Introduction
In developed countries, prostate cancer is the
second most frequently diagnosed cancer and the third most common
cause of mortality from cancer in males. Androgen ablation therapy
is crucial in the treatment of patients with advanced prostate
cancer. However, most patients relapse within 2–3 years after the
treatment. This treatment-resistant prostate cancer, known as
androgen-independent or castration-resistant prostate cancer, is
the final stage of this disease (1–3).
Therefore, identification of the molecules involved and novel
therapeutic strategies targeting these molecules in advanced
prostate cancer are required.
Seven angiopoietin-like proteins (ANGPTLs) have been
identified. These ANGPTLs structurally resemble angiopoietins: they
have a coiled-coil domain at the N-terminus and a fibrinogen-like
domain at the C-terminus. However, ANGPTLs do not bind to the
angiopoietin-specific receptor Tie2 (4–11).
Recently, Aoi et al reported that ANGPTL2 increases
inflammatory carcinogenesis in chemically induced skin squamous
cell carcinoma (12). Additionally,
Endo et al reported that ANGPTL2 expression in lung tumor
cells is highly correlated with the frequency of tumor cell
metastasis (13). Integrin α5β1,
which acts as functional receptor for ANGPTL2 in endothelial cells
and monocytes/macrophages (14,15),
is also expressed in several cancer cells, in which it regulates
tumor cell growth and invasion (16,17).
ANGPTL2 is expressed in certain tumor cell types (18). Cancer cell-derived ANGPTL2 is an
important factor in cancer development.
This study investigated the possible expression and
role of ANGPTLs in human prostate cancer cells. To the best of our
knowledge, this is the first study to demonstrate that a high
ANGPTL2 expression induces androgen-independent and malignant
behavior in human prostate cancer cells. By contrast, decreasing
ANGPTL2 levels in human prostate cancer cells attenuated cell
growth and malignant behavior. Our findings suggest that blocking
ANGPTL2 is useful as a therapeutic strategy against prostate cancer
progression.
Materials and methods
Cell line and culture conditions
The LNCaP, PC-3, DU145 and 22Rv1 human prostate
cancer cell lines were purchased from the American Type Culture
Collection (ATCC; Rockville, MD, USA). These cells were cultured at
37°C in a humidified incubator containing 5% CO2 and 95%
air. LNCaP, DU145 and 22Rv1 cells were cultured in RPMI-1640
(Sigma-Aldrich Corp. St. Louis, MO, USA) supplemented with 15%
fetal bovine serum (Sigma-Aldrich Corp.), 50 μg/ml streptomycin and
50 IU/ml penicillin (Gibco, Grand Island, NY, USA). PC-3 cells were
cultured in RPMI-1640 supplemented with 10% newborn calf serum
(Equitech-Bio Inc., Kerrville, TX, USA), 50 μg/ml streptomycin and
50 IU/ml penicillin.
For androgen [dihydrotestosterone (DHT)] ablation,
an androgen-independent prostate cancer cell line model LNCaP/AI
was cultured in phenol red free RPMI-1640 (Sigma-Aldrich Corp.)
supplemented with 15% charcoal/dextran-treated fetal bovine serum
(HyClone, Logan, UT, USA), 50 μg/ml streptomycin and 50 IU/ml
penicillin for 3 months.
RNA isolation and quantitative
reverse-transcription polymerase chain reaction (RT-qPCR)
Total Ribonucleic acid (RNA) was isolated using
TRIzol reagent (Invitrogen Life Technologies, Carlsbad, CA, USA)
according to the manufacturer’s instructions. Total RNA (1 μg) was
synthesized into cDNA using the ThermoScript RT-PCR System
(Invitrogen Life Technologies) according to the manufacturer’s
instructions. After the reverse transcription reaction,
first-strand cDNA (2 μg) was used for PCR with a
LightCycler® FastStart DNA Master SYBR-Green I reaction
mix (Roche Molecular Biochemicals, Mannheim, Germany) and
QuantiTect Primer Assays (Qiagen Inc., Hilden, Germany) on a
LightCycler system (Roche Diagnostics Corp., Indianapolis, IN,
USA). Each cycle included denaturation at 95°C for 15 sec,
annealing at 55°C for 5 sec and polymerization at 72°C for 10 sec.
The primers used were ANGPTL2 (HS_ANGPTL2_1_SG QuantiTect Primer
Assay; QT00091021) and β-actin (HS_ACTB_1_SG QuantiTect Primer
Assay; QT00095431). Messenger RNA (mRNA) expression quantification
was normalized by β-actin mRNA expression.
Immunohistochemical staining
Cultured cells were washed with phosphate-buffered
saline (PBS), fixed in methanol for 20 min and incubated in 10%
goat normal serum (Nichirei Corp., Tokyo, Japan) for 10 min at
37°C. The cells were incubated in the primary antibodies against
ANGPTL2 at a dilution of 1:500 (Atlas Antibodies AB, Stockholm,
Sweden), integrin α5β1 at a dilution of 1:500 (Biorbyt Ltd.,
Cambridge, UK) at room temperature in PBS with 1% BSA for 60 min.
After incubation with primary antibodies, the secondary antibody
(anti-rabbit; Dako, Kyoto, Japan) was added to PBS with 1% BSA for
30 min. After washing with PBS, the color was developed using the
Dako Cytomation Liquid DAB Substrate Chromogen System (Dako). The
cells were counterstained with hematoxylin.
Prostate cancer specimens were obtained from
prostate cancer patients (n=10) undergoing radical prostatectomy,
with (n=5) and without (n=5) androgen ablation therapy. The study
was approved by the Oita University Ethics Committee and informed
consent was obtained for experimentation with human subjects.
Tissue sections were cut from paraffin-embedded blocks and were
placed on silicone-coated slides. After deparaffinization in xylene
and rehydration using graded alcohol solutions, the sections were
incubated in 0.3% H2O2 for 10 min to
inactivate endogenous peroxidase with subsequent washing with PBS.
To block non-specific binding to sections and to eliminate
non-specific staining, 10% normal goat serum in PBS was applied to
the sections and incubated for 10 min. The slides were then
incubated in the primary antibody against ANGPTL2 at a dilution of
1:250 (Atlas Antibodies AB) at room temperature in PBS with 1% BSA
for 60 min. After incubation with primary antibodies, the secondary
antibody (anti-rabbit; Dako) was added to PBS with 1% BSA for 30
min. After washing with PBS, the color was developed using the Dako
Cytomation Liquid DAB Substrate Chromogen System (Dako). The cells
were counterstained with hematoxylin.
Transient transfection of ANGPTL2
small-interfering RNA (siRNA)
LNCaP and LNCaP/AI cells were transiently
transfected with a ANGPTL2 siRNA duplex [si-ANGPTL2; final
concentration, 60 nmol/l (Qiagen Inc.)] or control siRNA [random
scrambled sequence: si-Scr; final concentration, 60 nmol/l (Qiagen
Inc.)] using Lipofectamine RNAiMAX (Invitrogen Life Technologies)
according to the manufacturer’s instructions. The sequence of the
siRNA against ANGPTL2 generated by Invitrogen was:
5′-AACCTGAGAGCGAGTATTATA(dT)(dT)-3′,
5′-CTCGCGGGTCACGCAGCTCTA(dT)(dT)-3′,
5′-ACCGGCCGTATAGATAATGTA(dT)(dT)-3′,
5′-CAGAATGTCTACAATGCTAAT(dT)(dT)-3′.
