Introduction
Colorectal cancer (CRC) is one of the most common
malignancies with high incidence in Western countries (1). At present, 5-fluorouracil-based
regimens have been performed as international standard chemotherapy
for CRC treatment (2). However,
5-FU-based regimens are often coupled with serious toxicity and
side effects such as anemia, leucopenia, thrombocytopenia and
peripheral neuropathy (3–5). Thus, the development of novel
therapeutic drugs is necessary. Natural products, including
traditional Chinese medicines (TCM), have been considered as
alternative cancer remedies for many years. Numerous plants and
their constituents have been shown to possess beneficial
therapeutic effects for various diseases including cancer (6,7).
Patrinia scabiosaefolia (PS), a perennial plant natively
distributed in Eastern Asia, has long been used in China to
clinically treat edema, appendicitis, endometritis and other
inflammatory diseases (8–10). More importantly, PS has also been
used as a major component in several TCM formulas for the treatment
of gastrointestinal cancers (11,12).
We recently reported that PS can inhibit CRC growth through
induction of apoptosis and inhibition of tumor angiogenesis
(12–14). To further elucidate the mode of
action of PS, in the present study we used a CRC mouse xenograft
model and a human CRC cell line HT-29 to evaluate the effect of the
ethanol extract of PS (EEPS) on cancer cell proliferation and
investigated the underlying molecular mechanisms.
Materials and methods
Materials and reagents
Dulbecco’s modified Eagle’s medium (DMEM), fetal
bovine serum (FBS), penicillin-streptomycin, trypsin-EDTA, and
TRIzol reagent were purchased from Life Technologies (Carlsbad, CA,
USA). SuperScript II Reverse Transcriptase was obtained from
Promega Corporation (Madison, WI, USA). PCNA assay kit was
purchased from R&D Systems (Minneapolis, MN, USA). CDK4 and
CyclinD1, β-actin antibodies, and horseradish peroxidase
(HRP)-conjugated secondary antibodies were obtained from Cell
Signaling Technology, Inc. (Beverly, MA, USA). A cell cycle kit was
purchased from BD Biosciences (San Jose, CA, USA). All other
chemicals, unless otherwise stated, were obtained from
Sigma-Aldrich (St. Louis, MO, USA).
Preparation of ethanol extract from
PS
Ethanol extract from Patrinia scabiosaefolia
(EEPS) was prepared as described previously (12). For animal experiments, EEPS powder
was dissolved in saline to a working concentration of 250 mg/ml. In
cell-based experiments, EEPS powder was dissolved in 50% DMSO to a
stock concentration of 250 mg/ml, and the EEPS working
concentrations were obtained by diluting the stock solution in the
culture medium. The final concentration of DMSO in the medium for
all cell experiments was <0.5%.
Cell culture
Human CRC HT-29 cells were obtained from the cell
bank of the Chinese Academy of Science (Shanghai, China). The cells
were grown in DMEM containing 10% (v/v) FBS, 100 U/ml penicillin,
and 100 μg/ml streptomycin. Cells were cultured at 37°C in a
humidified incubator with 5% CO2.
Animals
Male BALB/c athymic (nude) mice (with an initial
body weight of 20–22 g) were obtained from the Shanghai SLAC
Laboratory Animal Co., Ltd. (Shanghai, China) and housed under
pathogen-free conditions with controlled temperature (22°C),
humidity, and a 12-h light/dark cycle. Food and water were given
ad libitum throughout the experiment. All animal treatments
were strictly in accordance with international ethical guidelines
and the National Institutes of Health Guide concerning the Care and
Use of Laboratory Animals. The experiments were approved by the
Institutional Animal Care and Use Committee of Fujian University of
Traditional Chinese Medicine.
In vivo tumor xenograft study
HT-29 cells (1.5×106) mixed with Matrigel
(1:1) were subcutaneously injected into the right flank of the mice
to initiate tumor growth. After 5 days of xenograft implantation,
mice were randomly divided into 2 groups (n=6) and given
intragastric administration with 1.93 g/kg/day of EEPS or saline
daily, 5 days per week for 3 weeks. Tumor size was determined by
measuring the major (L) and minor (W) diameter with a caliper. The
tumor volume was calculated according to the following formula:
Tumor volume = π/6 × L × W2.
