Introduction
Non-small cell lung cancer (NSCLC) patients
harboring activating mutations in epidermal growth factor receptor
(EGFR) initially respond well to treatment with EGFR-tyrosine
kinase inhibitors (TKIs) such as gefitinib or erlotinib (1,2).
However, drug resistance to treatment with EGFR-TKIs eventually is
inevitable and limits the clinical benefits. The mechanisms
underlying EGFR-TKIs resistance are multi-factorial and many have
been reported, for example, the T790M mutation in EGFR, MET
amplification, NF-κB activation and emergence of SCLC
(3–5). However, ~30% resistant mechanisms are
unknown. Thus, elucidating the molecular mechanisms underlying
EGFR-TKIs resistance is essential for the identification of key
biomarkers.
Recent findings have shown that non-coding RNAs
(ncRNAs) are involved in the pathogenesis of NSCLC, providing new
insights into the biology of diseases (6–9). Long
non-coding RNAs (lncRNAs) are non-protein coding transcripts,
>200 nucleotides (nt) in length. lncRNAs play important
regulatory roles in the cancer development, metastasis and
chemotherapy resistance of multiple types of cancer (10–12).
Accumulating evidence suggested that lncRNAs are abnormally
expressed in various types of human cancer (10,13–15).
Studies have reported that lncRNAs influence chemotherapy
resistance (16–18). For example, lncRNA-AK126698 is
associated with cisplatin resistance in NSCLC (19). Another study has shown that
cisplatin-based chemotherapy results in the upregulation of UCA1
expression in patients with bladder cancer (17). Therefore, lncRNAs are possible novel
candidate biomarkers and potential targets for EGFR-TKIs
therapy.
To ascertain whether lncRNAs expression signatures
can differ between gefitinib-sensitive PC9 and gefitinib-resistant
PC9/R cells, lncRNAs expression profiles were identified using
microarray analysis. Subsequent to obtaining expression-profile
results for these samples, we investigated the relationship between
lncRNAs and EGFR-TKIs resistance in NSCLC.
Materials and methods
Cell culture
PC9 (EGFR exon 19 deletion), H1975 (L858R/T790M),
H1299 and A549 (EGFR wild-type) human lung adenocarcinoma cell line
were obtained from the American Type Culture Collection (ATCC;
Vanassas, MA, USA). Gefitinib-induced acquired resistant lung
cancer cells from PC9 (PC9/R) were provided by the Shanghai
Pulmonary Hospital. The cells were cultured in a 37°C humidified
incubator with 5% CO2 in Dulbecco’s modified Eagle’s
medium (DMEM) supplemented with 10% fetal bovine serum (FBS) (both
from Gibco Life Technologies, Carlsbad, CA, USA).
Cell proliferation assay
Cells were seeded in 96-well plates at
5×103 cells/well, and treated with various concentration
of gefitinib for 72 h. At the end of incubation, the cell
proliferation reagent Cell Counting Kit-8 (CCK-8; Dojindo
Laboratories, Japan) (10 μl) was added to each well and incubated
for 1 h at 37°C. Viable cell numbers were estimated by measurement
of optical density (OD) at 450 nm.
LncRNA microarray
The Arraystar Human LncRNA Microarray v2.0 has been
designed for the global profiling of human lncRNAs and
protein-coding transcripts. A total of 33,045 lncRNAs and 30,215
coding transcripts can be detected by second-generation lncRNA
microarray. The lncRNAs were carefully collected from the most
authoritative databases, including RefSeq, UCSC knowngenes, Ensembl
and a number of related studies. Each transcript is represented by
a specific exon or splice junction probe that can accurately
identify individual transcripts. Positive probes for housekeeping
genes and negative probes were also printed onto the array for
hybridization quality control.
RNA labeling and array hybridization
Sample labeling and array hybridization were
performed according to the Agilent One-Color Microarray-Based Gene
Expression Analysis protocol (Agilent Technologies, Inc., Santa
Clara, CA, USA) with minor modifications. Briefly, mRNA was
purified from total RNA following removal of rRNA (mRNA-ONLY™
Eukaryotic mRNA Isolation kit; Epicentre). Each sample was
amplified and transcribed into fluorescent cRNA along the entire
length of the transcripts without 3′ bias utilizing a random
priming method. The labeled cRNAs were purified using an RNeasy
Mini kit (Qiagen, Valencia, CA, USA). The concentration and
specific activity of the labeled cRNAs (pmol Cy3/μg cRNA) were
measured by NanoDrop ND-1000. Each labeled cRNA (1 μg) was
fragmented by adding 5 μl 10X blocking agent and 1 μl of 25X
fragmentation buffer. The mixture was heated at 60°C for 30 min,
and 25 μl 2X GE hybridization buffer was added to dilute the
labeled cRNA. Hybridization solution (50 μl) was dispensed into the
gasket slide and assembled to the lncRNA expression microarray
slide. The slides were incubated for 17 h at 65°C in an Agilent
hybridization oven. The hybridized arrays were washed, fixed and
scanned using the Agilent DNA Microarray Scanner (part no.
G2505C).
RNA extraction and quantitative PCR
Total RNA was extracted from lung cancer cell lines
using TRIzol reagent (Takara, Japan). The expression of lncRNAs in
lung cancer cell lines was measured by qPCR using SYBR Premix
Ex Taq (Takara code: DRR420A) on MX3000P instrument and
using the following cycling parameters: initial denaturation at
95°C for 30 sec, followed by 40 cycles of 95°C for 5 sec, 60°C for
20 sec and 95°C for 60 sec, and 60°C for 30 sec. Primers were
designed by Sangon Biotech (China). GAPDH was used as a control.
