Introduction
Endothelin (ET) was first identified as a
vasoconstrictive 21-amino acid S-peptide derived from vascular
endothelial cells. It is currently known to exert diverse
biological effects on a wide variety of tissues and cell types
through its own receptors (1,2). Three
ETs (ET-1, ET-2 and ET-3), 2 G-protein-coupled receptors (ETRA and
ETRB) and 2 ET-converting enzymes (ECE-1 and ECE-2) comprise the ET
axis (2). The ET-1 gene contains 5
exons located on chromosome 6, and expresses a 212-amino acid
precursor, preproendothelin-1 (PPET-1). PPET-1 is converted by
protease to form an intermediate peptide consisting of 38 amino
acids, big-ET-1, which is further cleaved by ECE-1 or ECE-2 to
release the mature form of ET-1 (21 amino acids) (3,4).
An increase in ET-1 expression has been detected in
various malignancies including lung, prostate, colorectal, liver,
breast and ovarian cancers, as well as melanomas, and may
contribute to cell growth, metastasis and angiogenesis, and
suppression of apoptosis (4–11).
ET-1 typically induces proliferation through the ETRA receptor, and
decreased ETRB expression has frequently been implicated in the
pathogenesis of esophageal squamous cell carcinoma and
nasopharyngeal carcinoma, as well as prostate and ovarian cancers
(12). The promoter regulatory
region of the ETRB gene contains a 5′ CpG island which is commonly
(70%) methylated, suggesting that ETRB down-regulation occurs
through DNA hypermethylation in human carcinomas (13–17).
In gastric cancer, a high level of ET-1 has been detected in
plasma, which may be a reliable marker for predicting recurrence
(18,19). Stow et al (20) summarized previous studies and
revealed that ET-1 transcription can be enhanced using numerous
physical and chemical stimuli such as phorbol esters, thrombin,
angiotensin II and insulin, by activating signaling transduction
pathways including the protein kinase C (PKC) and JUN-terminal
kinase (JNK) pathways. In addition, GATA family transcription
factors, β-catenin, transforming growth factor β (TGFβ) and
hypoxia-inducible factor-1 (HIF-1) can enhance ET-1 transcriptional
activity by modulating its promoter (8,12).
Therefore, unbalanced stimulators and impaired signaling that
frequently lead to ET-1 overexpression trigger the activation of
the ET axis, and promote cancer cell progression through autocrine
and paracrine signaling. However, the detailed functions and
regulatory mechanisms of ET-1 in gastric cancer remain unknown. The
number of studies has recently been increasing regarding small
non-coding short RNA microRNAs (miRNAs) in cancer development
through post-transcriptional regulation of the expression of
oncogenes or tumor-suppressor genes. miRNAs are deregulated in
numerous disease states, particularly in cancer, making them novel
molecular indicators that are critical for early diagnosis,
prognosis, and therapy. Previous studies have demonstrated several
instances of abnormal miRNA expression in gastric cancer cells
compared with adjacent normal cells (21,22).
These dysfunctional miRNAs may disrupt the balance of their target
genes, resulting in the downregulation of tumor-suppressor genes
and oncogene overexpression in gastric cancer.
Previous studies have revealed that miRNAs play a
critical role in modulating ET-1 expression by targeting the
3′untranslated region (3′UTR), including miR-1, miR-125a, miR-125b,
miR-155 and miR-199 (23–28). To date, the biological function and
regulating mechanisms of ET-1 by miRNAs in gastric cancer remain
unknown. In the present study, we demonstrated that ET-1 plays an
oncogenic role in gastric cancer growth, and identified several
potential target sites for miRNA candidates harbored within the
3′UTR of ET-1 using a bioinformatics approach. We identified 5
dysfunctional miRNAs that may contribute to ET-1 overexpression in
gastric cancer, to provide a potential approach and novel targets
for therapy.
Materials and methods
Cell line and nuclear acid
extraction
Five human gastric cancer cell lines, AGS, AZ-521,
HR, NUGC and TSGH, were obtained from the American Type Culture
Collection, and maintained in Dulbecco’s modified Eagle’s medium
(DMEM) supplemented with 10% inactivated fetal bovine serum (FBS;
Invitrogen, Carlsbad, CA, USA). After 5-Aza-dC (10 μM) treatment
for 4 days, RNA was extracted using TRIzol reagent (Invitrogen),
according to the manufacturer’s protocol. After RNA extraction, the
interface was subjected to DNA extraction with a back extraction
buffer (4 M guanidine thiocyanate, 50 mM sodium citrate and 1 M
Tris). Genomic DNA was then precipitated with EtOH.
Constructed reporter of ET-1 and the
miRNA expression vector
ET-1 3′UTR was amplified from AGS cDNA by using the
specific primer. The amplified ET-1 3′UTR was cloned into the
pmiR-RB-Report vector, between the XhoI and NotI
restriction sites. The primary miRNA transcripts were amplified
from the genomic DNA of AGS cells by using miR-specific forward and
reverse primers. The amplified PCR products were then cloned into
the pLKO vector, between the AgeI and EcoRI
restriction sites. All constructs were further confirmed through
direct sequencing. The individual primers used in the present study
are shown in Table I.
 | Table IPrimer sequences. |
Table I
Primer sequences.