Immunohistochemical staining (IHS)
After being fixed with 10% formaldehyde for 12 h,
the tumor samples were processed conventionally for
paraffin-embedded 4-μm-thick tumor slides. The slides were
subjected to antigen retrieval, and the endogenous peroxidase
activity was quenched with hydrogen peroxide. After blocking
non-specific proteins with normal serum in phosphate-buffered
saline (PBS) (0.1% Tween-20), the slides were incubated with rabbit
polyclonal antibodies against PCNA (1:200 dilution) for 1 h. After
washing with PBS, slides were incubated with a biotinylated
secondary antibody followed by conjugated HRP-labeled streptavidin
(Dako), and then washed with PBS. The slides were then incubated
with diaminobenzidine (Sigma) as the chromogen, followed by
counterstaining with diluted Harris’ hematoxylin (Sigma). After
staining, five high-power fields (x400 magnification) were randomly
selected in each slide, and the average proportion of positive
cells in each field was counted using a true color multi-functional
cell image analysis management system (Image-Pro Plus; Media
Cybernetics, Rockville, MD, USA). To rule out any nonspecific
staining, PBS was used to replace the primary antibody as a
negative control.
Evaluation of cell viability by MTT
assay
The HT-29 cell viability was assessed by MTT
colorimetric assay. HT-29 cells were seeded into 96-well plates at
a density of 1×104 cells/well in 0.1-ml medium. The
cells were treated with various EEPS concentrations for 24 h. At
the end of the treatment, 100 μl MTT (0.5 mg/ml in PBS) was added
to each well, and the samples were incubated for an additional 4 h
at 37°C. The purple-blue MTT formazan precipitate was dissolved in
100 μl DMSO. The absorbance was measured at 570 nm using an ELISA
reader (BioTek, Model ELX800, BioTek Instruments, Inc., Winooski,
VT, USA).
Colony formation assay
HT-29 cells were seeded in 6-well plates at a
density of 2×105 cells/well in 2 ml medium. After
treatment with various EEPS concentrations for 24 h, the cells were
collected and diluted in fresh medium in the absence of EEPS and
then reseeded in 6-well plates at a density of 1×103
cells/well. Following incubation for 7 to 8 days in a 37°C
humidified incubator with 5% CO2, the formed colonies
were fixed with 4% paraformaldehyde for 15 min, stained with 0.01%
crystal violet, and counted. The numbers of colonies were observed,
and the data were normalized to the number of control cells
(100%).
Cell cycle analysis
Cell cycle was analyzed by flow cytometry using
fluorescence-activated cell sorting (FACSCalibur; Becton-Dickinson,
San Jose, CA, USA) and propidium iodide (PI) staining. Subsequent
to treatment with various EEPS concentrations (0, 0.5, 1 and 2
mg/ml) for 24 h, HT-29 cells were collected and adjusted to a
concentration of 5×105 cells/ml, and fixed in 70%
ethanol at 4°C overnight. The fixed cells were washed twice with
cold PBS and then incubated for 30 min with RNase (8 μg/ml) and PI
(10 μg/ml). The fluorescent signal was detected through the FL2
channel, and the DNA proportion in different phases was analyzed
using ModfitLT ver. 3.0 (Verity Software House, Topsham, ME,
USA).
RNA extraction and RT-PCR analysis
Total RNA was isolated from the tumor tissues or
HT-29 cells with TRIzol reagent. Oligo(dT)-primed RNA (1 μg) was
reverse-transcribed with SuperScript II reverse transcriptase
according to the manufacturer’s instructions. The obtained cDNA was
used to determine the mRNA amount of CDK4 and CyclinD1 by PCR.
GAPDH was used as an internal control. The sequences of the primers
used for amplification of CyclinD1 and CDK4 were as follows: Cyclin
D1 forward, 5′-GGA GCA GAA GTG CGA AGA-3′ and reverse, 5′-GGG TGG
GTT GGA AAT GAA-3′ (Tm = 57°C, 394 bp); CDK 4 forward, 5-CTT CCC
GTC AGC ACA GTTC-3 and reverse, 5′-GGT CAG CAT TTC CAG TAGC-3′ (Tm
= 55°C, 687 bp); GAPDH forward, 5′-CGA CCA CTT TGT CAA GCTCA-3′ and
reverse, 5′-AGG GGT CTA CAT GGC AACTG-3′ (Tm = 58°C, 240 bp).