Experiments were carried out in triplicate. The median in each
triplicate was used to calculate relative lncRNA concentrations
using the formula: ΔCt = Ctmedian lncRNAs −
Ctmedian GAPDH. Expression fold changes were calculated
using 2−ΔΔCt methods.
Data analysis
The Agilent Feature Extraction software (version
11.0.1.1) was used to analyze acquired array images. Quantile
normalization and subsequent data processing were performed using
the GeneSpring GX v11.5.1 software package (Agilent Technologies).
Differentially expressed LncRNAs and mRNAs were identified through
fold-change filtering. Hierarchical clustering was performed using
the Agilent GeneSpring GX software (version 11.5.1). Gene ontology
(GO) and pathway analysis were performed in the standard enrichment
computation method.
Statistical analysis
Statistical analysis was performed using SPSS
version 17.0 software (SPSS, Inc., Chicago, IL, USA). Results were
presented as means ± standard deviation (SD) of three separate
assays. Differences between groups were assessed using the t-test
(two-tailed). P<0.05 was considered to indicate a statistically
significant result.
Results
Differentially expressed lncRNAs and
mRNAs
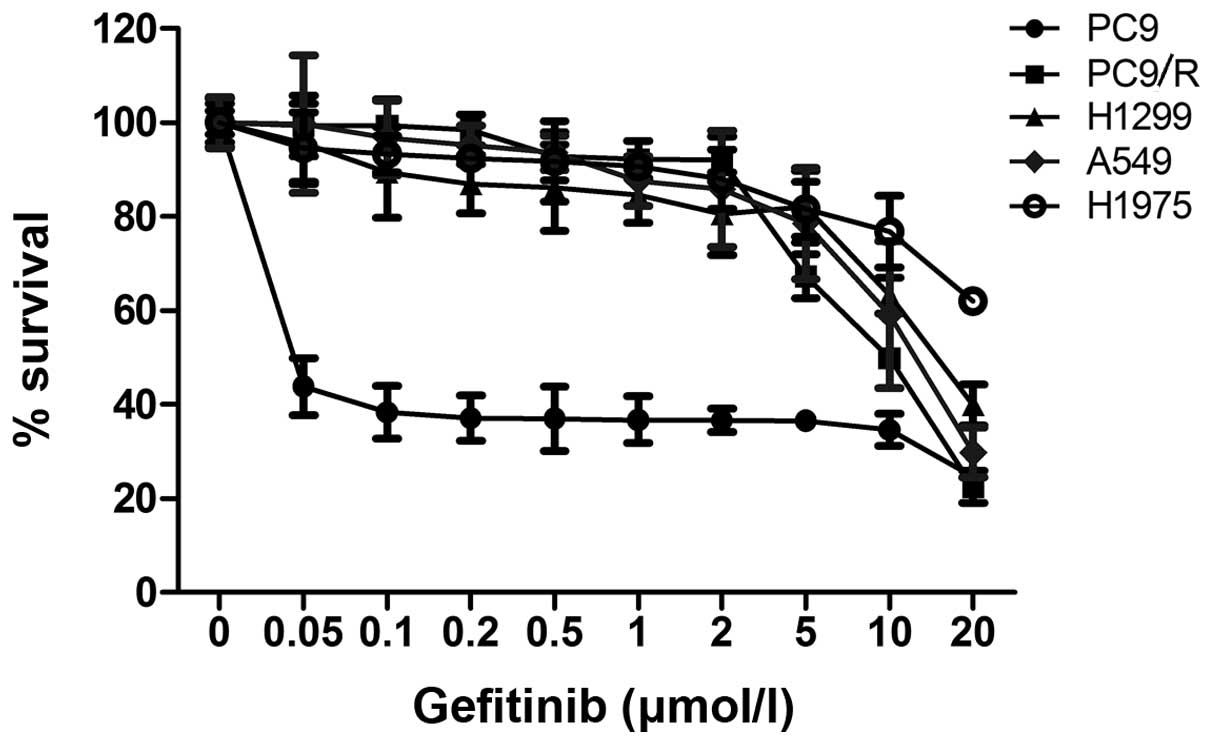
Gefitinib-resistant PC9/R cells were identified by
evaluating the IC50 value of PC9/R against the PC9 cell
line. The IC50 value of gefitinib for the drug resistant
PC9/R cell line was 15 μmol/l was 300-fold higher than that of the
PC9 cell line (0.05 μmol/l) (Fig.
1). This result demonstrated that PC9/R cells were more
resistant to gefitinib than PC9 cells.
