Introduction
Head and neck cancer refers to a group of
biologically similar cancers that start in the lip, oral cavity,
nasal cavity, paranasal sinuses, pharynx and larynx. Ninety percent
of head and neck cancers are squamous cell carcinomas, originating
from the epithelium of these regions. Head and neck squamous cell
carcinomas (HNSCCs) represent approximately 2.4% of all cancers
diagnosed and were implicated in 7,600 deaths in 2009 (1). Although early-stage head and neck
cancers, particularly laryngeal and oral cavity cancers, have high
cure rates, up to 50% of head and neck cancer patients present with
advanced disease (2). The patients
with locally advanced disease have a 5-year survival rate of
approximately 50%, and 10% of patients who suffer from distant
metastatic disease have a 5-year survival rate of approximately 25%
(3). Despite the advancement
achieved in surgical methods and radiation therapy, the prognosis
of head and neck cancers has not shown a satisfactory improvement
(1–6). Therefore, in order to achieve more
effective treatment of patients with HNSCC, particularly patients
with advanced disease, new treatment modalities such as
molecular-targeted therapy are clearly needed.
The family of inhibitor of apoptosis protein (IAP)
consists of a group of structurally related proteins with
anti-apoptotic properties (7,8).
Currently, eight human IAP members have been identified, including
cIAP1, cIAP2, NAIP, Survivin, XIAP, BRUCE, ILP-2 and Livin
(9–14). Livin is a novel member of the IAP
family. In contrast to other members of the IAP family, such as
cIAP1, cIAP2, XIAP and NIAP, it is expressed at low levels in
normal adult tissues, but is over-expressed in a wide variety of
human malignancies, such as colon cancer, leukemia, bladder cancer,
neuroblastoma. malignant pleural methothelioma, pancreatic cancer,
osteosarcoma, renal cell carcinoma, gastric cancer, breast
carcinoma and lung cancer (15–29).
It has been reported that Livin may be involved in the progression
of tumors, and high levels of this molecule are correlated with
tumor progression and patient outcome (22,30).
However, the impact and expression of Livin in the progression of
HNSCC are still unknown, and this issue has not been reported in
any studies, except Livin in nasopharyngeal carcinoma (32).
In order to analyze the pathobiological significance
of Livin in human oral squamous cell carcinomas (OSCCs), we
examined expression of Livin and investigated the potential
relevance between its expression and various clinicopathological
variables in a well-defined series of OSCCs. We also determine
whether Livin affects tumor cell invasion, migration, proliferation
and apoptosis in OSCC cell lines. This study demonstrated Livin
expression and its association with tumor progression in OSCC and
may provide the basis for applying Livin in the molecular-targeted
therapy for OSCCs.
Materials and methods
Patients and tumor specimens
To evaluate Livin protein expression,
paraffin-embedded tissue sections were collected from 18 patients
who underwent definitive surgery for OSCC at Chonnam National
University Hwasun Hospital (Jeonnam, Korea) between March 2006 and
August 2009. All 18 patients underwent appropriate definitive
surgery. In 6 patients, surgery was followed by radiation therapy
alone or cisplatin-based concurrent chemoradiation therapy.
Patient characteristics including age, gender, tumor
location, stage, depth of tumor invasion, lymph node metastasis,
distant metastasis, treatment, recurrence, survival and follow-up
information were obtained by review of the hospital records. All
patients were staged according to the 7th edition of the American
Joint Committee on Cancer (AJCC) staging system (33). Overall survival was calculated from
the first surgical date to the date of death or the date of last
follow-up.
Immunohistochemistry
Livin protein expression was investigated
immunohistochemically in formalin-fixed, paraffin-embedded tissue
blocks obtained from 18 OSCC patients. Tissue sections (5 μm) were
cut from each paraffin block, mounted, and dried on glass slides.
Tissues were deparaffinized, rehydrated, and retrieved with
retrieval buffer. Endogenous peroxidase activity was blocked with
peroxidase-blocking solution (Dako, Carpinteria, CA, USA), followed
by incubation with polyclonal rabbit anti-human Livin (Santa Cruz
Biotechnology, Santa Cruz, CA, USA) overnight at 4°C. After washing
in Tris-buffered saline-Tween-20 buffer (TBST), tissues were
stained using the Dako Real™ Envision horseradish peroxidase
(HRP)/3,3′-diaminobenzidine (DAB) detection system (Dako). Tissues
were counterstained with hematoxylin and mounted with coverslips.
Stained tissues were viewed and photographed using a light
microscope. Assessment of staining was interpreted by two
independent observers who were without knowledge regarding the
clinical information. The percentage of positive immunostaining
cells was graded according to three levels: 0, <5%; 1, 5–50%;
and 2, ≥50%.
Cell culture and transfection
Cells of the human OSCC cell line, PCI 50, were
cultured in RPMI-1640 (Invitrogen, Carlsbad, CA, USA) supplemented
with 10% fetal bovine serum (Hyclone, Logan, UT, USA), 50 U/ml
penicillin, and 50 μg/ml streptomycin (Gibco, Grand Island, NY,
USA) in a humidified atmosphere of 5% CO2 at 37°C. The
PCI 50 cell line originated from oral cavity cancer. For
transfection, cells were prepared and maintained in culture dishes
with medium, and cells were seeded on 6-well plates at
2×105 cells per well at the time of transfection. To
study the biological role of Livin in OSCC, small interfering RNA
(siRNA) was used to knock down endogenous Livin gene expression in
the PCI 50 cells. Cells were transfected with Livin-specific siRNA
(Bioneer, Daejeon, Korea) and negative control siRNA (Qiagen,
Valencia, CA, USA) using Lipofectamine™ 2000 (Invitrogen). Forty
eight hours after transfection, the viable transfected cells were
enumerated and re-seeded in the individual experimental
apparatus.
Cell invasion assay
Cell invasion was measured using a Transwell
invasion apparatus (Costar, Cambridge, UK). Transwell filters
(8.0-μm pore size) were coated overnight with 1% gelatin solution
on both the top and bottom surfaces and dried at room temperature.
Cells transfected with Livin siRNA or negative control siRNA were
seeded at 3×105 cells in 120 μl of a 0.2% bovine serum
albumin (BSA) suspension in the upper chamber. Subsequently, 400 μl
of 0.2% BSA containing 7 μg/ml fibronectin (Calbiochem, La Jolla,
CA, USA) as the chemoattractant was loaded into the lower chamber.
After incubation for 24 h, cells on the top surface of the filter
were wiped off with cottonballs. Cells that had migrated to the
bottom surface of the Transwell were stained with Diff Quik
solution (Sysmex, Kobe, Japan) and counted in five random squares
in the microscopic field of view. Results are expressed as the mean
± standard error of the number of cells/field in three individual
experiments.
Cell migration assay (wound healing
assay)
Cells transfected with Livin siRNA or negative
control siRNA were seeded in each well of
Culture-Inserts® (Ibidi, Bonn, Germany) at
1.5×105 cells/well. After incubation for 24 h, each
insert was detached and the progression of cell migration was
ascertained by photography at 0, 8, 12 and 24 h using an inverted
microscope. The distance between gaps was normalized to 1 cm after
capture of three random sites.