Cell proliferation assays
The cells were seeded in 24-well plates at a density
of 5×104 cells/well. The cells were trypsinized,
collected and counted at 24, 48 and 72 h using hemocytometry.
Migration and invasion assays
Cell migration was assessed using a 24-well BioCoat
Control Insert Chamber (BD Biosciences, Franklin Lakes, NJ, USA)
with polycarbonate filters containing 8-μm pores. The cells were
plated at 5×104 cells/well in 0.5 ml of serum-free
medium. The outer chambers were filled with 0.75 ml of media
containing 15% fetal bovine serum. After 48 h, cells migrating to
the undersurface of the filters were counted. The top surface of
the membrane was scrubbed gently with a cotton swab. The cells on
the undersurface were then fixed in 99.8% methanol and stained with
0.05% toluidine blue prior to undergoing a series of washes. The
cells passing to the undersurface of each filter were counted using
a cell counter system (BZ-9000; Keyence Co., Osaka, Japan).
For invasion assays, the control insert chambers
were replaced with BioCoat Matrigel Invasion Chambers (BD
Biosciences) treated with a Matrigel Matrix reconstituted basement
membrane layer.
For migration inhibition analysis by integrin α5β1
immunoneutralization, the outer chambers were filled with 0.75 ml
of media containing ANGPTL2 recombinant protein (Adipogen Corp. San
Diego, CA, USA) of various concentrations (0, 1.0, 2.5 and 5.0
μg/ml) with 25 μg/ml mouse anti-human integrin α5β1 monoclonal
antibody (Millipore Corp. Billerica, MA, USA) or 25 μg/ml control
mouse IgG (Zymed Laboratories Inc., San Francisco, CA, USA).
Protein extraction and western blot
analysis
Proteins were extracted from cell plates with lysis
buffer [50 mmol/l Tris (pH 8.0), 150 mmol/l NaCl, 0.02%
NaN3, 0.1% sodium dodecyl sulfate, 1% NP-40, 0.5% sodium
deoxycholate and 1 mmol/l phenylmethylsulfonyl fluoride] in the
presence of a protease inhibitor cocktail (Roche Applied Science,
Indianapolis, IN, USA). Samples containing equal amounts of protein
(20 μg) were electrophoresed on 4–20% Tris-glycine gels (Tefco
Corp., Tokyo, Japan) and transferred to nitrocellulose membranes.
After blocking with Blocking One-P solution (Nacalai Tesque Inc.,
Kyoto, Japan), the membranes were incubated with mouse monoclonal
antibodies against β-tubulin at a dilution of 1:5,000
(Sigma-Aldrich Corp.) or rabbit polyclonal antibodies against
ANGPTL2 at a dilution of 1:1,000 (Proteintech Group, Chicago, IL,
USA), Caspase-9, Caspase-3, poly adenosine diphosphate-ribose
polymerase (PARP), Bcl-2 at a dilution of 1:1,000 (Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA), Bcl-xL, Bad, E-Cadherin,
N-Cadherin, vimentin, snail, slug at a dilution of 1:1,000 (Cell
Signaling Technology, Inc., Danvers, MA, USA) and integrin α5β1 at
a dilution of 1:1,000 (Biorbyt Ltd.) at 4°C overnight. After
washing with Tween-Tris-buffered saline (T-TBS), the membranes were
incubated with the corresponding secondary antibodies conjugated
with horseradish peroxidase in T-TBS for 1 h at room temperature.
Immunoreactive bands were visualized using a western blotting
detection system (ECL Plus; Amersham Pharmacia Biotech, Little
Chalfont, UK).
Stable ANGPTL2 transfection
LNCaP cells were transfected with ANGPTL2 expression
vector (OriGene Technologies Inc. Rockville, MD, USA) or empty
control vector (Neo; OriGene Technologies Inc.) using Optifect
reagent (Invitrogen Life Technologies) according to the
manufacturer’s instructions. Briefly, LNCaP cells were seeded in
six-well plates at a density of 2×105 per well 24 h
prior to transfection in an FBS-supplemented medium. The cells were
transfected using 4 μg of ANGPTL2 expression vector or Neo and 18
μg Optifect reagent/well and cultured in the presence of 500 μg/ml
geneticin sulfate to obtain stable transfectants.
Statistical analysis
Values were presented as means ± SD. Statistical
analyses were performed using the Student’s t-test. P<0.05 were
considered statistically significant.
Results
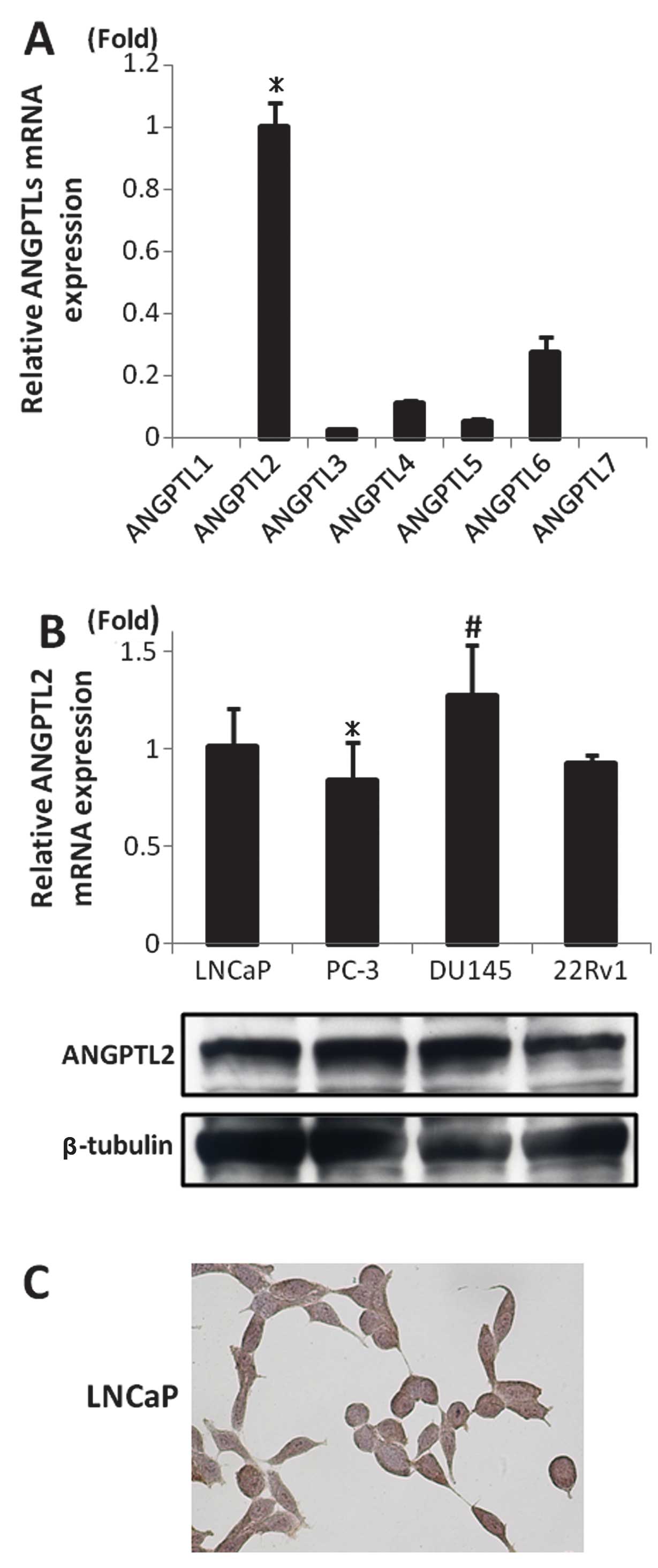
Expression of ANGPTLs in prostate cancer
cell lines
The expression of ANGPTL families in LNCaP cells was
analyzed using RT-PCR. The expression of ANGPTL2 mRNA was increased
significantly (p<0.05; Fig. 1A).