Samples were analyzed by gel electrophoresis (1.5% agarose). The
DNA bands were examined using a Gel Documentation system (Model Gel
Doc 2000; Bio-Rad, Hercules, CA, USA).
Western blot analysis
Tumor tissues were homogenized in non-denaturing
lysis buffer and centrifuged at 14,000 × g for 15 min. Protein
concentrations of the clarified supernatants were determined by BCA
protein assay. HT-29 (1×106) cells were seeded into
culture flasks in 5 ml medium and treated with various
concentrations of EEPS for 24 h. Treated cells were lysed in
mammalian cell lysis buffer (M-PER, Thermo Scientific, Rockford,
IL, USA) containing protease (EMD Biosciences) and phosphatase
inhibitor (Sigma-Aldrich) cocktail and centrifuged at 14,000 × g
for 15 min. Protein concentrations in the cell lysate supernatants
were determined by BCA protein assay and resolved on 12%
Tris-glycine gels. Equal amounts of protein from each tumor or cell
lysate were separated by 10% SDS-PAGE gels and transferred onto
PVDF membranes. Membranes were blocked for 2 h with blocking
solution at room temperature, washed in TBS with 0.25% Tween-20
(TBS-T), and exposed to primary antibodies against CDK4, CyclinD1
and β-actin (all at 1:1,000 dilutions) overnight at 4°C. After the
membranes were washed in TBS-T, secondary HRP-conjugated antibodies
(anti-rabbit or anti-mouse) were added at a 1:2,000 dilution for 1
h at room temperature, and the membranes were washed again in TBS-T
followed by enhanced chemiluminescence detection.