Cell viability
Cell viability was determined with the EZ-CyTox
(tetrazolium salt, WST-1) cell viability assay kit (Daeil Lab Inc.,
Seoul, Korea). After applying the WST-1 reagent at 37°C, cell
viability was measured using a microplate reader (Infinite M200;
Tecan, Austria GmbH, Vienna, Austria) with Magellan V6 data
analysis software (Tecan). Triplicate wells were used for each
experiment, and all experiments were conducted at least in
triplicate.
Cell apoptosis assay
Apoptosis was determined by Annexin V-fluorescein
isothiocyanate (FITC) assay. Cells transfected with Livin siRNA or
negative control siRNA were collected using trypsin, washed twice
in phosphate-buffered saline (PBS) twice and re-suspended in
binding buffer (BD Biosciences®, San Diego, CA, USA).
Annexin V-FITC and 7-amino-actinomycin D (7-AAD; BD Biosciences)
were added to the cells, which were then incubated in the dark for
15 min, then added and re-suspended in 400 ml of binding buffer.
Cells were analyzed using a FACSCalibur flow cytometer (Becton
Dickinson, San Jose, CA). Data analysis was performed using
standard Cell Quest software (Becton Dickinson).
Protein isolation and western blot
analysis
Cells and tissues were lysed in RIPA buffer (1 M
Tris-HCl, 150 mM NaCl, 1% Triton X-100, 2 mM EDTA) with 1 mM
phenylmethanesulfonyl fluoride (PMSF), Halt™ phosphatase inhibitor
and Halt™ protease inhibitor cocktail (Thermo, Rockford, IL, USA).
Resolved proteins (10–20 μg) were subjected to sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), and the
resolved proteins were electrophoretically transferred to
polyvinylidene fluoride (PVDF) membranes (Millipore, Billerica, MA,
USA). After blocking with 5% BSA in TBST at room temperature for 1
h, the PVDF membranes were sequentially blotted with the primary
antibodies: polyclonal anti-human Livin, cleaved caspase-3, cleaved
caspase-7, cleaved poly-ADP ribose polymerase (PARP) and β-actin
(Santa Cruz Biotechnology, Santa Cruz, CA, USA) and polyclonal
anti-human glyceraldehyde 3-phosphate dehydrogenase (GAPDH; Santa
Cruz Biotechnology) at 4°C overnight. After rinsing in TBST, each
membrane was incubated with the anti-rabbit or anti-mouse
HRP-conjugated secondary antibody (Santa Cruz Biotechnology) at
room temperature for 1 h. Blots were detected with
chemiluminescence (ECL) HRP substrate (Millipore) and recorded by a
Ras-4000 image reader (Fujifilm, Tokyo, Japan).
RNA isolation and reverse
transcriptase-polymerase chain reaction (RT-PCR)
The RNA from cells was isolated using TRIzol reagent
(Invitrogen), reverse transcribed, and amplified using specific
primers for Livin and GAPDH. Primer sequences were: Livin
5′-CACACAGGCCATCAGGAC AAG-3′/5′-ACGGCACAAAGACGATGGAC-3′ and GAPDH
5′-ACCACAGTCCATGCCATCAC-3′/5′-TCCACCACCCTG TTGCTGTA-3′. The size of
the amplified products was 456 bp for Livin α and 403 bp for Livin
β. For cDNA synthesis, 1 μg of mRNA was mixed with 50 ng/μl
oligo-dT (Promega, Madison, WI, USA), MMLV reverse transcriptase
(Invitrogen) and RNAsin (Takara Bio, Shiga, Japan) and incubation
was continued at 42°C for 1 h and at 72°C for 15 min. PCR
amplification of cDNA was performed using specific primers and
GoTaq® DNA polymerase (Promega). PCR products were
separated by electrophoresis on a 1% agarose gel containing
ethidium bromide.
Statistical analysis
Experimental differences between the Livin knockdown
group and the control group were tested with the Student’s t-test.
The statistical software program SigmaPlot Software (version 6.0;
Systat Software, San Jose, CA, USA) was used. Survival curves were
calculated using the Kaplan-Meier method, and comparison of the
curves was performed using the log-rank test. The statistical
software program used was Statistical Package for the Social
Sciences (SPSS) version 18.0 (Microcal Software Inc., Chicago, IL,
USA) was used. A p-value <0.05 was considered to indicate a
statistically significant result.
Results
Expression of Livin is increased in the
OSCC tissues compared with normal mucosa
The patient data are summarized in Table I. The study group of patients
included 12 males and 6 females. The mean age was 59.8±10.8 years
(mean ± standard deviation), with a range from 41 to 79 years. The
mean follow-up period was 50.5 months, with a range from 11.2 to
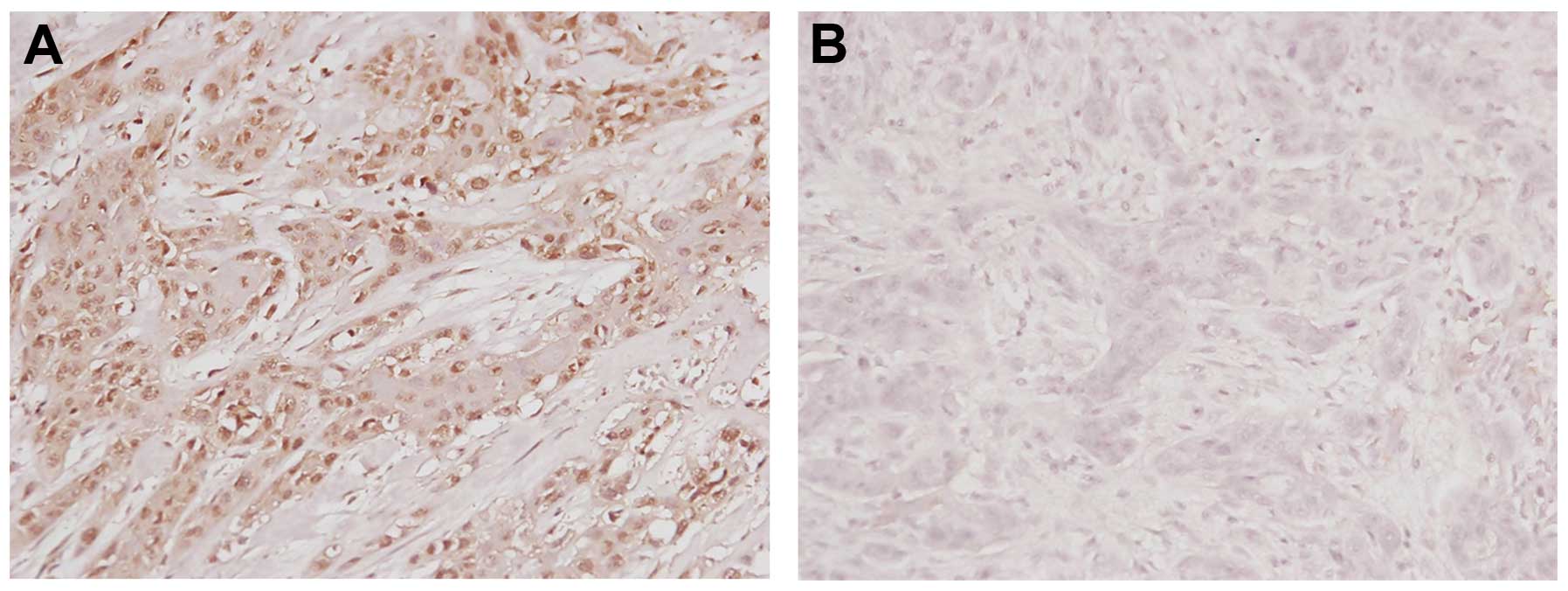
84.7 months. Immunohistochemical analysis revealed that 8 (44.4%)
of the 18 OSCC tissues overexpressed Livin relative to the adjacent
normal mucosa. A pattern of strong nuclear staining was evident
(Fig. 1A). In contrast, there was
no staining in another OSCC tissue sample as negative control
(Fig. 1B). The number of positive
tumor cells for each tissue was recorded. Three tissues (16.7%)
showed 2+ staining and five tissues (27.8%) had 1+ staining. Ten
tissues (55.6%) had negative staining.
