast cancer cells
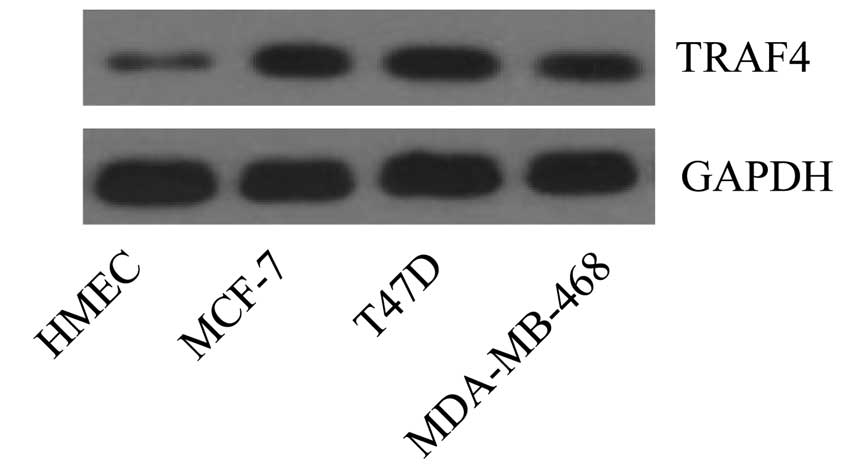
To verify the expression profiles of TRAF4 in human
breast cancer, we delineated the protein expression level of TRAF4
in different breast cancer cell lines in vitro. The results
showed that, compared with the normal human mammary epithelial
cells (HMECs), TRAF4 was particularly overexpressed in the human
breast cancer cell lines MCF-7, T47D and MDA-MB-468 (Fig. 1). The data indicate that TRAF4 has a
critical role in human breast cancer development.
TRAF4 promotes cancer cell
proliferation
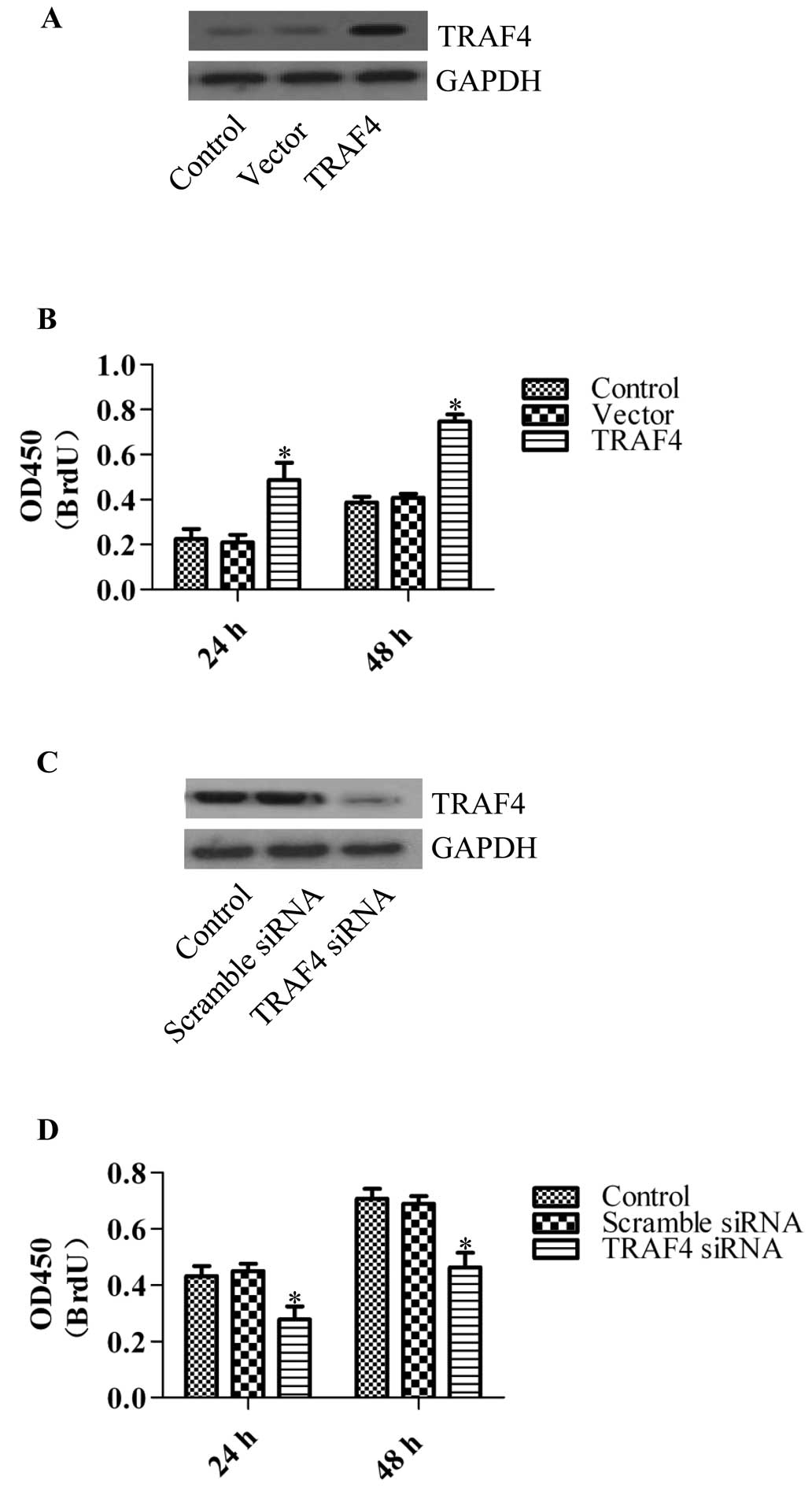
To investigate the role of TRAF4 in breast cancer
cells, we applied the TRAF4 overexpression vector or the TRAF
target siRNA to generate TRAF4-overexpressing or stable knockdown
cell lines. We first generated TRAF4-overexpressing cells by
transfecting HMECs with the TRAF4 overexpression vectors (Fig. 2A). The BrdU assay was used to
determine the effects on cell proliferation. The results showed
