Introduction
Mucins are the major glycoproteins of the
gastrointestinal tract; secretory mucin 2 (MUC2) is the main
component of the protective mucus layer. Although MUC2 expression
is decreased in colorectal adenocarcinoma, colonic mucinous
carcinomas are characterized by overexpression or ectopic
expression of MUC2 (1). In
contrast, the suppressive effect of MUC2 in colorectal cancer (CRC)
was demonstrated in MUC2-null mice (2), suggesting that it has a protective
effect in the colon. Although MUC2 overexpression was specifically
associated with mucinous carcinoma, its effects on tumor
progression remain unclear.
Intestinal inflammation is a crucial component of
tumor development and metastasis (3), and polymorphonuclear neutrophils and
macrophages may affect tumor development and progression in CRC
(3). The effects of interleukin
(IL)-6 on tumor growth are multifaceted. In previous studies,
tumor-infiltrating macrophages (TIMs) were found to express a high
level of IL-6 (4,5), which is implicated in CRC
carcinogenesis. In prostatic carcinoma, IL-6 acts as a growth
factor (6,7). In CRC patients, IL-6 levels were
significantly higher than these levels in healthy controls
(8,9) and were correlated with tumor stage,
tumor size, liver metastasis and poor survival (8,10–12).
Moreover, macrophage secretion of IL-6 decreased MUC2 expression in
a human colon cancer cell line through phosphorylation of STAT3
(13). IL-6 can also regulate the
immune microenvironment surrounding the tumor (14). However, the effects of MUC2
expression on IL-6 secretion by colon cancer cells have not been
determined. Therefore, we employed RNA interference (RNAi) to
suppress MUC2 expression in mouse colon carcinoma cells, and
evaluated the effects of MUC2 suppression on IL-6 production and
tumor growth.
Materials and methods
Cell culture
CT26 colon carcinoma cells (BALB/c mouse origin)
were kindly provided by Dr M.D. Lai of the National Cheng Kung
University (Tainan, Taiwan). CT26 cells were maintained in high
glucose Dulbecco’s modified Eagle’s medium (DMEM)
(Gibco-Invitrogen) containing 10% FBS (HyClone Laboratories, Logan,
UT, USA) and 1% penicillin/streptomycin. All cell lines were stored
in a humidified 37°C incubator with 5% CO2.
Mice
Eight-week-old BALB/c (H2d)
mice and non-obese diabetes/severe combined immunodeficiency
(NOD/SCID) mice, NOD.CB17-PRKDC, were purchased from the Laboratory
Animal Center of National Cheng Kung University and were maintained
under pathogen-free conditions. All animal experiments were
conducted with the approval of the Institutional Animal Care and
Use Committee (IACUC) of the National Cheng Kung University.
RNA interference
siRNA targeting mouse MUC2 was constructed within
the pHsU6 vector as described previously (15) using the following target sequences:
5′-GCTATGT GCCTGGCTCTAA-3′ (RNAi-1), 5′-GCAACAAGTGCACC TT CTT-3′
(RNAi-2) and 5′-GCTGCCCTACAAACTGTTT-3′ (RNAi-3). Cells were
cotransfected with the pHsU6 vectors containing the shRNA target
sequences, a nonspecific sequence (scrambled shRNA), and pCMV-neo
using Lipofectamine 2000 transfection reagent (Invitrogen,
Carlsbad, CA, USA). The stable transfectants were selected with
G418 (Calbiochem, San Diego, CA, USA). We established three clones
of MUC2-suppressed cells, including CT26 MUC2 RNAi-1, CT26 MUC2
RNAi-2 and CT26 MUC2 RNAi-3. The clonal cell lines were maintained
in complete medium with 250 μg/ml G418. To monitor the efficacy of
MUC2 silencing, the expression of MUC2 in CT26 stable transfectants
was analyzed by western blotting.
Western blot analysis
A MUC2-specific antibody was obtained by inoculating
rabbits with the mouse MUC2 peptide, CVRTRRSSPRFLGRK (c-terminal
position 911–924). Peptides used in this study were synthesized and
purified by Genemed Synthesis (San Antonio, TX, USA). Total cell
lysates were prepared and analyzed by SDS-PAGE as previously
described (16). Immunodetection
was performed using a horse-radish peroxidase (HRP)-based
SuperSignal Chemiluminescent Substrate (Pierce, Rockford, IL, USA).
For quantification, the bands were measured by the AlphaImager 2200
system (Alpha Innotech, San Leandro, CA, USA) and normalized by the
density obtained for β-actin.
Cell proliferation assay
Cells (1×103/well) were seeded in
quadruplicate onto 96-well plates and incubated at 37°C under 5%
CO2. At 24, 48 and 72 h, viable cell numbers were
measured using the CellTiter 96 Aqueous One Solution Cell
Proliferation assay (Promega, Madison, WI, USA) according to the
manufacturer’s instructions. The proliferation curves were
constructed by calculating the mean value of absorbance at 490 nm
with an Ultra Multifunctional Microplate Reader (Tecan, Durham, NC,
USA).
Orthotopic model of colon
adenocarcinoma
Eight-week-old female BALB/c mice were anesthetized
by intraperitoneal injection of Zoletil (50 mg/kg; Parnell
Laboratories, Alexandria, NSW, Australia) and xylazine (10 mg/kg;
Troy Laboratories, Glendenning, NSW, Australia). After making a
small median abdominal incision in the mice under anesthesia,
cecums were exteriorized and 1×106 cells (scramble RNA,
MUC2 RNAi-1, RNAi-2 or RNAi-3 tumor cells) in 0.05 ml of PBS were
injected into the middle wall of the greater curvature of the cecal
wall using a 1-cc U-100 insulin disposable syringe
(Becton-Dickinson, Franklin Lakes, NJ, USA). Bipolar (ICC, ER BE)
coagulation was used when the needle was removed from the wall of
the cecum. Each cecum was then returned to the peritoneal cavity,
and the abdominal wall and skin were closed with a Dexon 4-0
surgical suture (Ethicon, Bridgewater, NJ, USA). The mice were
sacrificed 11–17 days after the tumor cell implantation or when
moribund. The whole cecums from mice were excised, washed, removed
of remaining diet, and weighed. For IL-6 antibody neutralization,
BALB/c mice were pretreated with an injection of 100 μg/mouse of
IL6-neutralizing antibody or isotype control antibody (both from BD
Pharmingen, San Diego, CA, USA) 2 days prior to tumor cell
injection. After tumor cell injection, mice were treated with IL-6
neutralizing antibody or control antibody every 3 days by
intraperitoneal injection as previously described (17).
