Introduction
As a member of the kinin group, bradykinin (BK) is
an active peptide that is generated by the kallikrein-kinin system
(KKS) (1). It is reported that BK
is involved in the regulation of various cellular processes in
cancer cells including tumorigenesis, angiogenesis, cell cycle and
proliferation (2). BK exerts its
biological functions mainly via the binding of bradykinin
receptors, which are pharmacologically characterized as bradykinin
B1 receptor (B1R) and bradykinin B2 receptor (B2R) (3). B1R and B2R both belong to the
G-protein coupled receptors (4).
B1R induces COX-2 production and cell migration in glioma cells
(5), whereas B2R promotes VEGF
expression and increases angiogenesis in prostate cancer cells
(6). Therefore, activation of these
two receptors by bradykinin may be essential for the development
and progression of cancer.
Colorectal cancer is one of the most common
malignancies worldwide. Over 600,000 deaths from colorectal cancer
are estimated to occur annually worldwide, thus ranking it as the
fourth cause of death among cancer patients (7). Although therapeutic strategies have
been well developed, colorectal cancer often progresses, along with
cell invasion and metastasis (8).
Invasion and metastasis are still the major cause of cancer-related
mortality (9). Studies have found
that BK can induce MMP-3 expression in human colonic
myofibroblasts, which may then contribute to the pathophysiology
underlying colitis-associated cancer (10). However, the role of BK in colorectal
cancer remains elusive. In this study, we examined the effect of BK
treatment on the invasion and migration of colorectal cancer cells.
We further characterized the function of B2R in BK-mediated
invasion and migration, and determined the possible underlying
mechanisms.
Materials and methods
Reagents
The antibodies against B1R (rabbit monoclonal
antibody), B2R (mouse monoclonal antibody), β-actin (mouse
monoclonal antibody) and ERK1/2 (rabbit polyclonal antibody) were
obtained from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA).
The antibody against phospho-ERK1/2 (rabbit polyclonal antibody)
was obtained from Cell Signaling Technology, Inc. (Beverly, MA,
USA). BK and PD98059 were purchased from Sigma (St. Louis, MO,
USA). IL-6 ELISA kit was purchased from Boster (Wuhan, China).
Cell culture
Human colorectal cancer cell lines, T84, Caco-2,
HT-29, HCT116 and SW480, were purchased from the American Type
Culture Collection (Manassas, VA, USA). Cells were maintained in
Dulbecco’s modified Eagle’s medium (DMEM; Gibco-BRL, Rockville, MD,
USA) with 10% fetal bovine serum (FBS; Gibco-BRL) at 37°C in a
humidified CO2 incubator.
Invasion assay
A 24-well Transwell plate, containing PET membranes
with 8-μm pores, was purchased from Costar (San Diego, CA,
USA). The upper chambers of the plate were coated with thin layers
of Matrigel (BD Biosciences, Franklin Lakes, NJ, USA). Cells
(1.0×105 cells in 0.2 ml DMEM/well) were pretreated with
different dosages of bradykinin, and then were added to the upper
chambers. The chemoattractant (20% FBS) was placed into the lower
chambers. After incubation in a CO2 incubator for 16 h,
the cells that invaded through the membrane were fixed and stained
with eosin. The invaded cells were observed under a microscope
(magnification, ×100), and the number of the invaded cells was
counted from five random fields. All experiments were conducted in
triplicate, and the data are presented as a percentage of the
invaded cells in the control.
Migration assay
The procedure for the migration assay was similar to
the invasion assay described above, except that the upper chambers
were coated without Matrigel. Briefly, cells were adjusted to the
concentration of 5.0×105 cells/ml. Two hundred
microliters of the cell suspensions was placed in the upper
chambers, while 20% FBS was placed into the lower chambers. Sixteen
hours later, the migrated cells were stained with eosin. The number
of migrated cells was counted from five random fields, and the data
are presented as a percentage of the migrated cells in the
control.
Western blot analysis
Cells were lysed in 200 μl lysis buffer,
which contained 50 mM Tris-HCl (pH 7.5), 1% NP-40, 150 mM NaCl, 1
mg/ml aprotinin, 1 mg/ml leupeptin, 1 mM
Na3VO4 and 1 mM NaF. The protein
concentrations were determined by BCA assay. Equal amounts of cell
lysate (50 μg) were resolved by 10% SDS-PAGE and transferred
to a PVDF membrane (Pall Corp., Alcobendas, Spain). After blocking
with 5% non-fat milk (for detection of B1R, B2R, β-actin and
ERK1/2) or 5% BSA (for detection of p-ERK1/2) for 1 h at room
temperature, the membranes were incubated with primary antibodies
overnight at 4°C. Then the membranes were washed with TBST for
three times and further incubated with the secondary antibodies for
1 h at room temperature. The bands were detected with an enhanced
chemiluminescence kit (Applygen Technologies Inc., Beijing, China),
according to the manufacturer’s instructions. The density of the
bands was quantified with Quantity One software.
siRNA transfection
siRNAs were designed and chemically synthesized by
Genepharma (Shanghai, China). The sequences of the siRNAs are
listed as follows: control siRNA, 5′-UGG UUUACAUGUUUUCUGA-3′; B1R
siRNA, 5′-GCUCCC UGAAUCCAGUAAU-3′; B2R siRNA, 5′-GGCAGAGGA
AGAUAUUUCU-3′; IL-6 siRNA, 5′-CCCGGACCAUAU UUAUUAU-3′. Cells were
transfected with siRNAs using Lipofectamine 2000™ (Invitrogen,
Carlsbad, CA, USA) when the cell confluency reached 30–40%. The
knockdown efficiency was determined by western blotting or
real-time PCR.