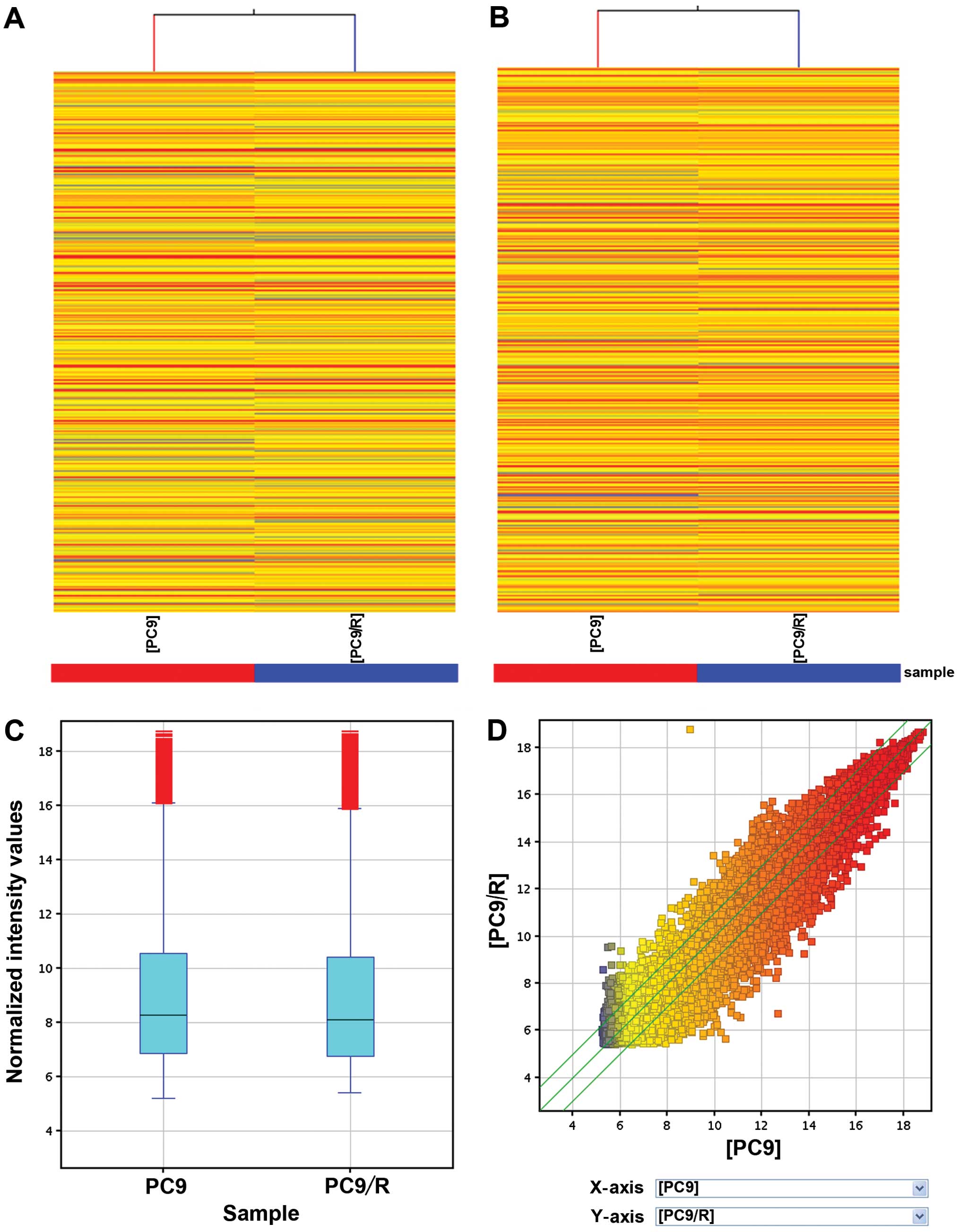
For the microarray analysis, 22,587 lncRNAs
(Fig. 2A, C and D) and 17,479 mRNAs
(Fig. 2B) were differentially
expressed in PC9 and PC9/R with gefitinib-sensitive and
acquired-resistant cells, respectively. Of these, 1,731 lncRNAs
were upregulated >2-folds in PC9/R compared to PC9 cells, while
2,936 were downregulated (P<0.01) (Table I). In addition, as compared to PC9
cells, 2,349 mRNAs were increased and 1,307 were decreased in the
expression level in PC9/R cells (fold-change >2, P<0.01).
 | Table IThe differentially expressed lncRNAs
in gefitinib-resistant and gefitinib-sensitive lung cancer
cells. |
Table I
The differentially expressed lncRNAs
in gefitinib-resistant and gefitinib-sensitive lung cancer
cells.
| Up- or
Downregulated | Fold-change
>2 | Fold-change
>5 | Fold-change
>10 |
|---|
| Up | 1,731 | 79 | 6 |
| Down | 2,936 | 326 | 43 |
GO analysis
GO analysis is a functional analysis associating
differentially expressed mRNAs with GO categories. In the present
study, GO analysis was performed to determine the gene and gene
product enrichment. The Fisher’s exact test was used to determine
whether there was more overlapping between the DE and the GO
annotation list than would be expected by chance.
