Introduction
Apoptosis is an evolutionary conserved cellular
suicide program first proposed in 1972 by Kerr et al
(1). The characteristics of
apoptosis include cell shrinkage, depolarization of the
mitochondria, condensation of chromatin, fragmentation of genomic
DNA, membrane blebbing and the formation of apoptotic bodies, and
these morphological changes make apoptosis obviously different from
necrosis. In general, apoptosis occurs through two pathways: the
death receptor pathway and the mitochondrial-dependent pathway. In
the mitochondrial-initiated pathway, the loss of mitochondrial
integrity is an important event (2). It has been clearly demonstrated that
the mitochondrial membrane potential (ΔΨm) decreases during
apoptosis (3). The intracellular
cysteine protease of the caspase family, which is divided into two
classes including initiators and effectors, is an important
component of apoptosis (4).
Caspase-3, one of the downstream executioner caspases, is widely
recognized as a key member of the caspase family and its activation
is a typical characteristic of apoptosis (5). Tumor-suppressor protein p53 is one of
the major regulators of the apoptotic process in response to DNA
damage from both endogenous and exogenous sources. p53 is involved
in the regulation of B-cell lymphoma 2 (Bcl-2) and Bcl-2-associated
X protein (Bax) (6). Bcl-2, an
anti-apoptotic protein present on the mitochondrial membrane, can
block the release of cytochrome c from mitochondria and
prevent the activation of caspase-9 by binding and inactivating
proapoptotic Bax, preventing activation of the mitochondrial
pathway (7). The expression of
apoptotic regulatory proteins (p53, Bax and Bcl-2) activates the
initiator caspases which in turn directly activate the effector
caspases such as caspase-3 (8). The
activation of the caspase cascade leads to the cleavage of
regulatory and structural proteins which results in the biochemical
and morphological changes associated with apoptotic cell death.
Apoptotic cell death is an essential process in the development and
homeostasis of metazoans (9). The
induction of apoptosis, which promotes cell death automatically
without damage, is also a crucial strategy in anticancer drug
research and development (10).
Therefore, apoptosis has been used as a marker to evaluate the
anticancer activities of phytochemical components during the
development of potential anticancer drugs (8,11).
Medicinal plants for the development of potential
cancer chemopreventive or cancer therapeutic agents are important
due to their minimal adverse side-effects and anti-multidrug
resistance (12,13). Previous studies have indicated that
natural products are rich in anticancer compounds (14). Some plant-derived antitumor drugs
such as paclitaxel, camptothecin, vincristine and vinblastine are
used in cancer chemotherapy (15).
The Commiphora genus in the family of Burseraceae, which
consists of more than 150 species, is mainly distributed in Eastern
Africa, Arabia and India (16,17).
This genus has been extensively researched due to its many
different medicinal uses, and the large number of compounds that
have been isolated from the plants show potential in the treatment
of a wide range of diseases (18).
Based on the theory of traditional Chinese medicine, the resinous
exudates of C. myrrha have been applied for the treatment of
blood stagnation and inflammatory diseases, as well as for the
relief of swelling and pain (19).
Phytochemical investigations have revealed that a series of
terpenoids have been isolated from the resinous exudates of C.
opobalsamum, including cycloartane-type triterpenoids,
sesquiterpenoids, an aliphatic alcohol glycoside and a steroid
(20–22). Some of these isolated compounds have
been shown to possess cytotoxic and anti-proliferative activities
on several cancer cells, particularly the cycloartane-type
triterpenoids (23,24).
Prostate cancer is one of the most commonly
diagnosed malignancies, with high incidence among elderly men
(25–27). In China, the morbidity and mortality
associated with prostate cancer are increasing. Androgen
deprivation therapy is the major treatment for metastatic prostate
cancer. Unfortunately, this form of treatment ultimately leads to
the development of resistance to anti-androgen therapy and the
continuation of tumor metastasis (28,29).
Therefore, it is critically important to develop novel anticancer
agents for the treatment of androgen-independent prostate cancer.
Many natural compounds with therapeutic potential for prostate
cancers are widely found in herbal medicines, including flavonoids
(30), lignans (31,32),
monoterpenoids (33,34), diter-penoids (35,36)
and triterpenoids (21,22,37,38).
Some reported cycloartane-type triterpenoids (21,22),
including cycloartan-24-ene-1α,2α,3β-triol (MY-1) from C.
opobalsamum were found to have a cytotoxic effect on the human
prostate cancer PC-3 cell line. However, the mechanism of apoptosis
that cycloartane-type triterpenoids induce is still unclear.