| Constructed
primers | Sequences |
|---|
| ET-1-3′UTR-F |
5′-CCGCTCGAGCAGACCTTCGGGGCCTGTCTGAAG-3′ |
| ET-1-3′UTR-R |
5′-GAATGCGGCCGCACAGTAAGGAAAAAAATATTTATTTTC-3′ |
| miR-1-1-F |
5′-TCTACCGGTGGACACCAGGCAGCAGTGGC-3′ |
| miR-1-1-R |
5′-TCTGAATTCACAATGCTGGCGGGGACACG-3′ |
| let-7b-F |
5′-TCTACCGGTGGGCCTCTGCCTGTGGAGGA-3′ |
| let-7b-R |
5′-TCTGAATTCTCACTGAGGTAGGGGGCGGC-3′ |
| let-7c-F |
5′-TCTACCGGTTGGCAGGTTAGATGGTCAGAAGACA-3′ |
| let-7c-R |
5′-ACCTTCTTGCACACAAATTGGCTCA-3′ |
| miR-33a-F |
5′-TCTACCGGTCCCATAGCCTCTGTAAGCCC-3′ |
| miR-33a-R |
5′-TCTGAATTCGCTAAGGACATGTTCCCCGT-3′ |
| miR-101-F |
5′-TCTACCGGTTGCCTCCTCACGTCTCCAACCA-3′ |
| miR-101-R |
5′-TCTGAATTCTGGCTGCACCAACAACTACCCC-3′ |
| miR-125a-F |
5′-TCTACCGGTTTCTGTCCTTGTCCCTGCATC-3′ |
| miR-125a-R |
5′-TCTGAATTCCCATCGTGTGGGTCTCAAGG-3′ |
| miR-125b-F |
5′-TCTACCGGTTGCGCCCCCAGATACTGCGT-3′ |
| miR-125b-R |
5′-TCTGAATTCGGGGCATGCTGGGACTTCAGC-3′ |
| miR-134-F |
5′-TCTACCGGTCGGGCCATGGACAATGCGCT-3′ |
| miR-134-R |
5′-TCTGAATTCAGGGGCTGCCAAGGCTGACT-3′ |
| miR-135a-F |
5′-TCTACCGGTTCTTGTTTCCCGGTCCTTGT-3′ |
| miR-135a-R |
5′-TCTGAATTCACCCGACTGGTGGGCTATTA-3′ |
| miR-135b-F |
5′-TCTACCGGTTTTATGGCCAGGAAGCCACC-3′ |
| miR-135b-R |
5′-TCTGAATTCCAGCACACCCTGAAGGTCTC-3′ |
| miR-141-F |
5′-TCTACCGGTGGACACAATGGGCCCCAGCC-3′ |
| miR-141-R |
5′-TCTGAATTCAAAGCAGACGTCGCAGCCCC-3′ |
| miR-144-F |
5′-TCTACCGGTACACTGGCCCTGGGTCCCTA-3′ |
| miR-144-R |
5′-TCTGAATTCTGCCCTGGCAGTCAGTAGGTT-3′ |
| miR-203-F |
5′-TCTACCGGTGTCGGGGGCTCCTCTCTCCG-3′ |
| miR-203-R |
5′-TCTGAATTCCACCGCCAGTTCCTCGCTGG-3′ |
| miR-224-F |
5′-TCTACCGGTGCAGAGGGCTGGGCTACCTT-3′ |
| miR-224-R |
5′-TCTGAATTCCCCAGGGCCCAACTGGAAGAGT-3′ |
| miR-340-F |
5′-TCTACCGGTTCCAGCTTGAGTCTTCAAGAG-3′ |
| miR-340-R |
5′-TCTGAATTCAGAGTTGTGATCAGTAAATTAGA-3′ |
| miR-379-F |
5′-TCTACCGGTGCCTGCTTCCAATGCCAAAT-3′ |
| miR-379-R |
5′-TCTGAATTCGCCCAAGTTGCATCACTTCC-3′ |
| miR-410-F |
5′-TCTACCGGTGCCCTTTTGAGGGTAGGAGC-3′ |
| miR-410-R |
5′-TCTGAATTCAATGATTCAATGGCGGGGGT-3′ |
| miR-425-F |
5′-TCTACCGGTGTGCCCCTGACCCCCAGACA-3′ |
| miR-425-R |
5′-TCTGAATTCAGCAGGGAAACCCAGGGGCA-3′ |
| miR-494-F |
5′-TCTACCGGTACCGTCAGGAAAGCTCCAAT-3′ |
| miR-494-R |
5′-TCTGAATTCTCAGGAACAGGAAGTGCCTC-3′ |
| miR-495-F |
5′-TCTACCGGTCGCCTCTGCTCAGTGTCAGCC-3′ |
| miR-495-R |
5′-TCTGAATTCTCAGGGTCCCGTCGGGGATG-3′ |
| miR-590-F |
5′-TCTACCGGTGAACGTCAGCACCCTCCCCCCA-3′ |
| miR-590-R |
5′-TCTGAATTCTTGAGCGCCAGTGGCCAAGC-3′ |
| miR-873-F |
5′-TCTACCGGTGTGACCAGTGTCTGGGATGC-3′ |
| miR-873-R |
5′-TCTGAATTCCCTTGGTGGGATTCAACACCT-3′ |
| Gene-specific
primer |
| miR-1-RT |
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGATACATAC-3′ |
| miR-1-GSF |
5′-CGGCGGTGGAATGTAAAGAAGT-3′ |
Luciferase reporter assay
HEK293T cells (2.5×104 cells/well) were
seeded in a 24-well plate and cotransfected with 0.3 μg of either
the control vector or various miRNA constructs in conjunction with
0.1 μg of the ET-1-3′UTR reporter construct (pmiR-RB-REPORT™;
RiboBio) by using the Lipofectamine 2000 reagent (Invitrogen).
After 48 h, the luciferase activity was measured using the Dual-Glo
Luciferase Assay System kit (Promega, Madison, WI, USA). Luciferase
readings were corrected for background, and firefly luciferase
values were normalized to Renilla values to control for
transfection efficiency. All of the assays were performed in
triplicate in 3 independent experiments.
Methylation status of CpG islands
The genomic DNA of 5 gastric cancer cell lines, and
normal and tumor tissue pairs from 50 gastric cancer patients were
subjected to bisulfite conversion using the EZ DNA Methylation-Gold
kit (Zymo Research Corporation, Orange, CA, USA). The bisulfite
conversion reaction was conducted at 98°C for 10 min, and then at
64°C for 2.5 h, with a final incubation at 4°C for up to 20 h in a
PCR thermocycler. Then, the modified genomic DNA was used for the
methylation analysis of CpG25 and CpG81 of the mir-1-1 upstream
using the specific methylation primers. The PCR conditions were as
follows: 94°C for 10 min, followed by 35 cycles of 94°C for 1 min,
60°C for 1 min, and 72°C for 30 sec, with a final extension at 72°C
for 10 min using a PCR thermocycler and HotStart Taq DNA
polymerase (Qiagen, Hilden, Germany). The methylation status of the
genomic DNA of individual samples was also examined with the
BstUI digestion (New England Biolabs, Beverly, MA, USA). The
digested PCR fragments were then separated on 2% agarose gel.
Detailed information is provided in a previous study (29). The individual primers used in the
present study were as follows: CpG25-F,
5′-GGAGGGGTAGGATAGTAGTTTGAGT-3′ and CpG25-R,
5′-AAAAAAACCTAACCTAAAAAACCAAAA-3′; CpG81-F, 5 ′-G G T G AG T T T T
G T T TAG T T TAT TAT TAT-3 ′ and CpG81-R,
5′-ATCAAAATTCCTACCTCCCAACTA-3′.
Stem-loop RT-PCR of miR-1
Total RNA (2 μg) was reverse-transcribed with
miR-1-specific stem-loop RT primers using SuperScript III Reverse
Transcriptase (Invitrogen). The reaction was performed under the
following incubation conditions: 30 min at 16°C, followed by 20°C
for 30 sec, 42°C for 30 sec and 50°C for 1 sec, totaling 50 cycles.
The enzyme was subsequently inactivated by incubation at 85°C for 5
min. Afterward, cDNA was diluted 10X for subsequent real-time PCR
analysis using the miR-1-GSF primer. Gene expression was detected
using a SYBR-Green I assay (Applied Biosystems, Foster City, CA,
USA). The U6 expression was used as the internal control, and the
expression levels of miR-1 were normalized to that of U6 (ΔCt =
CtmiR-1 - CtU6).
Ectopic expression of miRNA
candidates
Stable or transient gastric cell cultures expressing
miR-1 candidates were generated using the transfected gastric cells
with pLKO-pre-miRNA by Lipofectamine 2000. After 24 h of
transfection, the stably expressed miRNA cells were generated by
puromycin selection for 14 days. The shLuc targeting the luciferase
gene provided puromycin resistance as the control. The expression
levels of miRNAs were confirmed using real-time PCR.
Expression levels of ET axis genes and
miRNAs were analyzed using The Cancer Genome Atlas (TCGA) data
The TCGA project collects RNA transcriptome and
small RNA transcriptome data, which are obtained using the
next-generation sequencing approach from various types of human
cancers. For the present study, we downloaded all level-3
expression data for gastric cancer from the TCGA data portal
(https://tcga-data.nci.nih.gov/tcga/dataAccessMatrix.htm).