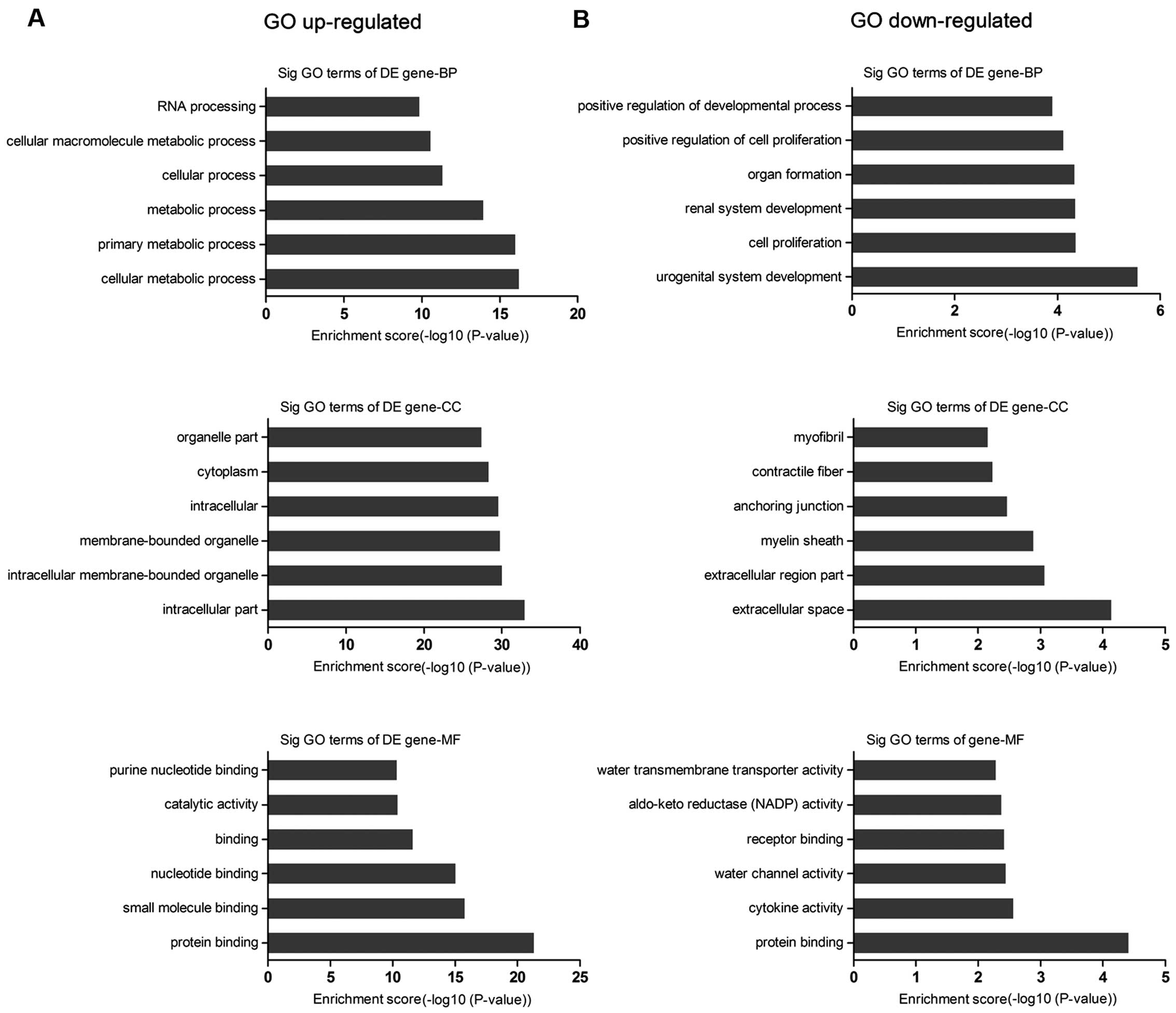
We found that the highest enriched GOs targeted by
upregulated transcripts were cellular metabolic process (GO:
biological processes), intracellular component (GO: cellular
components), and protein binding (GO: molecular functions)
(Fig. 3A), while the downregulated
transcripts were urogenital system development (GO: biological
processes), extracellular space (GO: cellular components), and
protein binding (GO: molecular functions) (Fig. 3B). Of note, the lower the P-value,
the more significant the GO term (P-value cut-off of ≤0.05 was
recommended). We also found that 699 genes were associated with
cell proliferation and cell apoptosis (Table II).
| Table IIGene ontology categories ranked by
cell proliferation and apoptosis. |
Table II
Gene ontology categories ranked by
cell proliferation and apoptosis.
| GO.ID | Term | Count | Fold.
enrichment | P-value |
|---|
| Upregulated |
| 0006915 | Apoptotic
process | 216 | 1.252336427 | 0.000174419 |
| 0008283 | Cell
proliferation | 190 | 1.181451791 | 0.00654506 |
| 0006917 | Induction of
apoptosis | 56 | 1.259490591 | 0.040513474 |
| Downregulated |
| 0097190 | Apoptotic signaling
pathway | 8 | 2.724513133 | 0.008043735 |
| 2001233 | Regulation of
apoptotic signaling pathway | 7 | 2.872964169 | 0.009633905 |
| 0097193 | Intrinsic apoptotic
signaling pathway | 4 | 3.201302932 | 0.032869143 |
| 0008283 | Cell
proliferation | 124 | 1.410666537 | 4.12154E-05 |
| 0042127 | Regulation of cell
proliferation | 94 | 1.415439678 | 0.000335816 |
Pathway analysis
Based on the latest Kyoto Encyclopedia of Genes and
Genomes (KEGG) database, we provided pathway analysis for
differentially expressed mRNAs. This analysis allowed us to
determine the biological pathway, which showed a significant
enrichment of differentially expressed mRNAs.
