Introduction
Histone acetylation modification is an important
epigenetic modification under the dual regulation of histone
acetyltransferases (HATs) and histone deacetylases (HDACs)
(1). HATs are important in the
regulation of genetic transcription, chromosome constitution, DNA
damage repair and cell cycle regulation (2–4). It is
closely associated with the occurrence and development of several
diseases such as cancer, neurodegeneration and inflammatory lung
disease (5–7). The p300 acetyltransferase serves as a
tumor suppressor and is mutated in epithelial malignancies
(8). HDAC1 is significantly
increased in hormone-refractory prostate cancer cell lines and
promotes the malignant transformation of cancer cells (9).
HATs are classified as: GNAT family
(Gcn5-related-N-acetyltransferase family), p300 (E1A-associated
protein of 300 kDa) or CBP (CREB-binding protein) and MYST family
members (10). As a MYST family
member, males absent on the first (MOF) is part of the dosage
compensation complex on the X chromosome of male Drosophila
and an important component of the male-specific lethal complex
(11,12). MOF impacts several physiological and
pathological processes such as mammalian embryonic development, the
maintenance of chromatin structure, and tumorigenesis. MOF
influences the chromatin structure of mouse embryonic cells and the
implantation of early embryos (13). MOF also plays an essential role in
the maintenance of embryonic stem cell self-renewal and
pluripotency (14).
Human MOF (hMOF) and Drosophila MOF are
orthologous, and hMOF specifically acetylates the histone H4K16
(15). The substrate specificity of
hMOF depends on its formation of different complexes. For example,
hMOF containing NSL complex can acetylate histone H4K16, H4K5 and
H4K8 (16). When the hMOF
gene is knocked down, H4K16 acetylation levels significantly
decrease (15,17). hMOF participates in basic
physiological processes of mammalian cells, including gene
transcription, cell proliferation and differentiation, nuclear
morphology, chromatic constitution and DNA repair (18). Therefore, the inactivation of hMOF
may be an important step in the malignant transformation of cells.
After the knockdown of the hMOF gene expression in HeLa and
HepG2 cells, nuclear morphology obviously changed into a
multi-leaved shape (17). However,
whether the nuclear morphological change was the result or cause of
the cellular malignant transformation remains to be determined.
The role of hMOF in the occurrence, development and
prognosis of malignant tumors has been investigated. Pfister et
al (19) found that hMOF and
H4K16 are frequently downregulated in primary breast carcinoma and
medulloblastoma. Furthermore, they found that hMOF protein
expression is a prognostic marker for medulloblastoma, with
patients harboring tumors with a low hMOF expression having a
significantly worse survival (19).
In a study on human renal cell carcinoma, Wang et al found
that the expression of hMOF in cancer tissues was significantly
lower than that in adjacent kidney tissues (20). However, hMOF was more frequently
found to be highly expressed in non-small cell lung cancer (NSCLC)
than corresponding normal tissues, and hMOF promoted the
proliferation, metastasis and adhesion of NSCLC cell lines
(21).
Although the low expression of hMOF in ovarian
cancer has been identified (22),
the relationship between the hMOF expression and ovarian cancer
prognosis has yet to be reported. hMOF may be important in the
occurrence, development and prognosis of ovarian cancer. Thus, the
expression of hMOF in different ovarian tissues and its
relationship with ovarian cancer prognosis were investigated.
Materials and methods
Patients and follow-up
Experiments were approved by the Institutional
Review Board of Shengjing Hospital at China Medical University. A
total of 45 samples were collected from tissues removed during the
surgical removal of cervical cancers at the Department of
Gynecology, Shengjing Hospital, China. The 45 samples comprised 30
samples of ovarian epithelial cancer tissues and 15 samples of
normal ovarian tissues. All the tissues were removed during surgery
whereupon they were immediately frozen in liquid nitrogen and
stored at −80°C. Based on the characteristics of ovarian carcinoma,
it is extremely difficult to obtain para-carcinoma tissue;
therefore, normal ovarian tissues were used as a control group.
Patients anonymity was maintained. The tissue samples were examined
by specialists to obtain a final diagnosis, and no patients were
administered chemotherapy or radiotherapy prior to surgery. The
clinicopathological characteristics of 30 patients of epithelial
ovarian cancer are shown in Table
I.
 | Table IClinicopathological characteristics of
30 patients with ovarian cancer. |
Table I
Clinicopathological characteristics of
30 patients with ovarian cancer.
| Characteristics | No. |
|---|
| Pathologic type |
| Serous | 20 |
| Mucinous | 2 |
| Clear cell
carcinoma | 1 |
| Poorly
differentiated adenocarcinoma | 7 |
| Surgical stage |
| I–II | 9 |
| III–IV | 21 |
| Differentiation |
| Well | 5 |
| Moderate-poor | 25 |
| Lymph node
metastasis |
| No | 27 |
| Yes | 3 |
In total, 45 fresh tissue samples were embedded into
paraffin sections and an additional 136 different paraffin slices
were obtained from the ovarian tissues resected during operations
at the Department of Gynecology, Shengjing Hospital, China Medical
University, China, from 2008 to 2012. A total of 181 paraffin
sections were thus obtained: 112 of primary malignant ovarian
tumors, 23 of borderline ovarian tumors, 26 of benign ovarian
tumors and 20 of normal ovarian tissues. The tissue sections were
examined by experienced specialists to obtain a final diagnosis.