We used 29 tumor samples and 29 normal samples to analyze the
expression levels of each miRNA and ET axis-related gene.
Cell proliferation assay
For cell proliferation analysis, we seeded 3,000
living AGS cells or AGS cells stably expressing miR-1 in 96-well
plates. After exogenous ET-1 stimulation, we determined cell growth
at 0, 1, 2, 3 and 4 days using WST-1 (Roche, Mannhein, Germany).
All experiments were repeated 3 times.
Results
ET-1 is upregulated in gastric
cancer
We first assessed the expression pattern of the ET
axis genes (ET-1, ET-2, ET-2, ETRA, ETRB, ECE-1 and ECE-2) in
gastric cancer by analyzing data from the database of TCGA. We
obtained RNA transcriptome data on cancer tissues and the
corresponding adjacent normal tissues from information on 29
gastric cancer patients. As shown in Fig. 1A, the expression levels of ET-1,
ECE-1 and ECE-2 were significantly increased in gastric cancer
compared to corresponding normal tissues (P<0.001). Conversely,
ET-2 expression was decreased significantly in gastric cancer
(P<0.05). We further examined the expression levels of ET-1 in 3
gastric cancer cell lines using the RT-PCR approach. As shown in
Fig. 1B, we detected a greater
abundance of ET-1 RNA in the AZ-521 and HR cells, and low
expression in AGS cells. To investigate the putative function of
ET-1 on gastric cancer, we further treated AGS cells with ET-1 (10
nM) for 4 days. We evaluated the potential influence of ET-1 on
gastric cancer cell proliferation through WST-1 for 96 h. As shown
in Fig. 1C, cell proliferation in
the ET-1 treatment groups was significantly increased compared with
that in the control groups. These results demonstrate that ET-1
plays an oncogenic role in promoting the proliferation of gastric
cancer cells. However, the mechanism that leads to ET-1
dysregulation in gastric cancer remains unclear. To further
investigate the underlying mechanisms of ET-1 dysfunction in
gastric cancer, we focused on the post-transcriptional regulation
of ET-1 expression by miRNAs during the progression of gastric
cancer.
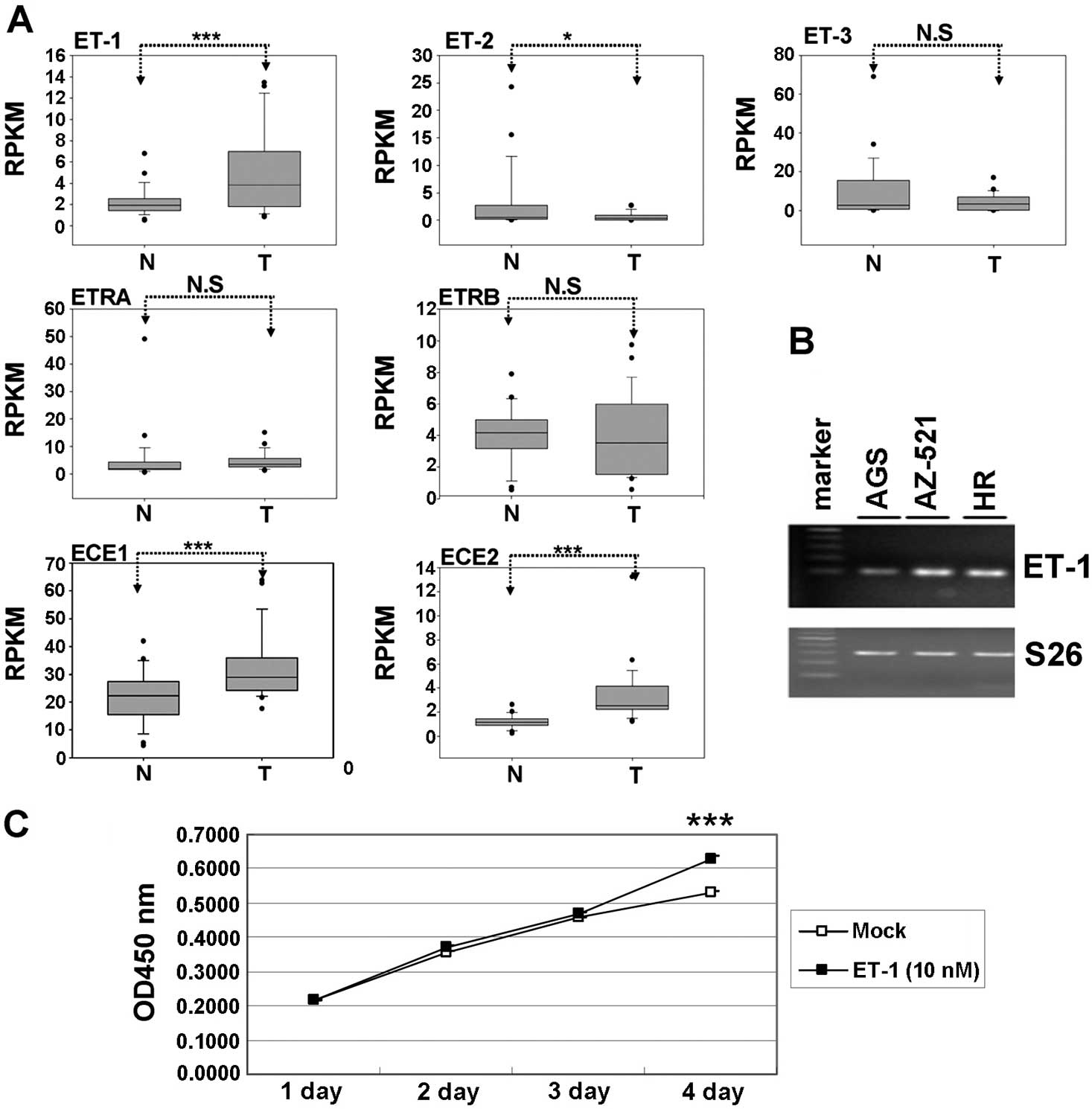 | Figure 1Expression levels and function of the
endothelin axis in gastric cancer cell lines. (A) The expression
levels of ET-1, ET-2, ET-3, ETRA, ETRB, ECE-1 and ECE-2 were
examined between gastric cancer and adjacent normal from 29
patients by analyzing the TCGA database. The expression levels of
the genes were presented in reads per kilobase per million reads
(RPKM). The expression level between the tumor and normal cells was
evaluated by conducting paired t-tests (P<0.05 was considered
significant; NS, non-significant; *P<0.05,
** P<0.01, ***P<0.001). (B) The mRNA
level of ET-1 was assessed in 3 human gastric cancer cell lines.
(C) Exogenous ET-1 stimulation enhanced AGS cell proliferation.
