Introduction
Bladder cancer is one of the most common urologic
malignant disease and shows an increasing tendency worldwide
(1). Tremendous difference in
prognosis is noted among patients with muscle-invasive bladder
cancer under the same condition of pathological stage and grade
(2). Therefore, heterogeneity plays
a crucial role in the metastatic potential risk for muscle-invasive
bladder cancer patients. In recent decades, even though much
research based on bladder cancer has been performed, the biological
basis of this disease remains enigmatic (3). Our research based on proteomics is one
way to explore the carcinogenic mechanism. In the present study, we
performed quantitative research on muscle-invasive bladder cancer.
The isobaric tag for relative and quantitation (iTRAQ) technique
was employed to quantitatively analyze the protein expression
levels of samples while covering the shortage of the traditional
shot-gun strategy. The quantitative analysis of proteins also
contributes to bladder cancer biomarker panel findings.
In recent years, the vital role of cancer stroma in
metastatic potential heterogeneity has been the focus of cancer
research (4). Research indicates
that the dynamic changes in the extracellular matrix and changes in
stromal biology signaling cascades are correlated with tumor
metastatic potential (5). Tumor
heterogeneity is influenced by several factors, such as the genetic
contribution, cancer stem cell differentiation and the tumor
microenvironment interaction. Regarding the tumor microenvironment,
several types of non-neoplastic cells, such as cancer-associated
fibroblasts, endothelial cells and immune cells, together
constitute the tumor stroma and influence the heterogeneity of the
tumor in various ways. Tumor heterogeneity can also be affected by
bioactive proteins, such as immune and inflammatory factors
(6). Due to changes in bioactive
proteins, the biological pathway is altered correspondingly and
reflects the features of heterogeneity in the tumor. In the present
study, we aimed to explore the muscle-invasive bladder cancer
heterogeneity by analyzing changes in the biological pathways.
Materials and methods
Patients and tissue samples
This study included a total of 30 patients, derived
from 3 risk groups. Thirty samples (cancerous and normal urothelial
tissues confirmed by 2 individual pathological diagnoses) were
obtained from patients treated at The Affiliated Hospital of
Qingdao University immediately after radical cystectomy due to
primary invasive bladder cancer (7). The cancerous and the adjacent
microscopically normal urothelial (5 cm away) samples were washed 3
times in 5 ml of sterile PBS and then flash frozen in liquid
nitrogen within 30 min of removal. Patient consent forms were
obtained and tissue-banking procedures were approved by the
Institutional Review Board.
Laser capture microdissection
Bladder cancer tissue sections were stained with
toluidine blue to guide microdissection. Sections (8-μm) were
air-dried and cut with a Leica AS LMD Laser Capture Microdissection
System (Sunnyvale, CA, USA). Approximately 300,000 shoots of tumor
or normal urothelial cells from each sample were microdissected and
stored on microdissection caps. The cells were captured within 120
min to protect protein from degradation. Cells of interest were
dissolved in lysis buffer (65 mM DTT, 4% CHAPS, 95 mM urea, 40% mM
Tris). Subesquently, specimens were solubilized via sonication
using 20-sec bursts, followed by ice cooling (20 sec), in a process
that was repeated 5 times. After completing the above steps, the
crude tissue extracts were centrifuged for 45 min at 15,000 rpm to
remove the biomembranes, DNA, and other undissolved materials.
These tissues were flash frozen in liquid nitrogen and stored at
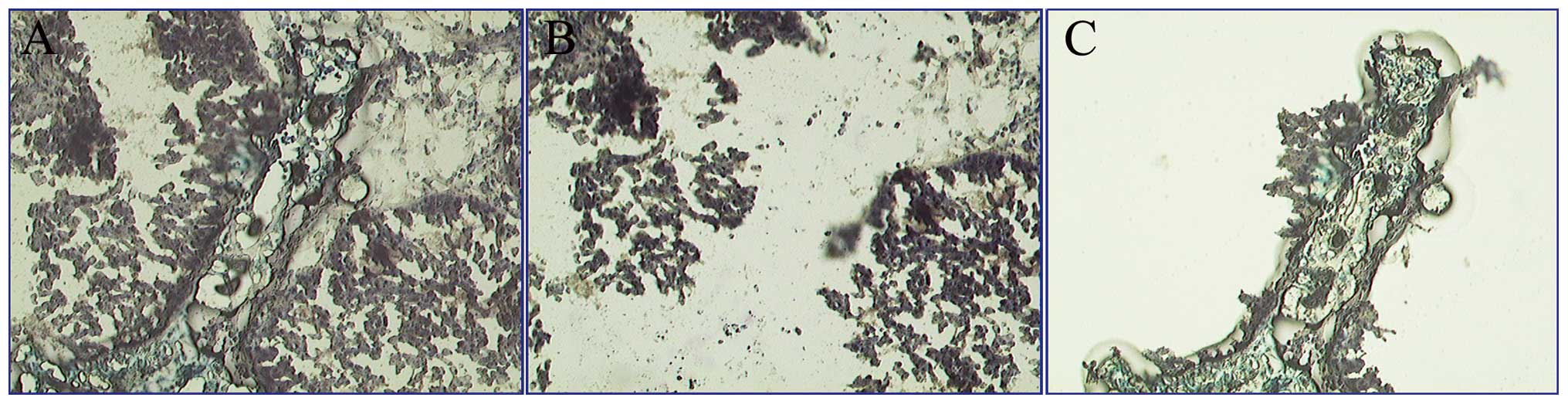
−80°C until use. Fig. 1 shows a
representative LCM process of a specimen.