The expression of mRNA and protein of ANGPTL2 was confirmed in the
other prostate cancer cell lines (PC-3, DU145 and 22Rv1) using
western blot analysis and immunohistochemical staining (Fig. 1B and C).
An androgen-independent prostate cancer cell line
model, LNCaP/AI, showed increased cell proliferation, migration and
invasion than the androgen-dependent prostate cancer cell line,
LNCaP (p=0.02; Fig. 2A, p<0.001;
Fig. 2B, p<0.05; Fig. 2C and D). RT-PCR and western blot
analysis revealed that ANGPTL2 mRNA and protein were expressed at
higher levels in LNCaP/AI cells than those in LNCaP cells
(p<0.05; Fig. 2E and F).
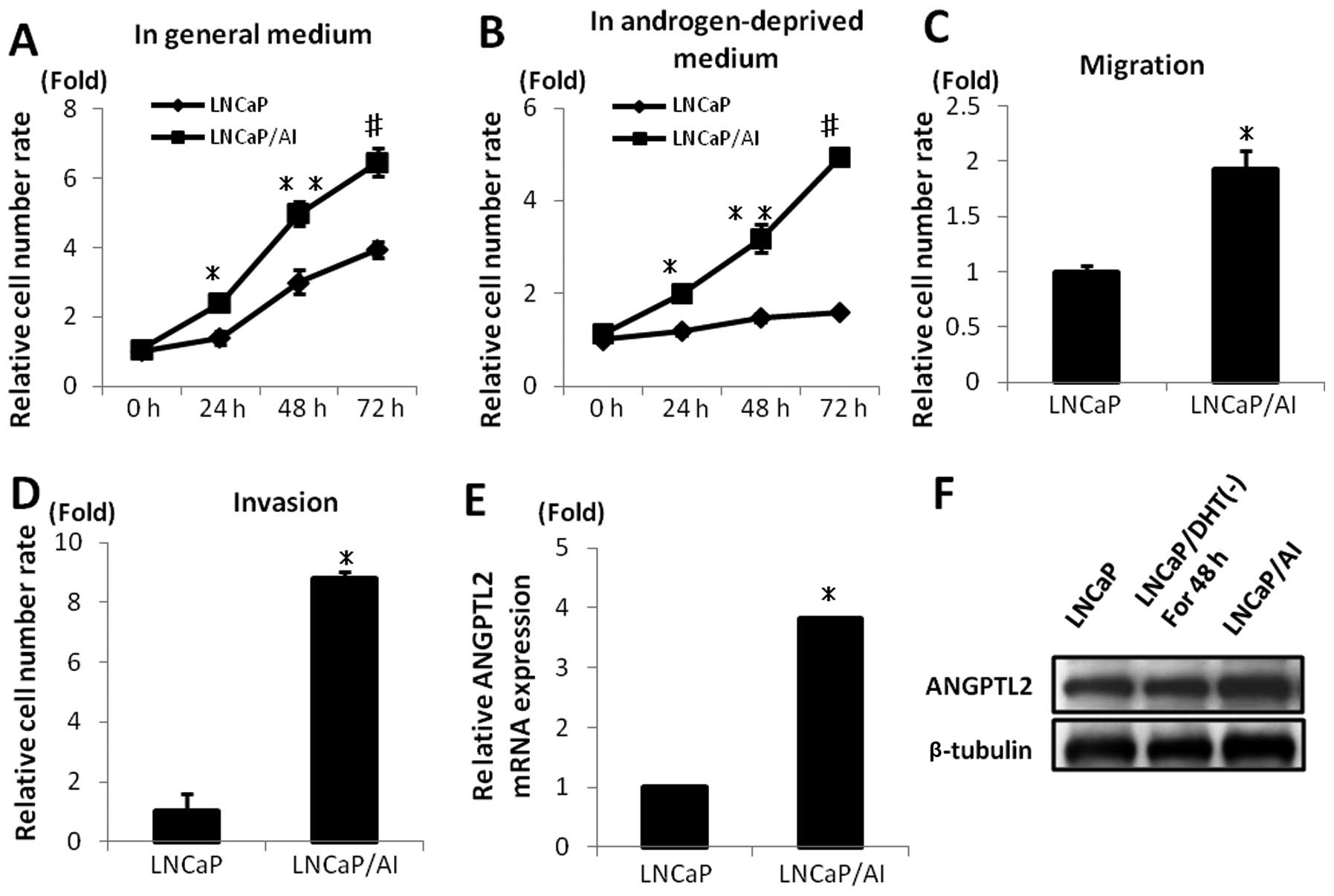 | Figure 2Angiopoietin-like proteins (ANGPTL2)
expression levels in LNCaP/AI cells. (A–D) LNCaP/AI cells showed
significantly increased cell proliferation, migration and invasion
compared with LNCaP cells (A, *p<0.05,
**p<0.03, #p=0.02, B,
*p<0.05, **p<0.01,
#p<0.001, C, D, *p<0.05). (E and F)
Quantitative reverse transcription polymerase chain reaction
(RT-qPCR) and western blot analysis revealed that ANGPTL2 was
significantly more expressed in LNCaP/AI cells than in LNCaP cells
(*p<0.05). β-tubulin.served as the control. |