ELISA assay
The level of IL-6 in the culture supernatant was
observed by ELISA assay, according to the manufacturer’s
instructions. Briefly, cells were treated with or without BK for 16
h, and then 50 μl of the cell supernatant was added into the
ELISA plate wells to detect the level of IL-6. The absorbance was
measured at 450 nm in a microplate reader. The concentration of
IL-6 was examined by comparing the absorbance values against the
standard curve. The level of IL-6 in each sample was quantified to
the total protein.
Real-time PCR
Total-RNA was isolated from the SW480 cells by using
TRIzol reagent (Invitrogen, Carlsbad, CA, USA). The concentrations
of RNA were measured with NanoDrop ND-1000 (Thermo Fisher
Scientific, Wilmington, DE, USA). The reverse transcription PCR
reaction was carried out with 2 μg of RNA and M-MLV reverse
transcriptase (Tiangen, Beijing, China) to obtain cDNA. Then the
real-time PCR reaction was performed with cDNA and primers.
Oligonucleotide primers are listed as follows: IL-6 sense,
5′-TGCGTCCGTAGTTTCCTTCT-3′ and antisense,
5′-GCCTCAGACATCTCCAGTCC-3′; β-actin sense,
5′-AGAAGGATTCCTATGTGGGCG-3′ and antisense,
5′-CATGTCGTCCCAGTTGGTGAC-3′. The reaction was performed for 5 min
at 95°C, followed by 40 cycles of 95°C for 15 sec, and 60°C for 1
min.
Statistical analysis
All experiments were repeated three to four times,
and the data are presented as the mean ± SEM. The data were
analyzed by the Student’s t-test between two groups or one-way
ANOVA between multiple groups. P<0.05 was considered to indicate
a statistically significant difference.
Results
BK enhances the invasion and migration of
colorectal cancer cells
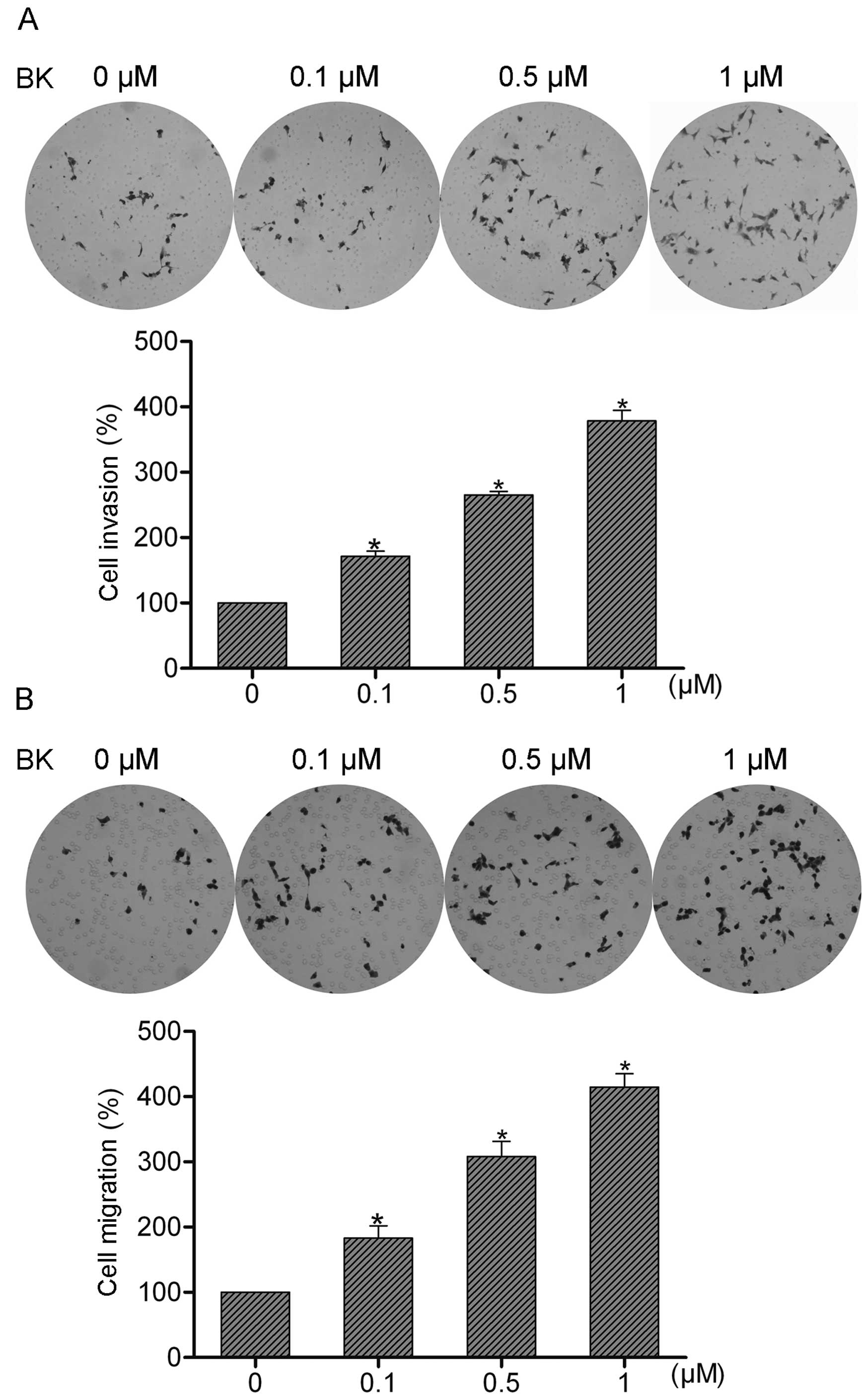
To examine the role of BK in colorectal cancer cell
invasion and migration, SW480 cells were treated with 0.1, 0.5 or 1
μM BK, respectively. Then the treated or untreated cells
were subjected to invasion and migration assays. The invaded or
migrated cells were observed and quantified 16 h later. The data
showed that BK treatment resulted in a concentration-dependent
increase in the invasive and migratory abilities of the SW480 cells
(Fig. 1A and B), indicating that BK
may be an important modulator of colorectal cancer cell invasion
and migration.