Sample preparation
Two hundred micrograms of cancer stromal tissue from
10 cases/group was prepared and digested with trypsin. A total of
100 μg of peptides from each group was labeled with iTRAQ reagents
following the manufacturer’s instructions (Applied Biosystems).
Normal group, high risk group, median risk group, and low risk
group were labeled with 114, 115, 116 and 117 tags,
respectively.
Strong cation exchange (SCX) separation
conditions
Each set of labeled specimens was dissolved in 10%
formic acid and diluted with SCX solvent A (25 mM ammonium formate
in 20% acetonitrile, pH 2.8). After that, samples were loaded onto
a PolyLC polysulfoethyl A SCX column (2.1-mm internal diameter ×
100-mm length, 5 μm beads with 300 Å pores). Separation was
performed using a linear gradient over 45 min on an ÄKTA purifier
system (GE Healthcare, Buckinghamshire, UK). The parameter settings
were as follows: flow rate 0.25 ml/min; 0% B (500 mM ammonium
formate, pH 2.8, 20% ACN) for 5 min; 0–40% B for 20 min; 40–100% B
for 10 min; and 100% B held for 10 min. These fractions were then
monitored with UV absorbance detection at 254 and 280 nm and dried
by speed vacuuming. The peptide-containing samples were desalted on
PepClean C18 spin columns following the manufacturer’s protocols
(Thermo Fisher Scientific).
2D LC-MS/MS analysis on LTQ-Orbitrap
Peptides were dried and resuspended in 0.1% formic
acid. LTQ-Orbitrap™ XL (Thermo Fisher Scientific) coupled with a
nano-LC system was employed to analyze the samples. The pre-column
(75-μm internal diameter × 45-mm length) and reversed phase column
(50-μm internal diameter × 45-mm length) were used to separate
peptides. The flow rate for the reversed phase column was 200
nl/min. Each run started from solvent A [(99.9% (v/v)
H2O, 0.1% (v/v) formic acid)] to 100% solvent B [(15.9%
(v/v) H2O, 84% (v/v) CH3CN and 0.1% (v/v)
formic acid)]. The LC eluent was nanosprayed into the LTQ-Orbitrap
XL. Collisional-induced dissociation (CID) was used to fragment the
precursor peptides and higher-energy collisional-induced
dissociation (HCD) was employed to quantify the selected peptides.
For LTQ-Orbitrap instruments, data were acquired with the following
parameters: MS1 scans and HCD MS2 scans were acquired in Orbitrap
with the resolution at 60,000 (m/z 400) and 7500 (m/z 400),
respectively; the instrument was operated in a data-dependent mode;
1 microscan for CID-MS2 with collision energy of 30%.
Database search and protein
quantification
MS data for each fraction were used to identify and
quantify proteins based on ProteomicsTools.3.1. (Thermo Fisher
Scientific). The database was searched using the Mascot™ search
engine. Protein identification was performed at a confidence
threshold of 95%. Precursor ion exclusion lists were used to
minimize redundancy. For protein quantification, the ratios of
iTRAQ reporter ion intensities from the raw data sets were used to
calculate fold changes between samples. The peptides (unique for a
given protein) were considered for relative quantitation, excluding
those common to other proteins within the same family. The ratios
were normalized to the mean value of the 50 ratios identified with
the highest number of peptides. Then the protein lists were
generated.
Pathway analysis
The names of the differentially expressed proteins
were converted from IPI database to SWISS-PROT database using Array
Track™ (Jefferson, AR, USA) software. Array Track offers a simple
query interface to retrieve information concerning human protein
expression profile, and provides direct connections to related
metabolic and regulatory pathways available from Kyoto Encyclopedia
of Genes and Genomes (KEGG) (8).
The SWISS-PROT names of the differentially expressed proteins were
used for pathway analysis based on Array Track. For statistical
analysis, a P-value for pathway enrichment was obtained using the
hypergeometric test, and P<0.05 was considered to indicate a
statistically significant difference.
Results
Identification of the differentially
expressed proteins
More than 1,000 proteins were identified and
quantified by iTRAQ technique and 2D LC-MS/MS. To obtain the
differentially expressed proteins, the protein profiles of
different metastatic potential risk groups were compared by paired
comparison (high risk vs. median, low risk; median vs. low risk), a
total of 3 comparisons. We accepted the differentially expressed
proteins as follows: i) proteins reappeared 3 times in the
triplicate experiments and ii) proteins were identified based on ≥2
peptides. Based on these criteria, a total of 1,049 proteins were
identified as differentially expressed proteins in our research. In
addition, when the protein expression level showed a ratio
fold-change ≥1.5 or ≤0.667 compared to the normal group
(high/median/low risk group vs. normal group, respectively, a total
of 3 comparisons), we accepted it as a significantly altered
protein. After the comparison, 510,549,548 proteins as
significantly altered proteins were present in the low/median/high
metastatic risk groups, respectively.
Analysis of the significant pathways
Differentially expressed proteins (363 of 510) with
SWISS-PROT numbers were analyzed by Array Track software (database
was accessed on Jun. 12, 2012); similarly 395 of 549 for the median
risk group and 388 of 548 for the high risk group. Pathway analysis
showed that 198 proteins were located in biological KEGG pathways
and were defined as potential biomarkers in the low risk group, and
220/205 proteins in the median/high risk groups, respectively.