Histopathological diagnoses were made using the World Health
Organization criteria. The classification of cancer stage and grade
was carried out according to the International Federation of
Gynecology and Obstetrics. The clinical and pathological
information concerning the patients was collected from their
clinical records, including their age, surgical stage, lymph node
metastasis, pathological tumor grade and subtype, and residual
tumor size.
The age range (median) was 16–77 years (52.7 years)
in the malignant ovarian tumor group; 21–78 years (41.5 years) in
the borderline ovarian tumor group; 24–61 years (43.2 years) in the
benign ovarian tumor group; and 45–68 years (58.9 years) in the
normal ovarian tissue group. There were no statistically
significant differences in the ages of these groups
(P>0.05).
Information was collected on the clinical
chemotherapeutic treatments received and the follow-up from 77
patients out of a total 112 patients with malignant ovarian cancer
(the information from 77 patients was complete and these 77
patients were followed-up for 24 months at least after surgery).
These 77 patients underwent treatments for ovarian cancer that
included surgical debulking followed by 6–8 postoperative cycles of
conventional chemotherapy, consisting of paclitaxel (Yangtze River
Pharmaceutical Group, Taizhou, China) and carboplatin (Qilu
Pharmaceutical Co., Ltd, Jinan, China). Overall survival (OS) was
defined as the date of surgery to the date of death or the last
follow-up. Disease-free survival (DFS) was defined as the interval
from the initial surgery to clinically or radiologically proven
recurrence/metastasis and deceased. Following surgery, the patients
were observed at 3-month intervals. As of May 2014, it has been 70
months since the first patient was recruited into the research
group, and it has been 24 months since the last patient was
recruited into our group. The median followup period was 48 months.
To determine the factors influencing survival after surgery and
standard chemotherapy, conventional variables, together with hMOF
expression, were assessed in 77 ovarian carcinoma patients.
Materials
The PrimeScript™ 1st Strand cDNA Synthesis and PCR
amplification kits were purchased from Takara (6110A, RR02A;
Dalian, China). Mouse monoclonal anti-hMOF antibody (GTX83065) was
obtained from GeneTex (Irvine, CA, USA). The anti-GAPDH
(glyceraldehyde-3-phosphate dehydrogenase) monoclonal antibody was
purchased from Boshide Biotech (BM1985; Wuhan, China).
Reverse transcription PCR (RT-PCR)
Total RNA from epithelial ovarian cancer and the
normal ovarian tissues were isolated using TRIzol® LS
reagent (Invitrogen, Carlsbad, CA, USA). RNA (500 ng) from each
sample was used as a template to produce cDNA using the
PrimeScript™ 1st Strand cDNA Synthesis kit. PCR reactions were
performed under the following conditions: an initial denaturation
step at 95°C for 2 min, followed by 30 cycles of denaturation step
at 95°C for 30 sec, annealing at 57°C for 30 sec and extension at
72°C for 30 sec. The primer sets used for the PCR were: GAPDH,
forward: 5′-ATCACTGCCACCCAGAAGAC-3′ and reverse:
5′-ATGAGGTCCACCACCCTGTT-3′, yielding a 433-bp product; hMOF,
forward: 5′-GACACTGTACTTTGACGTGGAGC-3′ and reverse:
5′-CACTGTGATGGGTGGTTTCTT-3′, yielding a 493-bp product.
Immunoblotting
Soluble proteins were isolated from tissues for
western blotting. The protein concentrations were measured by
bicinchoninic acid (23228; Thermo Fisher Scientific, Waltham, MA,
USA). Equal amounts of protein from each sample were separated by
electrophoresis on an SDS-10% polyacrylamide gel, transferred to a
polyvinylidene difluoride membranes, and blocked with 5% non-fat
dry milk in 1X TBS plus 0.1% Tween-20 at room temperature for 2 h.
The membranes were incubated overnight at 4°C with primary
antibodies in 1% bovine serum albumin in 1X TBS plus 0.1% Tween-20.
The primary anti-hMOF monoclonal antibody (diluted 1:1,000) was
purchased from GeneTex (GTX83065) and the anti-GAPDH monoclonal
antibody (diluted 1:2,000) was purchased from Boshide Biotech
(BM1985). Membranes were washed and incubated again for 2 h at room
temperature with horseradish-peroxidase-conjugated anti-mouse
secondary antibodies. Proteins were visualized with ECL reagent
(ECL Prime Western Blotting Detection Reagent; Amersham,
Pittsburgh, PA, USA). The experiments were repeated three
times.
Immunohistochemistry
Paraffin-embedded histological sections from each
group of ovarian tissues were cut into 5-μm slices.
Immunohistochemistry was used to analyze the hMOF protein
expression levels. Mouse monoclonal anti-hMOF antibody (GTX83065;
diluted 1:800) was purchased from the GeneTex. The staining
procedure was performed according to the manual of an
ultrasensitive streptavidin-peroxidase kit (KIT-9701; Maixin Bio,
Fuzhou, China) and Harris’s hematoxylin was used to stain the cell
nuclei. Tissues were treated with phosphate-buffered saline instead
of primary antibody as a negative control. Buff-colored granules in
the cell nucleus were considered a positive result. The tissues
were rated according to their chromatic intensity: no pigmentation,
0; light yellow, 1; buff, 2; and brown, 3. Five high-power fields
in serial sections from each slice were selected, scored, and the
mean percentage of chromatic cells was estimated: <5% chromatic
cells, 0; 5–25% chromatic cells, 1; 26–50% chromatic cells, 2;
51–75% chromatic cells, 3; and >75% chromatic cells, 4. The two
numbers (intensity score × percentage chromatic cells) were
multiplied and scored: 0–2 was considered (−), 3–4 (+), 5–8 (++)
and 9–12 (+++). The 112 patients of ovarian cancer were divided
into the high hMOF expression group (++/+++) and the low hMOF
expression group (−/+). Two observers read the sections to control
for systemic error.