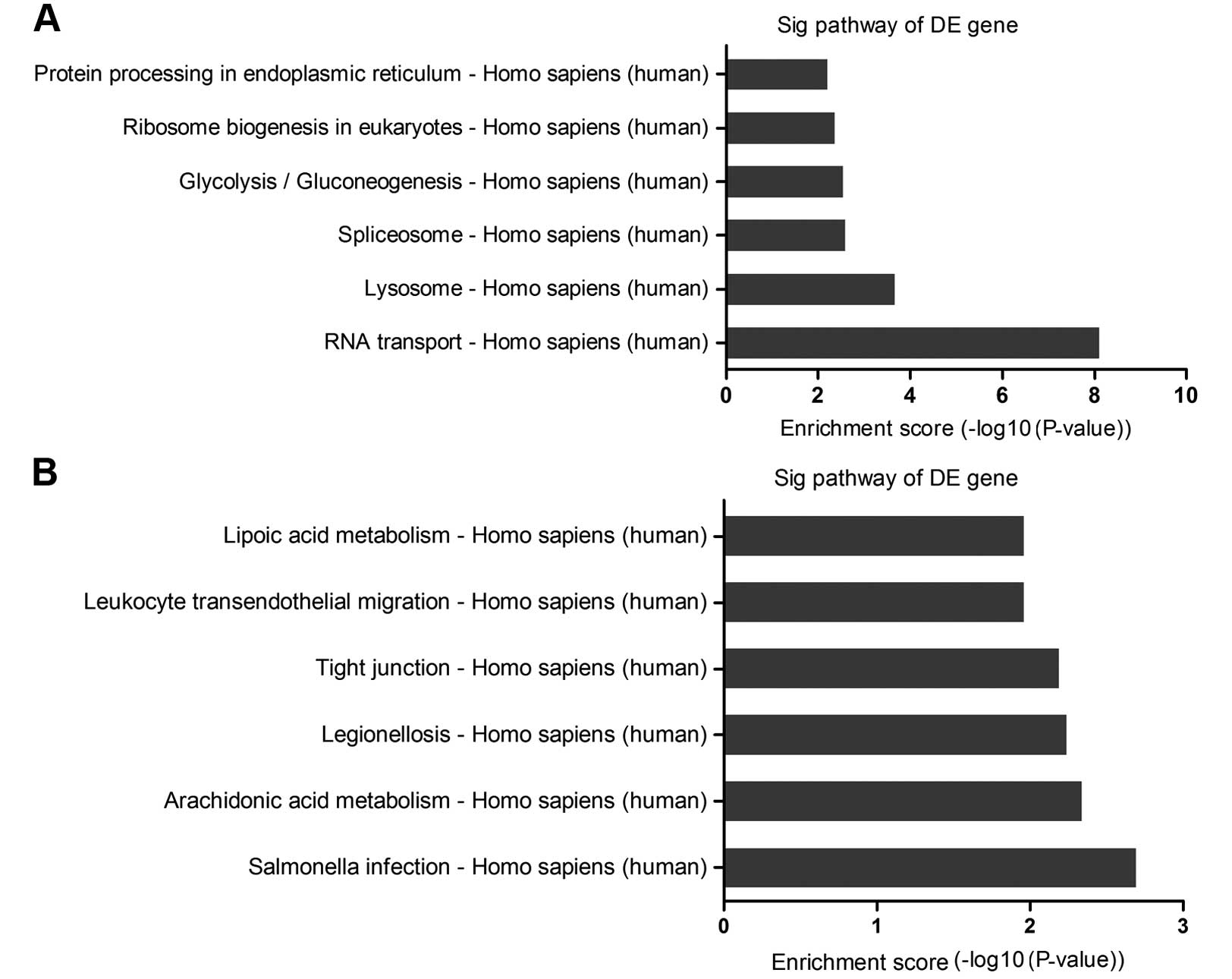
The pathway analysis indicated that there were 19
pathways corresponding to the upregulated transcripts. The
high-enrichment pathways targeted by overexpressed mRNAs were
involved in RNA transport, lysosome and spliceosome (Fig. 4A). By contrast, there were 17
pathways involved in the downregulated transcripts. Significant
pathways corresponding to downregulated mRNAs appeared to be
responsible for salmonella infection, arachidonic acid metabolism,
legionellosis, with the recommended P-value cut-off being 0.05)
(Fig. 4B). Of these, the
enriched-pathways associated with cell proliferation and apoptosis
played a critical role in EGFR-TKIs resistance.