 | Table IClinicopathological features of the
18 patients with oral squamous cell carcinoma. |
Table I
Clinicopathological features of the
18 patients with oral squamous cell carcinoma.
| Clinicopathological
features | Data |
|---|
| Age (years) |
| Mean ± SD | 59.8±10.8 |
| Range | 41–79 |
| Gender, n |
| Male | 12 |
| Female | 6 |
| Location, n |
| FOM | 4 |
| Oral tongue | 13 |
| Buccal mucosa | 1 |
| Stage, n |
| I | 9 |
| II | 4 |
| III | 3 |
| IV | 2 |
| T stage, n |
| T1 | 9 |
| T2 | 7 |
| T3 | 2 |
| N stage, n |
| N0 | 13 |
| N1 | 3 |
| N2 | 2 |
| Distant metastasis,
n |
| M0 | 17 |
| M1 | 1 |
| Recurrence, n |
| Negative | 13 |
| Positive | 5 |
| 3-year overall
survival (%) | 72 |
| Livin expression, n
(%) |
| Negative | 10 (55.6%) |
| Positive | 8 (44.4%) |
Correlations between Livin protein
expression and clinicopathological characteristics of the human
OSCC cases
Correlations between Livin protein expression and
clinicopathological characteristics in human OSCC are shown in
Table II. Livin protein expression
was not associated with age, gender, tumor location, stage, T
stage, N stage, distant metastasis and recurrence
(p>0.05) (Table II).
| Table IICorrelation between Livin expression
and the clinicopathological features of the patients with oral
squamous cell carcinoma. |
Table II
Correlation between Livin expression
and the clinicopathological features of the patients with oral
squamous cell carcinoma.
| Livin
expression | |
|---|
|
| |
|---|
| Features | Negative
(n=10) | Positive (n=8) | P-value |
|---|
| Age (years) | | | 0.19 |
| <60 | 6 | 2 | |
| ≥60 | 4 | 6 | |
| Gender | | | 0.32 |
| Male | 8 | 4 | |
| Female | 2 | 4 | |
| Location | | | 1.00 |
| FOM, buccal
mucosa | 3 | 2 | |
| Oral tongue | 7 | 6 | |
| Stage | | | 0.61 |
| I, II | 8 | 5 | |
| III, IV | 2 | 3 | |
| T stage | | | 1.00 |
| T1 | 5 | 4 | |
| T2, T3 | 5 | 4 | |
| N stage | | | 0.61 |
| N0 | 8 | 5 | |
| N1, N2 | 2 | 3 | |
| Distant
metastasis | | | 0.44 |
| M0 | 10 | 7 | |
| M1 | 0 | 1 | |
| Recurrence | | | 0.12 |
| Negative | 9 | 4 | |
| Positive | 1 | 4 | |
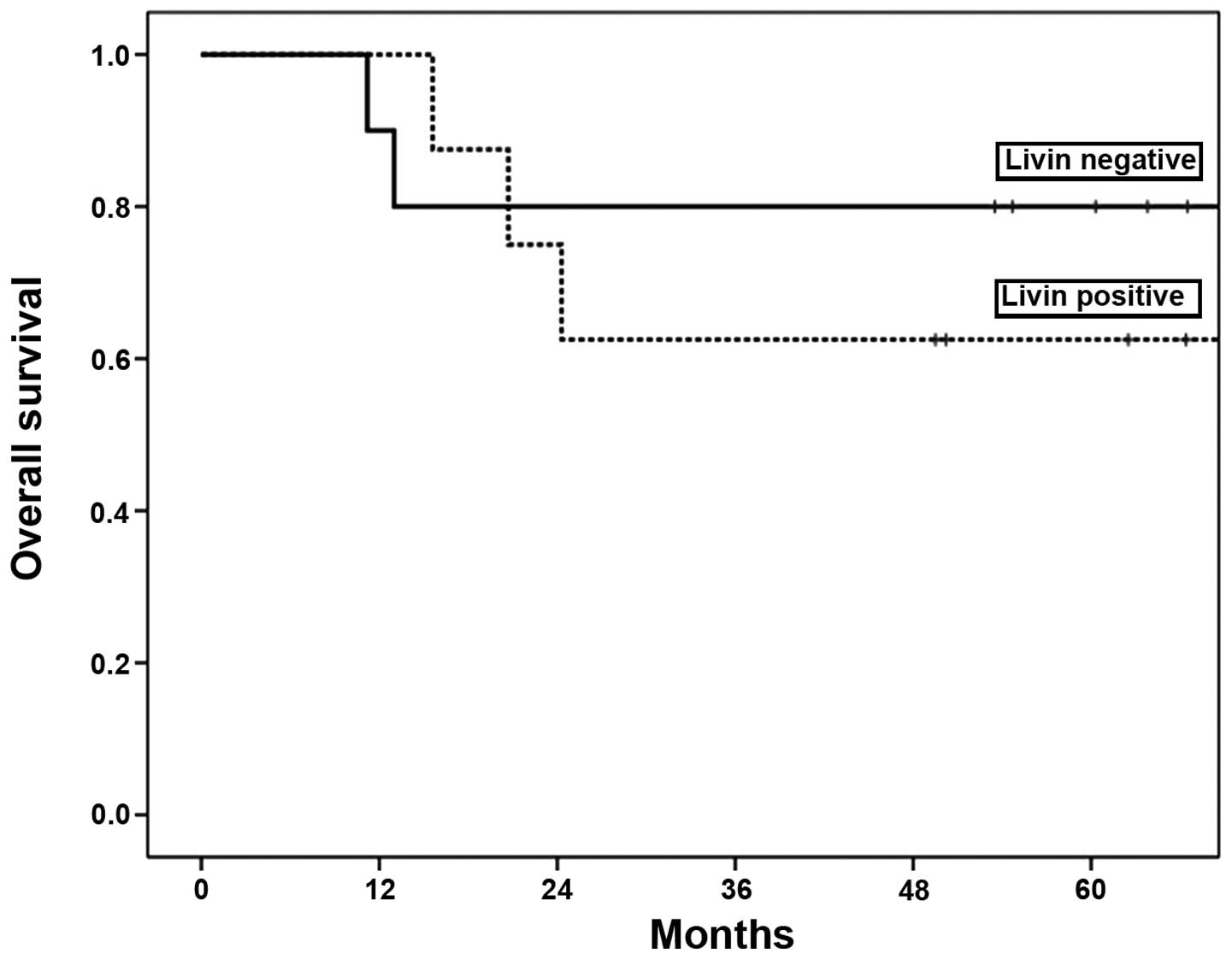
For the 18 patients with OSCC enrolled in this
study, the 3-year overall survival rate was 72%. The 3-year overall
survival was 80% in the negative Livin group, and 63% in the
positive Livin group. Livin expression was not associated with
patient overall survival (p=0.54) (Fig. 2).