Statistical analyses
The gene expression and western blot images were
scanned and quantified with ImageJ software (National Institutes of
Health, Bethesda, MD, USA). Differences in proportions were
evaluated using the χ2 test. The χ2 test or
Fisher’s exact test, whichever was appropriate, was used to analyze
the relationship between hMOF expression and clinicopathological
variables. A survival curve was generated using the Kaplan-Meier
method and compared by the log-rank test. Cox’s proportional hazard
regression model was used for multivariate survival analysis of
prognostic factors. Statistical analyses were performed using SPSS
v17.0 software (SPSS, Inc., Chicago, IL, USA). The Student’s
two-tailed t-test was used in all analyses. P<0.05 was
considered to indicate a statistically significant result.
Results
Reduced hMOF mRNA expression in ovarian
cancer
hMOF plays a role in the basic physiological
processes of mammalian cells and therefore, it may be involved in
different malignant tumors. Consequently, we hypothesized that hMOF
may be important in the occurrence and development of ovarian
cancer and that the role of hMOF may differ between ovarian cancer
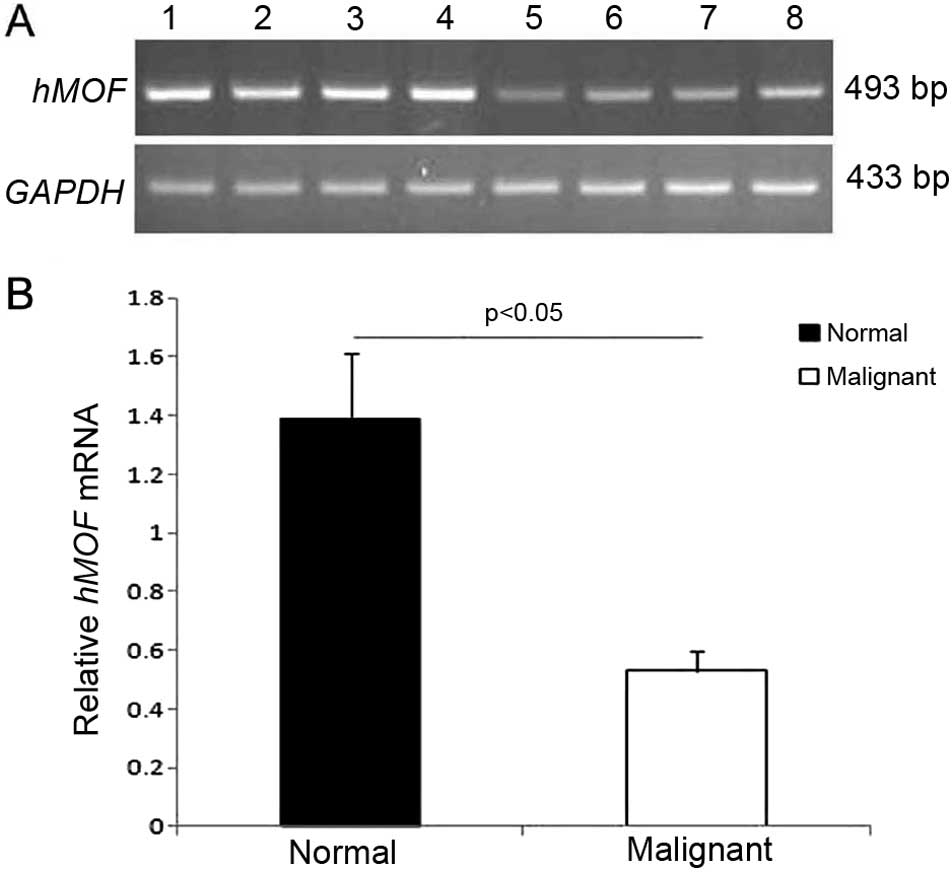
and normal ovarian tissues. RT-PCR showed that the expression of
hMOF mRNA in normal ovarian tissues (15 cases) was 2.62-fold higher
than that in ovarian cancer tissues (30 cases) (P<0.05; Fig. 1A and B).