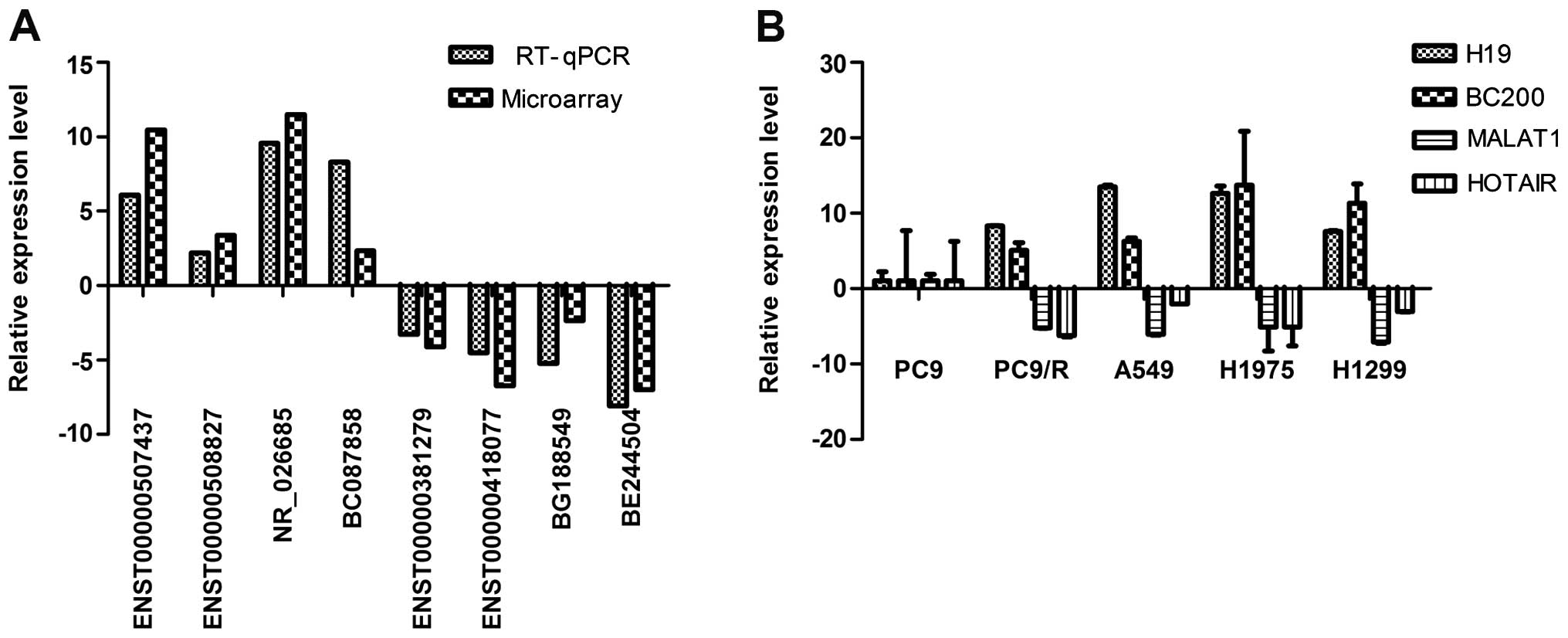
RT-qPCR validation
We examined the expression of four studied lncRNAs
(H19, BC200, MALAT1 and HOTAIR) in lung cancer cell lines using
RT-qPCR to validate the results of lncRNA profiles of which H19 and
BC200 were upregulated and MALAT1 and HOTAIR were downregulated.
The resistance of cell lines to gefitinib was assessed using CCK-8
assay and results are shown in Fig.
1. Overexpression of H19 and BC200 were observed in
gefitinib-resistant lung cancer cell lines (PC9/R, A549, H1299 and
H1975). However, in the gefitinib-sensitive cell line (PC9), MALAT1
and HOTAIR were downregulated (Fig.
5B).
Four upregulated and four downregulated lncRNAs from
differentially expressed lncRNAs were randomly selected. Using
RT-qPCR the results of microarray in PC9/R vs. PC9 were validated.
The expression levels of ENST00000507437, ENST00000508827,
NR_026685 and BC087858 were upregulated and ENST00000381279,
ENST00000418077, BG188549 and BE244504 were downregulated (Fig. 5A). Thus, the microarray data were
confirmed by RT-qPCR.
Discussion
Aberrant lncRNAs expression patterns have been
described in various types of cancer (10,20–22),
and alterations in expression of lncRNAs correlate with prognosis
and chemotherapy resistance of human cancer (16,17).
HOTAIR, HULC and H19 are crucial in the development and progression
of tumors and are associated with chemotherapy resistance (7,16,17).
However, the functions of lncRNAs should be investigated in
targeted therapy of human malignant tumors. In the present study,
we showed that differentially expressed lncRNAs are associated with
EGFR-TKIs resistance.
We first identified a total of 22,587 differentially
expressed lncRNAs in gefitinib-sensitive and gefitinib-induced
acquired-resistant lung cancer cells. Furthermore, to gain insight
into the underlying biology of the differentially expressed
transcripts, we utilized GO and pathway analyses to study the
biological functions to these lncRNAs in the resistance of
EGFR-TKIs. We found 699 genes associated with cell proliferation
and apoptosis. In the pathway analysis, 19 pathways were identified
to correspond to the upregulated transcripts. By contrast, there
were 17 pathways involved in the downregulated transcripts. The
enriched pathways associated with cell proliferation and apoptosis
suggested involvement in EGFR-TKIs resistance. Previous studies
reported that the mechanism of EGFR-TKIs resistance is associated
with cell proliferation and cell apoptosis (23–25).
Findings of our previous study showed that the activation of
PI3K/Akt, MEK pathway and BIM were involved in the resistance of
EGFR-TKIs (23,26). Those results indicate that some
lncRNAs may play critical roles in gefitinib-resistant lung cancer
cells through cell proliferation and apoptosis.
It has been reported that lncRNAs regulate the
neighboring protein-coding genes (27). Therefore, we analyzed aberrant
lncRNAs and their nearby coding genes. Some lncRNAs may play
critical roles in regulating nearby genes that encode markers in
EGFR-TKIs resistance. For example, upregulated lncRNAs-BC087858 is
a 1,322 bp intergenic lncRNAs and was found to be located near
forkhead box protein C1 (FOXC1), which is a member of the FOX
transcription factor family and is important in cancer development
(28). FOXC1 induces
epithelial-mesenchymal transition (EMT) through inhibition of
E-cadherin expression and promotes cell migration and invasion.