Statistical analysis
Data are presented as means ± SD for the indicated
number of independently performed experiments and analyzed using
the SPSS package for Windows (ver. 18.0). Statistical analysis of
the data was performed with the Student’s t-test and ANOVA.
Differences with P<0.05 were considered statistically
significant.
Results and Discussion
EEPS inhibits cancer cell proliferation
in CRC xenograft mice
Cancer cells are characterized by an uncontrolled
increase in cell proliferation (15); therefore inhibiting excessive
proliferation of tumor cells is one of the key approaches for
development of anticancer drugs. We first evaluated the efficacy of
EEPS against tumor growth in vivo in the CRC mouse xenograft
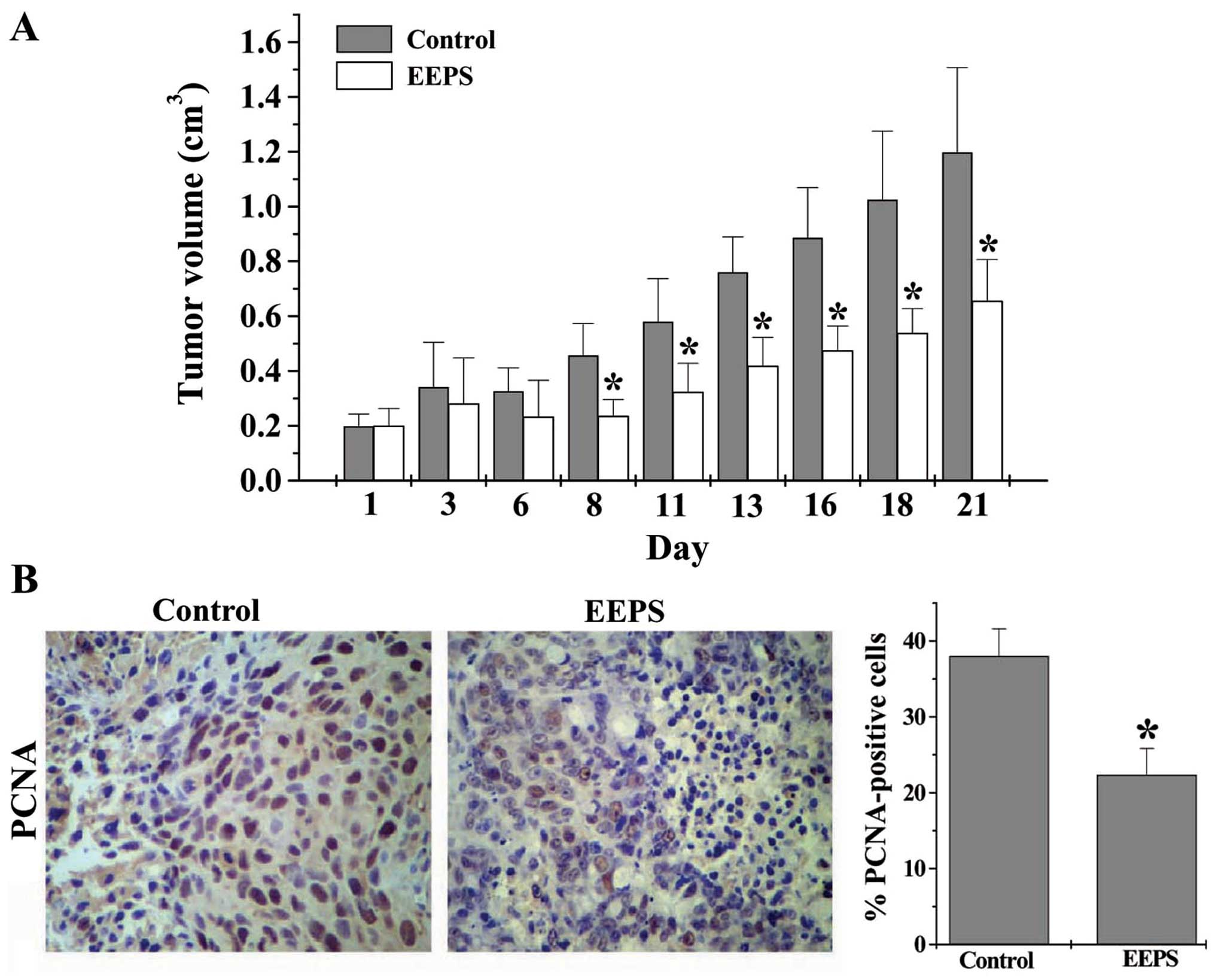
model. As shown in Fig. 1A, EEPS
treatment significantly reduced the tumor volume in CRC mice
(P<0.05, vs. controls), demonstrating its in vivo
anticancer activity. We next determined the cell proliferation in
CRC tumor tissues using IHS to examine the expression of
proliferating cell nuclear antigen (PCNA), a processivity factor
for DNA polymerase that has been recognized as a histologic marker
for cell proliferation (16). As
shown in Fig. 1B, the percentages
of PCNA-positive cells in the control and the EEPS-treated mouse
group were 38.0 and 22.3%, respectively (P<0.05), suggesting
that EEPS significantly suppressed CRC cell proliferation in
vivo.
EEPS inhibits cell proliferation in HT-29