These proteins were thought to be potential candidate biomarkers
for diagnosis or prognosis of bladder cancer. The distinctly
altered pathways mainly included focal adhesion, systemic lupus
erythematosus and ECM-receptor interaction pathway. Tables I and II show the major altered pathways and the
specifically expressed proteins in the different metastatic
potential groups.
 | Table IProteins located in the top 10 altered
pathways among the three metastatic potential risk groups. |
Table I
Proteins located in the top 10 altered
pathways among the three metastatic potential risk groups.
| SWISS-PROT name in
the 3 risk groups |
|---|
|
|
|---|
| Pathway | Low risk | Median risk | High risk |
|---|
| Systemic lupus
erythematosus | ACTN1,
ACTN4,a | ACTN1, C1S, C7,
H2AFY,a | C1QC, C1S,
C7,a |
| ECM-receptor
interaction | CD44,
THBS1,a |
THBS1,a | a |
| Amoebiasis | ACTN4, GNAS,
ITGAM,a | HSPB1,
a | |
| Focal adhesion | ACTN1, ACTN4, THBS1,
TLN1,a | ACTN1, THBS1,
TLN1,a |
GRB2,a |
| Dilated
cardiomyopathy (DCM) |
GNAS,a | a | a |
| Hypertrophic
cardiomyopathy (HCM) | a | a | a |
| Complement and
coagulation cascades | CD59,
PLG,a | C1S, C7, CFH, FGA,
FGB, FGG, SERPINA1,a | C1QC, C1R, C1S, C7,
CD59, CFH, FGA, FGB, FGG, PLG, SERPINA1, SERPIND1,
a |
| Protein processing in
endoplasmic reticulum | DNAJA2, DNAJB11,
EIF2S1, SKP1,a | | CAPN1, ERO1L,
HSP90AA1, HSP90AB1, HSP90B1, HYOU1, PDIA3, RPN1,
UBQLN1,a |
| Tight junction | a | | |
| Parkinson’s
disease | | a | |
| Alzheimer’s
disease | | a | |
| Cardiac muscle
contraction | | a | |
| Staphylococcus aureus
infection | | | a |
| Proteasome | | | a |
| Pertussis | | | a |
| Glutathione
metabolism | a | | |
| Table IICommonly expressed proteins in the
corresponding pathway. |
Table II
Commonly expressed proteins in the
corresponding pathway.
| Pathway | SWISS-PROT
namea |
|---|
| Systemic lupus
erythematosus | C3, C5, C9,
HIST1H2BL, HIST1H4A, HIST1H4B, HIST1H4C, HIST1H4D, HIST1H4E,
HIST1H4F, HIST1H4H, HIST1H4I, HIST1H4J, HIST1H4K, HIST1H4L,
HIST2H4A, HIST2H4B, HIST4H4, HLA-DRB3, SSB |
| ECM-receptor
interaction | COL1A1, COL6A1,
COL6A2, COL6A3, DAG1, FN1, LAMA2, LAMA4, LAMA5, LAMC1, TNXB |
| Amoebiasis | ACTN1, C9, COL1A1,
FN1, LAMA2, LAMA4, LAMA5, LAMC1, RAB5C, RAB7A, VCL |
| Focal adhesion | ACTB, COL1A1,
COL6A1, COL6A2, COL6A3, FLNA, FN1, LAMA2, LAMA4, LAMA5, LAMC1,
MYL12B, MYL9, PARVA, TNXB, VCL |
| Dilated
cardiomyopathy (DCM) | ACTB, ACTC1, DAG1,
DES, LAMA2, LMNA, TPM1, TPM2, TPM3, TPM4 |
| Hypertrophic
cardiomyopathy (HCM) | ACTB, ACTC1, DAG1,
DES, LAMA2, LMNA, TPM1, TPM2, TPM3, TPM4 |
| Complement and
coagulation cascades | A2M, C3, C4BPA, C5,
C9, F2, SERPINC1 |
| Protein processing
in endoplasmic | BAG2, BCAP31, CALR,
CRYAB, DDOST, UGGT1, ERP29, LMAN1, P4HB, reticulum PDIA6 |
| Tight junction | ACTB, ACTN1, ACTN4,
CTTN, EXOC4, MYH10, MYH9, MYL12B, MYL9, PPP2CA, RRAS, SPTAN1 |
| Parkinson’s
disease | ATP5D, ATP5H,
COX4I1, COX5B, COX6B1, COX7A2, NDUFA11, NDUFAB1, NDUFV1, SLC25A5,
PARK7, SNCA, UQCRC1, VDAC1, VDAC2, VDAC3 |
| Alzheimer’s
disease | APOE, ATP5D, ATP5H,
CALM1, CALM2, CALM3, CHP, COX4I1, COX5B, COX6B1, COX7A2, GAPDH,
NDUFA11, NDUFAB1, NDUFV1, SNCA, UQCRC1 |
| Cardiac muscle
contraction | ACTC1, COX4I1,
COX5B, COX6B1, COX7A2, TPM1, TPM2, TPM3, TPM4, UQCRC1 |
| Staphylococcus
aureus infection | C1QC, C1R, C1S, C3,
C5, CFH, FGG, HLA-DRB3, KRT10, PLG |
| Proteasome | PSMA7, PSMB8,
PSMC2, PSMC3, PSMC4, PSMD14, PSME3 |
| Pertussis | C1QC, C1R, C1S, C3,
C4BPA, C5, CALM1, CALM2, CALM3 |
| Glutathione
metabolism | GCLM, GPX3, GSR,
GSTM3, IDH1, MGST1, TXNDC12 |
Discussion
Bladder cancer is the most common urologic malignant
disease, and the features of the mutationally corrupted cells are
closely related to the tumorigenic microenvironment (5). Recently, most research concerning
bladder cancer has focused on the heterogeneity of the mutated
cells, but has neglected the vital role of the stroma which is one
of the crucial driving forces for tumor heterogeneity. After
protein detection by iTRAQ technique and 2D-LC-MS/MS analysis,
1,049 proteins were identified as being differentially expressed
among 3 metastatic risk groups. The smallest and largest molecular
weight (MW) values observed in the proteins were 5.5 and 629.1 kDa,
and the proteins were distributed across a wide isoelectric point
range (3.67–11.98). Several proteins have been reported previously
and a significantly altered expression level of these proteins is
closely related to tumor characteristics. For example, isocitrate
dehydrogenase cytoplasmic enzyme (IDHC) may defend against
oxidative damage of cells, and was significantly differentially
expressed among the 3 metastatic potential risk groups. Although
the specific role for the deregulation of IDHC in bladder cancer is
not clear, research indicates that the early loss of IDHC may
decrease the antioxidant capacity of cells and finally promote
tumor progression and metastasis (9).