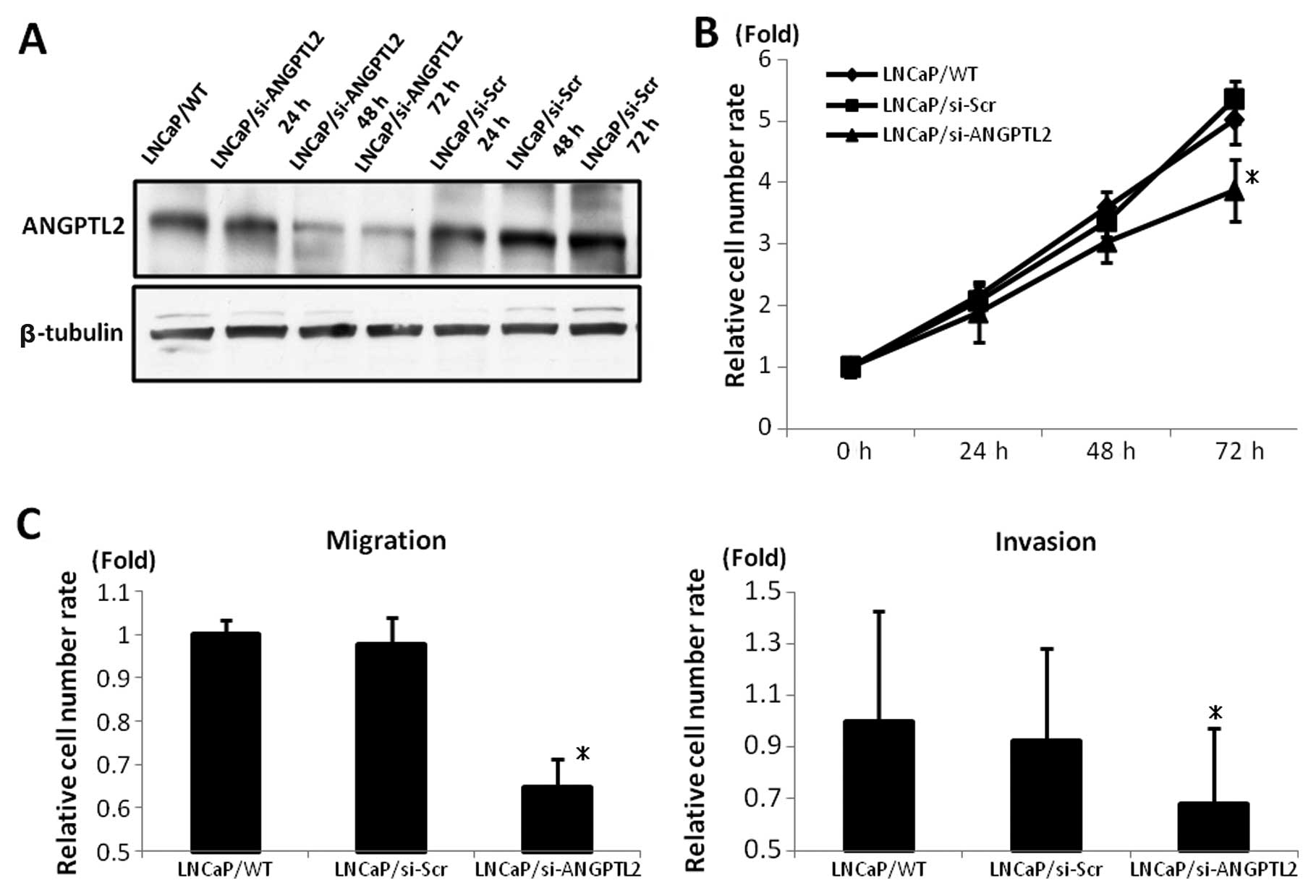
Effects of ANGPTL2 siRNA on cell
proliferation, migration, invasion, apoptosis of LNCaP and LNCaP/AI
cells
ANGPTL2 siRNA reduced the expression of ANGPTL2
protein in LNCaP cells from 24 to 72 h after transfection (Fig. 3A). The proliferation assay revealed
that the cell growth of LNCaP/si-ANGPTL2 was more suppressed than
that of LNCaP/wild-type (WT) or LNCaP/si-Scr (p<0.05; Fig. 3B).
Cell migration and invasion of LNCaP/si-ANGPTL2 were
reduced compared with those of LNCaP/WT and LNCaP/si-Scr
(p<0.05; Fig. 3C).
These results suggested that ANGPTL2 is involved in
the proliferation, migration and invasion of LNCaP cells and that
ANGPTL2 siRNA negatively affected these behaviors.
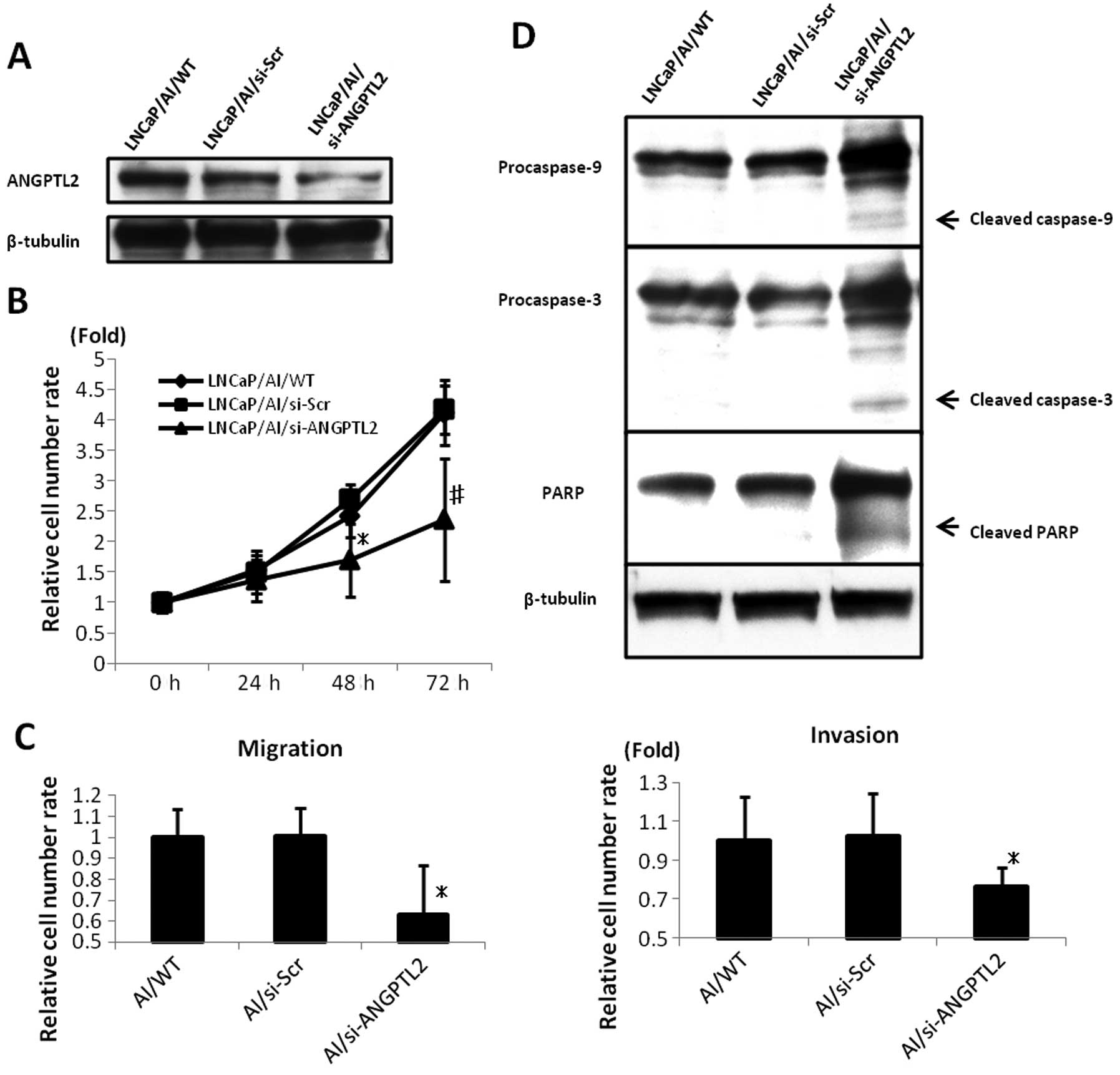
ANGPTL2 siRNA downregulated the ANGPTL2 protein
expression 72 h after transfection in LNCaP/AI cells (Fig. 4A). The LNCaP/AI/si-ANGPTL2 cell
growth was more restricted than that of LNCaP/AI/WT or
LNCaP/AI/si-Scr (p<0.02; Fig.
4B).
Cell migration and invasion were also assessed.
LNCaP/AI/si-ANGPTL2 reduced cell migration and invasion more than
LNCaP/AI/WT or LNCaP/AI/si-Scr (p<0.05; Fig. 4C).