that overexpression of TRAF4 significantly promoted the
proliferation of HMECs following transfection (Fig. 2B), whereas TRAF4 knockdown
significantly inhibited cell proliferation in the breast cancer
cell line MCF-7 (Fig. 2C and D).
Similar results were obtained in the siRNA-transfected T47D and
MDA-MB-468 cell lines (data not shown). These results imply that
TRAF4 plays an important role in cancer cell proliferation.
Knockdown of TRAF4 impairs tumor cell
migration and invasion
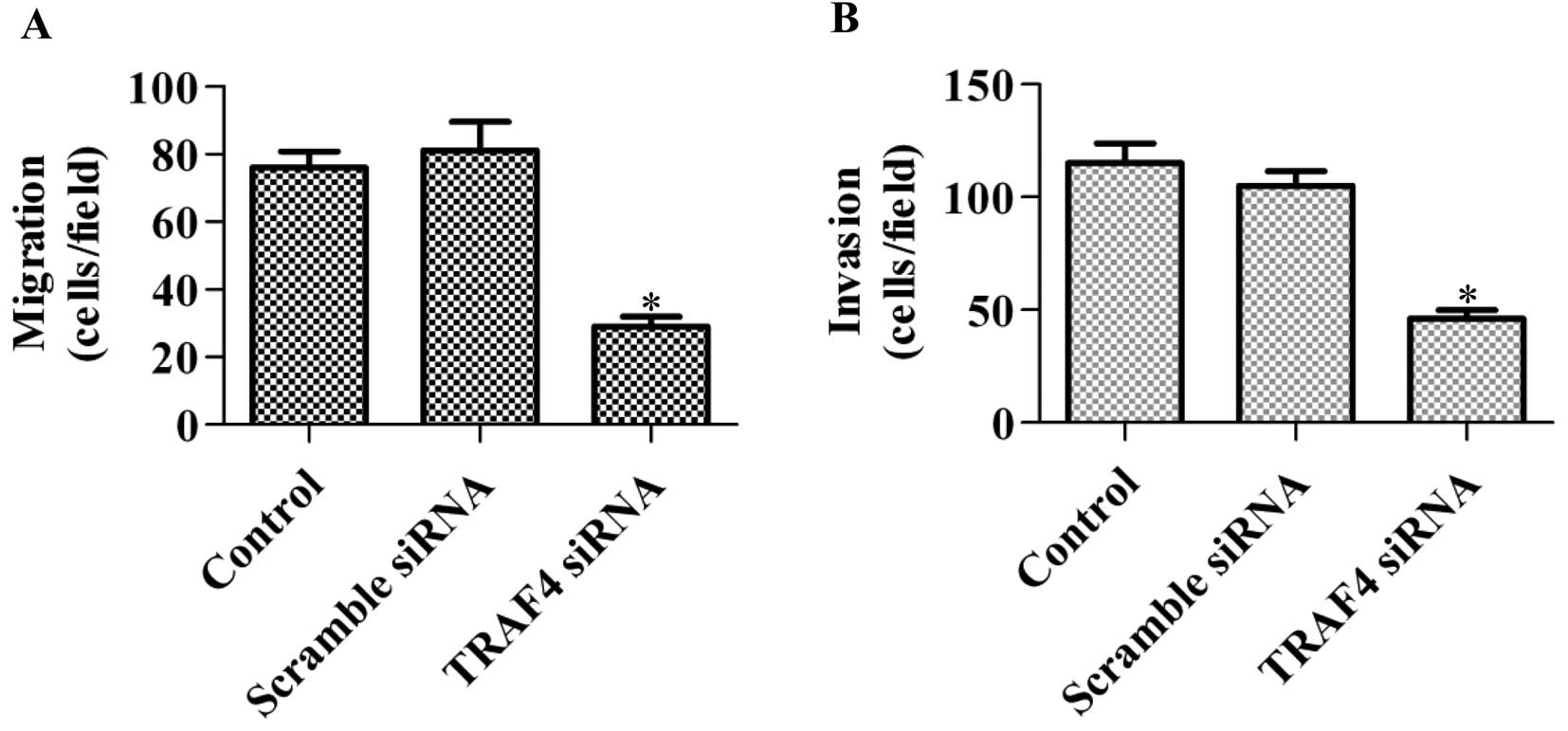
To further explore the function of TRAF4 in cancer
cells, the role of TRAF on cell migration and invasion was
determined. Knockdown of TRAF4 in MCF-7 cells significantly
decreased the cell migratory and invasive abilities (Fig. 3A and B). Furthermore, the same
results were obtained when using the T47D and MDA-MB-468 cell lines
(data not shown). Overall, the data suggest that TRAF4 has various
tumorigenic roles.