Knockdown of Livin suppresses tumor cell
invasion, migration and proliferation in human OSCC cells
Expression and knockdown of Livin in a
human OSCC cell line
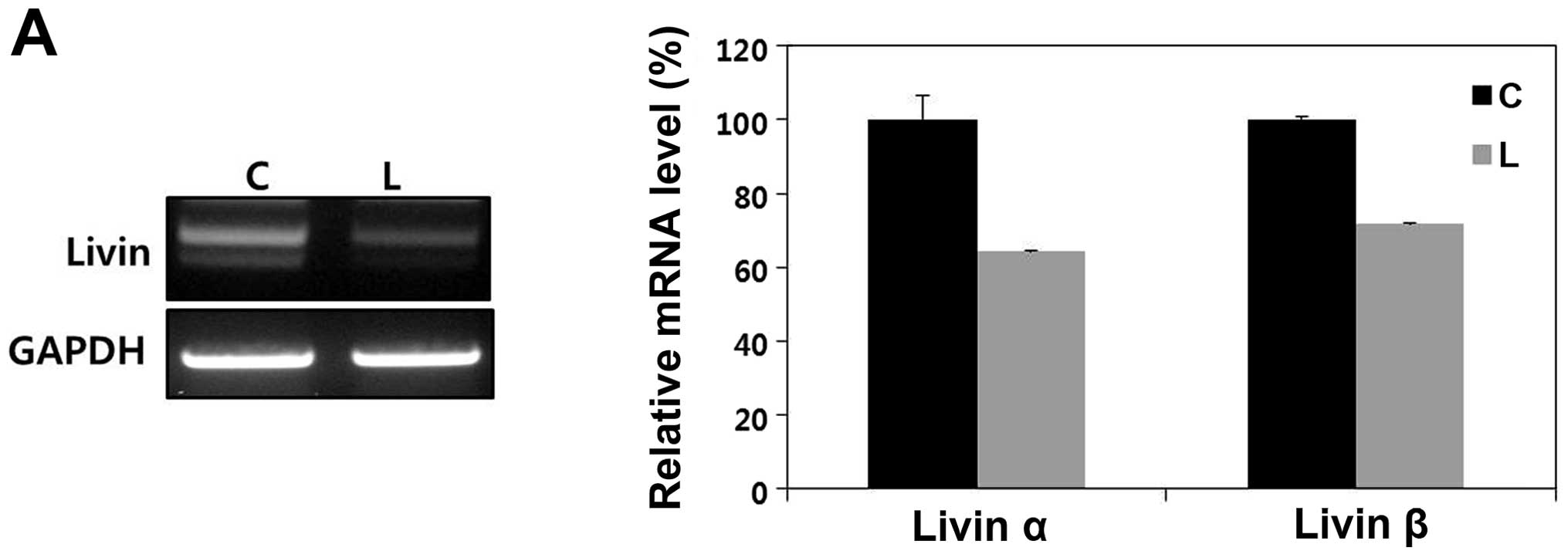
RT-PCR analysis demonstrated Livin mRNA expression
in the OSCC cell line (Fig. 3A).
Western blot analysis confirmed high Livin expression in the PCI 50
cells (Fig. 3B).
Knockdown of Livin results in
significantly reduced cell invasiveness in human OSCC cells
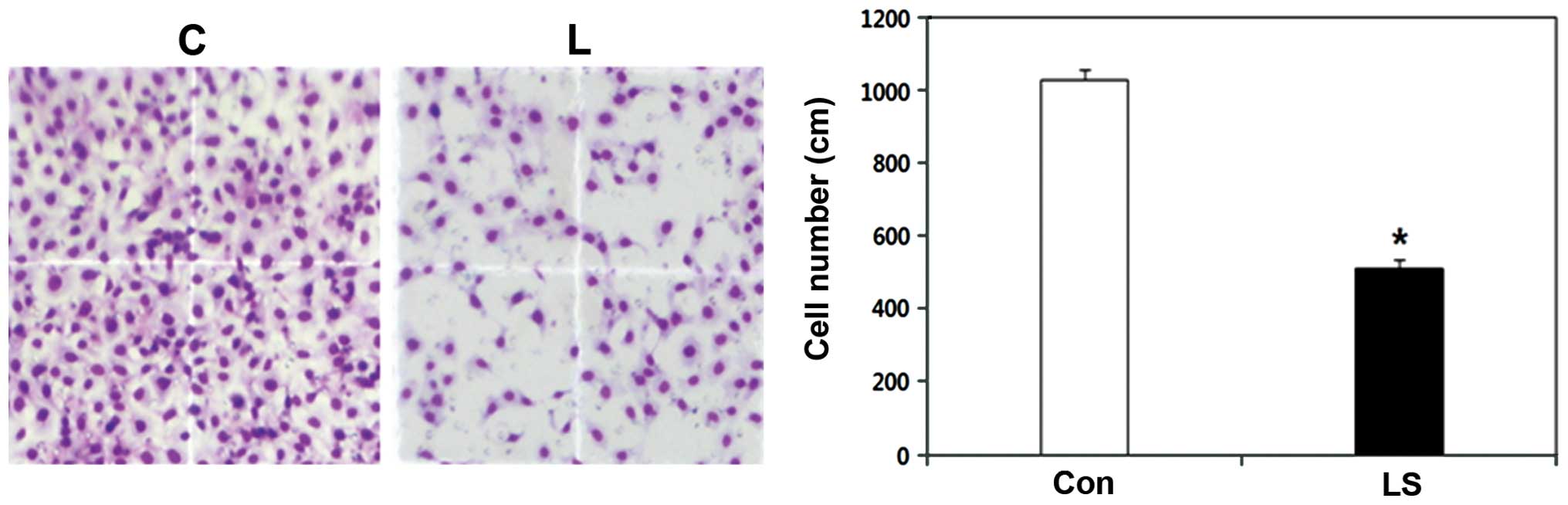
In the cell invasion assay, the number of invasive
Livin siRNA-transfected PCI 50 cells was 1029.0±28.7, whereas the
number of invasive cells in the negative control siRNA-transfected
PCI 50 cells was 511.0±24.1. The difference between the two groups
was statistically significant (p<0.01) (Fig. 4). Livin knockdown resulted in
significantly reduced cell invasiveness in the human OSCC
cells.