Expression of bradykinin receptors in
colorectal cancer cells and suppression of bradykinin receptors by
siRNA
It is known that BK acts via bradykinin receptors,
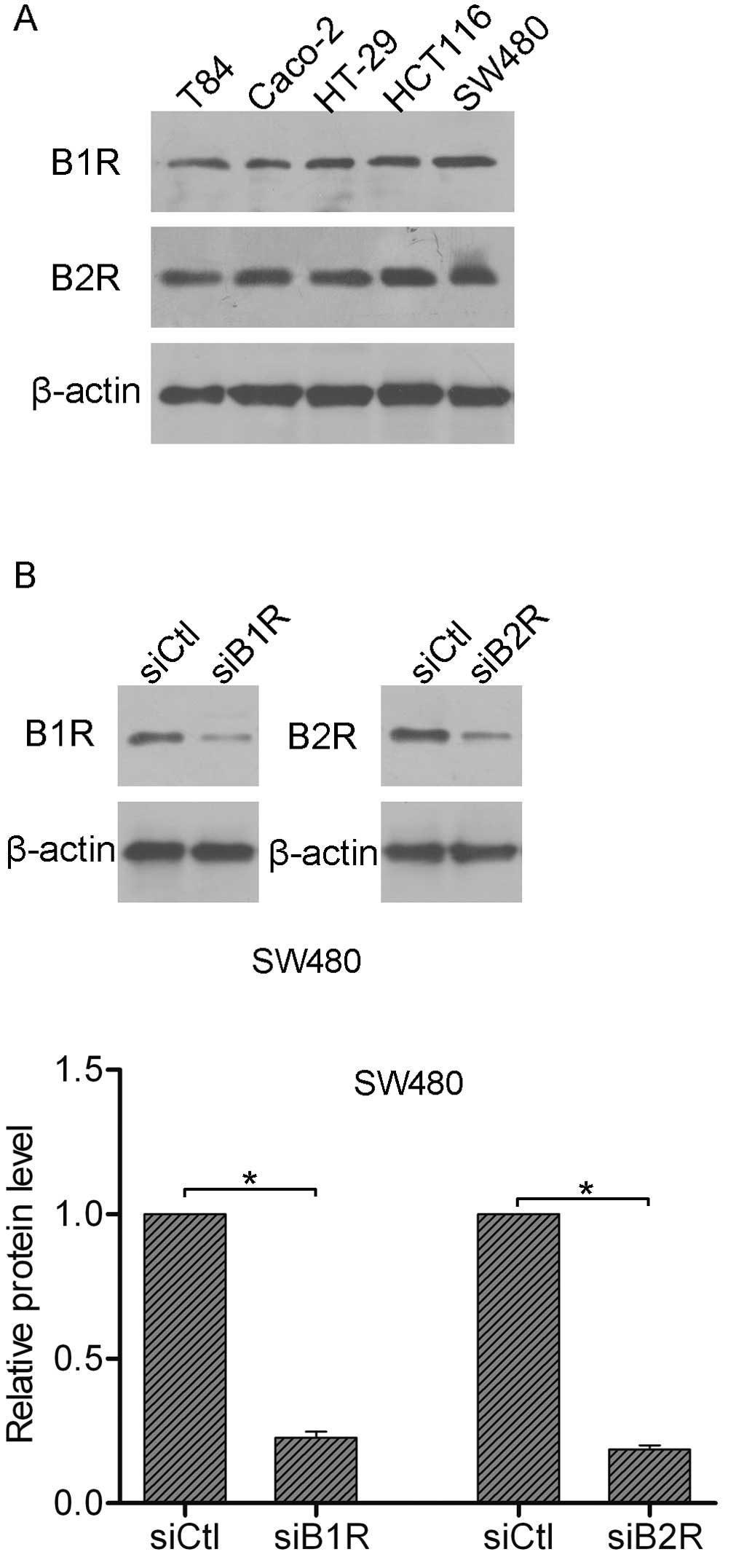
which are further characterized as B1R and B2R (3). Here, we detected the expression of B1R
and B2R in colorectal cancer cells. As shown in Fig. 2A, all tested cell lines expressed
significant amounts of B1R and B2R protein. Next, a B1R siRNA was
generated to silence the expression of B1R, while a B2R siRNA was
generated to silence the expression of B2R. As observed by western
blot analysis, both B1R siRNA and B2R siRNA achieved prominent
knockdown efficiency in the SW480 cells (Fig. 2B).