TRAF4 increases Akt membrane recruitment
and activation
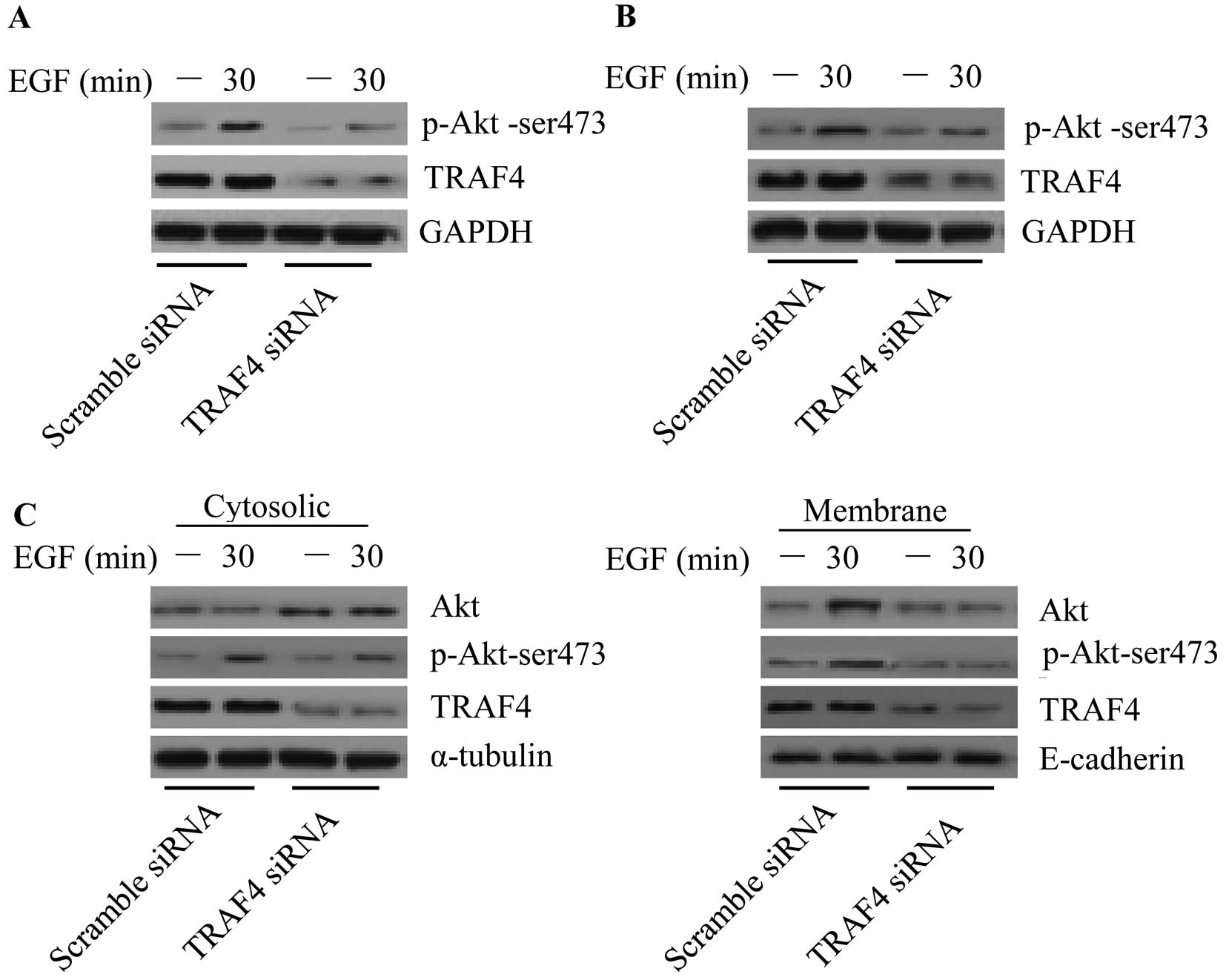
To further investigate the underlying mechanism of
TRAF4 in tumorigenesis, we aimed to identify the signaling pathway
in which TRAF4 is involved. A previous study demonstrated that TRAF
family members such as TRAF6 (27)
are involved in the Akt pathway. As the TRAF family members share
similar functional domains, we investigated whether TRAF4 is
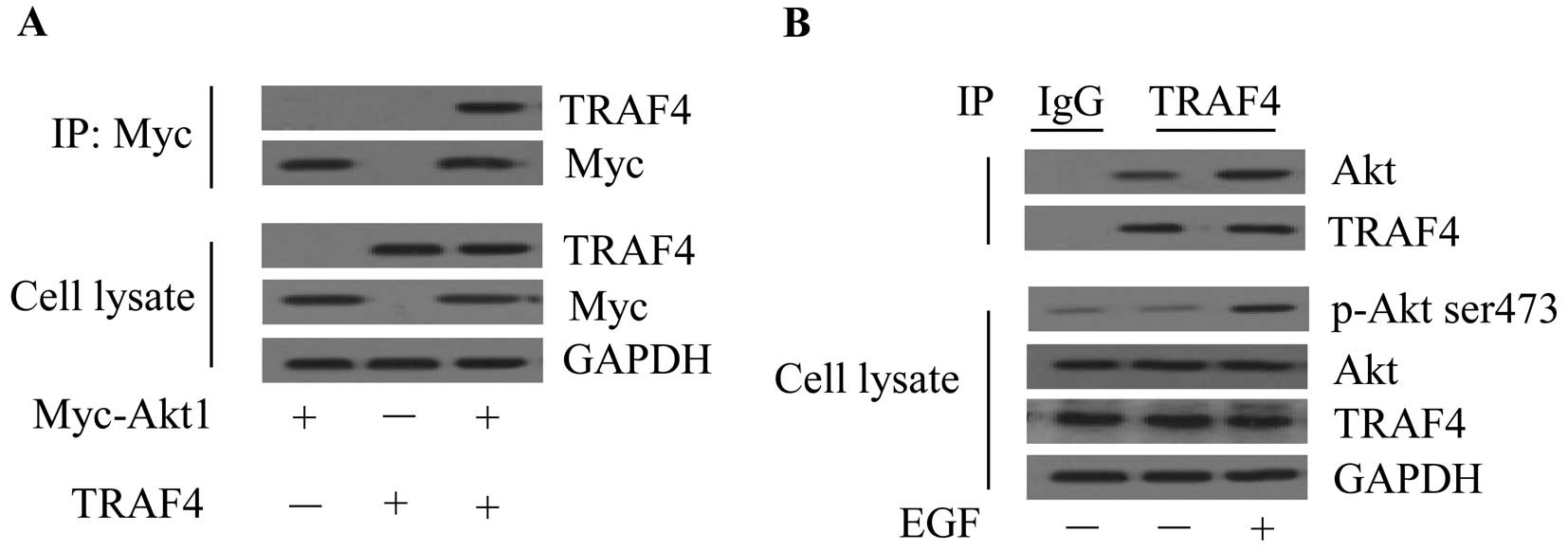
capable of activating Akt. Upon EGF treatment, Akt phosphorylation
(Ser473) was upregulated in the control cells. However, EGF-induced
phosphorylation of Akt was markedly inhibited in the
TRAF4-knockdown MCF-7 (Fig. 4A) and
T47D breast cancer cells (Fig. 4B).
Akt membrane recruitment is critical for Akt activation. We next
examined whether TRAF4 plays an important role in Akt membrane
translocation. The results showed that knockdown of TRAF4 caused
Akt cytosolic accumulation, and the phosphorylation of Akt was
markedly disturbed both in the cytoplasm and membrane (Fig. 4C). The data imply that TRAF4 plays
an important role in Akt membrane recruitment in breast cancer
cells.
TRAF4 displays an interaction with
Akt
To explore the underlying mechanism of TRAF4 in
regulating Akt, we examined whether a direct interaction is
observed between the two proteins. Myc-Akt1 and TRAF4 were
co-transfected in 293T cells and immunoprecipitated with the
anti-Myc antibody. The results showed that the TRAF4 protein was
only detected in the immunoprecipitation complex from the Myc-Akt1
and TRAF4 co-transfected cells (Fig.