Reduced hMOF protein expression in
ovarian cancer
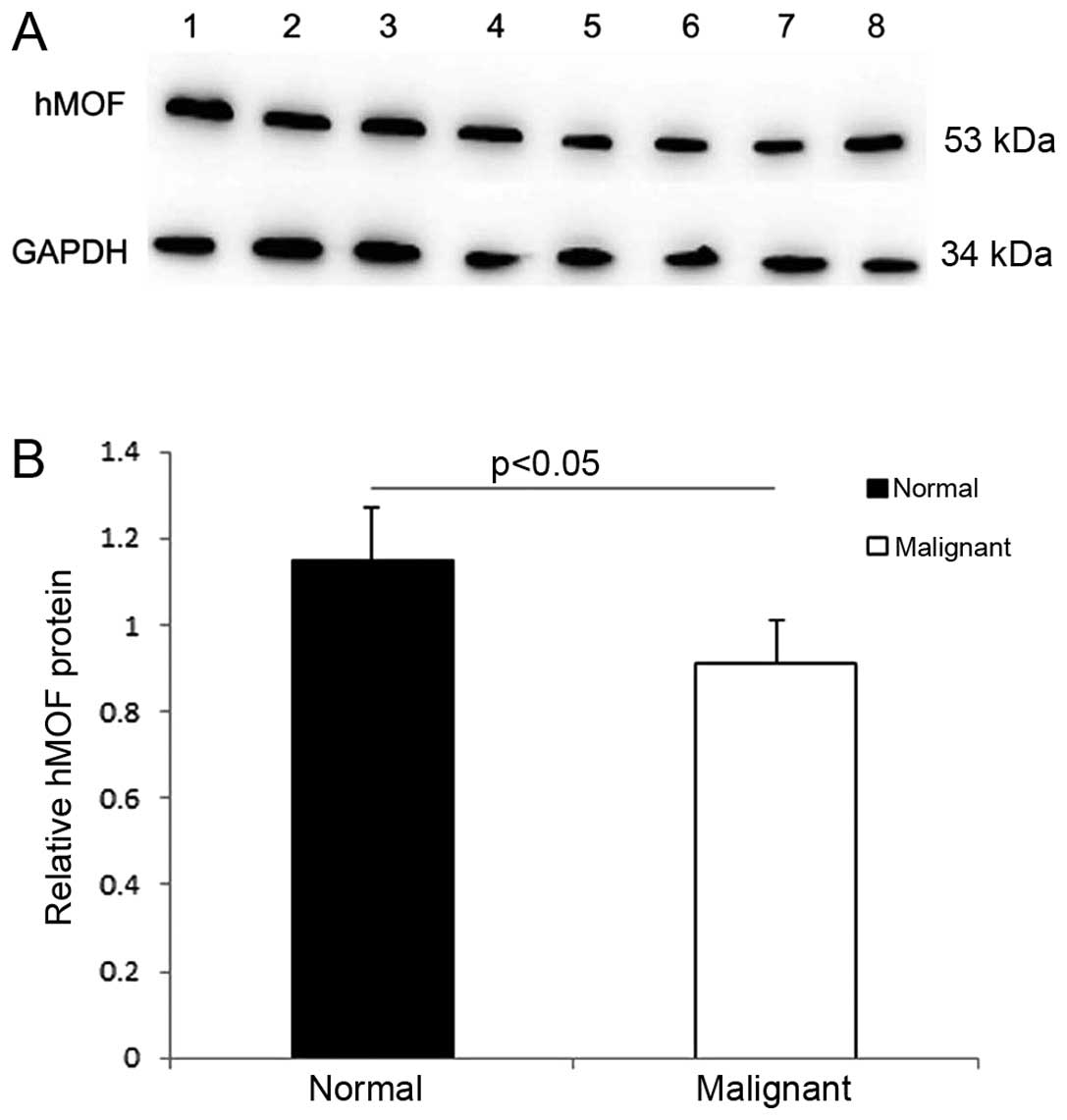
The results from the western blot analysis and
immunohistochemical methods used to investigate hMOF protein
expression in different ovarian tissues, confirmed the results from
the mRNA expression analysis. Western blot analysis showed that
hMOF protein expression in the normal ovarian tissues was 1.28-fold
higher than that in ovarian cancer tissues (P<0.05; Fig. 2A and B).
Immunohistochemical analysis of hMOF
protein expression in different ovarian tissues
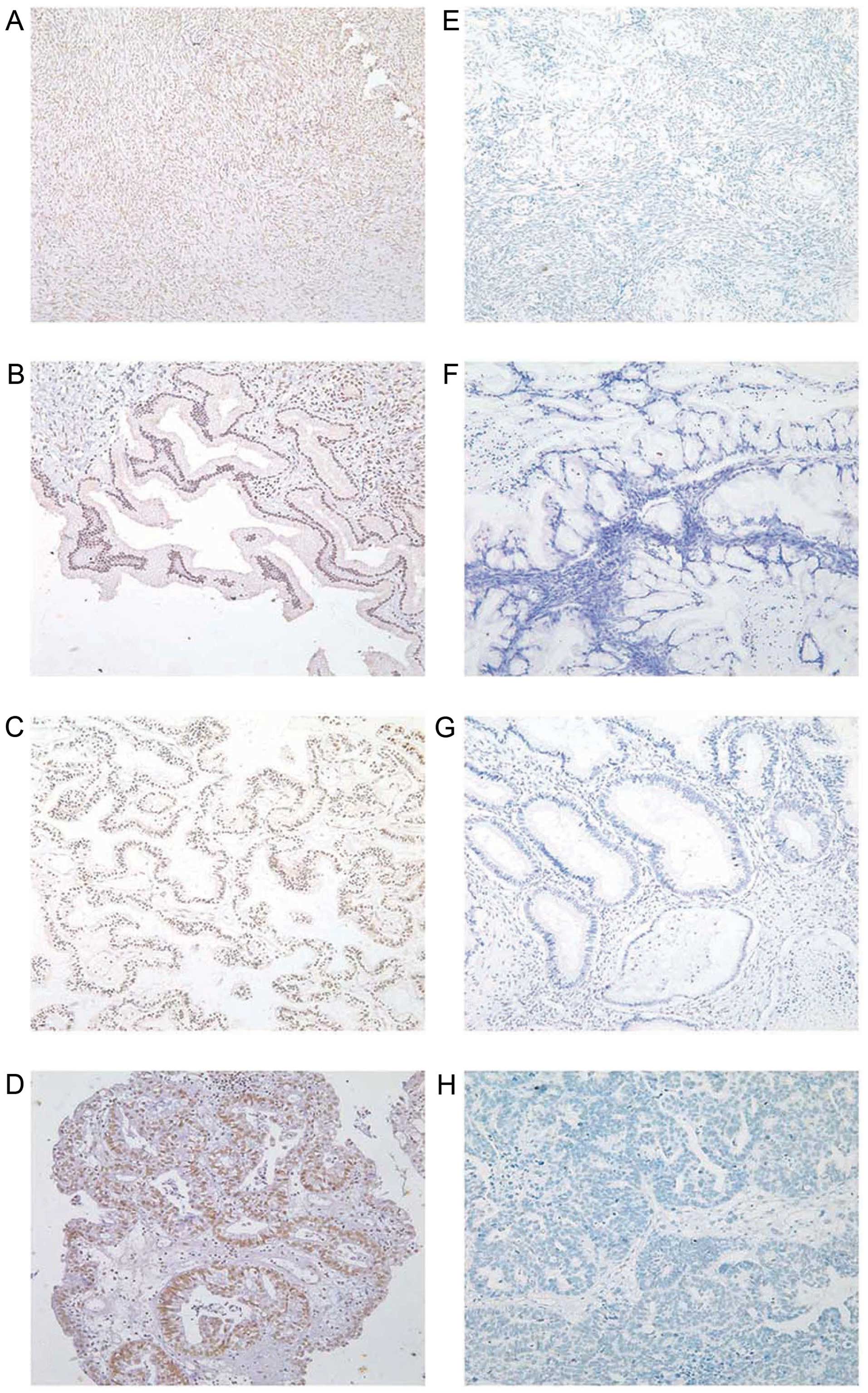
hMOF immunoreactivity was identified in the nucleus
(Fig. 3) and in all tissue types,
and hMOF was expressed in the nucleus. The positive hMOF protein
expression rates in ovarian epithelial cancer, borderline tumor,
benign tumor and normal ovarian tissues were 80.36, 65.22, 84.62
and 80.00%, respectively (Table
II). The positive rates did not significantly differ between
groups. Data were divided into a low hMOF expression group
(including negative and weak positive expression) and a high hMOF
expression group (including moderate and strong positive
expression). According to the standard scoring of paraffin slices,
3–4 was defined as weak expression and 5–8 was defined as moderate
expression. The specific scoring criteria was as described in
Materials and methods. Further analysis of the data found that the
proportions of high hMOF expression in ovarian epithelial cancer,
borderline tumor, benign tumor and normal ovarian tissues, were
27.68, 30.43, 57.69 and 50.00%, respectively (Table II). Therefore, hMOF protein
expression in ovarian benign tumor tissues and normal ovarian
tissues was much higher than that in ovarian epithelial cancer
tissues (P<0.05).
| Table IIhMOF in different ovarian tissues. |
Table II
hMOF in different ovarian tissues.
| | hMOF expression | | hMOF expression |
|---|
| |
| |
|
|---|
| Groups | Cases | − | + | ++ | +++ | Positive rates
(%) | Low (−/+) | High (++/+++) |
|---|
| Malignant | 112 | 22 | 59 | 16 | 15 | 80.36 | 81 (72.32%) | 31 (27.68%)a |
| Borderline | 23 | 8 | 8 | 3 | 4 | 65.22 | 16 (69.57) | 7 (30.43%) |
| Benign | 26 | 4 | 7 | 6 | 9 | 84.62 | 11 (42.31%) | 15 (57.69%) |
| Normal | 20 | 4 | 6 | 4 | 6 | 80.00 | 10 (50.00%) | 10 (50.00%) |
Relationship between hMOF expression and
clinicopathological data on ovarian cancer
A total of 112 ovarian cancer patients were divided
into the high (++/+++) and low (−/+) hMOF protein-expression