B2R suppression leads to inhibition of
BK-mediated invasion and migration in colorectal cancer cells
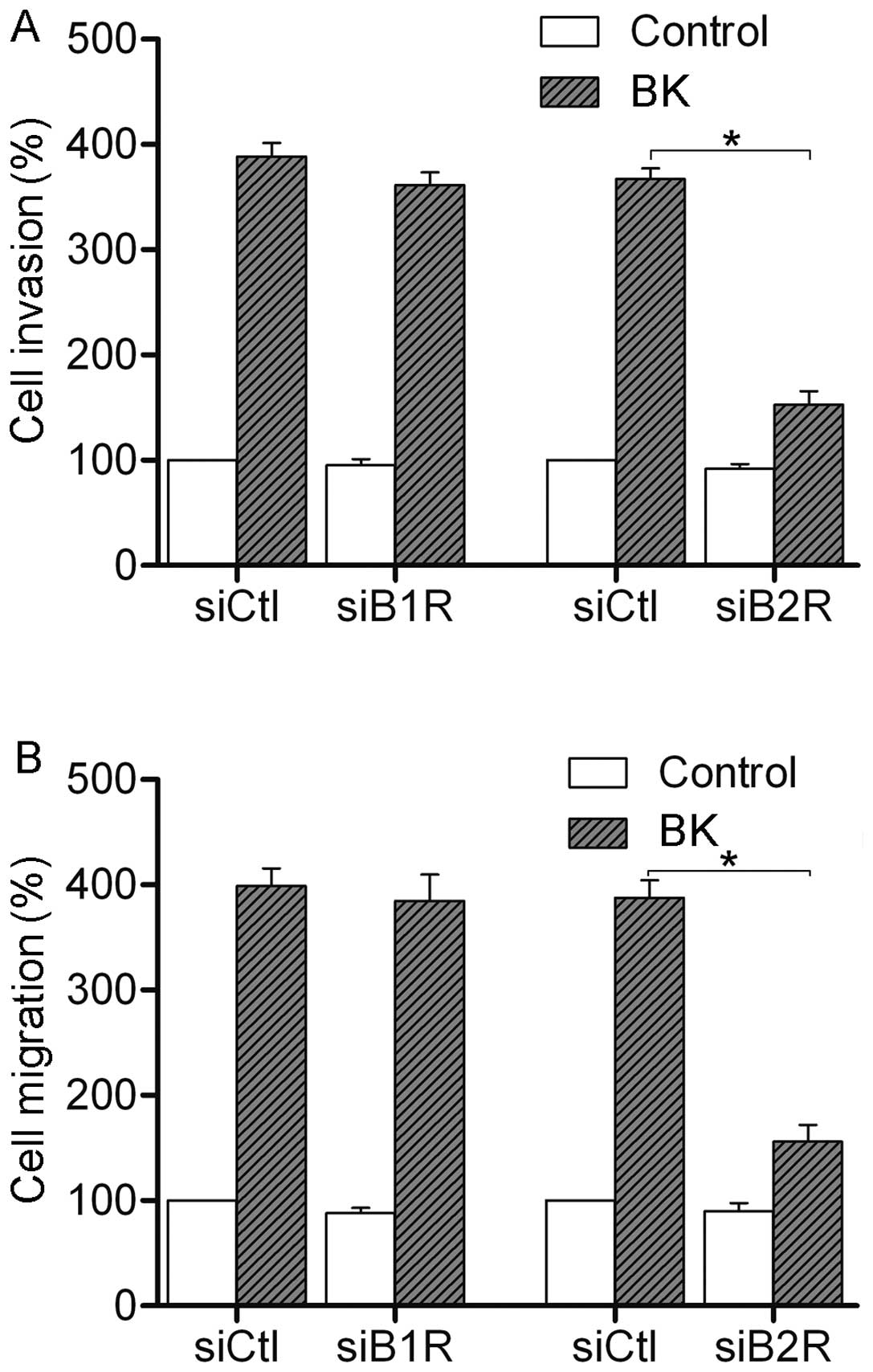
We further tested the effect of RNAi-mediated B1R
and B2R suppression on the invasion and migration of colorectal
cancer cells. Invasion and migration assays were performed after BK
treatment. As shown in Fig. 3, 1
μM BK treatment promoted the invasion and migration in the
control siRNA cells. Suppression of B1R had little effect on the
BK-mediated invasion and migration. However, suppression of B2R
resulted in a reduction in BK-mediated invasion and migration in
SW480 cells. Together, these data suggest that it is B2R that
participates in the BK-mediated invasion and migration of
colorectal cancer cells.
B2R is required for BK-mediated ERK1/2
activation
Previous studies have shown that BK can induce
multiple intracellular signaling pathways of cancer cells including
the ERK pathway (11). However, the
relationship between BK and the ERK pathway in colorectal cancer
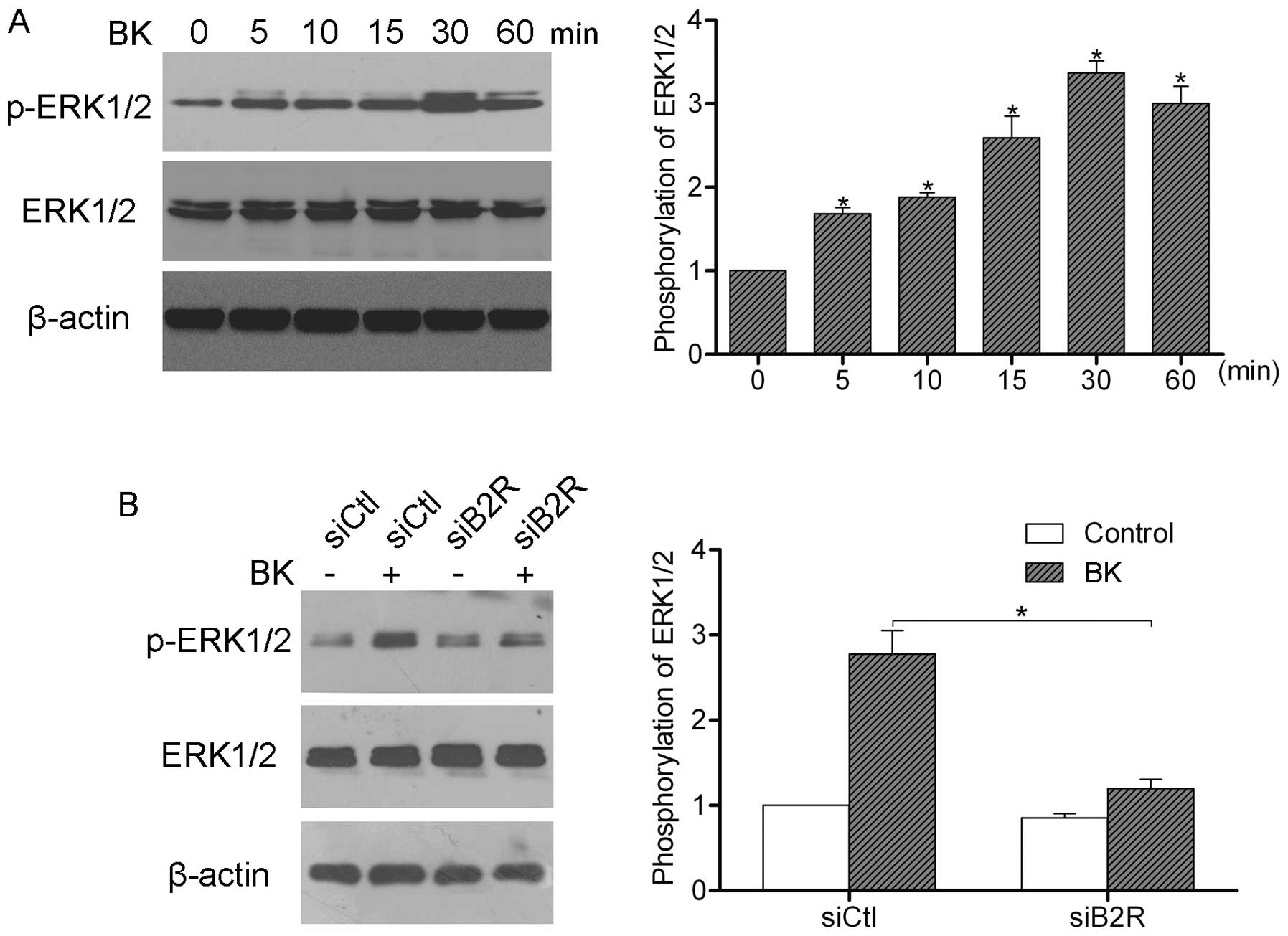