5A). Next, we aimed to determine the interaction of endogenous
Akt1 and TRAF4 in the MCF-7 cells. The results showed that
endogenous Akt interacted with TRAF4 in breast cancer cells and the
interaction was enhanced upon EGF treatment (Fig. 5B). These data indicate that TRAF4
undergoes an interaction with Akt in breast cancer cells.
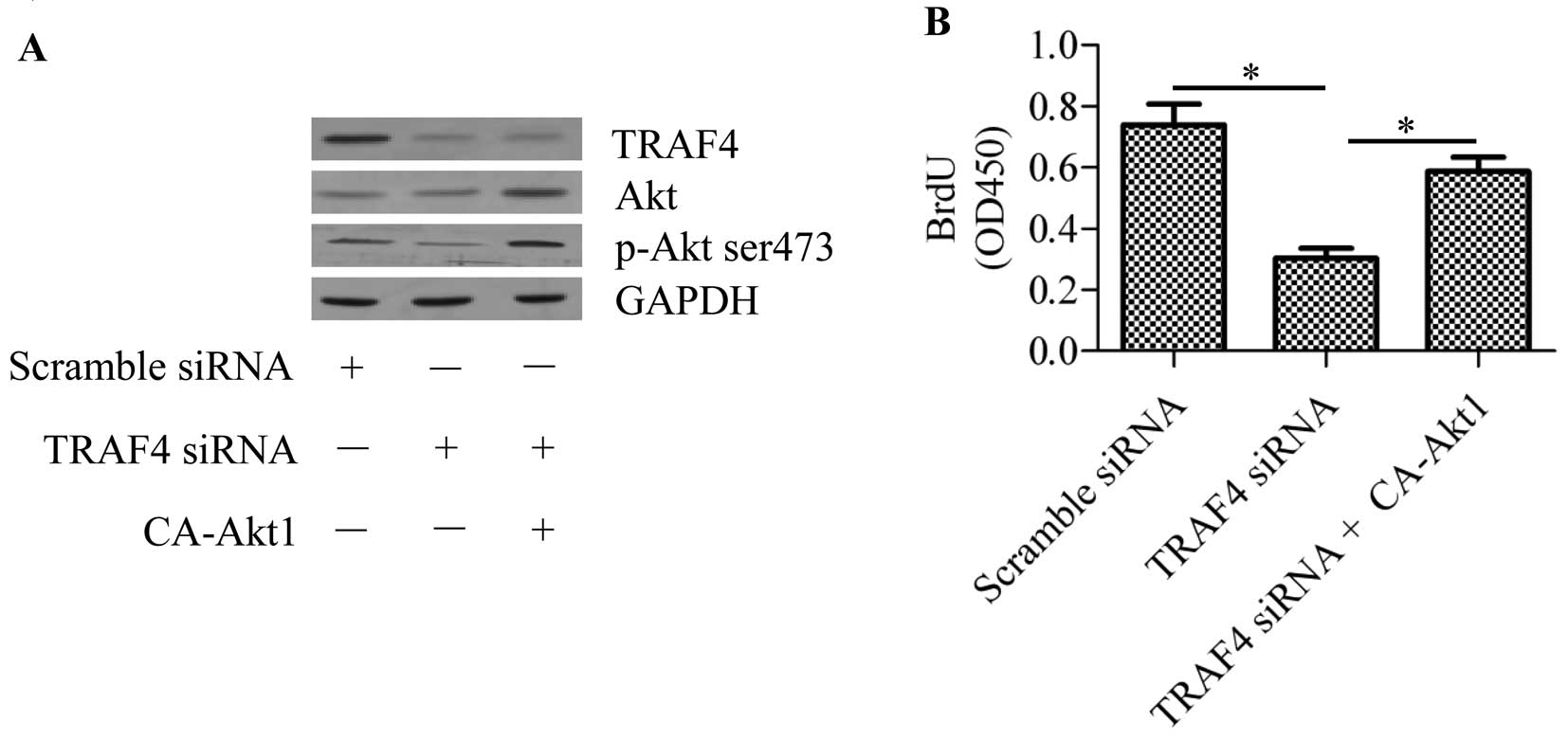
TRAF4 promotes cell proliferation through
Akt
To further confirm the function of TRAF4 in the Akt
signaling pathway, we transfected constitutively activated Akt1
(CA-Akt1) into TRAF4-knockdown MCF-7 breast cancer cells. The
results showed that transfection of CA-Akt1 significantly
upregulated the phosphorylation of Akt in the TRAF4-knockdown cells
(Fig. 6A). Cell growth arrest
induced by TRAF4 silencing was reversed by CA-Akt1 transfection
(Fig. 6B). These data suggest that
TRAF4 regulation on cell proliferation is dependent on Akt
activity.