ET-1, endothelin-1; ETRA and ETRB, G-protein-coupled receptors;
ECE-1 and ECE-2, ET-converting enzymes; TCGA, The Cancer Genome
Atlas. |
Discovery of putative miRNAs that
regulate ET-1 genes
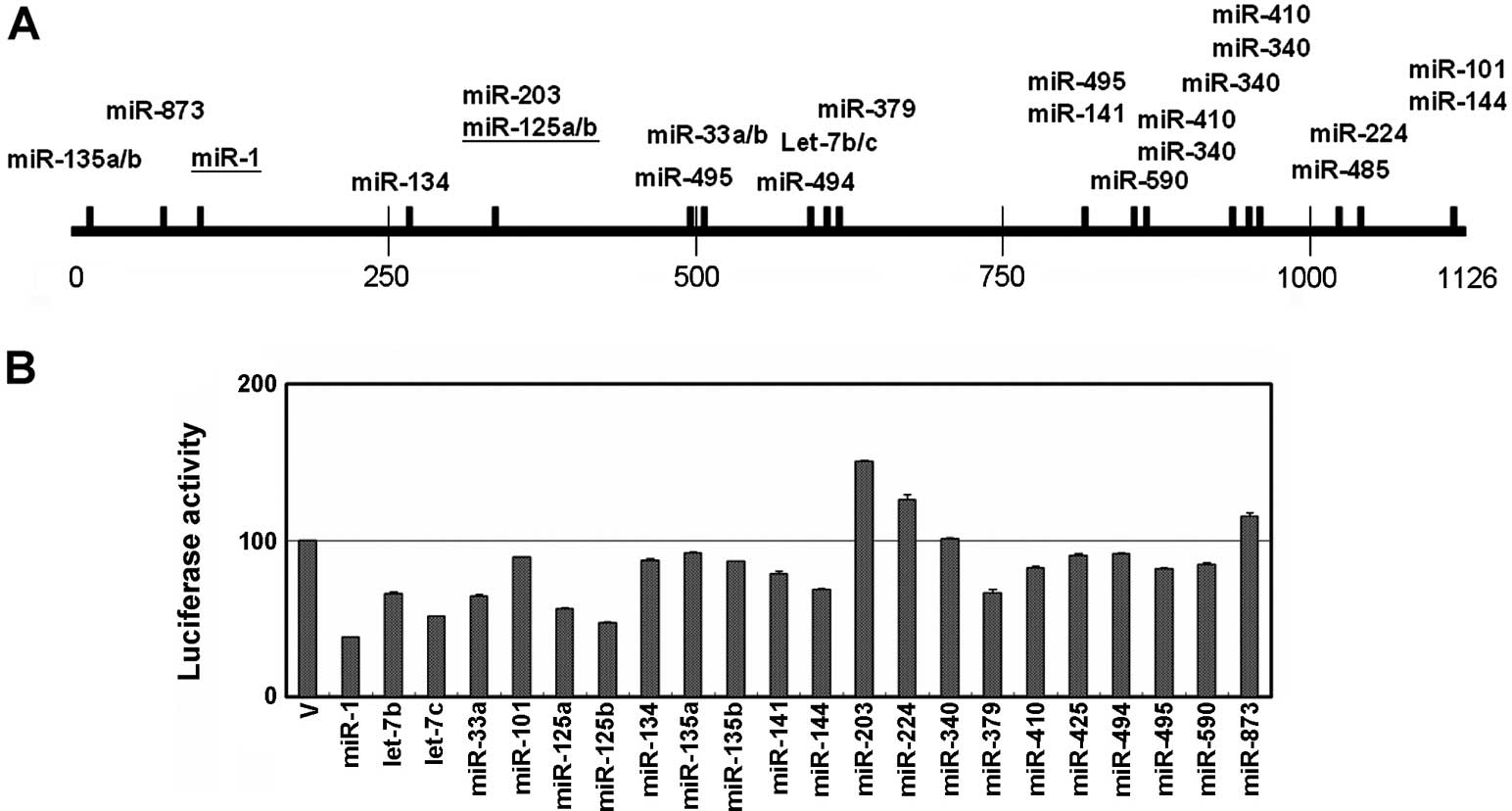
Using a prediction program available on
microRNA.org, we predicted that several miRNA candidates could
suppress ET-1 expression by targeting its 3′UTR sequence. Among
them, we selected 22 miRNA candidates, and constructed their
pre-miRNA sequence into a pKLO expression vector (Fig. 2A). In addition, we cloned the full
length of the 3′UTR (i.e., 1,126 bp) of ET-1 into the
pmiR-RB-Report vector. Due to the fact that the HEK293T cells have
a high transfection efficiency, we conducted a luciferase assay
using HEK293T cells. As shown in Fig.
2B, miR-1, let-7c and miR-125b substantially inhibited
luciferase activity by >50% compared to the control group after
transfection for 48 h. Fifteen miRNA candidates slightly suppressed
the luciferase activity, including let-7b, miR-33a, miR-101,
miR-125a, miR-134, miR-135a/b, miR-141, miR-144, miR-379, miR-410,
miR-425, miR-494, miR-495 and miR-590. Consistent with previous
studies, our data indicated that miR-1 and miR-125a/b both
suppressed ET-1 expression through post-transcriptional regulation.
After using a reporter assay, we found that 18 miRNA candidates may
be involved in ET-1 post-translational regulation through the
targeting of its 3′UTR sequence.
Expression level of miRNA candidates from
the TCGA database
After completing the reporter assay, we selected 18
miRNA candidates to further examine their expression levels in
gastric cancer by analyzing data from the TCGA database. As shown
in Fig. 3A, we identified 6 miRNAs
(miR-134, miR-135b, miR-141, miR-425, miR-494 and miR-590) and 5
miRNAs (miR-1, let-7c, miR-101, miR-125a and miR-144) that were
significantly upregulated and downregulated, respectively, in
gastric cancer. Since the expression level of ET-1 was upregulated
in the gastric cancer cells compared with the corresponding normal
cells, miRNA candidate expression should decrease in gastric
cancer. According to their expression levels in gastric cancer, we
selected 6 miRNAs (miR-1, miR-101, miR-125a/b, miR-144 and let-7c)
for further confirmation in the AGS cells. These miRNA candidates
were ectopically expressed in the AGS cells, and let-7c, miR-1,
miR-101, miR-125a/b and miR-144 were found to significantly
suppress luciferase activity (Fig.
3B). In addition, we found that the endogenous protein levels
of ET-1 were inhibited by let-7c, miR-1, miR-101, miR-125a and
miR-144 overexpression (Fig.
3C).