cells is, to our knowledge, not yet clear. We, therefore,
stimulated SW480 cells with 1 μM BK for 60 min, and then
examined the phosphorylation of ERK1/2 by western blot analysis.
The results showed that BK treatment induced the activation of
ERK1/2, and the peak activation was observed after 30 min (Fig. 4A). Furthermore, we silenced B2R
expression by siRNA and stimulated cells with 1 μM BK for 30
min. Western blot analysis showed that suppression of B2R inhibited
the BK-mediated activation of ERK1/2, suggesting that BK induces
ERK1/2 activation via B2R (Fig.
4B).
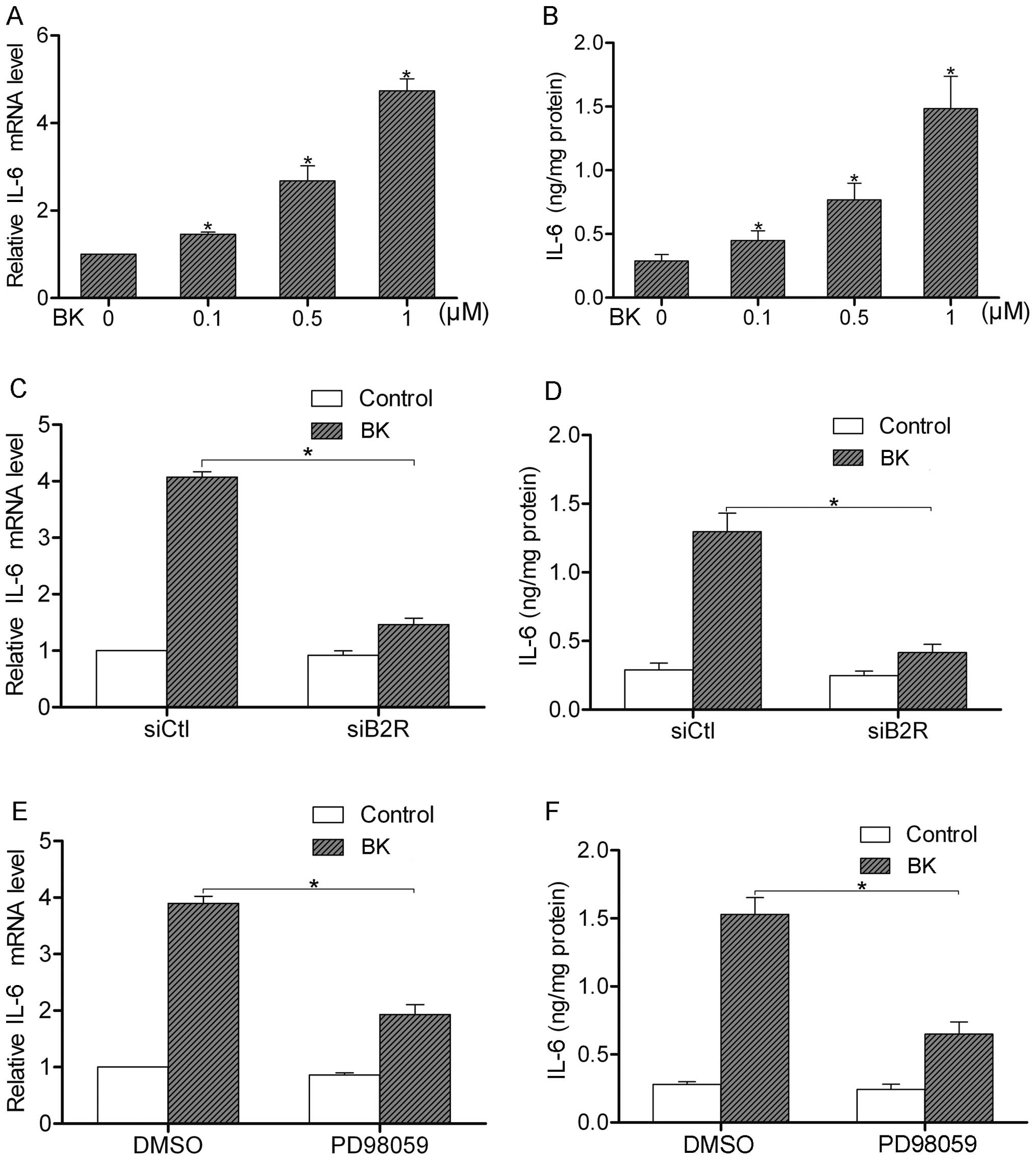
BK stimulates IL-6 production through the
B2R-ERK pathway
BK treatment is able to induce the secretion of
various cytokines such as IL-1β, IL-8 and IL-12 (12). Using real-time PCR and ELISA assay,
we found that BK treatment (0.1–1 μM) increased the
expression and secretion of IL-6 in the SW480 cells (Fig. 5A and B). We, then, silenced the
expression of B2R by siRNA to analyze the effect of B2R on the
BK-induced IL-6 production. We found that BK upregulated the
expression and secretion of IL-6 in the control cells. However,