Discussion
Breast cancer is currently the second leading cause
of cancer-related death, and is the most commonly diagnosed cancer
in women worldwide (28). Although
a better understanding of the biological mechanisms that underlie
breast cancer development has been achieved with the development of
advanced molecular biology techniques in recent years, a major
current challenge in the treatment of breast cancer is to discover
novel and effective targets that can complement current therapies.
In the present study, we demonstrated that TRAF4 plays an important
role in the activation of the Akt signaling pathway in breast
cancer. Considering the important function of TRAF4 in the
development of breast cancer, TRAF4 may be regarded as a potential
target for breast cancer therapy.
Our data revealed that TRAF4 was overexpressed in
breast cancer cell lines. Moreover, RNAi-mediated knockdown of
TRAF4 significantly inhibited cell proliferation, invasive and
migratory abilities of breast cancer cells. These results suggest
that TRAF4 plays an important role in human breast cancers.
Although TRAF4 overexpression in a wide range of human cancers has
been verified (14), TRAF4
overexpression in squamous cell carcinoma of the head and neck has
an anti-tumor effect through the induction of cell apoptosis and
the suppression of colony formation (29,30).
It has been demonstrated that TRAF4 is a p53-regulated
pro-apoptotic gene in p53 temperature-sensitive cells (31). In contrast, a recent study
demonstrated that TRAF4 was a downstream gene of steroid receptor
coactivator 3 (SRC-3), which is an oncogenic nuclear receptor
coactivator that increases p53 destabilization leading to
resistance to cytotoxic stress and poor prognosis in breast cancer
patients (20). Therefore, it seems
that TRAF4 overexpression has different outcomes in different
cancers. The apparent discrepancy implies that TRAF4 may have
different biological functions upon stimulation and cell type.
More recently, TRAF4 has been suggested to
participate in breast cancer migration through destabilizing tight
junctions in mammary epithelial cells (32). Wang et al showed that Smad
ubiquitin regulatory factors (Smurfs) promoted the ubiquitination
of TRAF4 which is essential for the proper localization of TRAF4 to
tight junctions in confluent epithelial cells (33). In prostate cancer, TRAF4 expression
was found to be regulated by tumor-suppressor microRNA-29a, and an
inverse correlation was identified in tumor tissues from radical
prostatectomy (34). In addition,
Zhang et al demonstrated that TRAF4 promotes the
transforming growth factor (TGF)-β signaling pathway, which
contributes to the pathogenesis of breast cancer (35). All of these data indicate that TRAF4
plays a critical role in tumorigenesis. In the present study, we
found that TRAF4 regulated the activation of Akt in breast cancer
cells. Our data are consistent with a more recent study which
indicated that TRAF4, possessing ubiquitin-protein ligase activity,
activates Akt through the ubiquitination of Akt in lung cancer
(36).
It is well-known that the Akt pathway is closely
related to tumorigenesis by regulating cell growth and survival,
cell cycle and metabolism (37–39).
Akt is deregulated in many types of cancer, including breast
cancer, and contributes to cancer cell growth and survival, and
resistance to chemotherapy or radiotherapy (40,41).
Akt activation is dependent on the phosphorylation of Thr308 and
Ser473 upon various stimuli such as insulin and epidermal growth
factor (EGF) (42). Studies suggest
that Akt recruitment to the membrane upon growth factor stimuli is
essential for activation (27,43).
In the present study, we found that TRAF4 regulated Akt membrane
recruitment in breast cancer cells upon EGF stimulation, whereas
knockdown of TRAF4 caused a decline in Akt membrane recruitment and
activation. We further revealed a direct interaction between TRAF4
and Akt, which was the basis for TRAF4 in promoting Akt membrane
recruitment. In accordance with this, Li et al demonstrated
that TRAF4 functions as an E3 ligase which regulates Akt
ubiquitination and activation (36). Previous studies suggest that
Lys63-mediated ubiquitination of Akt is necessary for Akt
activation (44). However, we did
not further investigate the ubiquitination of Akt by TRAF4 activity
in breast cancer cells. Whether TRAF4 promotes Akt activation by
ubiquitination in breast cancer cells therefore remains uncertain.
Additionally, we found that overexpression of constitutively active
Akt1 reversed the cell growth arrest caused by TRAF4 knockdown,
implying that TRAF4 regulates cell growth through the Akt signaling
pathway.
Taken together, we found that TRAF4 promoted Akt
activation in human breast cancer cells. Given the important roles
of Akt in regulating tumorigenesis, targeting TRAF4 to inhibit
activated Akt in breast cancer is a potential therapeutic strategy
for the prevention and treatment of human breast cancers.