ACTB (β-actin) is a cytoskeleton structural protein
and it regulates motility in several types of cells. In our study,
ACTB protein was distinctly weakly expressed in the stroma of
muscle-invasive bladder cancer. However, in regards to the
intracellular expression level, ACTB was reported to be highly
expressed in non-small cell lung cancer cell lines at the mRNA and
protein levels. Therefore, the different expression level of ACTB
inside and outside cancer cells might be a clue for tumorigenesis
(10–12). In our research, the expression level
of integrin α(M)β2 (ITAM) increased in turn from the
high risk group to the low risk groups (Table IV). Recently, Ma et al
provided a new insight for ITAM in the aspect of tumor metastatic
mechanism. ITAM, a type of immune adhesion molecule, is transferred
from innate immune cells to tumor cells via microparticles (derived
from innate immune cells). Then the tumor cells acquire a transient
immune phenotype and metastasize to distant organs. Due to the
protein derived from stromal cells of bladder cancer, the level of
ITAM had an inverse relationship between inside the cells and
outside the cells (13). Thus, the
different expression levels of ITAM in the 3 metastatic potential
risk groups may account for the metastatic potential heterogeneity
of bladder cancer.
| Table IVRelative expression levels of various
proteins in the 3 metastatic potential risk groups. |
Table IV
Relative expression levels of various
proteins in the 3 metastatic potential risk groups.
| Proteins | High/normal | Medium/normal | Low/normal |
|---|
| IDHC | 1.43 | 0.99 | 0.66 |
| ACTN4 | 1.27 | 0.75 | 0.54 |
| EXOC4 | 1.21 | 0.53 | 0.05 |
| MYH10 | 0.50 | 0.21 | 0.18 |
| ITAM | 0.71 | 1.12 | 1.66 |
| MMP-9 | 0.62 | 1.32 | 2.21 |
| MIF | 1.42 | 2.86 | 2.87 |
| FN1 | 1.57 | 1.68 | 2.33 |
| ACTB | 0.66 | 0.52 | 0.41 |
| A2M | 0.35 | 0.44 | 0.56 |
| CD44 | 0.75 | 0.74 | 0.45 |
| GRB2 | 0.62 | 0.83 | 0.74 |
Multiple altered biological pathways were present in
the muscle-invasive bladder cancer and might play a vital role in
regulating the metastatic potential heterogeneity of malignant
disease. In our study, 510,549,548 proteins were identified as
being significantly altered in the low, median and high metastatic
potential risk groups, respectively. Two hundred and seventy-one
(low risk), 264 (median risk), 208 (high risk) proteins were
located in KEGG pathways using Array track software. As shown in
Table III, our results
illustrated that the divergently altered biological pathways
presented in the 3 metastatic potential risk groups may play a key
role in regulating the metastatic potential heterogeneity of
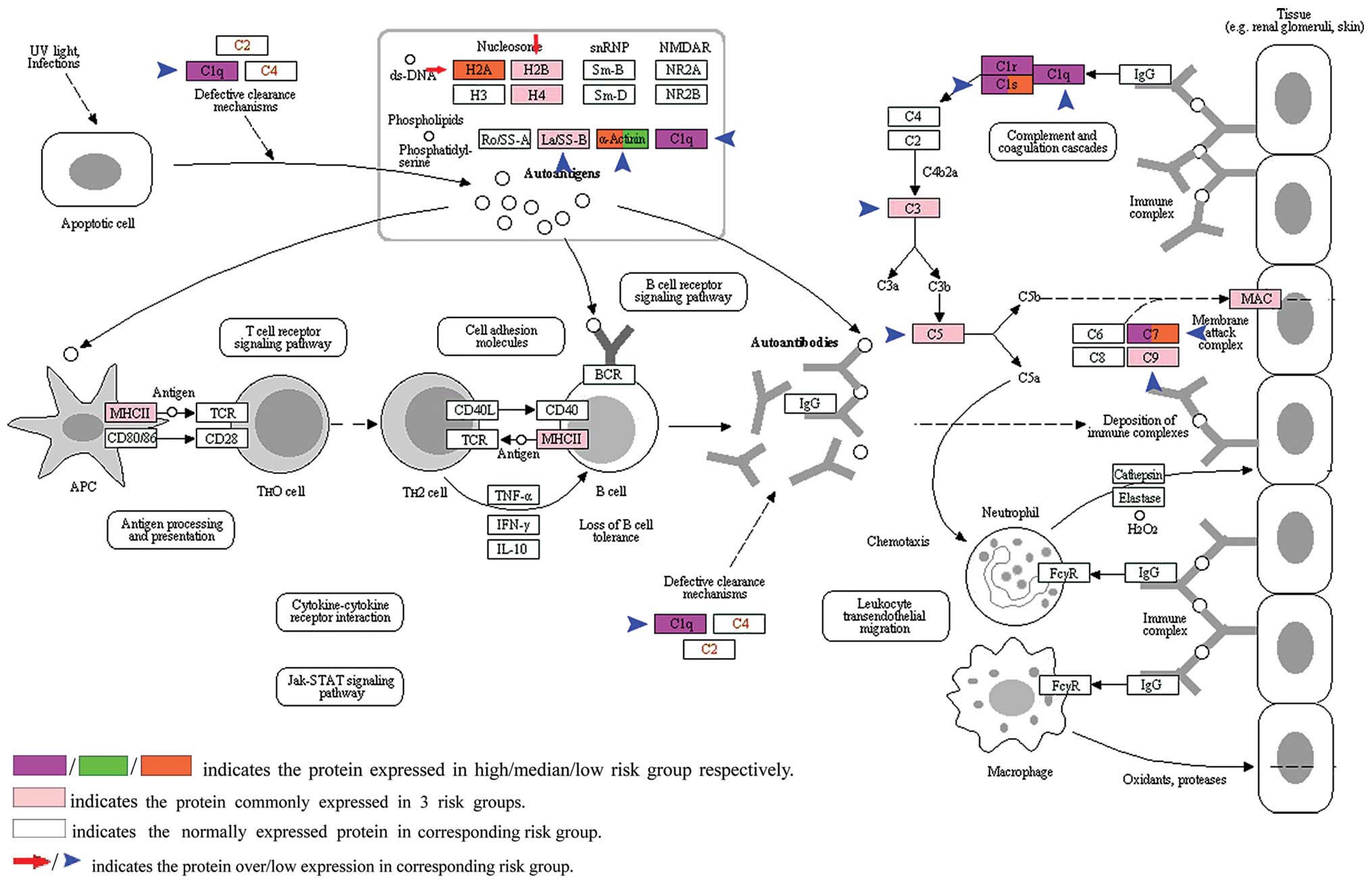
muscle-invasive bladder cancer. The KEGG pathway of systemic lupus
erythematosus (SLE) was one of the significantly altered biological
pathways. Immune system dysfunction has been reported to play a
vital role in the metastasis of neoplasms (13). Thus, the differentially altered KEGG
pathway of SLE among the 3 risk groups (Fig. 3 and Table III) illustrate that immunological