To investigate whether ANGPTL2 siRNA-induced growth
inhibition was triggered by increased apoptosis, we investigated
caspase-9 and caspase-3 activation and subsequent PARP cleavage
using western blot analysis. Treatment with ANGPTL2 siRNA induced
caspase-9 and caspase-3 activation and PARP cleavage in LNCaP/AI
cells (Fig. 4D).
These results suggested that ANGPTL2 induced the
malignant potential in androgen-dependent prostate cancer cell line
LNCaP cells and in androgen-independent prostate cancer cell line
model LNCaP/AI cells.
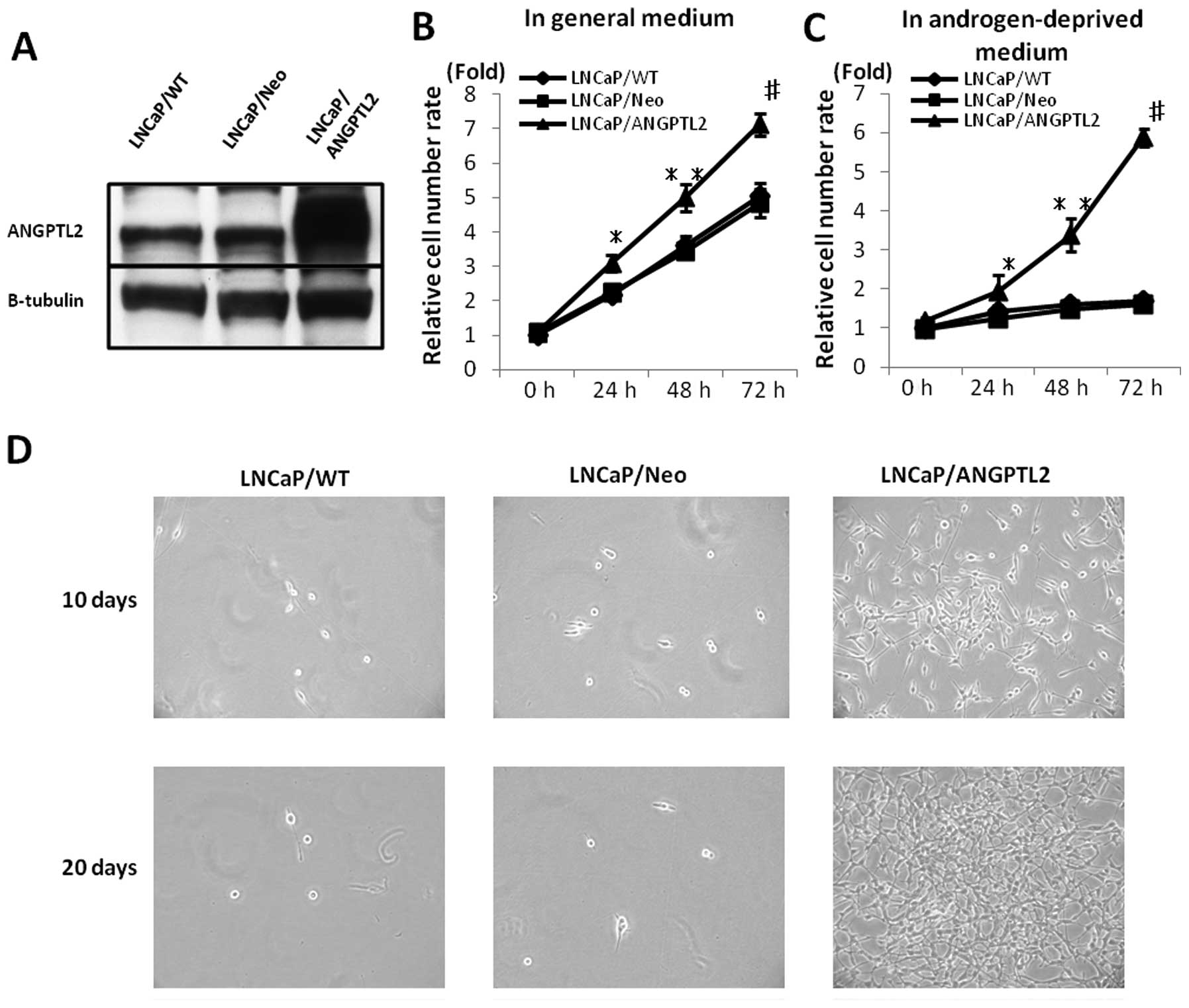
Effects of ANGPTL2 overexpression on cell
proliferation, migration, invasion and acquisition of androgen
independence in LNCaP cells
The results showed that LNCaP cells were stably
transfected with the human ANGPTL2 cDNA expression vector. Clonal
ANGPTL2-overexpressing LNCaP cell lines were established. LNCaP
stably transfected with Neo serving as a control. The human ANGPTL2
cDNA expression vector upregulated ANGPTL2 expression following
transfection into LNCaP cells (Fig.
5A). The ANGPTL2-overexpressing transfectant growth rates were
higher than those of LNCaP/WT or LNCaP/Neo, in the general and
androgen-deprived media. The LNCaP/ANGPTL2 upregulated cell growth
for 20 days longer after placement into androgen-deprived medium
than LNCaP/WT or LNCaP/Neo (p<0.01; Fig. 5B, p<1.0E-07; Fig. 5C and D).
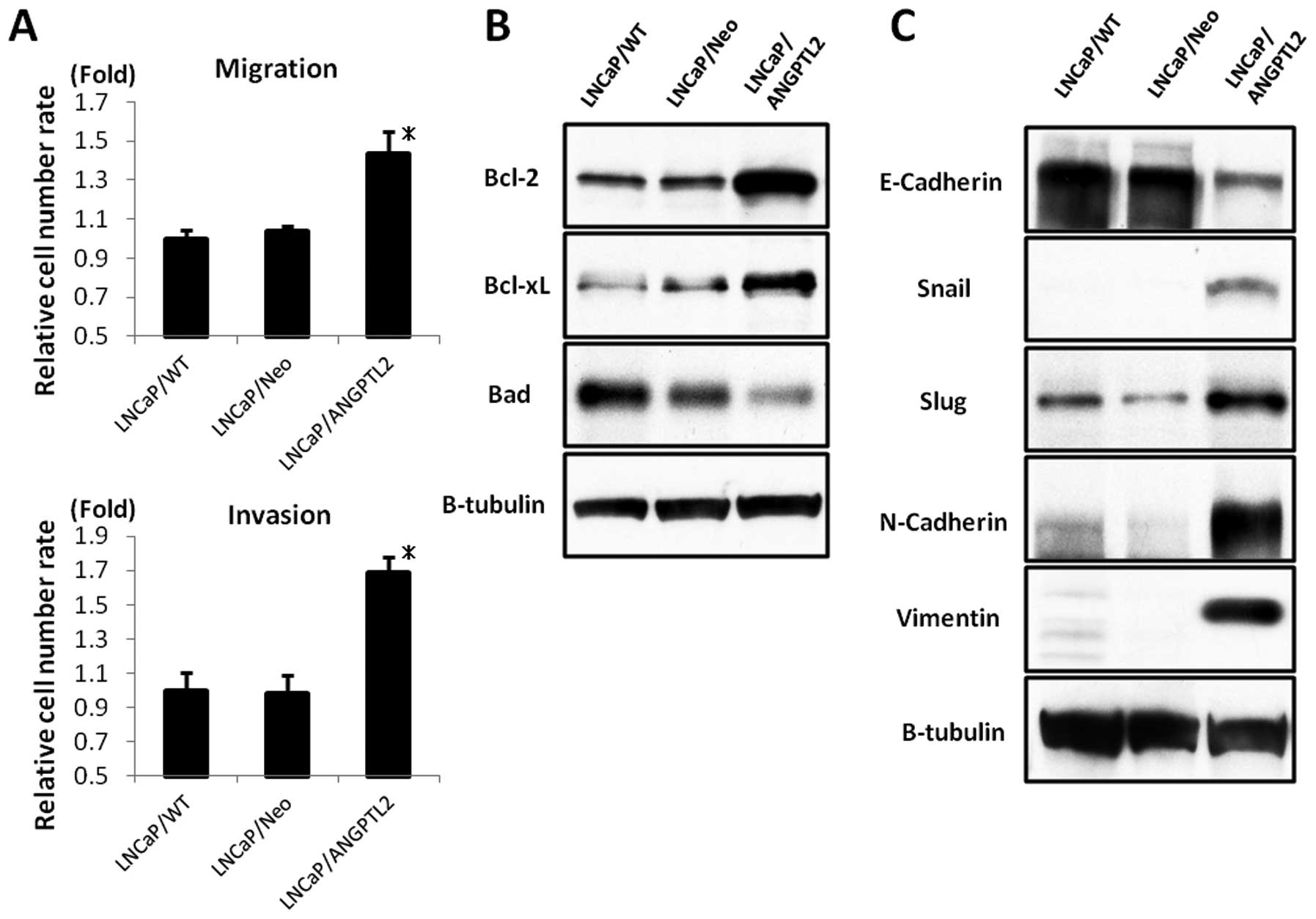
We also analyzed cell migration and invasion. The
ANGPTL2-overexpressing transfectants increased cell migration and
invasion more than that of LNCaP/WT or LNCaP/Neo (p<0.01;
Fig. 6A).