groups. The rate of high hMOF protein expression in stage I ovarian
cancer tissues was 33.33%, significantly higher than its expression
in stage IV ovarian cancer tissues (P<0.05). The protein
expression of hMOF in ovarian endometrioid adenocarcinoma tissues
was significantly higher than its expression in ovarian serous
adenocarcinoma tissues (P<0.05). There was no relationship
between hMOF protein expression with cell differentiation and lymph
node metastasis (P>0.05; Table
III).
| Table IIIRelationships between hMOF protein
expression and clinicopathological characteristics of 112 patients
with malignant ovarian cancer. |
Table III
Relationships between hMOF protein
expression and clinicopathological characteristics of 112 patients
with malignant ovarian cancer.
| | hMOF
expression | |
|---|
| |
| |
|---|
|
Characteristics | No. cases | Low
(−/+)
(%) | High
(++/+++)
(%) | P-value |
|---|
| Pathological
type | | | | <0.05 |
| Serous | 48 | 37 (77.08) | 11 (22.92)a | |
| Mucinous | 19 | 13 (68.42) | 6 (31.58) | |
| Endometrioid | 8 | 3 (37.50) | 5 (62.50) | |
| Clear cell
carcinoma | 10 | 8 (80.00) | 2 (20.00) | |
| Poorly
differentiated adenocarcinoma | 27 | 20 (74.07) | 7 (25.93) | |
| Surgical stage | | | | <0.05 |
| I | 36 | 24 (66.67) | 12 (33.33)b | |
| II | 13 | 11 (84.61) | 2 (11.39) | |
| III | 57 | 40 (70.18) | 17 (29.82) | |
| IV | 6 | 6 (100.00) | 0 (0.00) | |
|
Differentiation | | | | >0.05 |
| Well | 16 | 10 (62.50) | 6 (37.50) | |
| Moderate | 30 | 24 (80.00) | 6 (20.00) | |
| Moderate-poor | 19 | 13 (68.42) | 6 (31.58) | |
| Poor | 47 | 34 (72.34) | 13 (27.66) | |
| Lymph node
metastasisc | | | | >0.05 |
| No | 90 | 62 (68.89) | 28 (31.11) | |
| Yes | 17 | 14 (82.35) | 3 (17.65) | |
Multivariate prognostic analysis of
ovarian cancer
A total of 77 patients with complete follow-up data
were divided into the low (−/+) and high (++/+++) hMOF
protein-expression groups. The COX proportional-hazards regression
model was applied with survival time as an independent variable and
dependent variables including the age of ovarian cancer patients,
surgical pathological staging, differentiation, pathological
category, lymph-node metastasis, residual tumor size and hMOF
expression. Multivariate analysis revealed that the age of ovarian
cancer patients and hMOF expression were independent prognostic
risk factors for OS (Table IV),
while residual tumor size and hMOF expression were the independent
risk factors closely associated with DFS (Table IV).