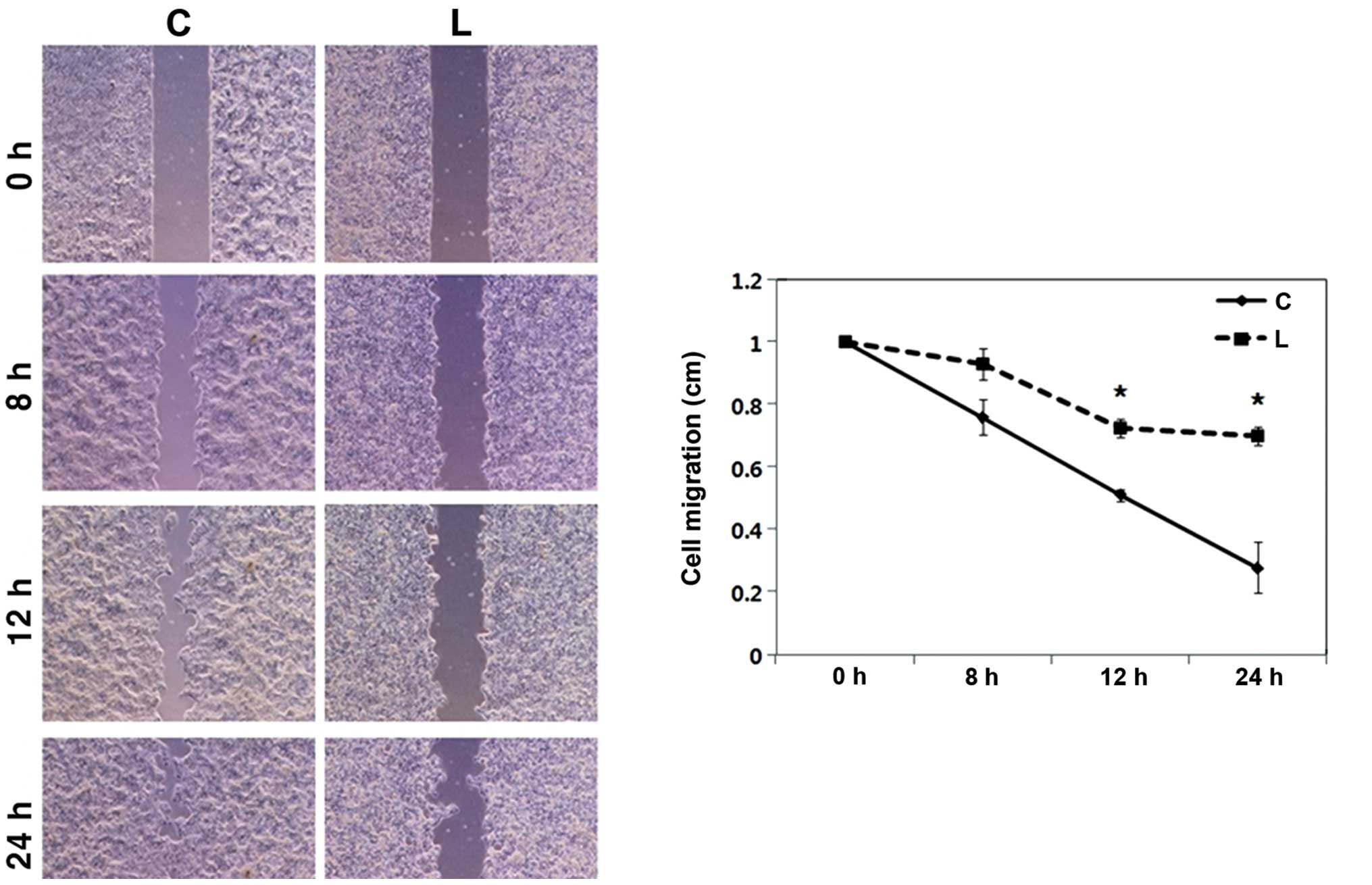
Knockdown of Livin results in
significantly diminished cell migration in human OSCC cells
In the cell migration assay, the artificial wound
gap became significantly narrower in the plates of the negative
control siRNA-transfected PCI 50 cells when compared with that of
the Livin siRNA-transfected PCI 50 cells at 12 and 24 h
(p<0.01) (Fig. 5). After
24 h, the wound gaps were filled in the plates of the negative
control siRNA-transfected OSCC cells. In contrast, the wound gaps
were wide open in the plates of Livin siRNA-transfected OSCC cells
after 24 h. The results clearly showed that Livin knockdown
resulted in significantly diminished cell migration in the human
OSCC cells.
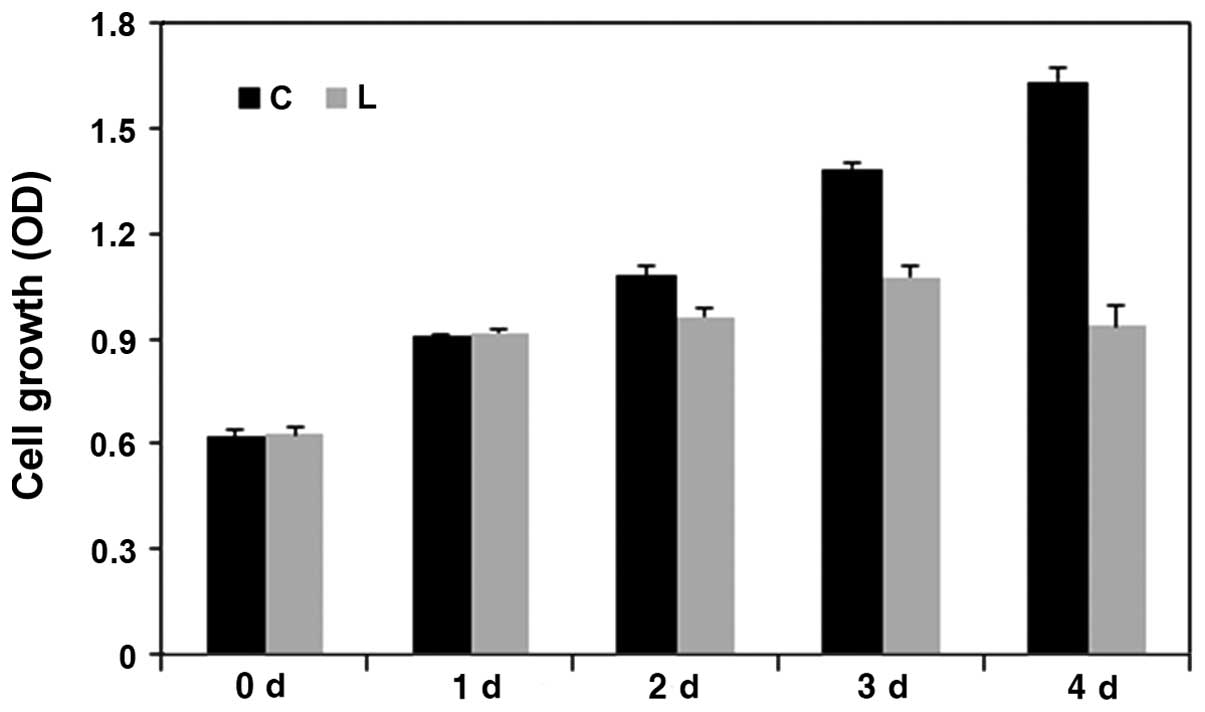
Knockdown of Livin results in
significantly decreased cell proliferation in human OSCC cells
The cell viability was assessed at 1, 2, 3 and 4
days in the negative control siRNA-transfected PCI 50 cells and
Livin siRNA-transfected PCI 50 cells to assessed the effect of
Livin on cell proliferation (Fig.