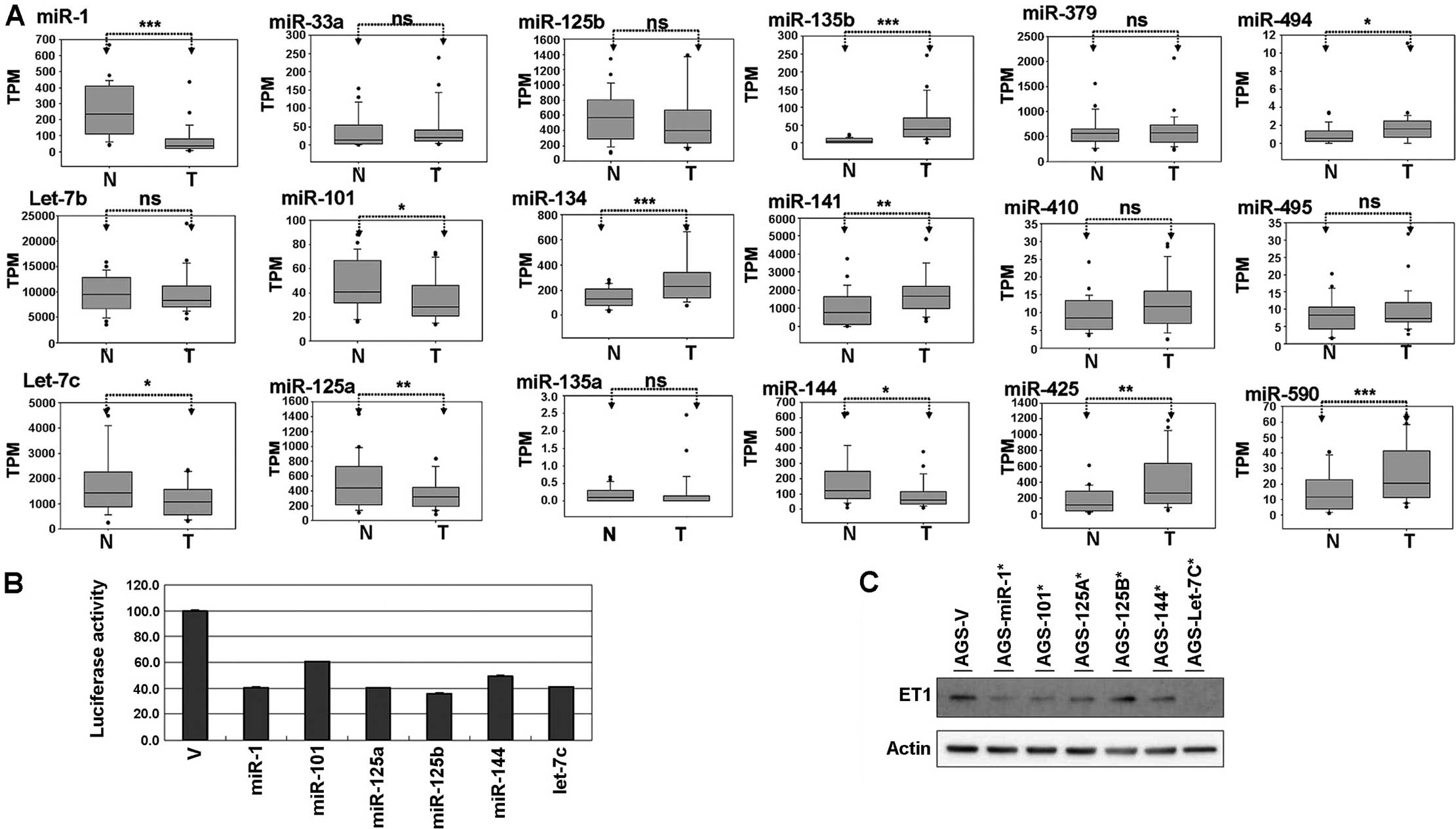 | Figure 3Suppression of ET-1 expression by
miRNAs in gastric cancer cells. (A) Expression levels of the miRNA
candidates in gastric cancer. Comparison of the miRNA expression
levels between the tumor (T) and corresponding normal (N) tissues
obtained from 29 gastric cancer patients, which were analyzed using
the TCGA dataset. The expression levels of the miRNAs are presented
in transcripts per million (TPM). The expression level between
tumor and normal cells was evaluated by conducting paired t-tests
(P<0.05 was considered significant; NS, non-significant,
*P<0.05, **P<0.01,
***P<0.001). (B) The reporter of ET-1 3′UTR was
cotransfected with various miRNA constructs [vector (V), miR-1,
miR-101, miR-125a/b, miR-144 and let-7c] into AGS cells. Luciferase
assay was conducted after 48 h of cotransfection in AGS cells using
the Dual-Glo Luciferase Reporter Assay System kit. (C) After 48 h
transfection, the endogenous protein levels of ET-1 were examined
by western blot analysis. ET-1, endothelin-1; 3′UTR, 3′
untranslated region. |
DNA hypermethylation silences miR-1
expression in gastric cancer
Among the listed miRNAs, we selected miR-1 to
further examine the biological function and the regulatory
mechanisms in gastric cancer. The expression of miR-1 originates
from 2 paralogous genes, mir-1-1 and mir-1-2, located on
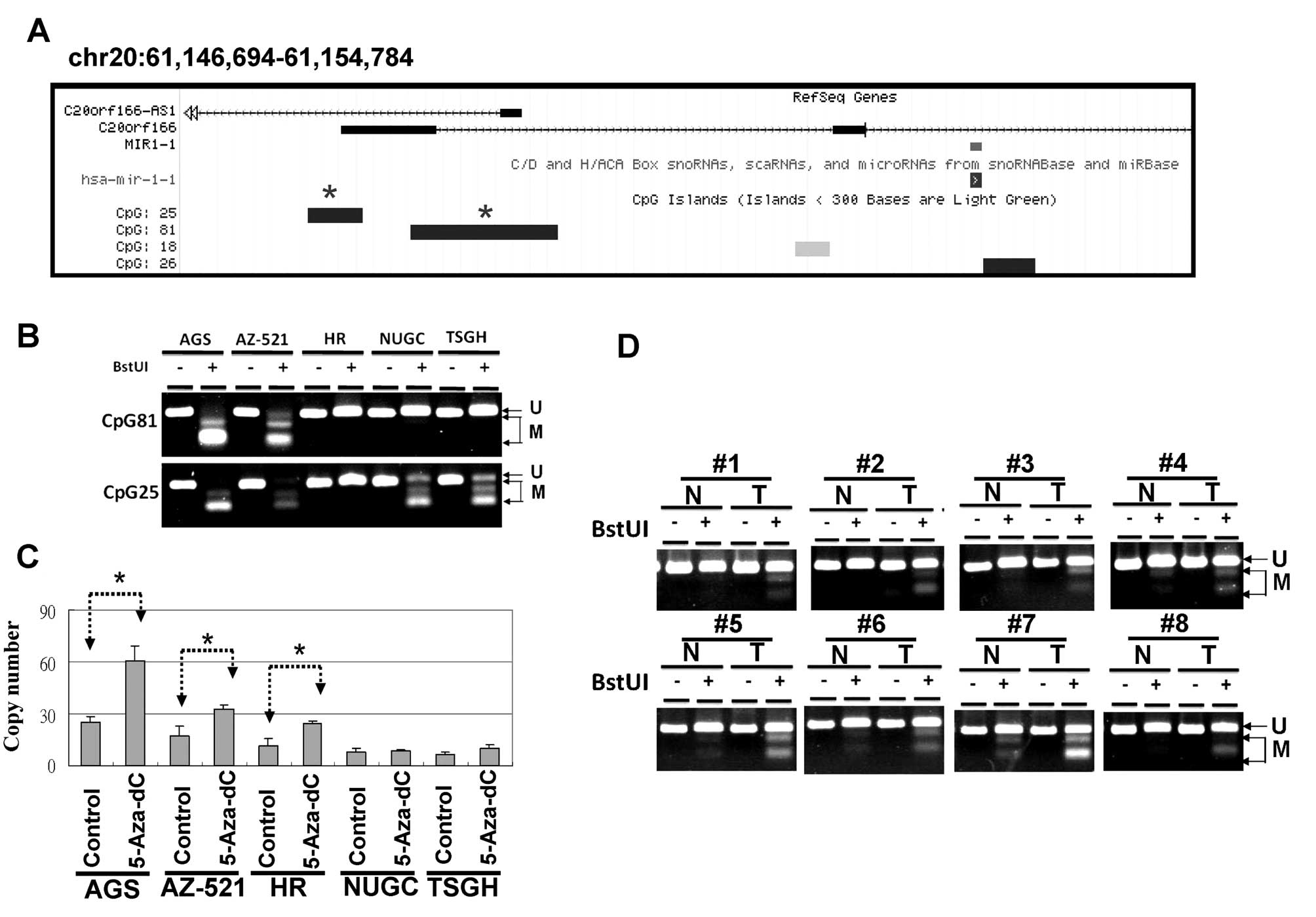
chromosomes 20 and 18, respectively. As shown in Fig. 4A, 4 CpG islands are located at the
putative transcription start sites of the miR-1-1 loci, according
to the UCSC database. We analyzed the methylation statuses of the
CpG25 and CpG81 regions in 5 human gastric cancer cell lines by
using a COBRA approach. As shown in Fig. 4B, the CpG81 regions were completely
methylated in the AGS and AZ-521 cells, yet unmethylated in the HR
cells. After treatment for 4 days with 5-Aza-dC, the expression
levels of miR-1 were restored in the AGS, AZ-521 and HR cells
(Fig. 4C). These data indicate that
DNA methylation contributes to the modulation of miR-1
transcriptional activity in gastric cancer cell lines.