Acknowledgements
This research was supported in part by the Natural
Science Foundation (no. 2011A310005), the Science and Technique
Foundation (no. 112102310206) and a University Key Teacher grant
from the Ministry of Education (no. 2012GGJS-136) of Henan.
Abbreviations:
|
TRAF4
|
tumor necrosis factor receptor
associated factor 4
|
|
RNAi
|
RNA interference
|
|
BrdU
|
bromodeoxyuridine
|
|
EGF
|
epidermal growth factor
|
References
|
1
|
Inoue J, Ishida T, Tsukamoto N, et al:
Tumor necrosis factor receptor-associated factor (TRAF) family:
adapter proteins that mediate cytokine signaling. Exp Cell Res.
254:14–24. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Arron JR, Walsh MC and Choi Y:
TRAF-mediated TNFR-family signaling. Curr Protoc Immunol. Nov
1–2002.Chapter 11(Unit 11): 9D View Article : Google Scholar
|
|
3
|
Xie P: TRAF molecules in cell signaling
and in human diseases. J Mol Signal. 8:72013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Keats JJ, Fonseca R, Chesi M, et al:
Promiscuous mutations activate the noncanonical NF-kappaB pathway
in multiple myeloma. Cancer Cell. 12:131–144. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Namjou B, Choi CB, Harley IT, et al:
Evaluation of TRAF6 in a large multiancestral lupus cohort.
Arthritis Rheum. 64:1960–1969. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zucchelli S, Codrich M, Marcuzzi F, et al:
TRAF6 promotes atypical ubiquitination of mutant DJ-1 and
alpha-synuclein and is localized to Lewy bodies in sporadic
Parkinson’s disease brains. Hum Mol Genet. 19:3759–3770.
2010.PubMed/NCBI
|
|
7
|
Kedinger V and Rio MC: TRAF4, the unique
family member. Adv Exp Med Biol. 597:60–71. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Masson R, Regnier CH, Chenard MP, et al:
Tumor necrosis factor receptor associated factor 4 (TRAF4)
expression pattern during mouse development. Mech Dev. 71:187–191.
1998. View Article : Google Scholar
|
|
9
|
Kedinger V, Alpy F, Tomasetto C, et al:
Spatial and temporal distribution of the traf4 genes during
zebrafish development. Gene Expr Patterns. 5:545–552. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Blaise S, Kneib M, Rousseau A, et al: In
vivo evidence that TRAF4 is required for central nervous system
myelin homeostasis. PLoS One. 7:e309172012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Regnier CH, Masson R, Kedinger V, et al:
Impaired neural tube closure, axial skeleton malformations, and
tracheal ring disruption in TRAF4-deficient mice. Proc Natl Acad
Sci USA. 99:5585–5590. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shiels H, Li X, Schumacker PT, et al:
TRAF4 deficiency leads to tracheal malformation with resulting
alterations in air flow to the lungs. Am J Pathol. 157:679–688.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cherfils-Vicini J, Vingert B, Varin A, et
al: Characterization of immune functions in TRAF4-deficient mice.
Immunology. 124:562–574. 2008. View Article : Google Scholar
|
|
14
|
Camilleri-Broët S, Cremer I, Marmey B, et
al: TRAF4 overexpression is a common characteristic of human
carcinomas. Oncogene. 26:142–147. 2007.PubMed/NCBI
|
|
15
|
Rhodes DR, Yu J, Shanker K, et al:
Large-scale meta-analysis of cancer microarray data identifies
common transcriptional profiles of neoplastic transformation and
progression. Proc Natl Acad Sci USA. 101:9309–9314. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Regnier CH, Tomasetto C, Moog-Lutz C, et
al: Presence of a new conserved domain in CART1, a novel member of
the tumor necrosis factor receptor-associated protein family, which
is expressed in breast carcinoma. J Biol Chem. 270:25715–25721.
1995. View Article : Google Scholar
|
|
17
|
Bieche I, Tomasetto C, Regnier CH, et al:
Two distinct amplified regions at 17q11-q21 involved in human
primary breast cancer. Cancer Res. 56:3886–3890. 1996.PubMed/NCBI
|
|
18
|
Glauner H, Siegmund D, Motejadded H, et
al: Intracellular localization and transcriptional regulation of
tumor necrosis factor (TNF) receptor-associated factor 4 (TRAF4).
Eur J Biochem. 269:4819–4829. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kedinger V, Alpy F, Baguet A, et al: Tumor
necrosis factor receptor-associated factor 4 is a dynamic tight
junction-related shuttle protein involved in epithelium
homeostasis. PLoS One. 3:e35182008. View Article : Google Scholar
|
|
20
|
Yi P, Xia W, Wu RC, et al: SRC-3
coactivator regulates cell resistance to cytotoxic stress via
TRAF4-mediated p53 destabilization. Genes Dev. 27:274–287. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Krajewska M, Krajewski S, Zapata JM, et
al: TRAF-4 expression in epithelial progenitor cells. Analysis in
normal adult, fetal, and tumor tissues. Am J Pathol. 152:1549–1561.