Cytokine array and enzyme-linked
immunosorbent assay (ELISA)
To collect culture supernatants, scrambled control
and CT26 MUC2 RNAi-1 cells were cultured for 48 h in serum-free
medium after which the supernatants were collected and measured
using Mouse Cytokine Antibody Array II kit (RayBiotech, Norcross,
GA, USA) and an IL-6 ELISA (R&D Systems, Minneapolis, MN, USA)
according to the manufacturers’ instructions. The signal intensity
of the array was scanned and quantified by densitometry using an
AlphaImager 2200 system (Alpha Innotech) and normalized to the
positive control.
Flow cytometric analysis
To characterize the immune cells of the peritoneal
fluid in vivo, peritoneal fluid was isolated and subjected
to flow cytometry as previously described (16,18).
The following fluorescent-labeled antibodies were purchased from BD
Bioscience: rat anti-Ly6G-FITC, anti-CD4-PE, anti-CD8-PE,
anti-CD11b-PE and isotype controls (FITC and PE). Data analysis was
evaluated using a flow cytometer (FACScan; Becton-Dickinson).
Statistical analysis
Data are expressed as means ± SD. Statistical
analyses were performed using the Student’s t-test. A P-value
<0.05 was considered significant for all comparisons.
Results
Suppression of MUC2 expression in mouse
CT26 colon carcinoma cells by RNAi
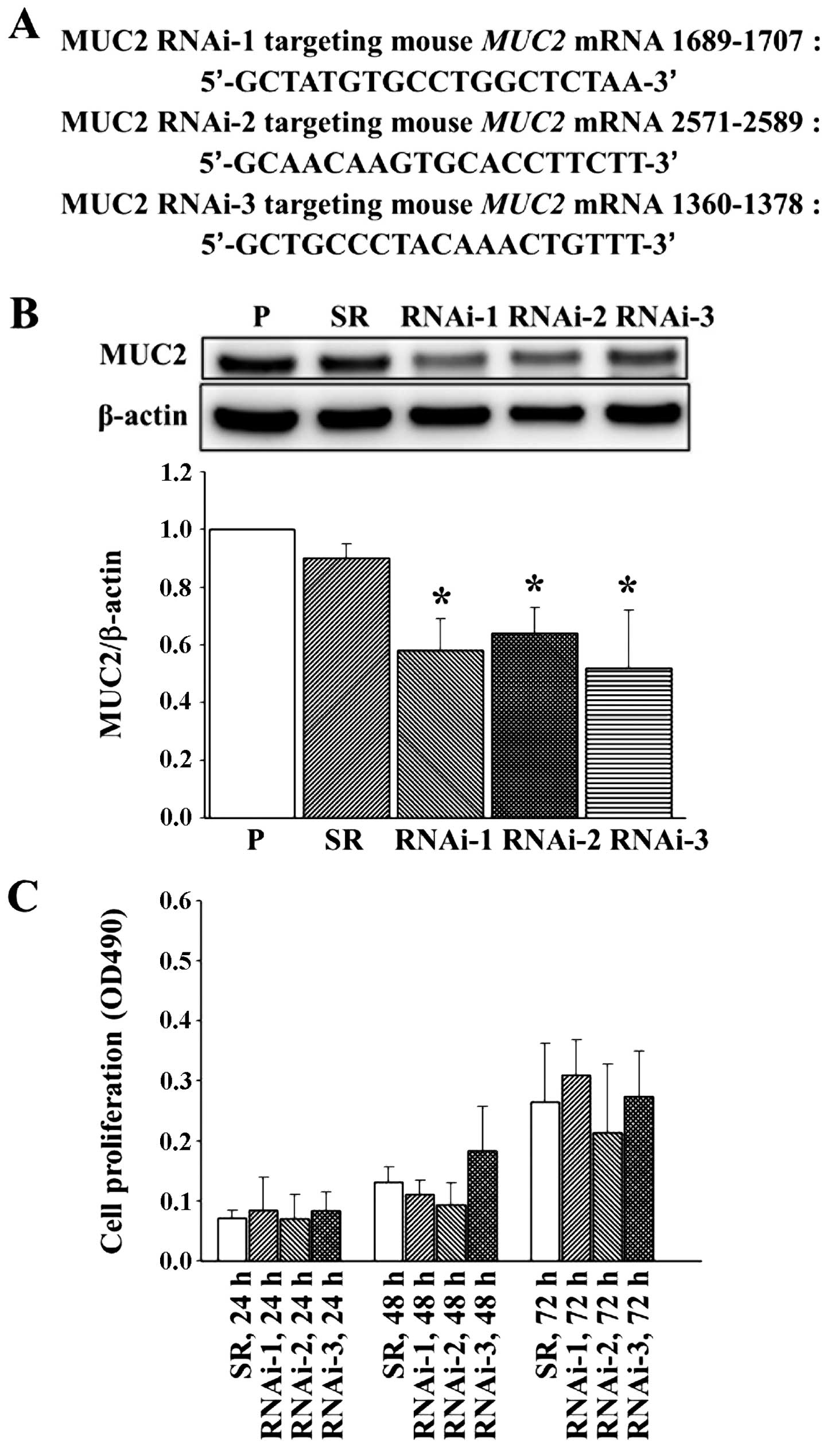
To study the role of MUC2 in colon cancer, we used
shRNA to suppress the expression of MUC2 in the CT26 murine colon
cancer cell line, which expresses high levels of MUC2. Three shRNAs
targeting different sites on MUC2 were transfected into CT26 cells,
and the stable transfectants were established by G418 selection.
The vector control was established by transfecting CT26 cells with
an empty vector, and the scrambled RNA (SR) transfectant contains a
different shRNA sequence with the same base composition of RNAi-1.