| Table IVCox proportional hazards regression
model. |
Table IV
Cox proportional hazards regression
model.
| Overall survival
(OS) | Disease-free
survival (DFS) |
|---|
|
|
|
|---|
| Variables | Hazard ratio (95%
CI) | P-value | Hazard ratio (95%
CI) | P-value |
|---|
| hMOF | 0.337
(0.125–0.903) | 0.031 | 0.307
(0.114–0.829) | 0.020 |
| Age (years) | 0.381
(0.158–0.919) | 0.032 | 0.451
(0.190–1.067) | 0.070 |
| Surgical stage | 1.588
(0.657–1.255) | 0.558 | 1.820
(0.701–4.729) | 0.219 |
|
Differentiation | 1.015
(0.390–2.642) | 0.976 | 1.073
(0.426–2.706) | 0.881 |
| Pathological
type | 0.908
(0.657–1.255) | 0.705 | 0.938
(0.673–1.307) | 0.705 |
| Lymph node
metastasis | 1.905
(0.724–5.012) | 0.191 | 2.394
(0.876–6.545) | 0.089 |
| Residual lesion
size | 1.719
(0.953–3.100) | 0.072 | 1.877
(1.029–3.461) | 0.040 |
Comparison of survival rate of ovarian
cancer patients
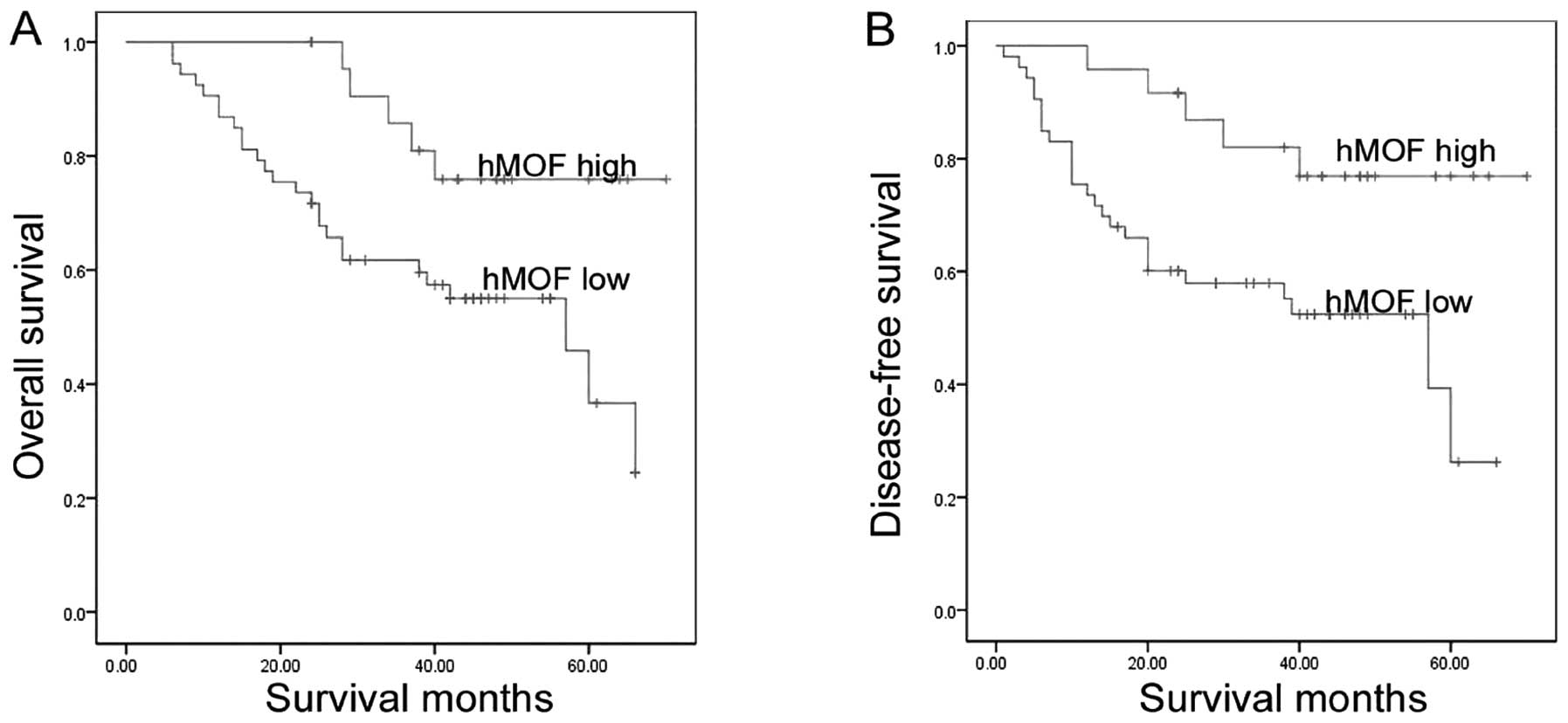
As of May 2014 (70 months since the recruitment of
the first patient and 24 months since recruitment of the last
patient), 26 patients in the low hMOF-expression group (comprising
53 patients) succumbed, while five patients in the high
hMOF-expression group (comprising 24 patients) succumbed. A
Kaplan-Meier analysis (Fig. 4)
showed that the OS and DFS rates were significantly higher in the
high hMOF expression group compared to the low hMOF expression
group (log-rank test; OS, P=0.018 and DFS, P=0.009).