6). The number of proliferating cells was significantly
decreased in the Livin siRNA-transfected PCI 50 cells when compared
to this number in the negative control siRNA-transfected PCI 50
cells at 2, 3 and 4 days (p=0.03, p=0.01, and
p=0.003, respectively). The results clearly showed that
Livin knockdown resulted in significantly decreased cell
proliferation in human OSCC cells.
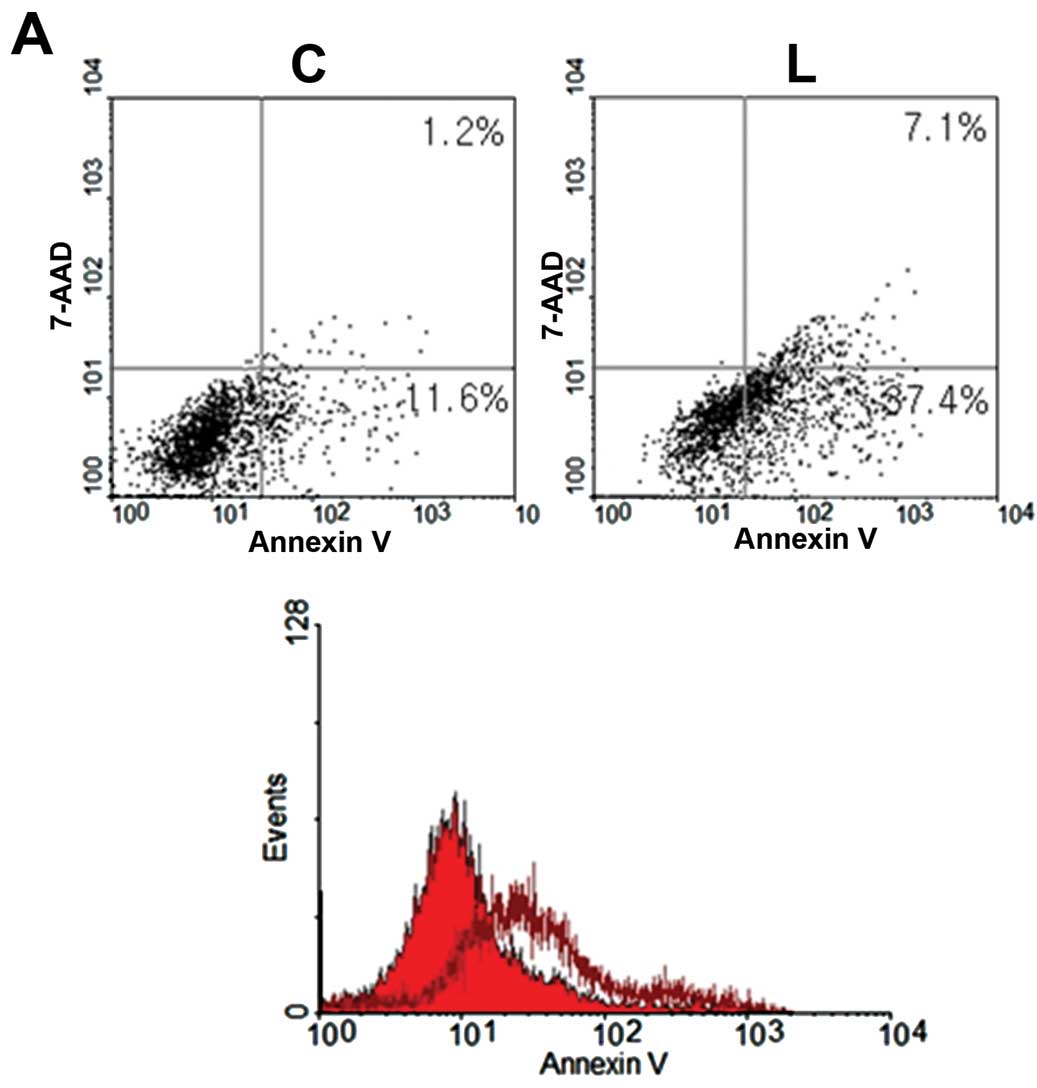
Knockdown of Livin induces cell apoptosis
in human OSCC cells
To determine the impact of Livin on cell apoptosis,
responses of Livin siRNA-transfected OSCC cells and negative
control siRNA-transfected OSCC cells were compared by an Annexin V
apoptosis assay. Livin-knockdown OSCC cells displayed increased
apoptosis, compared with the control cells (Fig. 7A). The proportions of early and late
apoptotic PCI 50 cells induced by transfection of Livin siRNA (37.4
and 7.1%, respectively) were greater than that induced following
transfection with negative control siRNA (11.6 and 1.2%,
respectively) (Fig. 7A).
Additionally, an increase in cleaved caspase-3, cleaved caspase-7
and cleaved PARP, which are key enzymes of apoptosis, was noted in
the Livin-knockdown OSCC cells (Fig.
7B). These results suggest that Livin knockdown-induced
apoptosis was associated with the modulation of apoptosis
regulatory proteins such as caspase-3, caspase-7 and PARP.
Discussion
The IAP family consists of a group of intracellular
proteins that are essential for the regulation of apoptosis
(19,34). The IAPs consist of one baculovirus
IAP repeat (BIR) domain and a COOH-terminal RING finger domain that
suppresses apoptosis induced by a variety of stimuli (29). Apoptosis plays an important role in
oncogenesis and tumor regression (35). Loss of apoptotic regulation can lead
to a variety of diseases including cancer. IAPs bind directly and
potentially inhibit a complex array of cysteine aspartyl-specific
proteases, caspase-3, caspase-7, which are responsible for
apoptosis and which are induced by diverse pro-apoptotic stimuli
(34).
Livin is one of the potent members of the IAP
family. Livin is undetectable in most normal tissues but is
upregulated in a wide variety of human cancers (15–29).
Livin expression contributes to tumor progression, a poor prognosis
and is correlated with more aggressive tumor behavior, such as
decreased response to radiotherapy and chemotherapy and reduced
survival time in many human cancers (18,22,23,36).
These findings represents a potential novel therapeutic target for
the treatment of human cancers.
However, whether Livin is related to head and neck
cancer has not been investigated. Upon review of the English
language literature, there is only one report of Livin expression
in biopsy samples of nasopharyngeal carcinoma. Xiang et al
reported that Livin was expressed in 48.75% of nasopharyngeal
carcinoma cases (32). In the
present study, Livin was expressed in 8 (44.4%) of 18 OSCC tissues.