By analyzing TCGA data, we found that the expression
levels of miR-1 were significantly decreased in the gastric cancer
cells compared with the corresponding adjacent normal cells
(Fig. 3A). Therefore, we further
examined the methylation status of the CpG81 regions in 50 primary
gastric cancers and the corresponding adjacent control, and
observed a high frequence of DNA hypermethylation in gastric cancer
(26 of 50). These results indicated that DNA hypermethylation leads
to low expression levels of miR-1 in gastric cancer (Fig. 4D).
miR-1 suppresses gastric cancer cell
growth
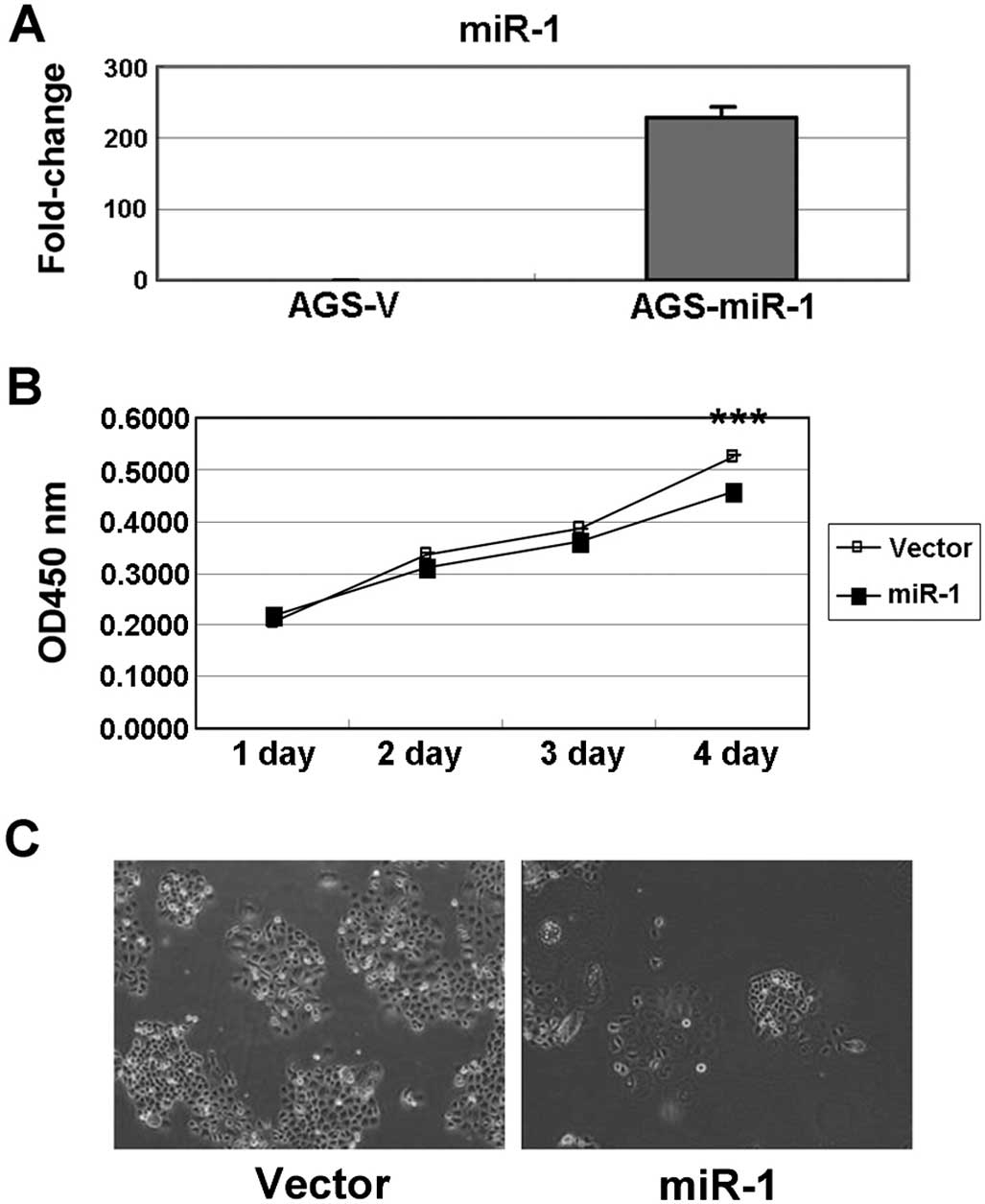
The ectopic expression of miR-1 in AGS cells
increased mature miR-1 expression by ~250-fold (Fig. 5A). The growth of AGS cells with
miR-1 overexpression was substantially inhibited compared with
those transfected with the vector (Fig.
5B). Furthermore, the clonogenic survival of AGS cells also
decreased after ectopic expression of miR-1 (Fig. 5C). Overall, our results imply that
ET-1 overexpression may be due to the silencing of miR-1 expression
by DNA hypermethylation in gastric cancer. In addition, we
identified several miRNA candidates that could modulate the ET-1
oncogenic function, and could also act as potential therapeutic
targets for therapy in the future.
Discussion
Dysfunction of the endothelin (ET) axis is known to
be a feature of human cancer, including lung, prostate, colorectal,
liver, breast, ovarian and oral cancer as well as melanomas.