Discussion
hMOF, a MYST (Moz-Ybf2/Sas3-Sas2-Tip60) family
member, specifically acetylates the histone H4K16 and regulates
basic physiological processes such as chromosome structure
maintenance, transcriptional regulation and DNA repair (12,13).
In recent years, the effects of hMOF on the occurrence and
development of malignant tumors has been a hot area of study in
international academia. Previous studies indicated that hMOF is
frequently downregulated in primary breast carcinoma and
medulloblastoma (19), human renal
cell (20), and colorectal
carcinoma, and gastric cancer (23). However, the expression of hMOF was
reportedly much higher in NSCLC compared to corresponding normal
tissues, and hMOF promoted the proliferation, metastasis and
adhesion of NSCLC cell lines (21).
Therefore hMOF may play different roles in various malignant
tumors. In the present study, the hMOF mRNA and protein expression
in ovarian epithelial cancer tissues was much lower than that of
normal ovarian tissues. The paraffin sections of different tissues
from 181 patients were analyzed by immunohistochemistry (primary
ovarian epithelial cancer tissues from 112 patients, borderline
tumor tissues from 23 patients, benign tumor tissues from 26
patients, and normal ovarian tissues from 20 patients). The results
indicated that hMOF protein expression in normal ovarian tissues
and benign tumor tissues was significantly higher compared to the
ovarian epithelial cancer tissues. The relationship between hMOF
expression and clinicopathological data of ovarian cancer patients
was further investigated and hMOF expression in stage I ovarian
cancer tissues was much higher than that in stage IV ovarian cancer
tissues. This finding suggests that the high expression of hMOF in
ovarian cancer tissues is associated with protection from advanced
ovarian cancer.
Pfister et al analyzed various factors that
may affect the prognosis of medulloblastoma, and found that hMOF
protein expression was a prognostic maker for medulloblastoma
(19). However, no previous study
has focused on the relationship between hMOF protein expression and
ovarian cancer prognosis. To the best of our knowledge, the present
study is the first to examine the relationship between hMOF protein
expression and ovarian cancer prognosis. The Cox
proportional-hazards regression model indicated that hMOF protein
expression was an independent risk factor closely associated with
ovarian cancer prognosis. In addition, the Kaplan-Meier survival
analysis based on the intensity of hMOF expression found that the
overall survival rate and disease-free survival rate in the high
hMOF expression group was significantly higher than that in the low
hMOF expression group. Thus, hMOF may serve as a
tumor-suppressor gene for ovarian cancer and its protein expression
may be a new clinical marker that reflects ovarian cancer
prognosis.
hMOF plays an important role in many aspects of
processes such as repair of DNA damage, maintenance of nuclear
structure and morphology, regulation of genetic transcription,
apoptosis and drug resistance of cells. A decrease in the
expression of hMOF may have important effects on the occurrence,
development and prognosis of malignant tumors. Therefore the
specific molecular mechanism responsible for the decreased
expression of hMOF in this study should be further investigated.
hMOF is important in the activation of ataxia telangiectasia
mutated (ATM). When the cells are exposed to ionizing radiation,
hMOF protein expression significantly increases and the ATM may
become activated. When the hMOF gene expression is decreased
by siRNAs, ATM autophosphorylation and ATM kinase activity may be
significantly reduced, leading to a decrease in DNA repair capacity
(24). The hMOF complex contains
the microporous component of the nucleus (25). Therefore, deficiencies in the
expression of hMOF may affect the morphology and stability of the
nuclear membrane, and convert cells into a malignant phenotype. In
addition, hMOF can acetylate p53 at K120, resulting in the
increased transcription of BAX and PUMA and cell
apoptosis (26). It has been
suggested that after hMOF expression decreases, multidrug-resistant
cancer cells are less sensitive to the topoisomerase II inhibitors
(27).
In the present study, the expression of hMOF mRNA
and protein was significantly downregulated in ovarian epithelial
cancer tissues, and patients with high hMOF levels showed improved
survival as compared to those with low hMOF levels. Future studies
should address the mechanisms hMOF may use to inhibit the
development of ovarian cancer and the potential of hMOF as a new
target for ovarian cancer treatment.
Acknowledgements
This study was supported by grants from the National
Natural Science Foundation of China (nos. 81172491 and 81101527),
the Education Department Doctor Project Fund (nos. 20112104110016
and 20112104120019); the Free Researcher Plan of Shengjing Hospital
(no. 201303), and the Scientific Research Projects of Liaoning
Province Department of Education (no. L2011129).