This result is similar to that for nasopharyngeal carcinoma. In
addition, OSCC tissues exhibited overexpression of Livin relative
to the adjacent normal mucosa. This suggests that Livin may be
involved in the development of OSCC. In addition, Livin is
specifically overexpressed in cancer tissues, but almost absent in
most normal adult tissues with the exception of the placenta
(37,38). This molecule represents a possible
target for developing drugs that selectively eliminate cancer
cells.
OSCC is characterized by a marked propensity for
local invasion and lymphatic metastasis. Understanding the
molecular mechanisms that mediate SCC invasion and metastasis may
enable identification of novel therapeutic targets for management
of tumor dissemination. Livin promotes the invasion, growth and
apoptotic resistance in a variety of human cancer cells (14,39–41).
In addition, Livin expression may be essential for survival of
certain cancer cells (17,21,23,41,42).
In this study, Livin knockdown inhibited tumor cell invasion,
migration and proliferation in OSCC cells. These results indicate
participation of Livin in tumor progression and metastasis in OSCCs
and support immunohistochemical data showing an association of
Livin expression with cervical lymph node metastasis in human OSCC
tissues. In addition, Livin expression was found to be
significantly associated with cell proliferation in the human OSCC
tissues.
Livin was previously found to inhibit apoptosis by
binding to caspase-3, -7 and -9, and its E3 ubiquitin-ligase
activity promotes the degradation of IAP antagonist SMAC/DIABLO
(second mitochondrial-derived activator of caspase/direct IAP
binding protein with low pI) (14,39–41).
In the present study, Livin knockdown induced cell apoptosis in
human OSCC cells. As knockdown of Livin suppression could lead to
apoptotic cell death in human OSCC cells, suppression of Livin
should have a benefit in cancer treatment. These results suggest
that knockdown of Livin could provide a potential therapeutic
strategy to induce apoptosis in OSCC cells and to significantly
improve antitumor responses.
To determine the mechanisms of cell apoptosis in
Livin siRNA-transfected OSCC cells, the change in apoptosis-related
proteins in Livin siRNA-transfected OSCC cell lysate was evaluated.
This study showed that the expression of cleaved caspase-3, -7 and
PARP was upregulated in human OSCC cells after knockdown of Livin.
Therefore, Livin inhibits apoptosis by suppressing the activity of
caspases in human OSCC cells.
Finally, we assessed the expression of Livin and its
prognostic relevance in a well-defined series of human OSCC tissues
with complete clinicopathological data including survival. No
significant correlation was found between Livin expression and
various clinicopathological parameters including age, gender, tumor
location, stage, T stage, N stage, distant metastasis, and
recurrence. Furthermore, Livin expression was not correlated with
survival. However, the 3-year overall survival was 80% in the
negative Livin group and 63% in the positive Livin group. Although
there was no statistically significant differences between Livin
expression and survival, Livin expression had a trend toward
predicting a reduced survival rate. This result may reflect the
relatively small sample size. In addition, the steps involved in
OSCC development and progression are not dependent on Livin
expression alone but may be regulated by many biological processes
including invasion and metastasis. Further studies are required to
clarify the impact of Livin on the biologic and prognostic
significance in OSCC.
In summary, knockdown of Livin inhibited tumor cell
invasion, migration and proliferation in human OSCC cells.
Knockdown of Livin induced cell apoptosis in human OSCC cells.
Livin inhibited apoptosis by suppressing the activity of caspases
in human OSCC cells. Livin was not related with the
clinicopathologic parameters of patients with OSCC. However, Livin
expression was predictive of a poor 3-year overall survival
rate.
In conclusion, Livin is associated with invasive and
oncogenic phenotypes such as tumor cell invasion, tumor cell
migration, tumor cell proliferation, and resistance to apoptosis in
human OSCC cells.
References
|
1
|
Jemal A, Siegel R, Ward E, Hao Y, Xu J and
Thun MJ: Cancer statistics, 2009. CA Cancer J Clin. 59:225–249.
2009. View Article : Google Scholar
|
|
2
|
Gourin CG and Podolsky RH: Racial
disparities in patients with head and neck squamous cell carcinoma.
Laryngoscope. 116:1093–1106. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jemal A, Siegel R, Ward E, Hao Y, Xu J,
Murray T and Thun MJ: Cancer statistics, 2008. CA Cancer J Clin.
58:71–96. 2008. View Article : Google Scholar
|
|
4
|
Adjuvant chemotherapy for advanced head
and neck squamous carcinoma. Final report of the Head and Neck
Contracts Program. Cancer. 60:301–311. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shah JP and Lydiatt W: Treatment of cancer
of the head and neck. CA Cancer J Clin. 45:352–368. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lavertu P, Adelstein DJ, Saxton JP, Secic
M, Eliachar I, Strome M, Larto MA and Wood BG: Aggressive
concurrent chemoradiotherapy for squamous cell head and neck
cancer: an 8-year single-institution experience. Arch Otolaryngol
Head Neck Surg. 125:142–148. 1999.PubMed/NCBI
|
|
7
|
Ambrosini G, Adida C and Altieri DC: A
novel anti-apoptosis gene, survivin, expressed in cancer and
lymphoma. Nat Med. 3:917–921. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lin JH, Deng H, Huang Q and Morser J:
KIAP, a novel member of the inhibitor of apoptosis protein family.
Biochem Biophys Res Commun. 279:820–831. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Badran A, Yoshida A, Ishikawa K, Goi T,
Yamaguchi A, Ueda T and Inuzuka M: Identification of a novel splice
variant of the human anti-apoptosis gene survivin. Biochem Biophys
Res Commun. 314:902–907. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Eckelman BP, Salvesen GS and Scott FL:
Human inhibitor of apoptosis proteins: why XIAP is the black sheep
of the family. EMBO Rep. 7:988–994. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Roy N, Deveraux QL, Takaahashi R, Salvesen
GS and Reed JC: The c-IAP-1 and c-IAP-2 proteins are direct
inhibitors of specific caspases. EMBO J. 16:6914–6925. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Vaux DL and Silke J: Mammalian
mitochondrial IAP binding proteins. Biochem Bioph Res Commun.
304:499–504. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wrzesień-Kuś A, Smolewski P, Sobczak-Pluta
A, Wierzbowska A and Robak T: The inhibitor of apoptosis protein
family and its antagonists in acute leukemias. Apoptosis.
9:705–715. 2004.PubMed/NCBI
|
|
14
|
Yan B: Research progress on Livin protein:
an inhibitor of apoptosis. Mol Cell Biochem. 357:39–45. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lv J and Chen ZC: Resent research about
Livin in cancer. Chin J Cancer Prev Treat. 13:1347–1350. 2006.
|
|
16
|
Ashhab Y, Alian A, Polliack A, Panet A and
Ben Yehuda D: Two splicing variants of a new inhibitor of apoptosis
gene with different biological properties and tissue distribution
pattern. FEBS Lett. 495:56–60. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yagihashi A, Ohmura T, Asanuma K,
Kobayashi D, Tsuji N, Torigoe T, Sato N, Hirata K and Watanabe N:
Detection of auto-antibodies to survivin and livin in sera from
patients with breast cancer. Clin Chim Acta. 362:125–130. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hariu H, Hirohashi Y, Toriegoe T, Asanuma
H, Hariu M, Tamura Y, Aketa K, Nabeta C, Nakanishi K, Kamiguchi K,
Mano Y, Kitamura H, Kobayashi J, Tsukahara T, Shijubo N and Sato
Nl: Aberrant expression and potency as a cancer immunotherapy
target of inhibitor of apoptosis protein family, Livin/ML-IAP in
lung cancer. Clin Cancer Res. 11:1000–1009. 2005.PubMed/NCBI
|
|
19
|
Chung CY, Park YL, Kim N, Park HC, Park
HB, Myung DS, Kim JS, Cho SB, Lee WS and Joo YE: Expression and
prognostic significance of Livin in gastric cancer. Oncol Rep.