cells
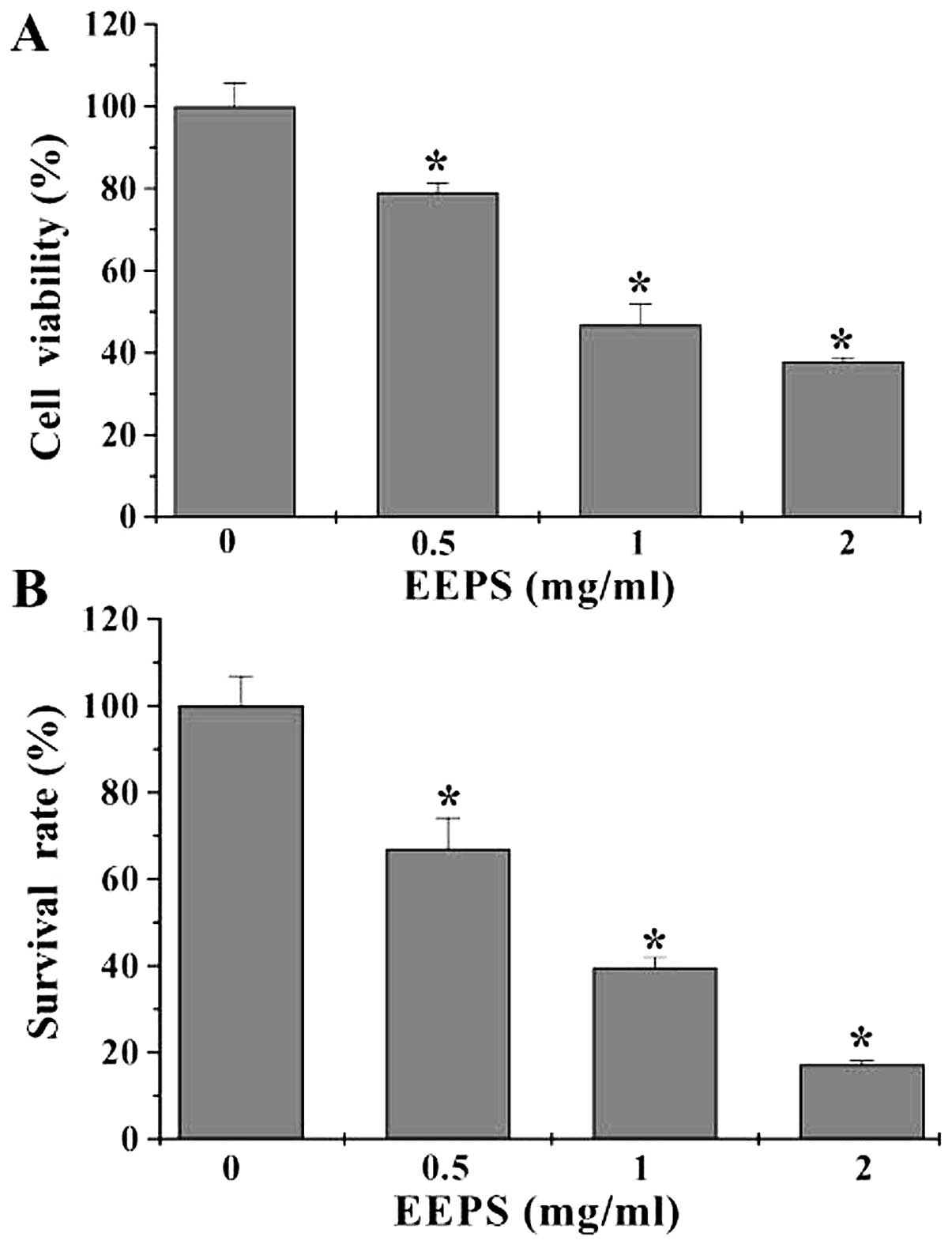
The in vitro effect of EEPS on CRC cell
proliferation was determined by MTT assay to compare the viability
of HT-29 cells in EEPS-treated monolayers to untreated controls. As
shown in Fig. 2A, treatment with
0.5 to 2 mg/ml of EEPS for 24 h dose-dependently reduced cell
viability by 21–62% compared with the untreated control cells
(P<0.05). These results were verified using a colony formation
assay. EEPS treatment reduced the survival rate of HT-29 cells in a
dose-dependent manner (P<0.05, Fig.
2B), suggesting that EEPS can inhibit CRC cell proliferation
in vitro.
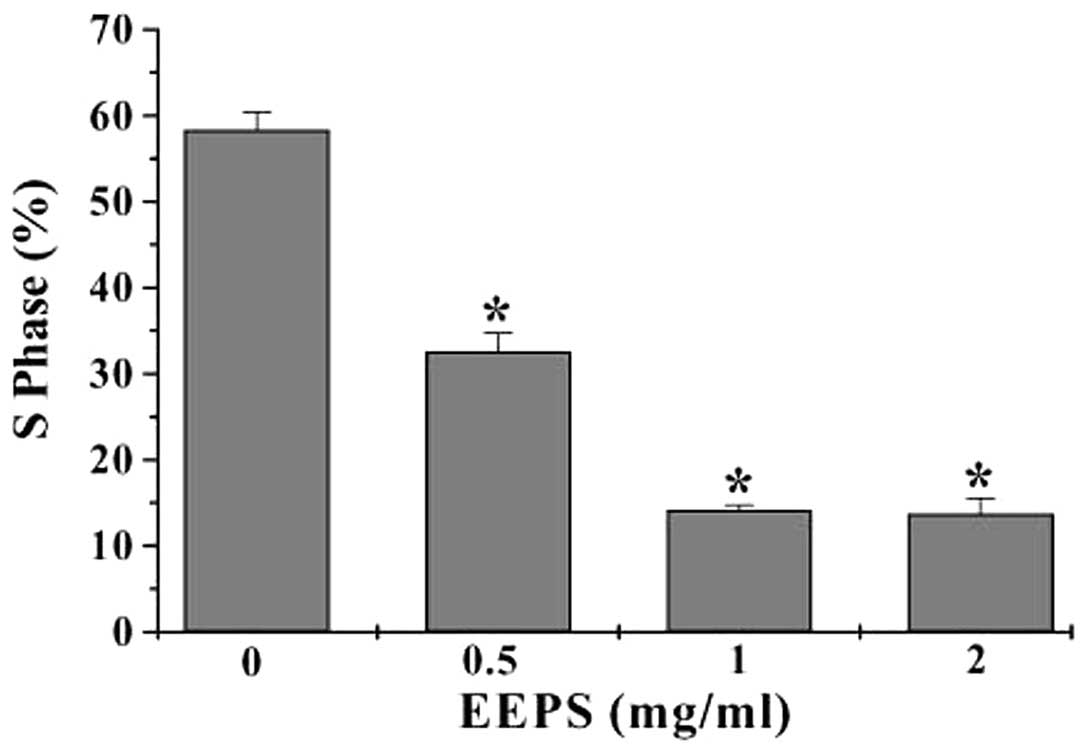
EEPS blocks cell cycle G1/S
progression in HT-29 cells
Eukaryotic cell proliferation is strongly regulated
by the cell cycle, which consists of S, M, G1 and
G2 phases. G1 to S transition is one of the
two main checkpoints of the cell cycle that is responsible for
initiation and completion of DNA replication (17). We, therefore, investigated the
effect of EEPS on the G1 to S phase progression. Using
PI staining followed by FACS analysis we found that the percentage
of HT-29 cells in the S-phase following treatment with 0, 0.5, 1 or
2 mg/ml of EEPS was 58.3, 32.6, 14.1 or 13.7%, respectively
(P<0.05, vs. untreated control cells) (Fig. 3), indicating that the inhibitory
effect of EEPS on CRC cell proliferation was mediated by
G1/S cell cycle arrest.
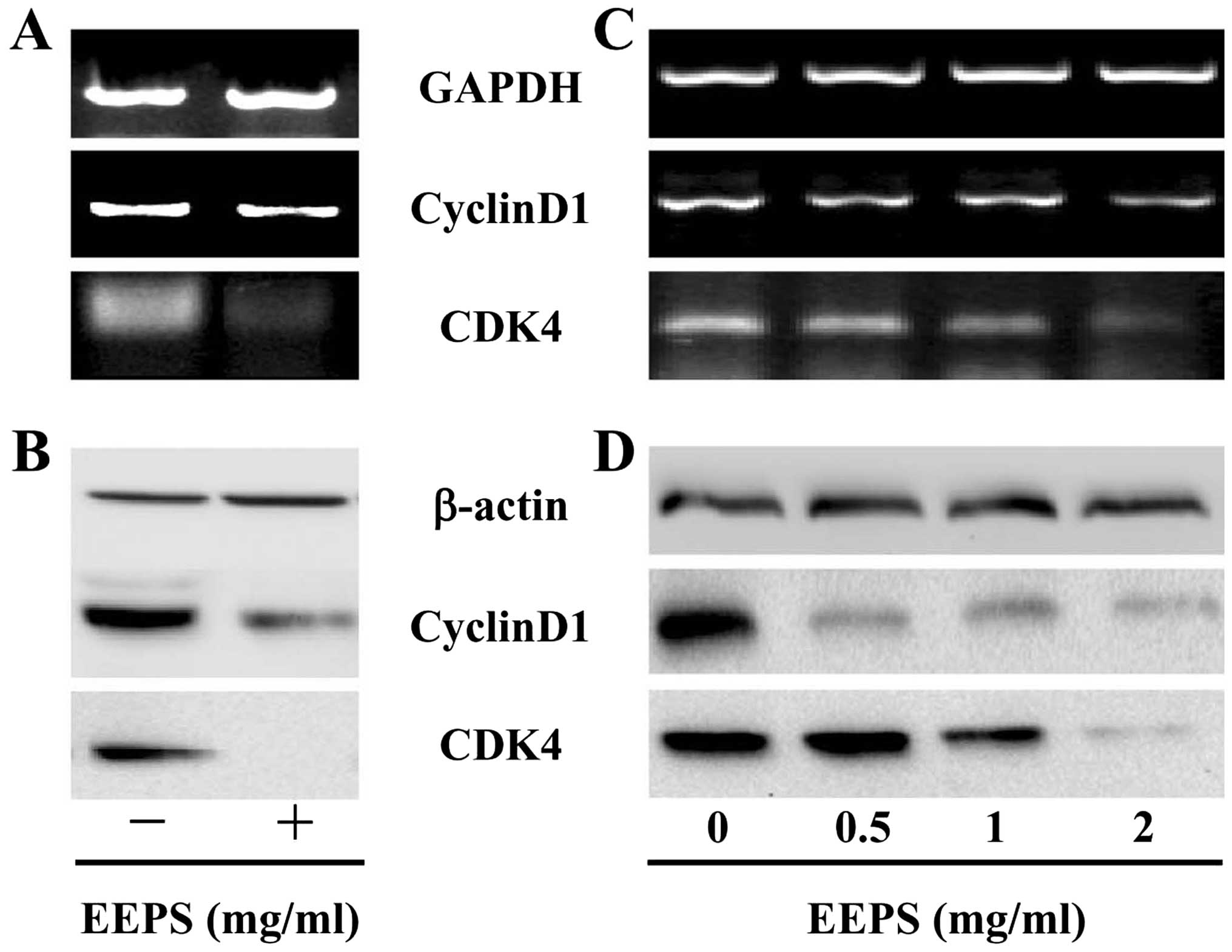
EEPS downregulates the expression of