Additionally, we used western blot analysis to
assess expression levels of anti-apoptotic proteins Bcl-2 and
Bcl-xL and pro-apoptotic protein Bad. The ANGPTL2- overexpressing
transfectants increased the expression levels of Bcl-2 and Bcl-xL
proteins and decreased the expression level of Bad protein more
than that of LNCaP/WT or LNCaP/Neo (Fig. 6B).
A critical event in tumor metastasis is a decrease
in cancer cell adhesion through acquisition of mesenchymal
phenotypes and invasive properties. The epithelial-to-mesenchymal
transition (EMT) correlated with ANGPTL2 expression levels. Western
blot analysis revealed that the ANGPTL2-overexpressing
transfectants increased the expression levels of snail, slug,
N-Cadherin and vimentin proteins, as mesenchymal markers, and
decreased the expression level of E-Cadherin protein, as an
epithelial indicator, more than that of LNCaP/WT or LNCaP/Neo
(Fig. 6C).
These results suggested that ANGPTL2 overexpression
induced androgen-independent progression and malignant behavior in
LNCaP cells via escape from apoptosis and promotion of EMT.
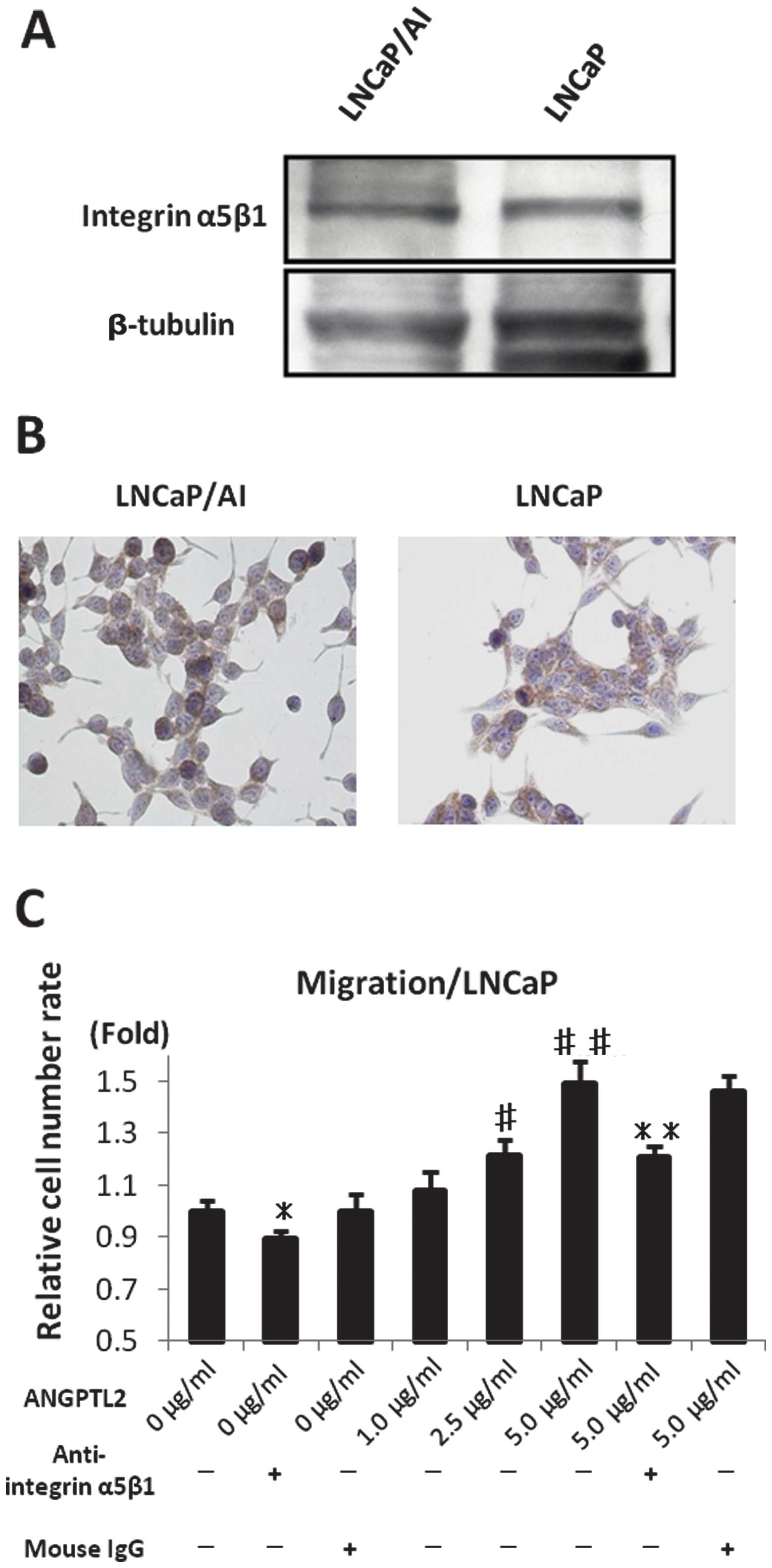
Integrin α5β1 expression levels in LNCaP
and LNCaP/AI cells
Integrin α5β1 is not specific receptor of ANGPTL2,
but a functional receptor of ANGPTL2. The expression of integrin
α5β1 protein in LNCaP and LNCaP/AI cells was confirmed using
western blot analysis and immunohistochemical staining (Fig. 7A and B).