30:2520–2528. 2013.PubMed/NCBI
|
|
20
|
Qiuping Z, Jei X, Youxin J, Wei J, Chun L,
Jin W, Qun W, Yan L, Chungsong H, Mingzhen Y, Qingping G, Kejian Z,
Qun L, Junyan L and Jinguan T: CC chemokine ligand 25 enhances
resistance to apoptosis in CD4+ T cells from patients
with T-cell lineage acute and chronic lymphocytic leukemia by means
of livin activation. Cancer Res. 64:7579–7587. 2004.PubMed/NCBI
|
|
21
|
Choi J, Hwang YK, Sung KW, Lee SH, Yoo KH,
Jung HL, Koo HH, Kim HJ, Kang HJ, Shin HY and Ahn HS: Expression of
Livin, an antiapoptotic protein, is an independent favorable
prognostic factor in childhood acute lymphoblastic leukemia. Blood.
109:471–477. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gazzaniga P, Gradilone A, Giuliani L,
Gandini O, Silvestri I, Nofroni I, Saccani G, Frati L and Aglianò
AM: Expression and prognostic significance of LIVIN, SURVIVIN and
other apoptosis-related genes in the progression of superficial
bladder cancer. Ann Oncol. 14:85–90. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kim DK, Alvarado CS, Abramowsky CR, Gu L,
Zhou M, Soe MM, Sullivan K, George B, Schemankewitz E and Findly
HW: Expression of inhibitor-of-apoptosis protein (IAP) livin by
neuroblastoma cells: correlation with prognostic factors and
outcome. Pediatr Dev Pathol. 8:621–629. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gordon GJ, Mani M, Mukhopadhyay L, Dong L,
Edenfield HR, Glickman JN, Yeap BY, Sugarbaker DJ and Bueno R:
Expression patterns of inhibitor of apoptosis proteins in malignant
pleural mesothelioma. J Pathol. 211:447–454. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lopes RB, Gangeswaran R, McNeish IA, Wang
Y and Lemoine NR: Expression of the IAP protein family is
dysregulated in pancreatic cancer cells and is important for
resistance to chemotherapy. Int J Cancer. 120:2344–2352. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Nedelcu T, Kubista B, Koller A, sutzbacher
I, Mosberger I, Arrich F, Trieb K, Kotz R and Toma CD: Livin and
Bcl-2 expression in high-grade osteosarcoma. J Cancer Res Clin
Oncol. 2:237–244. 2008.PubMed/NCBI
|
|
27
|
Crnković-Mertens I, Wagener N, Semzow J,
Grőne EF, Haferkamp A, Hohenfellner M, Butz K and Hoppe-Seyler F:
Targeted inhibition of Livin resensitizes renal cancer cells
towards apoptosis. Cell Mol Life Sci. 64:1137–1144. 2007.PubMed/NCBI
|
|
28
|
Kempkensteffen C, Hinz S, Christoph F,
Krause H, Koellemann J, Magheli A, Schrader M, Schostak M, Miller K
and Weikert S: Expression of the apoptosis inhibitor livin in renal
cell carcinomas: correlations with pathology and outcome. Tumor
Biol. 28:132–138. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Myung DS, Park YL, Chung CY, Park HC, Kim
JS, Cho SB, Lee WS, Lee KH, Lee JH and Joo YE: Expression of Livin
in colorectal cancer and its relationship to tumor cell behavior
and prognosis. PLoS One. 8:e732622013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu X, Chen N, Wang X, He Y, Chen X, Huang
Y, Yin W and Zhou Q: Apoptosis and proliferation markers in
diffusely infiltrating astrocytomas: profiling of 17 molecules. J
Neuropathol Exp Neurol. 65:905–913. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang R, Lin F, Wang X, Gao P, Dong K, Zou
AM, Cheng SY, Wei SH and Zhang HZ: Silencing Livin gene expression
to inhibit proliferation and enhance chemosensitivity in tumor
cells. Cancer Gene Ther. 15:402–412. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xiang Y, Yao H, Wang S, Hong M, He J, Cao
S, Min H, Song E and Guo X: Prognostic value of Survivin and Livin
in nasopharyngeal carcinoma. Laryngoscope. 116:126–130. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Edge S, Byrd DR, Compton CC, Fritz AG,
Greene FL and Trotti A: American Joint Committee on Cancer - Cancer
Staging Manual. 7th edition. Springer; New York: pp. 6492010
|
|
34
|
Kenneth NS and Duckett CS: IAP proteins;
regulators of cell migration and development. Curr Opin Cell Biol.
24:871–875. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Green D and Kroemer G: The central
executioners of apoptosis: caspases or mitochondrial. Trends Cell
Bio. 8:267–271. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Xi RC, Biao WS and Gang ZZ: Significant
elevation of survivin and livin expression in human colorectal
cancer: inverse correlation between expression and overall
survival. Onkologie. 34:428–432. 2011. View Article : Google Scholar
|
|
37
|
Kasof GM and Gomes BC: Livin, a novel
inhibitor of apoptosis protein family member. J Biol Chem.
276:3238–46. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sun JG, Liao RX, Zhang SX, Duan YZ, Zhuo
WL, Wang XX, Wang ZX, Li DZ and Chen ZT: Role of inhibitor of
apoptosis protein Livin in radiation response in nonsmall cell lung
cancer. Cancer Biother Radiopharm. 26:585–592. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang L, Zhang Q, Liu B, Han M and Shan B:
Challenge and promise: roles for Livin in progression and therapy
of cancer. Mol Cancer Ther. 7:3661–3669. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chang H and Schimmer AD: Livin/melanoma
inhibitor of apoptosis protein as a potential therapeutic target
for the treatment of malignancy. Mol Cancer Ther. 6:24–30. 2007.
View Article : Google Scholar
|
|
41
|
Liu B, Han M, Wen JK and Wang L:
Livin/ML-IAP as a new target for cancer treatment. Cancer Lett.
250:168–176. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wagener N, Crnković-Mertens I, Vetter C,
Macher-Gőppinger S, Bedke J, Grőne EF, Zentgraf H, Pritsch M,
Hoppe-Seyler K, Buse S, Haferkamp A, Autschbach F, Hohenfellner M
and Hoppe-Seyler F: Expression of inhibitor of apoptosis protein
Livin in renal cell carcinoma and non-tumorous adult kidney. Br J
Cancer. 97:1271–1276. 2007. View Article : Google Scholar : PubMed/NCBI
|