after suppression of B2R by siRNA, the BK-mediated IL-6 production
was greatly decreased (Fig. 5C and
D). Furthermore, we pretreated SW480 cells with PD98059, a
specific inhibitor of ERK1/2, to observe the function of the ERK
pathway in the regulation of IL-6 production. The results showed
that inactivation of ERK1/2 attenuated the BK-induced IL-6
production (Fig. 5E and F). Taken
together, these data indicate that the B2R-ERK pathway is necessary
for BK-induced IL-6 production.
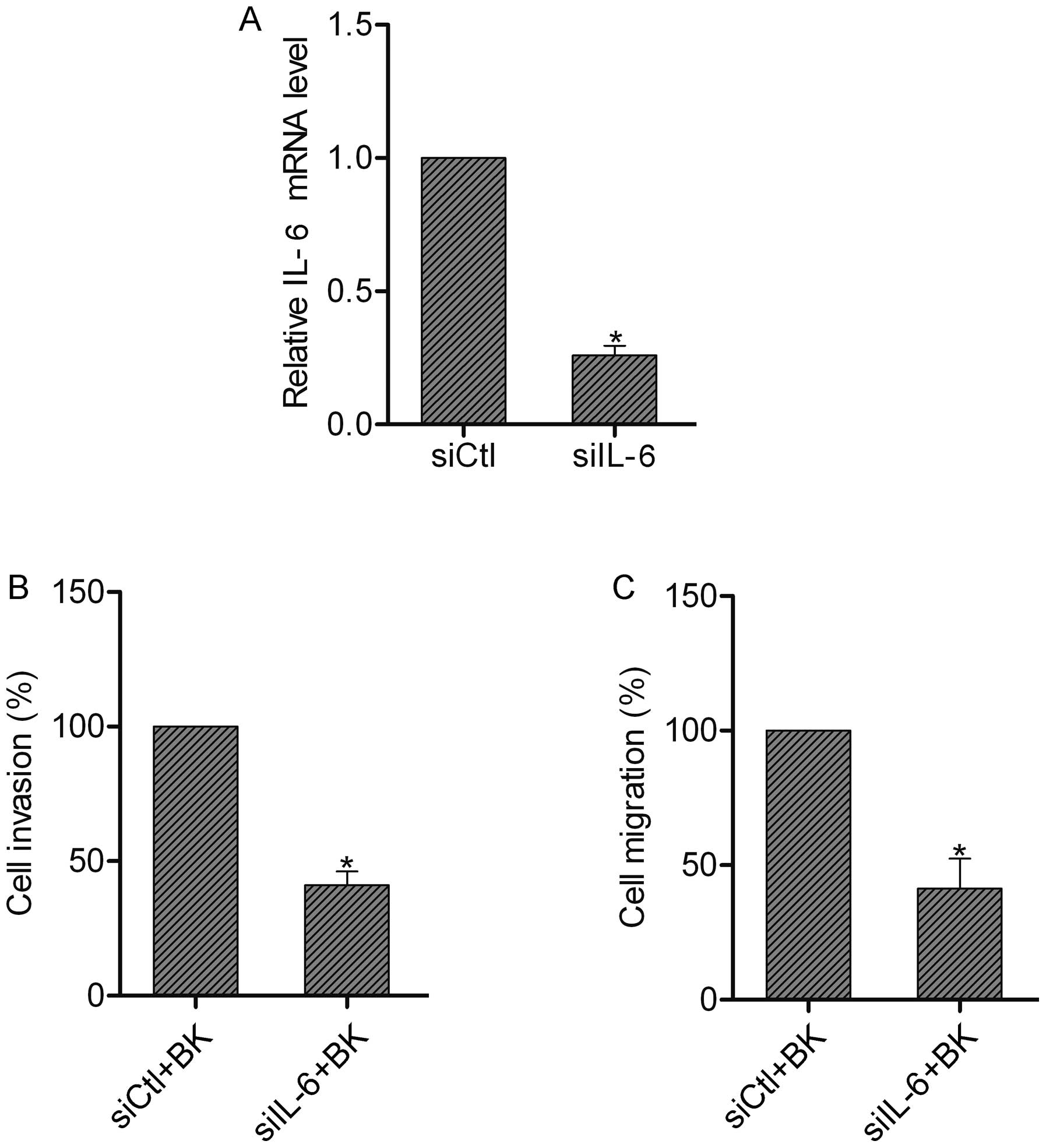
IL-6 contributes to the BK-mediated
invasion and migration of colorectal cancer cells
To reveal the role of IL-6 in invasion and
migration, we silenced the expression of IL-6 in SW480 cells by
siRNA (Fig. 6A). Then we subjected
these cells to invasion and migration assays following BK
treatment. We found that IL-6 suppression resulted in the
inhibition of invasion and migration in the SW480 cells pretreated
with BK, suggesting the involvement of IL-6 in the BK-mediated
invasion and migration of colorectal cancer cells (Fig. 6B and C).