Stable cells expressing three RNAi target sequences were
established: MUC2 RNAi-1, MUC2 RNAi-2 and MUC2 RNAi-3 (Fig. 1A). Western blot analysis confirmed
that the SR did not alter MUC2 protein levels (Fig. 1B). However, MUC2 expression was
significantly decreased in the MUC2 RNAi-1, MUC2 RNAi-2 and MUC2
RNAi-3 CT26 cells (Fig. 1B).
MUC2 suppression does not affect cell
proliferation in vitro
To determine the effects of MUC2 suppression on cell
growth, cells were seeded at a low density, and growth rates were
determined for the parental cell line and its derived cell clones
at 24, 48 and 72 h. MUC2 silencing did not inhibit the in
vitro proliferation of CT26 cells (Fig. 1C).
Effect of MUC2 suppression on CT26 tumor
growth in vivo
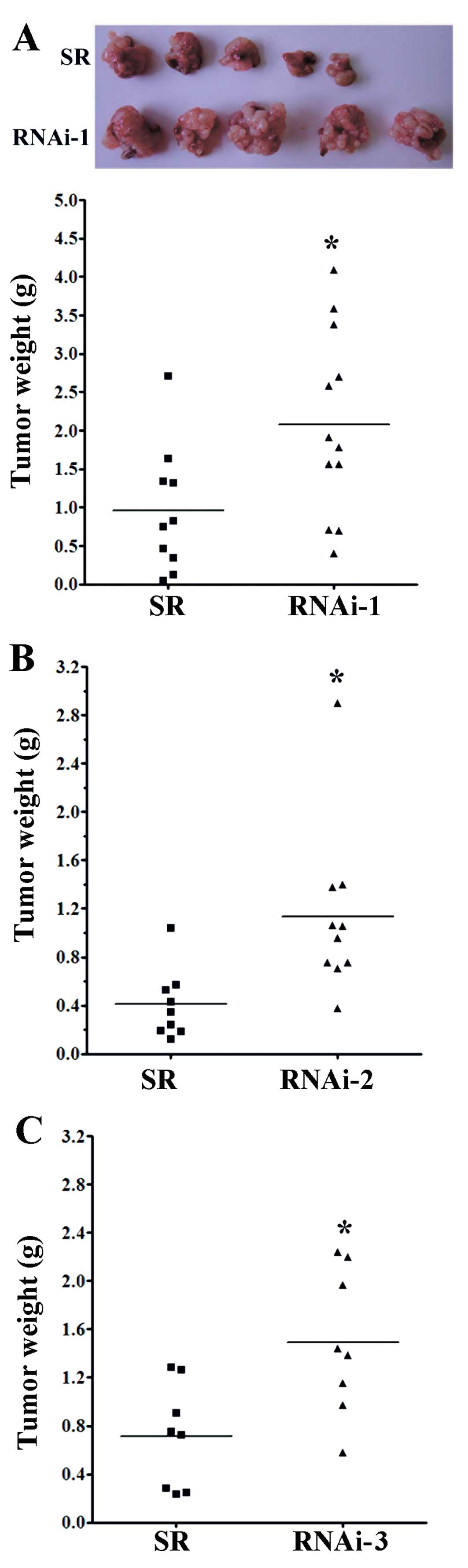
To investigate the role of MUC2 in a native tumor
environment, we examined the effects of MUC2 knockdown in an
orthotopic immune-competent animal model. One million SR control or
MUC2 shRNA-expressing cells were implanted orthotopically in BALB/c
mice, and macroscopic tumor nodules indicative of tumor formation
were detected (Fig. 2A). The tumor
weight of mice injected with SR cells was significantly lower than
the tumor weights of the mice injected with RNAi-1 (Fig. 2A), RNAi-2 (Fig. 2B) and RNAi-3 cells (Fig. 2C) at day 17. These results
demonstrated that knockdown of MUC2 promoted the tumor growth of
colon cancer cells in vivo, suggesting that MUC2 plays an
important role in the tumorigenicity of colon cancer.
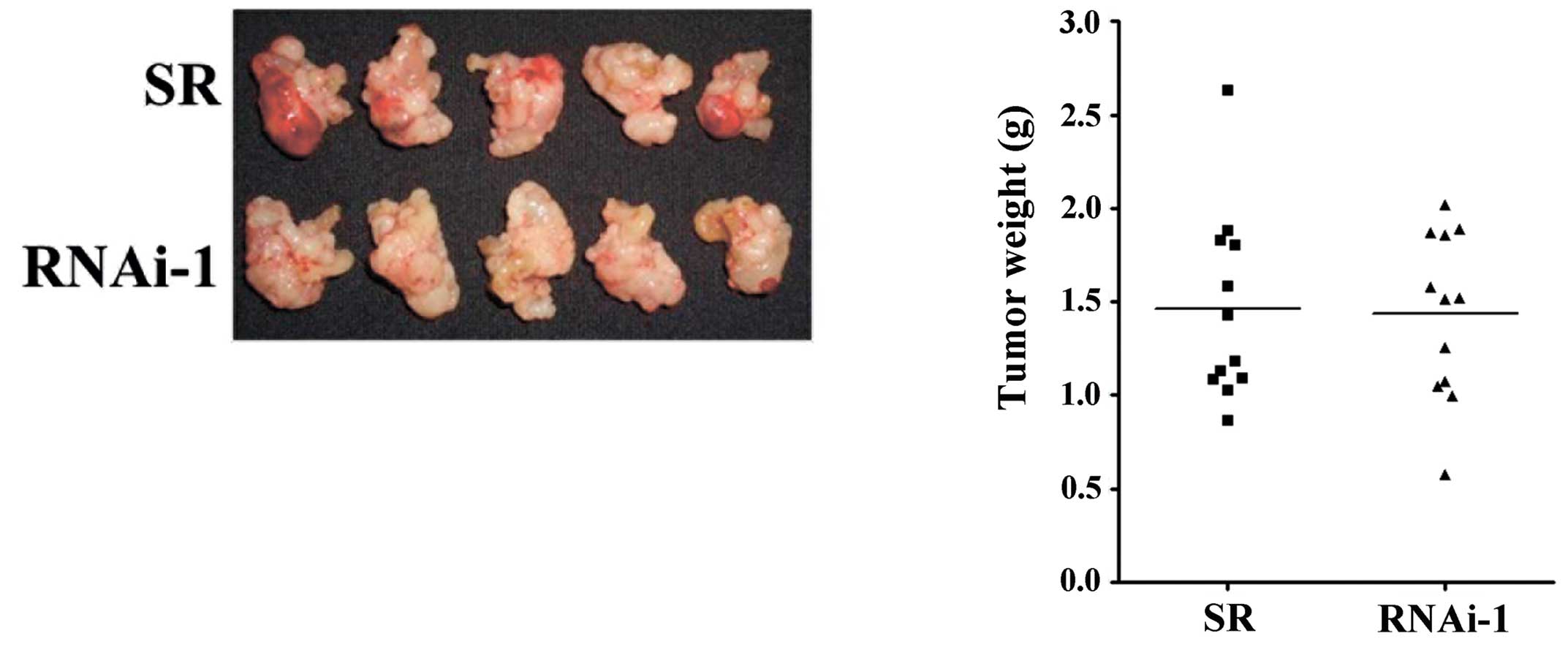
Since the growth of the MUC2 shRNA transfectants was
not altered in vitro, we hypothesized that the changes in
tumor growth in vivo may result from altered
tumor-microenvironment interaction. To assess the potential
immunological effects of MUC2 on tumor progression, tumor growth
was measured in the immune-deficient NOD/SCID mice. In the absence
of NK cells, macrophages, B and T cells, the mean tumor mass of
mice implanted with the SR cells was similar to the mean tumor mass
of mice implanted with MUC2 RNAi-1 cells (Fig. 3), suggesting that the effects of
MUC2 were dependent upon the presence of a competent immune
system.