1998.PubMed/NCBI
|
|
22
|
Ye X, Mehlen P, Rabizadeh S, et al: TRAF
family proteins interact with the common neurotrophin receptor and
modulate apoptosis induction. J Biol Chem. 274:30202–30208. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Takeshita F, Ishii KJ, Kobiyama K, et al:
TRAF4 acts as a silencer in TLR-mediated signaling through the
association with TRAF6 and TRIF. Eur J Immunol. 35:2477–2485. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Esparza EM and Arch RH: TRAF4 functions as
an intermediate of GITR-induced NF-kappaB activation. Cell Mol Life
Sci. 61:3087–3092. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Abell AN and Johnson GL: MEKK4 is an
effector of the embryonic TRAF4 for JNK activation. J Biol Chem.
280:35793–35796. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zepp JA, Liu C, Qian W, et al: Cutting
edge: TNF receptor-associated factor 4 restricts IL-17-mediated
pathology and signaling processes. J Immunol. 189:33–37. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yang WL, Wang J, Chan CH, et al: The E3
ligase TRAF6 regulates Akt ubiquitination and activation. Science.
325:1134–1138. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics for Hispanics/Latinos, 2012. CA Cancer J Clin.
62:283–298. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Gu X, Coates PJ, MacCallum SF, et al:
TRAF4 is potently induced by TAp63 isoforms and localised according
to differentiation in SCCHN. Cancer Biol Ther. 6:1986–1990. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Rozan LM and El-Deiry WS: Identification
and characterization of proteins interacting with Traf4, an
enigmatic p53 target. Cancer Biol Ther. 5:1228–1235. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sax JK and El-Deiry WS: Identification and
characterization of the cytoplasmic protein TRAF4 as a
p53-regulated proapoptotic gene. J Biol Chem. 278:36435–36444.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rousseau A, McEwen AG,
Poussin-Courmontagne P, et al: TRAF4 is a novel
phosphoinositide-binding protein modulating tight junctions and
favoring cell migration. PLoS Biol. 11:e10017262013. View Article : Google Scholar
|
|
33
|
Wang X, Jin C, Tang Y, Tang LY and Zhang
YE: Ubiquitination of tumor necrosis factor receptor-associated
factor 4 (TRAF4) by Smad ubiquitination regulatory factor 1
(Smurf1) regulates motility of breast epithelial and cancer cells.
J Biol Chem. 288:21784–21792. 2013. View Article : Google Scholar
|
|
34
|
Ahmed F, Shiraishi T, Vessella RL and
Kulkarni P: Tumor necrosis factor receptor associated factor-4: an
adapter protein overexpressed in metastatic prostate cancer is
regulated by microRNA-29a. Oncol Rep. 30:2963–2968. 2013.
|
|
35
|
Zhang L, Zhou F, Garcia de Vinuesa A, et
al: TRAF4 promotes TGF-β receptor signaling and drives breast
cancer metastasis. Mol Cell. 51:559–572. 2013.PubMed/NCBI
|
|
36
|
Li W, Peng C, Lee MH, et al: TRAF4 Is a
critical molecule for Akt activation in lung cancer. Cancer Res.
73:6938–6950. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Brazil DP, Park J and Hemmings BA: PKB
binding proteins. Getting in on the Akt. Cell. 111:293–303. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Manning BD and Cantley LC: AKT/PKB
signaling: navigating downstream. Cell. 129:1261–1274. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Liu P, Cheng H, Roberts TM and Zhao JJ:
Targeting the phosphoinositide 3-kinase pathway in cancer. Nat Rev
Drug Discov. 8:627–644. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
40
|
Bose S, Chandran S, Mirocha JM and Bose N:
The Akt pathway in human breast cancer: a tissue-array-based
analysis. Mod Pathol. 19:238–245. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Stal O, Perez-Tenorio G, Akerberg L, et
al: Akt kinases in breast cancer and the results of adjuvant
therapy. Breast Cancer Res. 5:R37–R44. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Hay N: The Akt-mTOR tango and its
relevance to cancer. Cancer Cell. 8:179–183. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Brazil DP and Hemmings BA: Ten years of
protein kinase B signalling: a hard Akt to follow. Trends Biochem
Sci. 26:657–664. 2001.PubMed/NCBI
|
|
44
|
Yang WL, Wu CY, Wu J and Lin HK:
Regulation of Akt signaling activation by ubiquitination. Cell
Cycle. 9:487–497. 2010.PubMed/NCBI
|