Additionally, FOXC1 expression can be activated by epidermal growth
factor/extracellular signal-related kinase (EGF/ERK) signaling
pathways (29). It has been
observed that NSCLC cells resistant to EGFR-TKIs exhibit EMT
features (30–32). Thus, our results suggest that
BC087858 may be involved in EGFR-TKIs resistance through EMT. The
lncRNA-RP11-15H7.2 is a 1,580 bp intergenic lncRNAs that was found
to be located near CITED2, a transcriptional modulator that is
involved in human oncogenesis. Wu et al reported that CITED2
was involved in the resistance of cancer cells to the
chemotherapeutic drug cisplatin (33). Therefore, lncRNAs may influence
EGFR-TKIs resistance by regulating the nearby genes.
Additionally, we selected the expression of some
studied lncRNAs (H19, HOTAIR, MALAT1 and BC200) to validate the
consistency of microarray. Accumulating evidence suggests that H19
may be an oncogene or a tumor-suppressor gene that plays an
important role in cancer development. Tsang and Kwok reported that
H19 can induce MDR-1-associated chemotherapy resistance in human
hepatocellular carcinoma cells (16). MALAT-1 was downregulated in
gefitinib-resistant cells. In some studies, their potential roles
of regulating EMT transcription have been reported (34). MALAT-1 was linked to EMT-associated
transcription factors ZEB1, ZEB2, slug and E-cadherin. Moreover,
MALAT-1 promoted EMT by activating the Wnt signaling pathway
(35,36). Our results suggested that those
lncRNAs may be involved in the EGFR-TKIs resistance in NSCLC and
may provide novel pathways for improved understanding of the
molecular mechanisms underlying EGFR-TKIs resistance. However,
future studies are required to fully elucidate the mechanisms by
which lncRNAs may promote resistance to EGFR-TKIs.
In conclusion, we reported that numerous lncRNAs
were differentially expressed between gefitinib-sensitive PC9 and
gefitinib-resistant PC9/R cells, many of which played important
roles in regulating EGFR-TKIs resistance through various mechanisms
including EMT and cell apoptosis. It is suggested that lncRNAs can
exert their functions through interactions with coding transcripts
and proteins in NSCLC EGFR-TKIs resistance. However, the exact
mechanisms of lncRNAs require further study and to determine
whether the differentially expressed lncRNAs were involved in
EGFR-TKIs resistance in NSCLC. Based on these results, future
studies focusing on lncRNAs expression and functions should be
performed to assist patients in the improvement of response to
EGFR-TKIs.
Acknowledgements
This study was supported by grants from the National
Natural Science Foundation of China (nos. 81372392 and 81172101),
and the Key Project of the Science and Technology Commission of
Shanghai Municipality (nos. 124119a800 and 11JC1411301).
References
|
1
|
Zhang Z, Lee JC, Lin L, et al: Activation
of the AXL kinase causes resistance to EGFR-targeted therapy in
lung cancer. Nat Genet. 44:852–860. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhou C, Wu YL, Chen G, et al: Erlotinib
versus chemotherapy as first-line treatment for patients with
advanced EGFR mutation-positive non-small-cell lung cancer
(OPTIMAL, CTONG-0802): a multicentre, open-label, randomised, phase
3 study. Lancet Oncol. 12:735–742. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bar J and Onn A: Overcoming molecular
mechanisms of resistance to first-generation epidermal growth
factor receptor tyrosine kinase inhibitors. Clin Lung Cancer.
13:267–279. 2012. View Article : Google Scholar
|
|
4
|
Turke AB, Zejnullahu K, Wu YL, et al:
Preexistence and clonal selection of MET amplification in EGFR
mutant NSCLC. Cancer Cell. 17:77–88. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bean J, Brennan C, Shih JY, et al: MET
amplification occurs with or without T790M mutations in EGFR mutant
lung tumors with acquired resistance to gefitinib or erlotinib.
Proc Natl Acad Sci USA. 104:20932–20937. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Qiu M, Xu Y, Yang X, et al: CCAT2 is a
lung adenocarcinoma-specific long non-coding RNA and promotes
invasion of non-small cell lung cancer. Tumour Biol. 35:5375–5380.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liu Z, Sun M, Lu K, et al: The long
noncoding RNA HOTAIR contributes to cisplatin resistance of human
lung adenocarcinoma cells via downregualtion of
p21WAF1/CIP1 expression. PLoS One. 8:e772932013.
View Article : Google Scholar
|
|
8
|
Gutschner T, Hämmerle M, Eissmann M, et
al: The noncoding RNA MALAT1 is a critical regulator of the
metastasis phenotype of lung cancer cells. Cancer Res.
73:1180–1189. 2013. View Article : Google Scholar :
|
|
9
|
Liu XH, Liu ZL, Sun M, et al: The long
non-coding RNA HOTAIR indicates a poor prognosis and promotes
metastasis in non-small cell lung cancer. BMC Cancer. 13:4642013.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Takahashi K, Yan I, Haga H and Patel T:
Long noncoding RNA in liver diseases. Hepatology. 60:744–753. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhao J and Lawless MW: Long noncoding RNAs
and their role in the liver cancer axis. Nat Rev Gastroenterol
Hepatol. Nov 19–2013.(Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ren S, Wang F, Shen J, et al: Long
non-coding RNA metastasis associated in lung adenocarcinoma
transcript 1 derived miniRNA as a novel plasma-based biomarker for
diagnosing prostate cancer. Eur J Cancer. 49:2949–2959. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Iacoangeli A, Lin Y, Morley EJ, et al:
BC200 RNA in invasive and preinvasive breast cancer.