Suppression of MUC2 increases IL-6
secretion by CT26 colon cancer cells
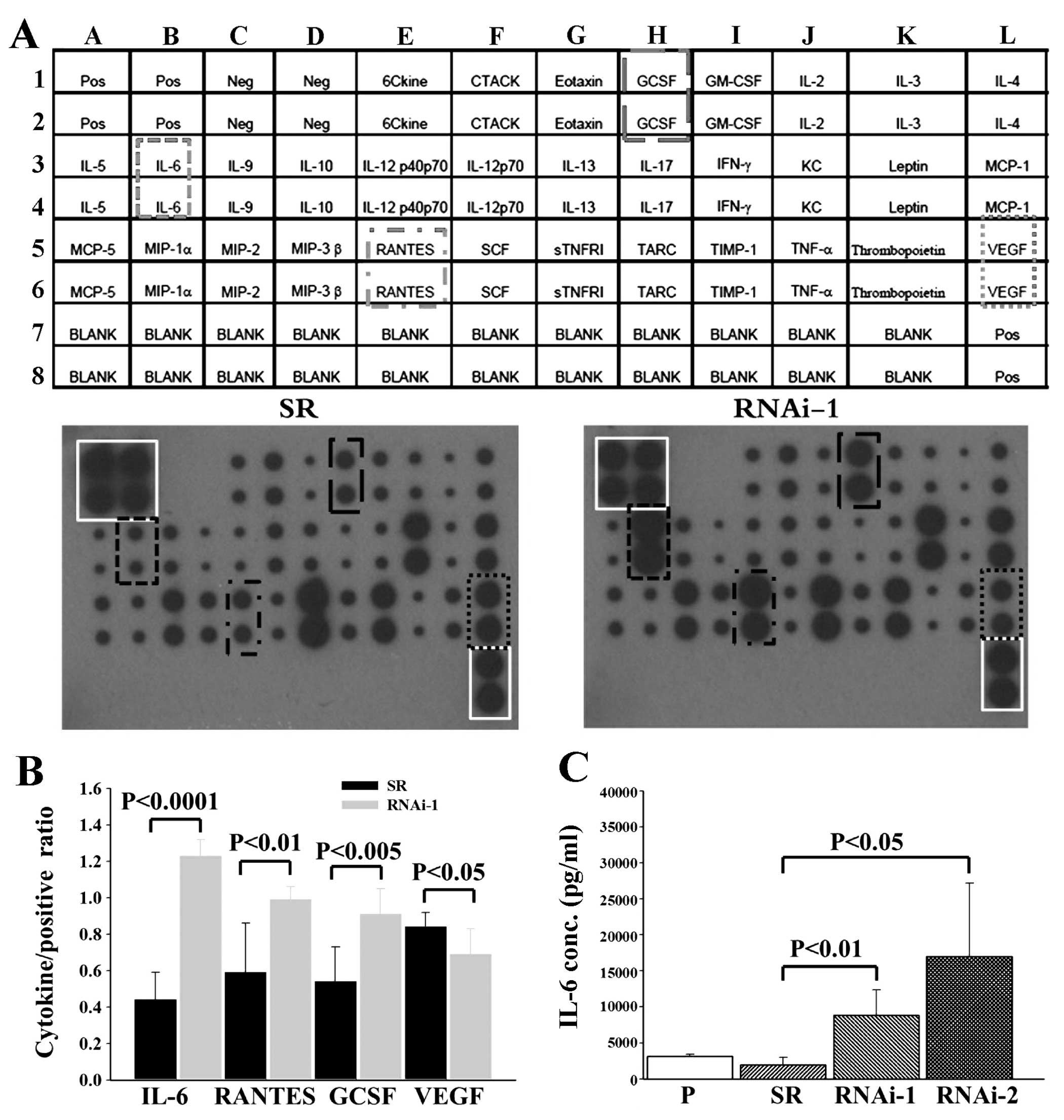
Given the importance of a functional immune system
for the effects of MUC2 on tumor growth, we further investigated
whether cancer cell-secreted cytokines were involved in the tumor
microenvironment. Specifically, the cytokine profile consisting of
32 different factors in the conditioned medium of SR and MUC2
RNAi-1 cells was compared 48 h after serum-deprivation. Conditioned
medium from MUC2 RNAi-1 cells had significantly increased IL-6,
regulated on activation, normal T cell expressed and secreted
(RANTES) and granulocyte colony-stimulating factor (GCSF)
expression and decreased vascular endothelial growth factor (VEGF)
expression compared to the SR conditioned medium (Fig. 4). IL-6 secretion by SR and MUC2
RNAi-1 cells 48 h after serum-deprivation was further quantified by
ELISA. MUC2 RNAi-1 and MUC2 RNAi-2 cells secreted significantly
higher levels of IL-6 than SR cells (Fig. 4C). Therefore, MUC2 expression by
colon cancer cells alters IL-6 secretion.