CyclinD1 and CDK4 in vivo and in vitro
G1/S progression is highly mediated by
CyclinD1, which forms an active complex with its CDK major
catalytic partners (CDK4/6) (18–20).
Overexpression of CyclinD1 and CDK4 is commonly found in many types
of human cancer (21–24). To further explore the mechanism of
the anti-proliferative activity of EEPS, we performed RT-PCR and
western blot analyses to examine the mRNA and protein levels of
CyclinD1 and CDK4. The results of the RT-PCR assay showed that EEPS
treatment significantly reduced the mRNA expression of
pro-proliferative CyclinD1 and CDK4 both in CRC tumor tissues and
in HT-29 cells (Fig. 4A and C); and
the pattern of protein expression of CyclinD1 and CDK4 was similar
to their respective mRNA level (Fig. 4B
and D).
In conclusion, in the present study, we report that
Patrinia scabiosaefolia inhibits colorectal cancer cell
proliferation through blockade of G1/S progression and
the modulation of cell cycle-regulatory gene expression, which may
be one of the mechanisms through which Patrinia
scabiosaefolia exerts its antitumor function.
Acknowledgements
This study was sponsored by the Developmental Fund
of Chen Keji Integrative Medicine (CKJ2014004).
Abbreviations:
|
CRC
|
colorectal cancer
|
|
TCM
|
traditional Chinese medicine
|
|
EEPS
|
ethanol extract of Patrinia
scabiosaefolia
|
|
MTT
|
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
|
|
IHS
|
immunohistochemical staining
|