Carcinogenesis. 25:2125–2133. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Svoboda M, Slyskova J, Schneiderova M, et
al: HOTAIR long non-coding RNA is a negative prognostic factor not
only in primary tumors, but also in the blood of colorectal cancer
patients. Carcinogenesis. 35:1510–1515. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yuan JH, Yang F, Wang F, et al: A long
noncoding RNA activated by TGF-β promotes the invasion-metastasis
cascade in hepatocellular carcinoma. Cancer Cell. 25:666–681. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tsang WP and Kwok TT: Riboregulator H19
induction of MDR1-associated drug resistance in human
hepatocellular carcinoma cells. Oncogene. 26:4877–4881. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tsang WP, Wong TW, Cheung AH, et al:
Induction of drug resistance and transformation in human cancer
cells by the noncoding RNA CUDR. RNA. 13:890–898. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jiang M, Huang O, Xie Z, et al: A novel
long non-coding RNA-ARA: adriamycin resistance-associated. Biochem
Pharmacol. 87:254–283. 2014. View Article : Google Scholar
|
|
19
|
Yang Y, Li H, Hou S, et al: The noncoding
RNA expression profile and the effect of lncRNA AK126698 on
cisplatin resistance in non-small-cell lung cancer cell. PLoS One.
8:e653092013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gainor JF and Shaw AT: Emerging paradigms
in the development of resistance to tyrosine kinase inhibitors in
lung cancer. J Clin Oncol. 31:3987–3996. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang EB, Yin DD, Sun M, et al:
P53-regulated long non-coding RNA TUG1 affects cell proliferation
in human non-small cell lung cancer, partly through epigenetically
regulating HOXB7 expression. Cell Death Dis. 5:e12432014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hauptman N and Glavač D: Long non-coding
RNA in cancer. Int J Mol Sci. 14:4655–4669. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li H, Schmid-Bindert G, Wang D, et al:
Blocking the PI3K/AKT and MEK/ERK signaling pathways can overcome
gefitinib-resistance in non-small cell lung cancer cell lines. Adv
Med Sci. 56:275–284. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sordella R, Bell DW, Haber DA and
Settleman J: Gefitinibsensitizing EGFR mutations in lung cancer
activate anti-apoptotic pathways. Science. 305:1163–1167. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ng KP, Hillmer AM, Chuah CT, et al: A
common BIM deletion polymorphism mediates intrinsic resistance and
inferior responses to tyrosine kinase inhibitors in cancer. Nat
Med. 18:521–528. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li Z, Zhou S, Zhang L, et al: BIM
induction of apoptosis triggered by EGFR-sensitive and resistance
cell lines of non-small-cell lung cancer. Med Oncol. 28:572–577.
2011. View Article : Google Scholar
|
|
27
|
Yang F, Zhang L, Huo XS, et al: Long
noncoding RNA high expression in hepatocellular carcinoma
facilitates tumor growth through enhancer of zeste homolog 2 in
humans. Hepatology. 54:1679–1689. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xia L, Huang W, Tian D, et al:
Overexpression of forkhead box C1 promotes tumor metastasis and
indicates poor prognosis in hepatocellular carcinoma. Hepatology.
57:610–624. 2013. View Article : Google Scholar
|
|
29
|
Wang J, Ray PS, Sim MS, et al: FOXC1
regulates the functions of human basal-like breast cancer cells by
activating NF-κB signaling. Oncogene. 31:4798–4802. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Uramoto H, Iwata T, Onitsuka T, et al:
Epithelial-mesenchymal transition in EGFR-TKI acquired resistant
lung adenocarcinoma. Anticancer Res. 30:2513–2517. 2010.PubMed/NCBI
|
|
31
|
Bryant JL, Britson J, Balko JM, et al: A
microRNA gene expression signature predicts response to erlotinib
in epithelial cancer cell lines and targets EMT. Br J Cancer.
106:148–156. 2012. View Article : Google Scholar :
|
|
32
|
Cufi S, Bonavia R, Vazquez-Martin A, et
al: Silibinin suppresses EMT-driven erlotinib resistance by
reversing the high miR-21/low miR-200c signature in vivo. Sci Rep.
3:24592013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wu ZZ, Sun NK and Chao CC: Knockdown of
CITED2 using short-hairpin RNA sensitizes cancer cells to cisplatin
through stabilization of p53 and enhancement of p53-dependent
apoptosis. J Cell Physiol. 226:2415–2428. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Samatov TR, Tonevitsky AG and Schumacher
U: Epithelial-mesenchymal transition: focus on metastatic cascade,
alternative splicing, non-coding RNAs and modulating compounds. Mol
Cancer. 12:1072013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ying L, Chen Q, Wang Y, et al: Upregulated
MALAT-1 contributes to bladder cancer cell migration by inducing
epithelial-to-mesenchymal transition. Mol Biosyst. 8:2289–2294.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sequist LV, Waltman BA, Dias-Santagata D,
et al: Genotypic and histological evolution of lung cancers
acquiring resistance to EGFR inhibitors. Sci Transl Med.
3:75ra262011.PubMed/NCBI
|