Effects of integrin α5β1
immunoneutralization and ANGPTL2 recombinant protein on cell
migration in LNCaP cells
To assess the effect of integrin α5β1
immunoneutralization in LNCaP cells, we analyzed cell migration.
Cell migration with integrin α5β1 immunoneutralization was more
restricted than in LNCaP/WT or LNCaP/control mouse IgG (p<0.05;
Fig. 7C). To assess the effects of
the autocrine/paracrine action of ANGPTL2, we analyzed cell
migration in LNCaP cells with various concentrations (0, 1.0, 2.5
and 5.0 μg/ml) of ANGPTL2 recombinant protein. The migration rates
increased along with the ANGPTL2 recombinant protein concentration
(p<0.01; Fig. 7C).
These results suggested that ANGPTL2 affected human
prostate cancer cells in an autocrine/paracrine manner via the
integrin α5β1 receptor.
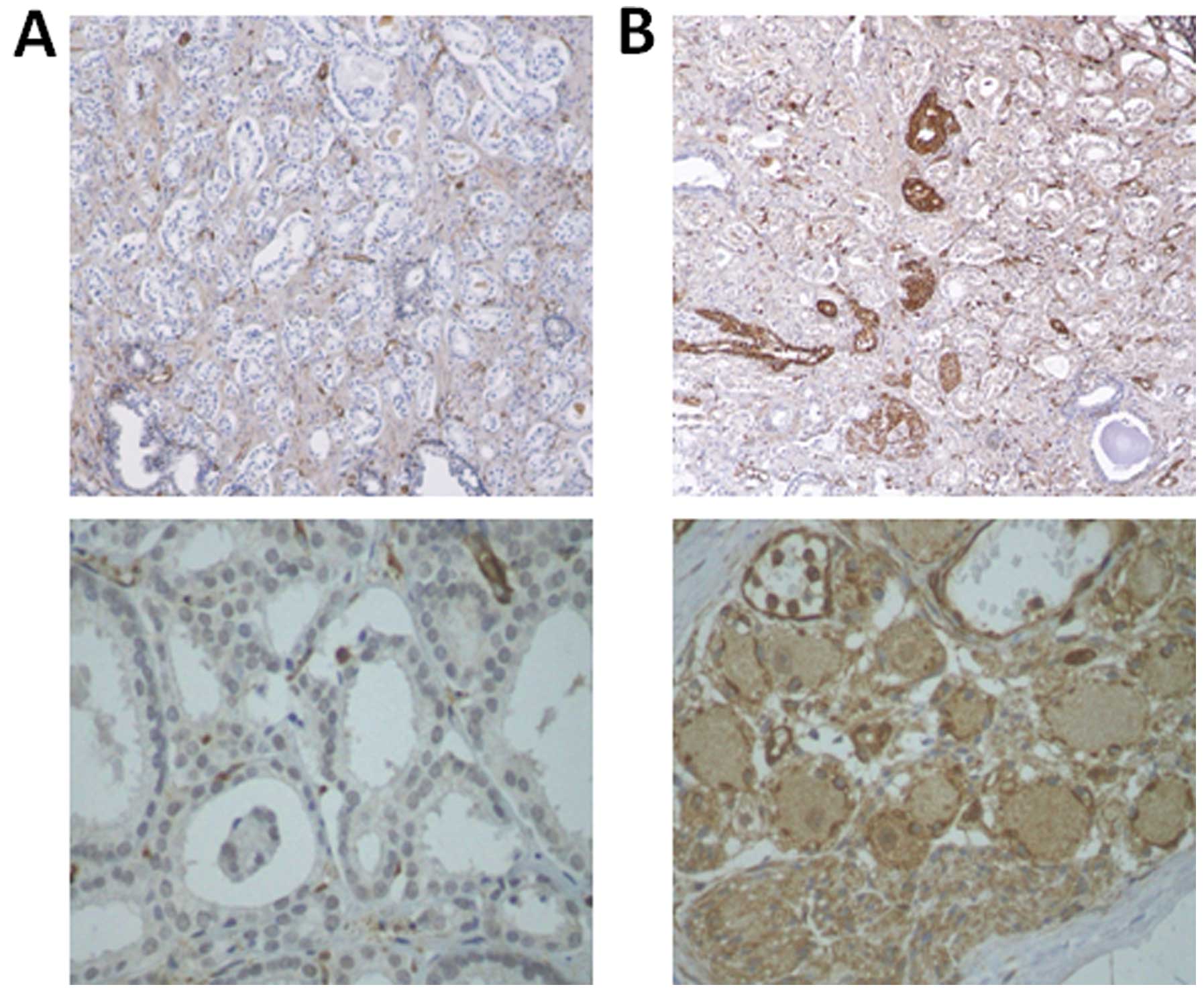
ANGPTL2 expression levels in human
prostate cancer tissues
Immunohistochemical staining revealed that ANGPTL2
was expressed at higher levels in human prostate cancer tissues
after androgen ablation therapy than in human prostate cancer
tissues without androgen ablation therapy (Fig. 8A and B).
These results suggested that androgen ablation
therapy induced more ANGPTL2 expression in human prostate cancer
tissues.
Discussion
Prostate cancer cells require androgens for growth
and survival. Androgen ablation therapy is therefore the gold
standard for advanced prostate cancer. However, androgen-dependent
prostate cancer becomes androgen-independent with long-term
androgen deprivation. Both androgen receptor (AR)-dependent and
AR-independent mechanisms are involved in progression to the
castration-resistant state (19,20).
The role of ANGPTL2 in prostate cancer development has not been
reported in connection with androgen independency and malignant
behavior. Therefore, the aims of this study were i) to define the
biological effects of ANGPTL2 in prostate cancer cells, and ii) to
examine the role of ANGPTL2 as a novel therapeutic target for
prostate cancer.
The results of this study demonstrate that ANGPTL2
expression is strongly induced by androgen deprivation and that it
is highly associated with androgen-independent and malignant
behavior in LNCaP cells in an autocrine and/or paracrine manner via
the integrin α5β1 receptor. The downregulation of ANGPTL2
expression by siRNA inhibits cell growth, migration and invasion of
LNCaP cells and androgen-independent prostate cancer cell line
model LNCaP/AI. Furthermore, stably overexpressed ANGPTL2 promotes
cancer cell growth, migration and invasion, in the general and
androgen-deprived media over the long term via escape from
apoptosis and promotion of EMT. Thus, ANGPTL2 is an effective
growth-promoting factor and an important determinant of the
potential malignancy of androgen-independent prostate cancer cells.
Additionally, we have demonstrated that strong immunostaining of
ANGPTL2 occurred in residual cancer cells in prostate specimens
obtained from patients who had undergone neoadjuvant hormonal
therapy. This finding suggests that ANGPTL2 is involved in the
hormone refractory mechanism in clinical prostate cancer.
ANGPTL2 was found to increase inflammatory
carcinogenesis in a chemically induced skin squamous cell
carcinoma. Furthermore, ANGPTL2 expression in lung tumor cells is
highly correlated with the frequency of tumor cell metastasis
through increased tumor angiogenesis. It was also reported that
ANGPTL2 increases EMT in cancer cells (12,13).
Additionally, ANGPTL2 stimulates the nuclear translocation of
nuclear factor-κB (NF-κB) via integrin α5β1 signaling (14). The NF-κB-dependent expression of
genes is extremely important for anti-apoptotic mechanisms in
cancer cells. Therefore, the activation of NF-κB induces high rates
of cancer cell proliferation via the producton of anti-apoptotic
proteins. The abovementioned studies supported the findings of this
study.