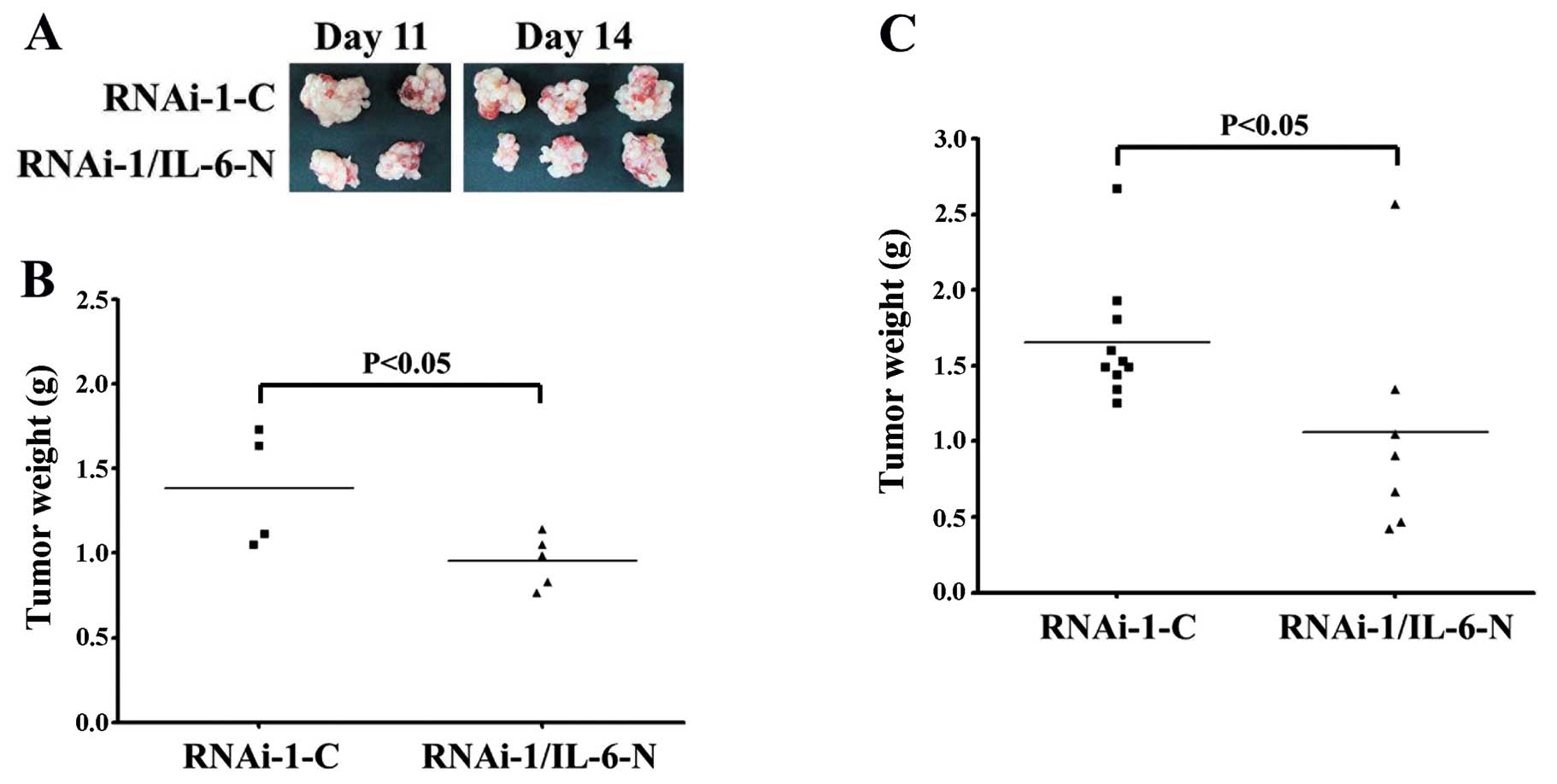
IL-6 neutralization attenuates tumor
formation by CT26 MUC2 knockdown cells
To confirm the biological effect of IL-6 in
vivo, we tested whether an IL-6 neutralization antibody could
inhibit tumorigenic growth. Mice pretreated with either an
IL-6-neutralizing antibody or control IgG antibody were injected
with MUC2 RNAi-1 cells, and then treated every 3 days with either
IL-6 or control antibody. The growth of MUC2 RNAi-1 tumors was
reduced with the IL-6 neutralizing antibodies on day 11 (Fig. 5A and B) and day 14 (Fig. 5A and C). These experiments suggest
that MUC2 knockdown enhanced IL-6 secretion and promoted tumor
growth.