Abnormal activation of the ET axis leads to promotion of cell
growth, metastasis and angiogenesis, and suppression of apoptosis
(4–11). By analyzing TCGA data, we found that
the expression levels of ET-1, ECE-1 and ECE-2 were significant
increased in the gastric cancer compared to the corresponding
normal tissues (Fig. 1). A previous
study reported that the plasma big-ET-1 level in gastric cancer was
significantly higher than that in the control (30). These findings imply that the
expression level of ET-1 is increased in gastric cancer. Although
ECE-1 (31) and ECE-2 were found to
be upregulated in a number of cancers, including prostate, ovarian
and oral cancer, thyroid carcinoma, and breast cancer (32–35),
the expression of ECE-1 and ECE-2 was not reported in gastric
cancer. Previous studies found that, in human cancer, ET-2 and ET-3
expression levels are silenced by DNA hypermethylation of their
promoter regions (36,37). In the present study, we observed
that the expression level of ET-2 was significant decreased in
gastric cancer. However, what mechanism is involved in the loss of
expression in gastric cancer remains unclear. These studies support
our finding that the ET-1 axis is overexpressed, and that ET-2 is
downregulated in gastric cancer.
In the past decade, miRNAs have emerged as playing
important roles via modulation of the cell cycle, cell growth and
cell motility in gastric cancer progression (21,22).
In the present study, we identified 15 miRNAs that may modulate
ET-1 expression by targeting 3′UTR. Among them, 6 miRNAs (miR-134,
miR-135b, miR-141, miR-425, miR-494 and miR-590) and 5 miRNAs
(miR-1, let-7c, miR-101, miR-125a and miR-144) showed significant
upregulation and downregulation, respectively, in gastric cancer.
Consistent with our findings, previous studies have revealed that
miR-135b and miR-425 act as oncomiRs involved in the tumorigenesis
of gastric cancer (38–40). Furthermore, miR-125a/b, miR-144 and
miR-101 have been reported to be significantly decreased and
display a tumor-suppressive function by silencing their oncongenic
targets in gastric cancer. Here, we demonstrated that miR-125a/b,
miR-101 and miR-144 suppressed ET1 expression by targeting the
3′UTR in gastric cancer. In addition, previous studies have
reported that miR-1, miR-125a/b, miR-155 and miR-199 suppress ET-1
expression by targeting 3′UTR (23–28).
In numerous cancers, miR-1 performs a tumor-suppressive function by
suppressing tumor cell growth and migration in prostate cancer,
hepatocellular, bladder and esophageal squamous cell carcinoma, and
lung cancer (29,41–48).
Furthermore, hypermethylation of the promoter region of mir-1-1
resulted in low expression in hepatocellular carcinoma and
colorectal cancer (29,48–50).
In the present study, we also demonstrated that miR-1 expression
levels were suppressed by DNA methylation and that ectopic miR-1
expression led to the suppression of gastric cancer cell growth.
Collectively, our results imply that ET-1 overexpression may be due
to DNA hypermethylation resulting in the silencing of miR-1
expression in gastric cancer.
In solid tumors, the ET-axis pathway plays a
critical oncogenic role in facilitating cancer growth and
metastasis. Various investigations have demonstrated that the
ET-1-ETRA axis is a potential therapeutic target for cancer
therapy. To date, several drugs have been designed as ETRA
antagonists for anticancer growth and metastasis. The ET-1 is a
short intercellular peptide derived from numerous tumor cells, and
has emerged as a potential therapeutic target or biomarker for
human cancer. In the present study, our identification of the
post-translational regulation of ET-1 may lead to novel insights
into the function of ET-1 in gastric cancer, and may provide a
potential therapeutic target.
Acknowledgements
This study was supported by grants from the
Kaohsiung Veterans General Hospital (VGHKS 102-020, VGHKS 102-074,
VGHKS 103-066 and VGHKS 103-108).
References
|
1
|
Yanagisawa M, Kurihara H, Kimura S, et al:
A novel potent vasoconstrictor peptide produced by vascular
endothelial cells. Nature. 332:411–415. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kedzierski RM and Yanagisawa M: Endothelin
system: the double-edged sword in health and disease. Annu Rev
Pharmacol Toxicol. 41:851–876. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rubanyi GM and Polokoff MA: Endothelins:
molecular biology, biochemistry, pharmacology, physiology, and
pathophysiology. Pharmacol Rev. 46:325–415. 1994.PubMed/NCBI
|
|
4
|
Grimshaw MJ: Endothelins in breast tumour
cell invasion. Cancer Lett. 222:129–138. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Asham E, Shankar A, Loizidou M, et al:
Increased endothelin-1 in colorectal cancer and reduction of tumour
growth by ETA receptor antagonism. Br J Cancer.
85:1759–1763. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Eberle J, Fecker LF, Orfanos CE and Geilen
CC: Endothelin-1 decreases basic apoptotic rates in human melanoma
cell lines. J Invest Dermatol. 119:549–555. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Douglas ML, Richardson MM and Nicol DL:
Endothelin axis expression is markedly different in the two main
subtypes of renal cell carcinoma. Cancer. 100:2118–2124. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kim TH, Xiong H, Zhang Z and Ren B:
β-Catenin activates the growth factor endothelin-1 in colon cancer
cells. Oncogene. 24:597–604. 2005. View Article : Google Scholar
|
|
9
|
Abdel-Gawad IA, Hassanein HM, Bahgat NA,
et al: Study of endothelin-1 and vascular endothelial growth factor
in patients with cancer colon. J Egypt Natl Canc Inst. 20:216–223.
2008.PubMed/NCBI
|
|
10
|
Said N, Smith S, Sanchez-Carbayo M and
Theodorescu D: Tumor endothelin-1 enhances metastatic colonization
of the lung in mouse xenograft models of bladder cancer. J Clin
Invest. 121:132–147. 2011. View
Article : Google Scholar :
|
|
11
|
Nelson JB, Udan MS, Guruli G and Pflug BR:
Endothelin-1 inhibits apoptosis in prostate cancer. Neoplasia.
7:631–637. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rosano L, Spinella F and Bagnato A:
Endothelin 1 in cancer: biological implications and therapeutic
opportunities. Nat Rev Cancer. 13:637–651. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nelson JB, Lee WH, Nguyen SH, et al:
Methylation of the 5′ CpG island of the endothelin B receptor gene
is common in human prostate cancer. Cancer Res. 57:35–37.
1997.PubMed/NCBI
|
|
14
|
Pao MM, Tsutsumi M, Liang G, Uzvolgyi E,
Gonzales FA and Jones PA: The endothelin receptor B (EDNRB)
promoter displays heterogeneous, site specific methylation patterns
in normal and tumor cells. Hum Mol Genet. 10:903–910. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cohen AJ, Belinsky S, Franklin W and Beard
S: Molecular and physiologic evidence for 5′CpG island methylation
of the endothelin B receptor gene in lung cancer. Chest.
121:27S–28S. 2002. View Article : Google Scholar
|
|
16
|
Lo KW, Tsang YS, Kwong J, To KF, Teo PM
and Huang DP: Promoter hypermethylation of the EDNRB gene in
nasopharyngeal carcinoma. Int J Cancer. 98:651–655. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhao BJ, Sun DG, Zhang M, Tan SN and Ma X:
Identification of aberrant promoter methylation of EDNRB gene in
esophageal squamous cell carcinoma. Dis Esophagus. 22:55–61. 2009.
View Article : Google Scholar
|
|
18
|
Mathieu MN and Chevillard C: Endothelin-1
and ETA receptor subtype are expressed in the gastric HGT-1 cell
line. J Cardiovasc Pharmacol. 26(Suppl 3): S508–S509. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ferrari-Bravo A, Franciosi C, Lissoni P,
Fumagalli L and Uggeri F: Effects of oncological surgery on
endothelin-1 secretion in patients with operable gastric cancer.