dysregulation is one driving force resulting in tumor metastatic
potential heterogeneity. The KEGG pathway of SLE was the most
prominently altered pathway noted in the list of altered pathways
in the low/median metastatic potential risk groups (Table III). There are several types of
immunocytes presented on the KEGG SLE pathway map, such as
macrophages, neutrophils, antigen-presenting cells.
| Table IIITop 5 altered pathways in the 3
metastatic potential risk groups. |
Table III
Top 5 altered pathways in the 3
metastatic potential risk groups.
| High risk
group | Medium risk
group | Low risk group |
|---|
| Complement and
coagulation cascades | Systemic lupus
erythematosus | Systemic lupus
erythematosus |
| Systemic lupus
erythematosus | Complement and
coagulation cascades | ECM-receptor
interaction |
| Protein processing
in endoplasmic reticulum | Parkinson’s
disease | Amoebiasis |
| Staphylococcus
aureus infection | ECM-receptor
interaction | Focal adhesion |
| ECM-receptor
interaction | Alzheimer’s
disease | Dilated
cardiomyopathy (DCM) |
Macrophage migration inhibitory factor (MIF),
upregulated in the 3 metastatic groups (Table IV), is thought to be released from
monocytes/macrophages (14) and
acts as a regulator of innate immunity. Jung et al reported
that overexpression of MIF can suppress the tumor-suppressor gene
p53, leading to uncontrolled cell growth (15). In addition, overexpression of MIF
can result in upregulation of hypoxia inducible factor-1 (HIF-1);
this change leads to the overexpression of several genes, such as
matrix metalloproteinases (MMPs) and vascular endothelial growth
factor (VEGF), consequently promoting tumor metastasis and
angiogenesis (16). The
differential expression level of MIF among the different risk
groups reflects the metastatic potential heterogeneity of
muscle-invasive bladder cancer. In the high metastatic potential
risk group, the KEGG complement and coagulation cascade pathway was
shown as the most prominently altered one (Table III). Nineteen proteins located in
this pathway are thought to be candidate biomarkers for bladder
cancer (Tables I and II). Among them, α-2-macroglobulin (A2M)
is one of the serum proteinase inhibitors located in this pathway
(Table II). Research has shown
that the serum level of A2M has an inverse relationship with PSA in
prostate cancer patients and A2M (activated state) can promote
tumor development and progression by mediating proliferation,
pro-migratory, and anti-apoptotic signaling mechanisms (17,18).
The other KEGG pathways, including protein processing in
endoplasmic reticulum, Staphylococcus aureus infection,
ECM-receptor interaction, focal adhesion and tight junction, were
also divergently ranked among the 3 metastatic risk groups
(Table III). It can be derived
from the analysis of the data that the simultaneous change of
multi-pathways is one of the effective ways to account for the
metastatic potential heterogeneity in terms of muscle-invasive
bladder cancer.