IL-6 neutralization enhances the
CD8-mediated immune response in CT26 cells after MUC2
silencing
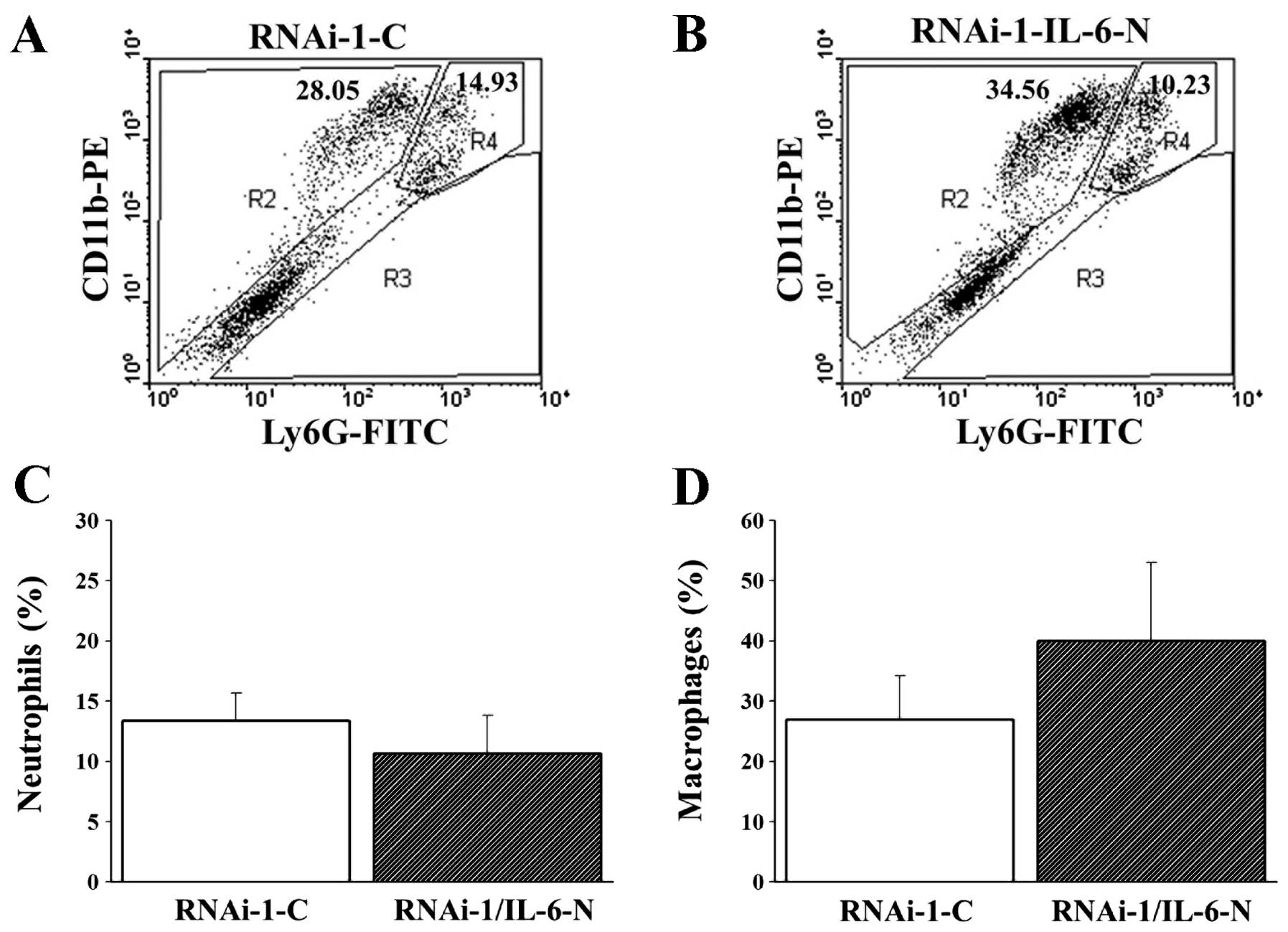
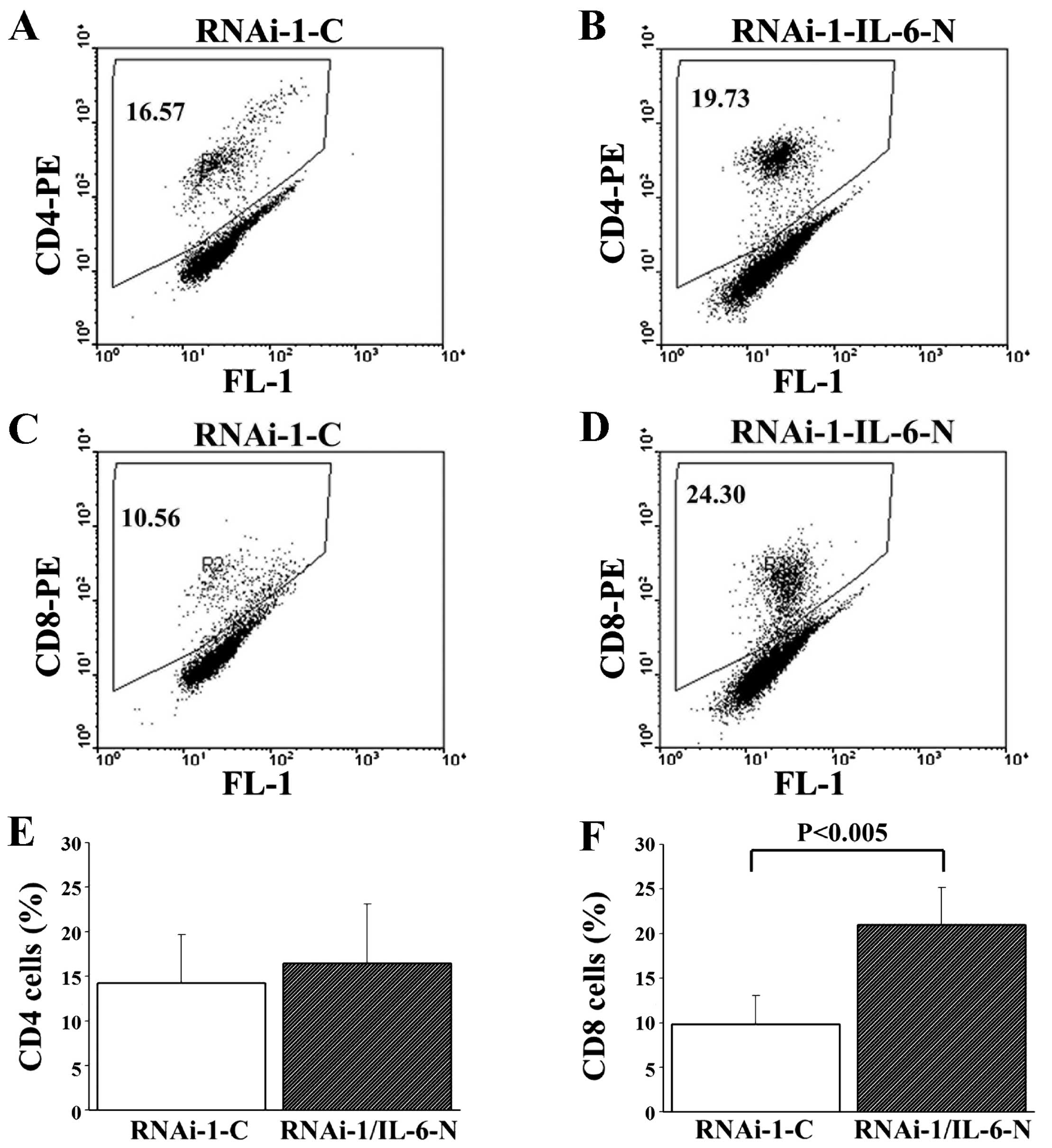
To elucidate the possibility that MUC2 regulates
IL-6 secretion and induces a local immune response in colon cancer
cells, we analyzed Ly6G+CD11b+ neutrophil,
Ly6G−CD11b+ macrophage, CD4 T cell and CD8 T
cell levels in the peritoneal fluid of mice with MUC2 RNAi-1 cell
tumors treated with control or IL-6 neutralization antibodies.