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gustin DM and Brenner DE: Chemoprevention
of colon cancer: current status and future prospects. Cancer
Metastasis Rev. 21:323–348. 2002. View Article : Google Scholar
|
|
3
|
Longley DB, Allen WL and Johnston PG: Drug
resistance, predictive markers and pharmacogenomics in colorectal
cancer. Biochim Biophys Acta. 1766:184–196. 2006.PubMed/NCBI
|
|
4
|
Sun Y, Zhao H, Guo Y, Lin F, Tang L and
Yao Y: Clinical study of combining chemotherapy of oxaliplatin or
5-fluorouracil/leucovorin with hydroxycamptothecine for advanced
colorectal cancer. Clin Oncol Cancer Res. 6:117–123. 2009.
View Article : Google Scholar
|
|
5
|
Boose G and Stopper H: Genotoxicity of
several clinically used topoisomerase II inhibitors. Toxicol Lett.
116:7–16. 2000. View Article : Google Scholar
|
|
6
|
Newman DJ, Cragg GM and Snader KM: The
influence of natural products upon drug discovery. Nat Prod Rep.
17:215–234. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gordaliza M: Natural products as leads to
anticancer drugs. Clin Transl Oncol. 9:767–776. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cho EJ, Shin JS, Noh YS, Cho YW, Hong SJ,
Park JH, Lee JY, Lee JY and Lee KT: Anti-inflammatory effects of
methanol extract of Patrinia scabiosaefolia in mice with ulcerative
colitis. J Ethnopharmacol. 136:428–435. 2011. View Article : Google Scholar
|
|
9
|
Gao L, Zhang L, Li N, Liu JY, Cai PL and
Yang SL: New triterpenoid saponins from Patrinia scabiosaefolia.