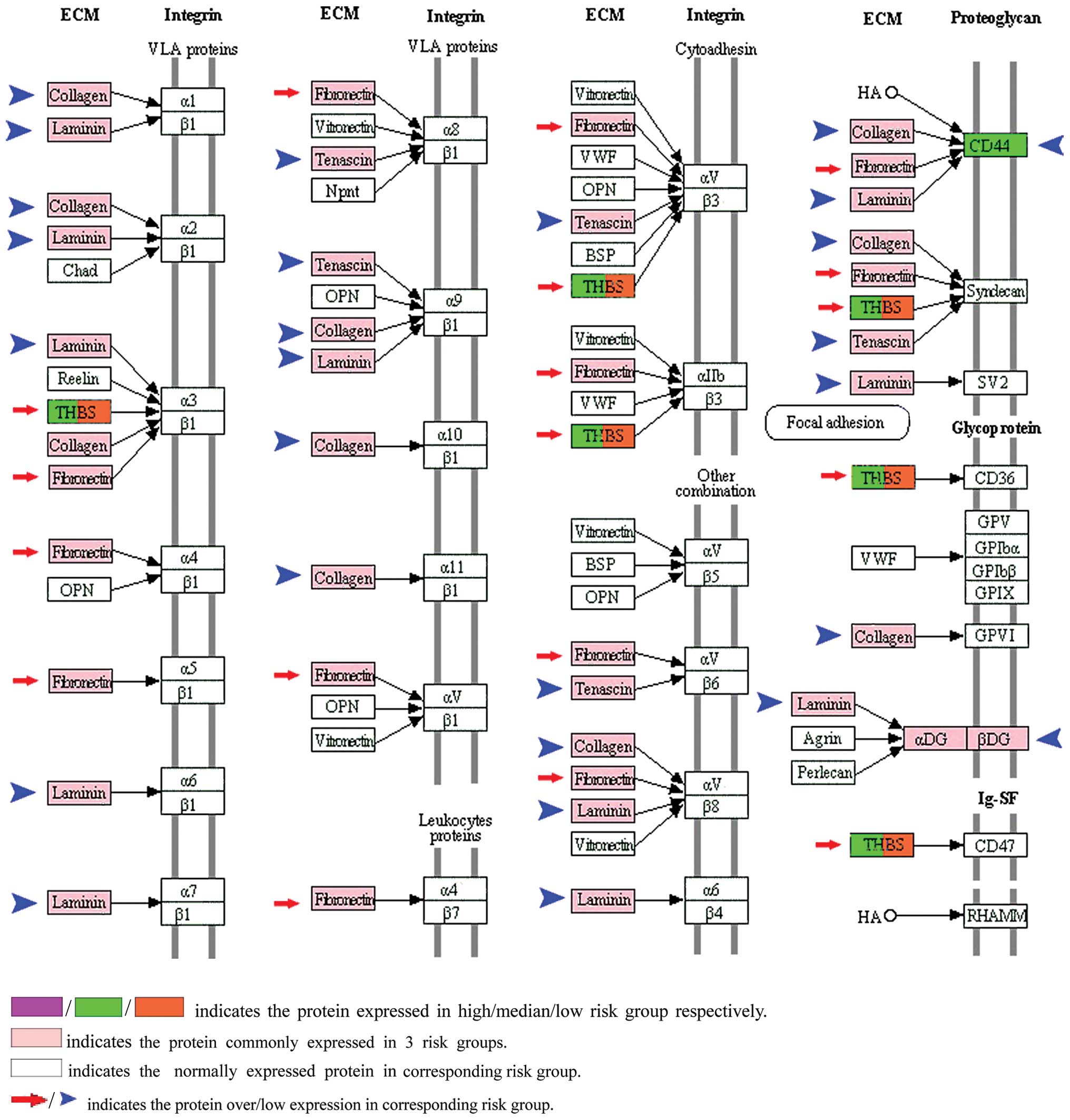
To clarify the alterations in pathways, it is
necessary to understand the heterogeneity of tumors. In our study,
the KEGG ECM-receptor interaction pathway was confirmed as one of
the significantly altered pathways among the 3 risk groups.
According to the P-value, all altered pathways were ranked in
ascending order. The KEGG ECM-receptor interaction pathway ranked
5th in the high risk group, 4th in the median risk group and 2nd in
the low risk group, respectively (Table III). Fibronection (FN1) is a
non-collagenous glycoprotein (19)
and was upregulated in our results (Table IV). The other proteins, such as
collagen, laminin and tenascin were downregulated in the protein
list (Fig. 2). All of these
proteins were located in the KEGG ECM-receptor interaction pathway.