Although there was no significant difference in the proportion of
Ly6G+CD11b+ neutrophils,
Ly6G−CD11b+ macrophages and CD4 T cells in
the peritoneal fluid between the two groups on day 14 (Figs. 6 and 7A,
B and E), the peritoneal fluid of the MUC2 RNAi-1 tumor-bearing
mice treated with IL6-neutralizing antibody had a significantly
greater proportion of CD8 T cells than MUC2 RNAi-1 tumor-bearing
mice treated with the control antibody on day 14 (Fig. 7C, D and F). Thus, an IL-6
neutralizing antibody could inhibit the in vivo growth of
MUC2 knockdown tumors, increasing CD8 T cell influx in the
peritoneal cavity.
Discussion
In the present study, we examined the effects of
MUC2 on tumor cell growth, IL-6 secretion and the immune response
in colon cancer. To the best of our knowledge, this is the first
study to demonstrate that downregulation of MUC2 expression
enhances IL-6 secretion as well as tumor growth. Thus, MUC2 had a
protective effect during tumorigenesis. This observation is
consistent with previous studies in which loss of MUC2 expression
was associated with progression and metastasis in CRC (19–23).
Moreover, CRC patients with a high MUC2/carcinoembryonic antigen
(CEA) mRNA ratio in their lymph nodes had a significantly better
prognosis than those with a low ratio (24). In addition, MUC2-positive CRC was
found to be correlated with reduced disease recurrence and
prolonged survival as well as a low incidence of liver and nodal
metastasis (1,25,26).
In previous studies, MUC2 suppressed inflammation in
the intestinal tract and inhibited intestinal tumorigenesis
(2,27). In the present study, the secretion
of IL-6, RANTES and GCSF was increased in the MUC2-knockdown tumor
cells in comparison to the SR tumor cells. Both IL-6 and RANTES
were associated with tumor progression, metastasis and macrophage
activation in previous studies (28–32).
In particular, IL-6 is an inflammatory cytokine released by T
cells, macrophages and several cancer cell types (6,7,33).
IL-6 production by tumor cells may act as an autocrine and/or
paracrine growth factor during carcinogenesis, and various studies
have proposed promising targets for CRC therapy using anti-IL-6 and
anti-IL-6R antibodies as well as soluble gp130Fc (sgp130Fc) and
selective small-molecule JAK inhibitors that suppress the
IL-6/STAT3 pathway (34). The
increased secretion of IL-6 by MUC2-knockdown tumor cells and
enhanced tumor growth observed in the present study suggest that
the protective mechanisms of MUC2 in colon cancer may be associated
with its effects on suppressing the IL-6/STAT3 signaling
pathway.
Inflammatory cells of the innate and adaptive immune
system may affect different stages of tumor progression or
metastasis in colon cancer. Since MUC2 silencing increased IL-6
secretion, we analyzed neutrophil, macrophage, CD4 T cell and CD8 T
cell levels in the peritoneal fluid of mice. In this study, mice
bearing MUC2-knockdown tumors and treated with IL-6 neutralizing
antibodies displayed increased CD8 T cell infiltration into the
peritoneal fluid. To the best of our knowledge, this is the first
study to demonstrate that the immunoregulatory effects of MUC2 may
represent a novel therapeutic method for treating cancer.
Alternatively, tumor-associated antigens (TAAs) have served as a
target for CTL immunotherapy as DNA vaccines have a therapeutic
effect on established tumors through activation of a CD8 T
cell-dependent pathway (35,36).
Thus, it is intriguing that MUC2 may induce a systemic antitumor
immune response and may have therapeutic value in the treatment of
IL-6-secreting cancers. In conclusion, the therapeutic effects of
MUC2 may be mediated at least in part by decreasing IL-6 secretion,
inhibiting tumorigenicity, and inducing CD8 T cells in
vivo.
Acknowledgements
This study was supported by grants from the National
Science Council (NSC100-2320-B041-006 and NSC102-2311-B041-001) and
the Department of Health, Executive Yuan, Taiwan
(101-TD-C-111-003).
References
|
1
|
Hanski C, Hofmeier M, Schmitt-Gräff A, et
al: Overexpression or ectopic expression of MUC2 is the common
property of mucinous carcinomas of the colon, pancreas, breast, and
ovary. J Pathol. 182:385–391. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Velcich A, Yang W, Heyer J, et al:
Colorectal cancer in mice genetically deficient in the mucin Muc2.
Science. 295:1726–1729. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Coussens LM and Werb Z: Inflammation and
cancer. Nature. 420:860–867. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Piancatelli D, Romano P, Sebastiani P,
Adorno D and Casciani CU: Local expression of cytokines in human
colorectal carcinoma: evidence of specific interleukin-6 gene
expression. J Immunother. 22:25–32. 1999. View Article : Google Scholar
|
|
5
|
Lieubeau B, Heymann MF, Henry F, Barbieux
I, Meflah K and Grégoire M: Immunomodulatory effects of
tumor-associated fibroblasts in colorectal-tumor development. Int J
Cancer. 81:629–636. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Giri D, Ozen M and Ittmann M:
Interleukin-6 is an autocrine growth factor in human prostate
cancer. Am J Pathol. 159:2159–2165. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Adler HL, McCurdy MA, Kattan MW, Timme TL,
Scardino PT and Thompson TC: Elevated levels of circulating
interleukin-6 and transforming growth factor-beta 1 in patients
with metastatic prostatic carcinoma. J Urol. 161:182–187. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Esfandi F, Mohammadzadeh Ghobadloo S and
Basati G: Interleukin-6 level in patients with colorectal cancer.