Carbohydr Res. 346:2881–2885. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ju HK, Baek SH, An RB, Bae K, Son KH, Kim
HP, Kang SS, Lee SH, Son JK and Chang HW: Inhibitory effects of
nardostachin on nitric oxide, prostaglandin E2, and tumor necrosis
factor-alpha production in lipopolysaccharide activated
macrophages. Biol Pharm Bull. 26:1375–1378. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chiu LC, Ho TS, Wong EY and Ooi VE: Ethyl
acetate extract of Patrinia scabiosaefolia downregulates
anti-apoptotic Bcl-2/Bcl-XL expression, and induces apoptosis in
human breast carcinoma MCF-7 cells independent of caspase-9
activation. J Ethnopharmacol. 105:263–268. 2006. View Article : Google Scholar
|
|
12
|
Peng J, Chen YQ, Lin JM, Zhuang QC, Xu W,
Hong ZF and Sferra TJ: Patrinia scabiosaefolia extract suppresses
proliferation and promotes apoptosis by inhibiting the STAT3
pathway in human multiple myeloma cells. Mol Med Rep. 4:313–318.
2011.PubMed/NCBI
|
|
13
|
Liu LY, Shen AL, Chen YQ, Wei LH, Lin JM,
Sferra TJ, Hong ZF and Peng J: Patrinia scabiosaefolia induces
mitochondrial-dependent apoptosis in a mouse model of colorectal
cancer. Oncol Rep. 30:897–903. 2013.PubMed/NCBI
|
|
14
|
Chen LW, Liu LY, Ye L, Shen AL, Chen YQ,
Sferra TJ and Peng J: Patrinia scabiosaefolia inhibits colorectal
cancer growth through suppression of tumor angiogenesis. Oncol Rep.
30:1439–1443. 2013.PubMed/NCBI
|
|
15
|
Evan GI and Vousden KH: Proliferation,
cell cycle and apoptosis in cancer. Nature. 411:342–348. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhong W, Peng J, He H, Wu D, Han Z, Bi X
and Dai Q: Ki-67 and PCNA expression in prostate cancer and benign
prostatic hyperplasia. Clin Invest Med. 31:E8–E15. 2008.PubMed/NCBI
|
|
17
|
Nurse P: Ordering S phase and M phase in
the cell cycle. Cell. 79:547–550. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen Y, Robles AI, Martinez LA, Liu F,
Gimenez-Conti IB and Conti CJ: Expression of G1 cyclins,
cyclin-dependent kinases, and cyclin-dependent kinase inhibitors in
androgen-induced prostate proliferation in castrated rats. Cell
Growth Differ. 7:1571–1578. 1996.PubMed/NCBI
|
|
19
|
Grana X and Redy EP: Cell cycle control in
mammalian cells: role of cyclins, cyclin dependent kinases (CDKs),
growth suppressor genes and cyclin-dependent kinase inhibitors
(CKIs). Oncogene. 11:211–219. 1995.PubMed/NCBI
|
|
20
|
Morgan DO: Principles of CDK regulation.
Nature. 374:131–134. 1995. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Harakeh S, Abu-El-Ardat K, Diab-Assaf M,
Niedzwiecki A, El-Sabban M and Rath M: Epigallocatechin-3-gallate
induces apoptosis and cell cycle arrest in HTLV-1-positive and
-negative leukemia cells. Med Oncol. 25:30–39. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kessel D and Luo Y: Cells in
cryptophycin-induced cell-cycle arrest are susceptible to
apoptosis. Cancer Lett. 151:25–29. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Purohit A, Hejaz HAM, Walden L,
MacCarthy-Morrogh L, Packham G, Potter BVL and Reed MJ: The effect
of 2-methoxyoestrone-3-O-sulphamate on the growth of breast cancer
cells and induced mammary tumours. Int J Cancer. 85:584–589. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zafonte BT, Hulit J, Amanatullah DF,
Albanese C, Wang C, Rosen E, Reutens A, Sparano JA, Lisanti MP and
Pestell RG: Cell-cycle dysregulation in breast cancer: breast
cancer therapies targeting the cell cycle. Front Biosci.
5:D938–D961. 2000. View
Article : Google Scholar : PubMed/NCBI
|