Kikuchi et al reported ANGPTL2 as a putative
tumor suppressor in ovarian cancer (18). In that study, lack of ANGPTL2
immunoreactivity was associated with poorer overall survival in
stage I and II disease, whereas ANGPTL2 positivity was associated
significantly with a poorer survival in stage III and IV disease.
The latter findings are consistent with those of this study,
suggesting that ANGPTL2 is critical for tumor progression and
malignant behavior. However, we did not observe a tumor suppressor
function for ANGPTL2 in human prostate cancer cells. Whether
ANGPTL2 function differs in various tumor cell types remains to be
determined (21–23). Additionally, ANGPTL1 represses lung
cancer cell motility by abrogating the expression of the EMT
mediator slug (24). However, it
has been reported that ANGPTL4 promotes cancer growth and
progression (25–27). The involvement of the ANGPTLs family
in cancer remains unclear and requires further investigation.
In conclusion, ANGPTL2, which is upregulated in
androgen-independent prostate cancer cells and human prostate
cancer tissue after androgen ablation therapy, may be important in
androgen-independent prostate cancer progression in an autocrine
and/or paracrine manner via the integrin α5β1 receptor.
Downregulation of ANGPTL2 by siRNA reduces migration and invasion
and inhibits cancer cell growth. Targeting ANGPTL2 may therefore be
an efficacious therapeutic modality for prostate cancer, especially
androgen-independent prostate cancer.
Acknowledgements
We gratefully acknowledge Ms. Noriko Hamamatsu, Ms.
Yuka Ito and Ms. Saiko Kato for their excellent technical support
of the experiment. The authors thank Fastek (www.fastekjapan.com) for English language
assistance.
References
|
1
|
Damber JE and Aus G: Prostate cancer.
Lancet. 371:1710–1721. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Grönberg H: Prostate cancer epidemiology.
Lancet. 361:1048–1053. 2003. View Article : Google Scholar
|
|
3
|
Scher HI, Buchanan G, Gerald W, et al:
Targeting the androgen receptor: Improving outcomes for
castration-resistant prostate cancer. Endocr Relat Cancer.
11:459–476. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Oike Y and Tabata M: Angiopoietin-like
proteins. Circ J. 73:2192–2197. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dhanabal M, LaRochelle WJ, Jeffers M,
Herrmann J, et al: Angioarretin: an antiangiogenic protein with
tumor-inhibiting properties. Cancer Res. 62:3834–3841.
2002.PubMed/NCBI
|
|
6
|
Kubota Y, Oike Y, Satoh S, et al:
Cooperative interaction of Angiopoietin-like proteins 1 and 2 in
zebrafish vascular development. Proc Natl Acad Sci USA.
102:13502–13507. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Camenisch G, Pisabarro MT, Sherman D, et
al: ANGPTL3 stimulates endothelial cell adhesion and migration via
integrin alpha vbeta 3 and induces blood vessel formation in vivo.
J Biol Chem. 277:17281–17290. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ito Y, Oike Y, Yasunaga K, et al:
Inhibition of angiogenesis and vascular leakiness by
angiopoietin-related protein 4. Cancer Res. 63:6651–6657.
2003.PubMed/NCBI
|
|
9
|
Zeng L, Dai J, Ying K, et al:
Identification of a novel human angiopoietin-like gene expressed
mainly in heart. J Hum Genet. 48:159–162. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Oike Y, Yasunaga K, Ito Y, et al:
Angiopoietin-related growth factor (AGF) promotes epidermal
proliferation, remodeling and regeneration. Proc Natl Acad Sci USA.
100:9494–9499. 2003. View Article : Google Scholar
|
|
11
|
Katoh Y and Katoh M: Comparative
integromics on Angiopoietin family members. Int J Mol Med.
17:1145–1149. 2006.PubMed/NCBI
|
|
12
|
Aoi J, Endo M, Kadomatsu T, et al:
Angiopoietin-like protein 2 is an important facilitator of
inflammatory carcinogenesis and metastasis. Cancer Res.
71:7502–7512. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Endo M, Nakano M, Kadomatsu T, et al:
Tumor cell-derived angiopoietin-like protein ANGPTL2 is a critical
driver of metastasis. Cancer Res. 72:1784–1794. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tabata M, Kadomatsu T, Fukuhara S, et al:
Angiopoietin-like protein 2 promotes chronic adipose tissue
inflammation and obesity-related systemic insulin resistance. Cell
Metab. 10:178–188. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Okada T, Tsukano H, Endo M, et al:
Synoviocyte-derived angiopoietin-like protein 2 contributes to
synovial chronic inflammation in rheumatoid arthritis. Am J Pathol.
176:2309–2319. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Avraamidea CJ, Garmy-Susini B and Varner
JA: Integrins in angiogenesis and lymphangiogenesis. Nat Rev
Cancer. 8:604–617. 2008. View
Article : Google Scholar
|
|
17
|
Desgrosellier JS and Cheresh DA: Integrins
in cancer: biological implications and therapeutic opportunities.
Nat Rev Cancer. 10:9–22. 2010. View
Article : Google Scholar
|
|
18
|
Kikuchi R, Tsuda H, Kozaki K, et al:
Frequent inactivation of a putative tumor suppressor,
angiopoietin-like protein2, in ovarian cancer. Cancer Res.
68:5067–5075. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Saraon P and Jarvi K: Molecular
alterations during progression of prostate cancer to androgen
independence. Clin Chem. 57:1366–1375. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Schröder FH: Progress in understanding
androgen-independent prostate cancer (AIPC): a review of potential
endocrine-mediated mechanisms. Eur Urol. 53:1129–1173. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Aoi J, Endo M, Kadomatsu T, et al:
Angiopoietin-like protein 2 accelerates carcinogenesis by
activating chronic inflammation and oxidative stress. Mol Cancer
Res. 12:239–249. 2014. View Article : Google Scholar
|
|
22
|
Endo M, Yamamoto Y, Masuda T, et al: Serum
ANGPTL2 levels reflect clinical features of breast cancer patients:
implications for the pathogenesis of breast cancer metastasis. Int
J Biol Markers. 29:239–245. 2014.
|
|
23
|
Odagiri H, Kadomatsu T, Endo M, et al: The
secreted protein ANGPTL2 promotes metastasis of osteosarcoma cells
through integrin α5β1, p38 MAPK and matrix metalloproteinases. Sci
Signal. 7:ra72014. View Article : Google Scholar
|
|
24
|
Kuo TC, Tan CT, Chang YW, et al:
Angiopoetin-like protein 1 suppresses SLUG to inhibit cancer cell
motility. J Clin Invest. 123:1082–1095. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tan MJ, Teo Z, Sng MK, et al: Emerging
Roles of Angiopoietin-like 4 in human cancer. Mol Cancer Res.
10:677–688. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li H, Ge C, Zhao F, et al:
Hypoxia-inducible factor 1 alpha-activated angiopoietin-like
protein 4 contributes to tumor metastasis via vascular cell
adhesion molecule-1/integrin β1 signaling in human hepatocellular
carcinoma. Hepatology. 54:910–919. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Okochi-Takada E, Hattori N, Tsukamoto T,
et al: ANGPTL4 is a secreted tumor suppressor that inhibits
angiogenesis. Oncogene. 24:2273–2278. 2014. View Article : Google Scholar
|