Int J Biol Markers. 15:56–57. 2000.PubMed/NCBI
|
|
20
|
Stow LR, Jacobs ME, Wingo CS and Cain BD:
Endothelin-1 gene regulation. FASEB J. 25:16–28. 2011. View Article : Google Scholar :
|
|
21
|
Pan HW, Li SC and Tsai KW: MicroRNA
dysregulation in gastric cancer. Curr Pharm Des. 19:1273–1284.
2013.
|
|
22
|
Wu HH, Lin WC and Tsai KW: Advances in
molecular biomarkers for gastric cancer: miRNAs as emerging novel
cancer markers. Expert Rev Mol Med. 16:e12014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jacobs ME, Wingo CS and Cain BD: An
emerging role for microRNA in the regulation of endothelin-1. Front
Physiol. 4:222013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yeligar S, Tsukamoto H and Kalra VK:
Ethanol-induced expression of ET-1 and ET-BR in liver sinusoidal
endothelial cells and human endothelial cells involves
hypoxia-inducible factor-1α and microRNA-199. J Immunol.
183:5232–5243. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li D, Yang P, Xiong Q, et al:
MicroRNA-125a/b-5p inhibits endothelin-1 expression in vascular
endothelial cells. J Hypertens. 28:1646–1654. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li D, Yang P, Li H, et al: MicroRNA-1
inhibits proliferation of hepatocarcinoma cells by targeting
endothelin-1. Life Sci. 91:440–447. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li D, He B, Zhang H, et al: The inhibitory
effect of miRNA-1 on ET-1 gene expression. FEBS Lett.
586:1014–1021. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Feng B, Cao Y, Chen S, Ruiz M and
Chakrabarti S: miRNA-1 regulates endothelin-1 in diabetes. Life
Sci. 98:18–23. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chen WS, Leung CM, Pan HW, et al:
Silencing of miR-1-1 and miR-133a-2 cluster expression by DNA
hypermethylation in colorectal cancer. Oncol Rep. 28:1069–1076.
2012.PubMed/NCBI
|
|
30
|
Teng XJ, Shen ZX, Xiang JJ, et al: Pre-
and post-operative plasma big endothelin-1 levels in patients with
gastric carcinoma undergoing radical gastrectomy. Anticancer Res.
26:2503–2507. 2006.PubMed/NCBI
|
|
31
|
Whyteside AR, Hinsley EE, Lambert LA,
McDermott PJ and Turner AJ: ECE-1 influences prostate cancer cell
invasion via ET-1-mediated FAK phosphorylation and ET-1-independent
mechanisms. Can J Physiol Pharmacol. 88:850–854. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Smollich M, Götte M, Yip GW, et al: On the
role of endothelin-converting enzyme-1 (ECE-1) and neprilysin in
human breast cancer. Breast Cancer Res Treat. 106:361–369. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Dawson LA, Maitland NJ, Berry P, Turner AJ
and Usmani BA: Expression and localization of endothelin-converting
enzyme-1 in human prostate cancer. Exp Biol Med. 231:1106–1110.
2006.
|
|
34
|
Awano S, Dawson LA, Hunter AR, Turner AJ
and Usmani BA: Endothelin system in oral squamous carcinoma cells:
specific siRNA targeting of ECE-1 blocks cell proliferation. Int J
Cancer. 118:1645–1652. 2006. View Article : Google Scholar
|
|
35
|
Van Beneden R, Michel L, Havaux X, Delos M
and Donckier J: Increased expression of endothelin-1 converting
enzyme in human thyroid carcinoma. Clin Endocrinol. 60:146–147.
2004. View Article : Google Scholar
|
|
36
|
Wiesmann F, Veeck J, Galm O, et al:
Frequent loss of endothelin-3 (EDN3) expression due to epigenetic
inactivation in human breast cancer. Breast Cancer Res. 11:R342009.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang R, Lohr CV, Fischer K, et al:
Epigenetic inactivation of endothelin-2 and endothelin-3 in colon
cancer. Int J Cancer. 132:1004–1012. 2013. View Article : Google Scholar
|
|
38
|
Lim JY, Yoon SO, Seol SY, et al:
Overexpression of miR-196b and HOXA10 characterize a poor-prognosis
gastric cancer subtype. World J Gastroenterol. 19:7078–7088. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ma J, Liu J, Wang Z, et al:
NF-kappaB-dependent microRNA-425 upregulation promotes gastric
cancer cell growth by targeting PTEN upon IL-1β induction. Mol
Cancer. 13:402014. View Article : Google Scholar
|
|
40
|
Peng WZ, Ma R, Wang F, Yu J and Liu ZB:
Role of miR-191/425 cluster in tumorigenesis and diagnosis of
gastric cancer. Int J Mol Sci. 15:4031–4048. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yoshino H, Chiyomaru T, Enokida H, et al:
The tumour-suppressive function of miR-1 and miR-133a targeting
TAGLN2 in bladder cancer. Br J Cancer. 104:808–818. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wu CD, Kuo YS, Wu HC and Lin CT:
MicroRNA-1 induces apoptosis by targeting prothymosin alpha in
nasopharyngeal carcinoma cells. J Biomed Sci. 18:802011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wang F, Song G, Liu M, Li X and Tang H:
miRNA-1 targets fibronectin1 and suppresses the migration and
invasion of the HEp2 laryngeal squamous carcinoma cell line. FEBS
Lett. 585:3263–3269. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Nohata N, Sone Y, Hanazawa T, et al: miR-1
as a tumor suppressive microRNA targeting TAGLN2 in head and neck
squamous cell carcinoma. Oncotarget. 2:29–42. 2011.PubMed/NCBI
|
|
45
|
Nasser MW, Datta J, Nuovo G, et al:
Down-regulation of micro-RNA-1 (miR-1) in lung cancer. Suppression
of tumorigenic property of lung cancer cells and their
sensitization to doxorubicin-induced apoptosis by miR-1. J Biol
Chem. 283:33394–33405. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Kinoshita T, Nohata N, Fuse M, et al:
Tumor suppressive microRNA-133a regulates novel targets: moesin
contributes to cancer cell proliferation and invasion in head and
neck squamous cell carcinoma. Biochem Biophys Res Commun.
418:378–383. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Hudson RS, Yi M, Esposito D, et al:
MicroRNA-1 is a candidate tumor suppressor and prognostic marker in
human prostate cancer. Nucleic Acids Res. 40:3689–3703. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Datta J, Kutay H, Nasser MW, et al:
Methylation mediated silencing of microRNA-1 gene and its role in
hepatocellular carcinogenesis. Cancer Res. 68:5049–5058. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Migliore C, Martin V, Leoni VP, et al:
MiR-1 downregulation cooperates with MACC1 in promoting MET
overexpression in human colon cancer. Clin Cancer Res. 18:737–747.
2012. View Article : Google Scholar
|
|
50
|
Suzuki H, Takatsuka S, Akashi H, et al:
Genome-wide profiling of chromatin signatures reveals epigenetic
regulation of microRNA genes in colorectal cancer. Cancer Res.
71:5646–5658. 2011. View Article : Google Scholar : PubMed/NCBI
|