Cancer Lett. 244:76–78. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Galizia G, Orditura M, Romano C, et al:
Prognostic significance of circulating IL-10 and IL-6 serum levels
in colon cancer patients undergoing surgery. Clin Immunol.
102:169–178. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ueda T, Shimada E and Urakawa T: Serum
levels of cytokines in patients with colorectal cancer: possible
involvement of interleukin-6 and interleukin-8 in hematogenous
metastasis. J Gastroenterol. 29:423–429. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kinoshita T, Ito H and Miki C: Serum
interleukin-6 level reflects the tumor proliferative activity in
patients with colorectal carcinoma. Cancer. 85:2526–2531. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chung YC and Chang YF: Serum interleukin-6
levels reflect the disease status of colorectal cancer. J Surg
Oncol. 83:222–226. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li YY, Hsieh LL, Tang RP, Liao SK and Yeh
KY: Macrophage-derived interleukin-6 up-regulates MUC1, but
down-regulates MUC2 expression in the human colon cancer HT-29 cell
line. Cell Immunol. 256:19–26. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Atsumi T, Singh R, Sabharwal L, et al:
Inflammation amplifier, a new paradigm in cancer biology. Cancer
Res. 74:8–14. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lu TJ, Lai WY, Huang CY, et al: Inhibition
of cell migration by autophosphorylated mammalian sterile 20-like
kinase 3 (MST3) involves paxillin and protein-tyrosine
phosphatase-PEST. J Biol Chem. 281:38405–38417. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shan YS, Fang JH, Lai MD, et al:
Establishment of an orthotopic transplantable gastric cancer animal
model for studying the immunological effects of new cancer
therapeutic modules. Mol Carcinog. 50:739–750. 2011. View Article : Google Scholar
|
|
17
|
Ancrile B, Lim KH and Counter CM:
Oncogenic Ras-induced secretion of IL6 is required for
tumorigenesis. Genes Dev. 21:1714–1719. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fridlender ZG, Sun J, Kim S, et al:
Polarization of tumor-associated neutrophil phenotype by TGF-beta:
‘N1’ versus ‘N2’ TAN. Cancer Cell. 16:183–194. 2009.PubMed/NCBI
|
|
19
|
Ajioka Y, Allison LJ and Jass JR:
Significance of MUC1 and MUC2 mucin expression in colorectal
cancer. J Clin Pathol. 49:560–564. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yonezawa S, Sueyoshi K, Nomoto M, et al:
MUC2 gene expression is found in noninvasive tumors but not in
invasive tumors of the pancreas and liver: its close relationship
with prognosis of the patients. Hum Pathol. 28:344–352. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li A, Goto M, Horinouchi M, et al:
Expression of MUC1 and MUC2 mucins and relationship with cell
proliferative activity in human colorectal neoplasia. Pathol Int.
51:853–860. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bresalier RS, Niv Y, Byrd JC, et al: Mucin
production by human colonic carcinoma cells correlates with their
metastatic potential in animal models of colon cancer metastasis. J
Clin Invest. 87:1037–1045. 1991. View Article : Google Scholar
|
|
23
|
Blank M, Klussmann E, Krüger-Krasagakes S,
et al: Expression of MUC2-mucin in colorectal adenomas and
carcinomas of different histological types. Int J Cancer.
59:301–306. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ohlsson L, Israelsson A, Oberg A, et al:
Lymph node CEA and MUC2 mRNA as useful predictors of outcome in
colorectal cancer. Int J Cancer. 130:1833–1843. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kang H, Min BS, Lee KY, et al: Loss of
E-cadherin and MUC2 expressions correlated with poor survival in
patients with stages II and III colorectal carcinoma. Ann Surg
Oncol. 18:711–719. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Elzagheid A, Emaetig F, Buhmeida A, et al:
Loss of MUC2 expression predicts disease recurrence and poor
outcome in colorectal carcinoma. Tumour Biol. 34:621–628. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Van der Sluis M, De Koning BA, De Bruijn
AC, et al: Muc2-deficient mice spontaneously develop colitis,
indicating that MUC2 is critical for colonic protection.
Gastroenterology. 131:117–129. 2006.PubMed/NCBI
|
|
28
|
Eichbaum C, Meyer AS, Wang N, et al:
Breast cancer cell-derived cytokines, macrophages and cell
adhesion: implications for metastasis. Anticancer Res.
31:3219–3227. 2011.PubMed/NCBI
|
|
29
|
Luboshits G, Shina S, Kaplan O, et al:
Elevated expression of the CC chemokine regulated on activation,
normal T cell expressed and secreted (RANTES) in advanced breast
carcinoma. Cancer Res. 59:4681–4687. 1999.PubMed/NCBI
|
|
30
|
Niwa Y, Akamatsu H, Niwa H, Sumi H, Ozaki
Y and Abe A: Correlation of tissue and plasma RANTES levels with
disease course in patients with breast or cervical cancer. Clin
Cancer Res. 7:285–289. 2001.PubMed/NCBI
|
|
31
|
Zhang GJ and Adachi I: Serum interleukin-6
levels correlate to tumor progression and prognosis in metastatic
breast carcinoma. Anticancer Res. 19:1427–1432. 1999.
|
|
32
|
Salgado R, Junius S, Benoy I, et al:
Circulating interleukin-6 predicts survival in patients with
metastatic breast cancer. Int J Cancer. 103:642–646. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kishimoto T: The biology of interleukin-6.
Blood. 74:1–10. 1989.
|
|
34
|
Jones SA, Scheller J and Rose-John S:
Therapeutic strategies for the clinical blockade of IL-6/gp130
signaling. J Clin Invest. 121:3375–3383. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yen MC, Lin CC, Chen YL, et al: A novel
cancer therapy by skin delivery of indoleamine 2,3-dioxygenase
siRNA. Clin Cancer Res. 15:641–649. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lin CC, Chou CW, Shiau AL, et al:
Therapeutic HER2/Neu DNA vaccine inhibits mouse tumor naturally
overexpressing endogenous neu. Mol Ther. 10:290–301. 2004.
View Article : Google Scholar : PubMed/NCBI
|