Discussion
As the most prominent member of the kinin group,
bradykinin (BK) is an important player in the regulation of
inflammation and cancer (13,14).
Accumulating research shows that BK treatment can induce the
activation of multiple intracellular pathways such as MAPK and
PI3K/AKT signaling pathways in cancer cells (5,15), and
thus contributes to the proliferation, angiogenesis and migration
of cancer cells (16–18). Our present study demonstrated for
the first time that BK treatment activated B2R and ERK1/2, which
then led to an increase in IL-6 production, which finally promoted
cell invasion and migration of colorectal cancer cells. These data
strongly support the conclusion that BK is a potent stimulator of
colorectal cancer cell invasion and migration.
BK exerts its effects via bradykinin receptors,
which are categorized as B1R and B2R. B1R and B2R are both
implicated in the progression of cancer. Studies have found that
B1R and B2R are highly expressed in the cytoplasm of lung cancer,
and B2R is overexpressed in head and neck squamous cell carcinomas
(19,20). Using western blot analysis, we
observed significant expression levels of B1R and B2R in all of the
examined colorectal cancer cells, indicating a role of bradykinin
receptors in colorectal cancer. It was also found that BK has a
higher affinity to B2R as compared to B1R (12). Activation of B2R by BK promotes the
growth and migration of prostate cancer cells (21,22).
In the present study, we found that B2R suppression attenuated the
BK-mediated invasion and migration of SW480 cells, whereas B1R
suppression did not have an impact on the effect of BK on
colorectal cancer cells. Thus, it appears that B2R, but not B1R,
participates in regulating the BK-enhanced invasion and migration
of colorectal cancer cells.
Numerous signaling pathways have been reported to be
activated by BK, notably the MAPK, PKC and NF-κB pathways (11,23,24).
Our results showed that BK treatment induced the activation of
ERK1/2 through B2R. These findings suggest that the ERK pathway may
contribute to the BK-mediated invasion and migration of colorectal
cancer cells, as it is well known that the ERK pathway plays a
pivotal role in colorectal cancer dissemination (25).
Of note, BK treatment is able to stimulate the
generation of various cytokines such as IL-1 and tumor necrosis
factor (TNF) (26). Studies have
found that BK induces IL-6 secretion in lung fibroblasts (27). However, little is known concerning
the effect of BK on IL-6 production in cancer cells. Here, we found
that BK induced the expression and secretion of IL-6 via the
B2R-ERK pathway. To the best of our knowledge, our findings are the
first to show that BK treatment increases IL-6 production in
colorectal cancer cells. IL-6 expression is frequently upregulated
in various types of human cancers (28–30).
Clinical data have shown that IL-6 expression in tumor tissues
correlates with lymph node metastasis of colorectal cancer
(31), whereas experimental studies
confirm that IL-6 promotes cell proliferation, invasion and
metastasis of cancers (32–34). We found that suppression of IL-6 in
SW480 cells decreased the BK-mediated invasion and migration,
confirming that IL-6 plays an important role in colorectal
cancer.
In conclusion, our study demonstrated that BK
treatment enhanced the invasion and migration of colorectal cancer
cells. Suppression of B2R attenuated the BK-mediated invasion and
migration. The possible mechanism involved BK-stimulated B2R,
subsequently leading to the activation of the ERK pathway and
upregulation of IL-6 production. Therefore, it is likely that B2R
may be a potential therapeutic target for the treatment of
colorectal cancer.
Acknowledgements
This research was supported by grants from the
National Natural Science Foundation of China (81201955).
References
|
1
|
Moreau ME, Garbacki N, Molinaro G, Brown
NJ, Marceau F and Adam A: The kallikrein-kinin system: current and
future pharmacological targets. J Pharmacol Sci. 99:6–38. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Maeda H, Wu J, Okamoto T, Maruo K and
Akaike T: Kallikrein-kinin in infection and cancer.
Immunopharmacology. 43:115–128. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Regoli D: Pharmacology of bradykinin and
related kinins. Adv Exp Med Biol. 156:569–584. 1983.PubMed/NCBI
|
|
4
|
Marceau F and Regoli D: Bradykinin
receptor ligands: therapeutic perspectives. Nat Rev Drug Discov.
3:845–852. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lu DY, Leung YM, Huang SM and Wong KL:
Bradykinin-induced cell migration and COX-2 production mediated by
the bradykinin B1 receptor in glioma cells. J Cell Biochem.
110:141–150. 2010.PubMed/NCBI
|
|
6
|
Yu HS, Wang SW, Chang AC, et al:
Bradykinin promotes vascular endothelial growth factor expression
and increases angiogenesis in human prostate cancer cells. Biochem
Pharmacol. 87:243–253. 2014. View Article : Google Scholar
|
|
7
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar
|
|
8
|
Glimelius B and Cavalli-Bjorkman N:
Metastatic colorectal cancer: current treatment and future options
for improved survival. Medical approach - present status. Scand J
Gastroenterol. 47:296–314. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dashwood RH: Early detection and
prevention of colorectal cancer (Review). Oncol Rep. 6:277–281.
1999.PubMed/NCBI
|
|
10
|
Yoo J, Rodriguez Perez CE, Nie W,
Sinnett-Smith J and Rozengurt E: Protein kinase D1 mediates
synergistic MMP-3 expression induced by TNF-α and bradykinin in
human colonic myofibroblasts. Biochem Biophys Res Commun.