Ioachim et al showed that the expression of tenascin,
fibronectin and collagen appears to be correlated with more
aggressive tumor behavior, and their interrelationship could
influence tumor progression by remodeling bladder cancer tissue
(20). In addition, CD44 was
abnormally expressed in the low risk group (Table I). CD44 is a transmembrane
glycoprotein which acts as a ‘node’ to interact with collagen,
fibronectin and laminin in the KEGG ECM-receptor interaction
pathway (Fig. 2). Henke et
al indicated that malignant cells used CD44-related chondroitin
sulfate proteoglycan as a matrix receptor to mediate metastasis
(21). Therefore, the divergently
altered KEGG ECM-receptor interaction pathways among the 3
metastatic risk groups reflect that the biological pathways may be
pivotal for regulating the metastatic heterogeneity of bladder
cancer. Moreover, the KEGG focal adhesion pathway functions as the
sub-pathway of the ECM-receptor interaction pathway, and was
cross-linked by integrin α and β, which makes it another
significantly altered pathway in our study. According to the
P-value, the pathway lists showed that the KEGG focal adhesion
pathway ranked 7th in the high risk group and 6th in the median and
4th in the low metastatic risk group. Actinin-4 (ACTN4) and growth
factor receptor-bound protein 2 (GRB2) act as candidate biomarkers
located in the focal adhesion pathway. ACTN4 was specifically
located in the low risk metastatic potential group, and GRB2 in the
high risk metastatic potential risk group, respectively. For ACTN4,
Yamamoto et al indicated that the ACTN4 gene, mapped to the
chromosomal band 19q13.1-q13.2, was commonly amplified in ovarian
cancer cells. The actinin-4 protein was found to play an important
role in the pathogenesis of ovarian cancer (22). In addition, GRB2 acts as a
downstream intermediary in several oncogenic signaling pathways.
The human GRB2 gene is located in chromosome 17q22 which is known
to be duplicated in solid tumors. GRB2 signaling has been
implicated in the pathogenesis of several human cancers (23). Other proteins located in the KEGG
focal adhesion pathway were thrombospondin- 1 (THBS1) and talin-1
(TLN1) (Tables I and II). From another perspective, mechanical
signaling plays a crucial role in the metastasis of tumors. Cells
receive mechanical signals from the extracellular matrix through
focal adhesions. Focal adhesions are the major sites of
interactions between extracellular mechanical signals and
intracellular biochemical signaling molecules. Cells respond to
signals through integrinrelated signaling pathways to maintain
cellular migration function. Accordingly, the above analysis
provides effective support for the theory which states that
biological pathway change may play a crucial role in regulating
bladder cancer metastatic heterogeneity (24).
Another contribution of pathway analysis is the
confirmation of candidate biomarkers. The protein exocyst complex
component 4 (EXOC4) is a key protein in the KEGG tight junction
pathway and was expressed differentially among the 3 metastatic
risk groups (Table IV). Yamamoto
et al found that EXOsec8 (aliases for EXOC4) was
overexpressed in oral squamous-cell carcinoma (OSCC) and plays a
crucial role in OSCC progression by promoting secretion of matrix
metalloproteinases (MMPs) (25).
MMPs can induce gaps in the ECM to promote cancer metastasis by
degrading associated proteins (24). The abnormal expression of protein
EXOC4 was first identified in muscle-invasive bladder cancer in our
study and EXOC4 may act as a candidate biomarker in bladder cancer.
In addition, MMP-9 is another protein whose expression level was
significantly altered among the 3 risk groups (Table IV). Some researchers have shown
that the MMP gene contained the E2F (E2F is a group of genes that
codifies a family of transcription factors) binding site in the
promoter region and is regulated by E2F transcription factors which
play a crucial role in promoting metastasis (26). Ramón de Fata et al found that
patients with bladder cancer had a higher level of MMP-9 in the
serum than controls, and serum MMP-9 has application in the
prediction of the progression of bladder cancer (27). Myosin-10 (MYH10), one of the
cytoskeletal regulators, was located in the KEGG tight junction
pathway. The expression level is shown in Table IV. MYH10 can regulate mitosis and
endothelial cell migration by interacting with microfilaments and
microtubules (28). Although little
information is available to show the role of myosin-10 in human
bladder cancer progression, based on our results, myosin-10 may
affect the prognosis of human bladder cancer and is thought to be a
candidate biomarker (28,29).
In conclusion, based on our results, 1,049 proteins
were identified as being differentially expressed and 510,549,548
proteins as being significantly altered proteins in the
low/median/high metastatic risk groups, respectively. Pathway
analysis shows that the differentially expressed proteins are
mainly located in the systemic lupus erythematosus pathway,
ECM-receptor interaction, focal adhesion, and tight junction
pathways. Our study confirmed that stroma is an important part of
the tumor and plays a vital role in regulating the heterogeneity of
muscle-invasive bladder cancer. Commonly altered biological
pathways determine the malignant phenotype of muscle-invasive
bladder cancer. Biomarker discovery should take into account both
neoplastic cells and the corresponding stroma.
Acknowledgements
This research was supported by a grant from the
National Natural Science Foundation of China (nos. 30901481,
81372752, 81101932), the Wu Jieping Medical Foundation
(320.6750.13261) and the Doctoral Science Foundation of Shandong
Province, China (BS2010YY009).
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
Statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar
|
|
2
|
Pasin E, Josephson DY, Mitra AP, Cote RJ
and Stein JP: Superficial bladder cancer: an update on etiology,
molecular development, classification, and natural history. Rev
Urol. 10:31–43. 2008.PubMed/NCBI
|
|
3
|
Knowles MA: Molecular subtypes of bladder
cancer: Jekyll and Hyde or chalk and cheese? Carcinogenesis.
27:361–373. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hasebe T: Tumor-stromal interactions in
breast tumor progression - significance of histological
heterogeneity of tumor-stromal fibroblasts. Expert Opin Ther
Targets. 17:449–460. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
van der Horst G, Bos L and van der Pluijm
G: Epithelial plasticity, cancer stem cells, and the
tumor-supportive stroma in bladder carcinoma. Mol Cancer Res.
10:995–1009. 2012.PubMed/NCBI
|
|
6
|
De Sousa E, Melo F, Vermeulen L, Fessler E
and Medema JP: Cancer heterogeneity - a multifaceted view. EMBO
Rep. 14:686–695. 2013.PubMed/NCBI
|
|
7
|
Niu HT, Shao SX, Zhang ZL, et al:
Quantitative risk stratification and individual comprehensive
therapy for invasive bladder cancers in China. Int Urol Nephrol.