References
|
1
|
Strahl BD and Allis CD: The language of
covalent histone modifications. Nature. 403:41–45. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Carrozza MJ, Utley RT, Workman JL and Côté
J: The diverse functions of histone acetyltransferase complexes.
Trends Genet. 19:321–329. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yamada T, Mizuno K, Hirota K, et al: Roles
of histone acetylation and chromatin remodeling factor in a meiotic
recombination hotspot. EMBO J. 23:1792–1803. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Eberharter A and Becker PB: Histone
acetylation: a switch between repressive and permissive chromatin.
Second in review series on chromatin dynamics. EMBO Rep. 3:224–229.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yang XJ: The diverse superfamily of lysine
acetyltransferases and their roles in leukemia and other diseases.
Nucleic Acids Res. 32:959–976. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Saha RN and Pahan K: HATs and HDACs in
neurodegeneration: a tale of disconcerted acetylation homeostasis.
Cell Death Differ. 13:539–550. 2006. View Article : Google Scholar
|
|
7
|
Ito K, Charron CE and Adcock IM: Impact of
protein acetylation in inflammatory lung diseases. Pharmacol Ther.
116:249–265. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gayther SA, Batley SJ, Linger L, et al:
Mutations truncating the EP300 acetylase in human cancers. Nat
Genet. 24:300–303. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Halkidou K, Gaughan L, Cook S, Leung HY,
Neal DE and Robson CN: Upregulation and nuclear recruitment of
HDAC1 in hormone refractory prostate cancer. Prostate. 59:177–189.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yang XJ and Seto E: Lysine acetylation:
codified crosstalk with other posttranslational modifications. Mol
Cell. 31:449–461. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hilfiker A, Hilfiker-Kleiner D, Pannuti A
and Lucchesi JC: mof, a putative acetyl transferase gene related to
the Tip60 and MOZ human genes and to the SAS genes of yeast, is
required for dosage compensation in Drosophila. EMBO J.
16:2054–2060. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Akhtar A and Becker PB: Activation of
transcription through histone H4 acetylation by MOF, an
acetyltransferase essential for dosage compensation in Drosophila.
Mol Cell. 5:367–375. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Thomas T, Dixon MP, Kueh AJ and Voss AK:
Mof (MYST1 or KAT8) is essential for progression of embryonic
development past the blastocyst stage and required for normal
chromatin architecture. Mol Cell Biol. 28:5093–5105. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li X, Li L, Pandey R, et al: The histone
acetyltransferase MOF is a key regulator of the embryonic stem cell
core transcriptional network. Cell Stem Cell. 11:163–178. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Smith ER, Cayrou C, Huang R, Lane WS, Côté
J and Lucchesi JC: A human protein complex homologous to the
Drosophila MSL complex is responsible for the majority of histone
H4 acetylation at lysine 16. Mol Cell Biol. 25:9175–9188. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cai Y, Jin J, Swanson SK, et al: Subunit
composition and substrate specificity of a MOF-containing histone
acetyltransferase distinct from the male-specific lethal (MSL)
complex. J Biol Chem. 285:4268–4272. 2010. View Article : Google Scholar :
|
|
17
|
Taipale M, Rea S, Richter K, et al: hMOF
histone acetyltransferase is required for histone H4 lysine 16
acetylation in mammalian cells. Mol Cell Biol. 25:6798–6810. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rea S, Xouri G and Akhtar A: Males absent
on the first (MOF): from flies to humans. Oncogene. 26:5385–5394.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pfister S, Rea S, Taipale M, et al: The
histone acetyltransferase hMOF is frequently downregulated in
primary breast carcinoma and medulloblastoma and constitutes a
biomarker for clinical outcome in medulloblastoma. Int J Cancer.
122:1207–1213. 2008. View Article : Google Scholar
|
|
20
|
Wang Y, Zhang R, Wu D, et al: Epigenetic
change in kidney tumor: downregulation of histone acetyltransferase
MYST1 in human renal cell carcinoma. J Exp Clin Cancer Res.
32:82013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhao L, Wang DL, Liu Y, Chen S and Sun FL:
Histone acetyltransferase hMOF promotes S phase entry and
tumorigenesis in lung cancer. Cell Signal. 25:1689–1698. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu N, Zhang R, Zhao X, et al: A potential
diagnostic marker for ovarian cancer: Involvement of the histone
acetyltransferase, human males absent on the first. Oncol Lett.
6:393–400. 2013.PubMed/NCBI
|
|
23
|
Cao L, Zhu L, Yang J, et al: Correlation
of low expression of hMOF with clinicopathological features of
colorectal carcinoma, gastric cancer and renal cell carcinoma. Int
J Oncol. 44:1207–1214. 2014.PubMed/NCBI
|
|
24
|
Gupta A, Sharma GG, Young CS, et al:
Involvement of human MOF in ATM function. Mol Cell Biol.
25:5292–5305. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mendjan S, Taipale M, Kind J, et al:
Nuclear pore components are involved in the transcriptional
regulation of dosage compensation in Drosophila. Mol Cell.
21:811–823. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sykes SM, Mellert HS, Holbert MA, et al:
Acetylation of the p53 DNA-binding domain regulates apoptosis
induction. Mol Cell. 24:841–851. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hajji N, Wallenborg K, Vlachos P,
Füllgrabe J, Hermanson O and Joseph B: Opposing effects of hMOF and
SIRT1 on H4K16 acetylation and the sensitivity to the topoisomerase
II inhibitor etoposide. Oncogene. 29:2192–2204. 2010. View Article : Google Scholar : PubMed/NCBI
|