413:30–35. 2011.PubMed/NCBI
|
|
11
|
Hsieh HL, Wu CY and Yang CM: Bradykinin
induces matrix metalloproteinase-9 expression and cell migration
through a PKC-delta-dependent ERK/Elk-1 pathway in astrocytes.
Glia. 56:619–632. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
da Costa PL, Sirois P, Tannock IF and
Chammas R: The role of kinin receptors in cancer and therapeutic
opportunities. Cancer Lett. 345:27–38. 2014.PubMed/NCBI
|
|
13
|
Colman RW: Regulation of angiogenesis by
the kallikrein-kinin system. Curr Pharm Des. 12:2599–2607. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kashuba E, Bailey J, Allsup D and Cawkwell
L: The kinin-kallikrein system: physiological roles,
pathophysiology and its relationship to cancer biomarkers.
Biomarkers. 18:279–296. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Greco S, Elia MG, Muscella A, Romano S,
Storelli C and Marsigliante S: Bradykinin stimulates cell
proliferation through an extracellular-regulated kinase 1 and
2-dependent mechanism in breast cancer cells in primary culture. J
Endocrinol. 186:291–301. 2005. View Article : Google Scholar
|
|
16
|
Searovic P, Alonso M, Oses C,
Pereira-Flores K, Velarde V and Saez CG: Effect of tamoxifen and
retinoic acid on bradykinin induced proliferation in MCF-7 cells. J
Cell Biochem. 106:473–481. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ikeda Y, Hayashi I, Kamoshita E, et al:
Host stromal bradykinin B2 receptor signaling facilitates
tumor-associated angiogenesis and tumor growth. Cancer Res.
64:5178–5185. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang WH, Chang JT, Hsu SF, et al:
Bradykinin enhances cell migration in human chondrosarcoma cells
through BK receptor signaling pathways. J Cell Biochem. 109:82–92.
2010.PubMed/NCBI
|
|
19
|
Chee J, Naran A, Misso NL, Thompson PJ and
Bhoola KD: Expression of tissue and plasma kallikreins and kinin B1
and B2 receptors in lung cancer. Biol Chem. 389:1225–1233. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang W, Bhola N, Kalyankrishna S, et al:
Kinin b2 receptor mediates induction of cyclooxygenase-2 and is
overexpressed in head and neck squamous cell carcinomas. Mol Cancer
Res. 6:1946–1956. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yu HS, Lin TH and Tang CH: Bradykinin
enhances cell migration in human prostate cancer cells through B2
receptor/PKCδ/c-Src dependent signaling pathway. Prostate.
73:89–100. 2013.PubMed/NCBI
|
|
22
|
Srinivasan D, Kosaka AH, Daniels DV, Ford
AP and Bhattacharya A: Pharmacological and functional
characterization of bradykinin B2 receptor in human prostate. Eur J
Pharmacol. 504:155–167. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sabatini F, Luppi F, Petecchia L, et al:
Bradykinin-induced asthmatic fibroblast/myofibroblast activities
via bradykinin B2 receptor and different MAPK pathways. Eur J
Pharmacol. 710:100–109. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lee CH, Shieh DC, Tzeng CY, et al:
Bradykinin-induced IL-6 expression through bradykinin B2 receptor,
phospholipase C, protein kinase Cdelta and NF-kappaB pathway in
human synovial fibroblasts. Mol Immunol. 45:3693–3702. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sosa MS, Avivar-Valderas A, Bragado P, Wen
HC and Aguirre-Ghiso JA: ERK1/2 and p38α/β signaling in tumor cell
quiescence: opportunities to control dormant residual disease. Clin
Cancer Res. 17:5850–5857. 2011.
|
|
26
|
Tiffany CW and Burch RM: Bradykinin
stimulates tumor necrosis factor and interleukin-1 release from
macrophages. FEBS Lett. 247:189–192. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hayashi R, Yamashita N, Matsui S, et al:
Bradykinin stimulates IL-6 and IL-8 production by human lung
fibroblasts through ERK- and p38 MAPK-dependent mechanisms. Eur
Respir J. 16:452–458. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Knüpfer H and Preiss R: Serum
interleukin-6 levels in colorectal cancer patients - a summary of
published results. Int J Colorectal Dis. 25:135–140.
2010.PubMed/NCBI
|
|
29
|
Dethlefsen C, Højfeldt G and Hojman P: The
role of intratumoral and systemic IL-6 in breast cancer. Breast
Cancer Res Treat. 138:657–664. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
De Vita F, Orditura M, Auriemma A,
Infusino S, Roscigno A and Catalano G: Serum levels of
interleukin-6 as a prognostic factor in advanced non-small cell
lung cancer. Oncol Rep. 5:649–652. 1998.
|
|
31
|
Ashizawa T, Okada R, Suzuki Y, et al:
Study of interleukin-6 in the spread of colorectal cancer: the
diagnostic significance of IL-6. Acta Med Okayama. 60:325–330.
2006.PubMed/NCBI
|
|
32
|
Li R, Li G, Deng L, et al: IL-6 augments
the invasiveness of U87MG human glioblastoma multiforme cells via
up-regulation of MMP-2 and fascin-1. Oncol Rep. 23:1553–1559.
2010.PubMed/NCBI
|
|
33
|
Zheng T, Hong X, Wang J, et al: Gankyrin
promotes tumor growth and metastasis through activation of
IL-6/STAT3 signaling in human cholangiocarcinoma. Hepatology.
59:935–946. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sun W, Liu DB, Li WW, et al: Interleukin-6
promotes the migration and invasion of nasopharyngeal carcinoma
cell lines and upregulates the expression of MMP-2 and MMP-9. Int J
Oncol. 44:1551–1560. 2014.PubMed/NCBI
|