41:571–577. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tong W, Harris S, Cao X, et al:
Development of public toxicogenomics software for microarray data
management and analysis. Mutat Res. 549:241–253. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Memon AA, Chang JW, Oh BR and Yoo YJ:
Identification of differentially expressed proteins during human
urinary bladder cancer progression. Cancer Detect Prev. 29:249–255.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Guo C, Liu S, Wang J, Sun MZ and Greenaway
FT: ACTB in cancer. Clin Chim Acta. 417:39–44. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Andersen CL, Jensen JL and Ørntoft TF:
Normalization of real-time quantitative reverse transcription-PCR
data: a model-based variance estimation approach to identify genes
suited for normalization, applied to bladder and colon cancer data
sets. Cancer Res. 64:5245–5250. 2004. View Article : Google Scholar
|
|
12
|
Saviozzi S, Cordero F, Lo Iacono M,
Novello S, Scagliotti GV and Calogero RA: Selection of suitable
reference genes for accurate normalization of gene expression
profile studies in non-small cell lung cancer. BMC Cancer.
6:2002006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ma J, Cai W, Zhang Y, et al: Innate immune
cell-derived microparticles facilitate hepatocarcinoma metastasis
by transferring integrin α(M)β2 to tumor cells. J
Immunol. 191:3453–3461. 2013.PubMed/NCBI
|
|
14
|
Nishihira J, Ishibashi T, Fukushima T, Sun
B, Sato Y and Todo S: Macrophage migration inhibitory factor (MIF):
Its potential role in tumor growth and tumor-associated
angiogenesis. Ann NY Acad Sci. 995:171–172. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jung H, Seong HA and Ha H: Critical role
of cysteine residue 81 of macrophage migration inhibitory factor
(MIF) in MIF-induced inhibition of p53 activity. J Biol Chem.
283:20383–20396. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Oda S, Oda T, Nishi K, et al: Macrophage
migration inhibitory factor activates hypoxia-inducible factor in a
p53-dependent manner. PLoS One. 3:e22152008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kanoh Y, Ohtani H, Egawa S, Baba S and
Akahoshi T: Changes of proteases and proteinase inhibitors in
androgen-dependent advanced prostate cancer patients with
alpha2-macroglobulin deficiency. Clin Lab. 58:217–225. 2012.
|
|
18
|
Misra UK, Payne S and Pizzo SV: Ligation
of prostate cancer cell surface GRP78 activates a proproliferative
and antiapoptotic feedback loop: a role for secreted
rostate-specific antigen. J Biol Chem. 286:1248–1259. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Brunner A and Tzankov A: The role of
structural extracellular matrix proteins in urothelial bladder
cancer (review). Biomark Insights. 2:418–427. 2007.PubMed/NCBI
|
|
20
|
Ioachim E, Michael M, Stavropoulos NE,
Kitsiou E, Salmas M and Malamou-Mitsi V: A clinicopathological
study of the expression of extracellular matrix components in
urothelial carcinoma. BJU Int. 95:655–659. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Henke CA, Roongta U, Mickelson DJ, Knutson
JR and McCarthy JB: CD44-related chondroitin sulfate proteoglycan,
a cell surface receptor implicated with tumor cell invasion,
mediates endothelial cell migration on fibrinogen and invasion into
a fibrin matrix. J Clin Invest. 97:2541–2552. 1996. View Article : Google Scholar
|
|
22
|
Yamamoto S, Tsuda H, Honda K, et al: ACTN4
gene amplification and actinin-4 protein overexpression drive
tumour development and histological progression in a high-grade
subset of ovarian clear-cell adenocarcinomas. Histopathology.
60:1073–1083. 2012. View Article : Google Scholar
|
|
23
|
Giubellino A, Burke TR Jr and Bottaro DP:
Grb2 signaling in cell motility and cancer. Expert Opin Ther
Targets. 12:1021–1033. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Seong J, Wang N and Wang Y:
Mechanotransduction at focal adhesions: from physiology to cancer
development. J Cell Mol Med. 17:597–604. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yamamoto A, Kasamatsu A, Ishige S, et al:
Exocyst complex component Sec8: a presumed component in the
progression of human oral squamous-cell carcinoma by secretion of
matrix metalloproteinases. J Cancer Res Clin Oncol. 139:533–542.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Johnson JL, Pillai S, Pernazza D, Sebti
SM, Lawrence NJ and Chellappan SP: Regulation of matrix
metalloproteinase genes by E2F transcription factors. Rb-Raf-1
interaction as a novel target for metastatic disease. Cancer Res.
72:516–526. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ramón de Fata F, Ferruelo A, Andrés G,
Gimbernat H, Sánchez-Chapado M and Angulo JC: The role of matrix
metalloproteinase MMP-9 and TIMP-2 tissue inhibitor of
metalloproteinasas as serum markers of bladder cancer. Actas Urol
Esp. 37:480–488. 2013.PubMed/NCBI
|
|
28
|
Woolner S, O’Brien LL, Wiese C and Bement
WM: Myosin-10 and actin filaments are essential for mitotic spindle
function. J Cell Biol. 182:77–88. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ju XD, Guo Y, Wang NN, et al: Both
Myosin-10 isoforms are required for radial neuronal migration in
the developing cerebral cortex. Cereb Cortex. 24:1259–1268.
2013.PubMed/NCBI
|