In the present study, human prostate cancer PC-3
cells were selected to evaluate the cytotoxic effect of MY-1 from
C. myrrha. Based on a previous report (24) that cycloartane triterpenoids induce
the apoptosis of MCF-7 cells through the mitochondrial signaling
pathway, we also investigated whether MY-1 induces apoptosis in
PC-3 cells through the mitochondrial pathway.
Materials and methods
Plant materials
C. myrrha exudates were purchased in December
2009 from the Anguo medicinal material market, Daqing, China. This
material was identified by Professor Taiming Wei of the Institute
of Biological Pharmacy at Harbin Medical University in China. A
voucher specimen (ZY089) was deposited in the Herbarium of the
Institute.
Extraction and isolation
The C. myrrha exudates (0.5 kg) were
pulverized and extracted with a 1-fold volume of 95% ethanol under
reflux (3 × 2 h), and the solvent was evaporated under vacuum to
obtain a residue. The residue was then suspended in H2O
and extracted successively with petroleum ether (PE), chloroform,
and ethyl acetate to yield PE-soluble, chloroform-soluble, and
ethyl acetate-soluble fractions. The chloroform-soluble fraction
was evaporated in vacuo to yield a residue (112.3 g), and
this residue was chromatographed on a silica gel column (63 g) and
eluted with PE-ethyl acetate gradient with increasing amounts of
ethyl acetate to obtain eight subfractions (Fr1–8). Fr7 (16.9 g)
was divided into five fractions (F71–F75) using a silica gel column
and eluted with a gradient of dichloromethane-methanol (100:0,
80:1, 40:1, 20:1 and 10:1). Fraction F73 was further purified to
yield MY-1 (24.1 mg). The structure of MY-1 was identified through
one dimensional nuclear magnetic resonance (1D NMR) spectroscopic
methods.
Cell culture
The human prostate cancer cell line PC-3 was
obtained from the American Type Culture Collection (ATCC;
Rockville, MD, USA) and maintained in Roswell Park Memorial
Institute (RPMI)-1640 medium supplemented with 10% fetal bovine
serum (FBS; Hangzhou Sijiqing Biological Engineering Materials Co.,
Ltd., Zhejiang Deqing County, China) in a humidified incubator with
an atmosphere of 95% air and 5% CO2 at 37°C.
Cytotoxicity assay
For the cytotoxicity assays in the PC-3 cells, the
cells were seeded in 96-well plates at a density of
1×105 cells/well and incubated for 24 h at 37°C in a
humidified 5% CO2 atmosphere prior to testing. The old
media were then replaced with fresh media containing various
concentrations of the tested compounds or the vehicle, dimethyl
sulfoxide (DMSO). After a 24-h incubation, 10 μl of
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT;
Sigma-Aldrich, St. Louis, MO, USA) solution [5 mg/ml in
phosphate-buffered saline (PBS)] was added to each well, and the
plates were then incubated at 37°C for an additional 4 h. After
incubation, the culture medium was replaced with 100 μl of DMSO,
the plates were shaken for 10 min to dissolve the crystals, and the
absorbance of each well was measured using a microplate reader
(Gen5; BioTek, Winooski, VT, USA) at a wavelength of 490 nm.
Briefly, the cell growth curves were generated by plotting the OD
value of each point against the concentration. In each experiment,
the determinations were performed at least three times.
Apoptosis assays
Flow cytometric analysis was employed to determine
the apoptotic cells after treatment with various concentrations of
MY-1 using an Annexin V-FITC/PI apoptosis detection kit (Beyotime
Institute of Biotechnology, Shanghai, China). Briefly, the cells
were seeded in 6-well plates and incubated for 24 h. After
treatment with 9.6 or 14.4 μM MY-1 for 24 h, both adherent and
floating cells were combined, subjected to a brief trypsinization,
washed with PBS, and stained with Annexin V-FITC/propidium iodide
(PI) according to the manufacturer’s instructions. Cells in early
apoptosis (AV+/PI−, lower right quadrant) and
late apoptosis eventually leading to secondary necrosis
(AV+/PI+, upper right quadrant) were
discriminated from the primary necrotic cells (PI+).
TUNEL assay
The terminal deoxynucleotidyl transferase dUTP nick
end-labeling (TUNEL) technique was used to detect the apoptotic
morphological changes in PC-3 cells treated with MY-1 according to
the manufacturer’s instructions (Roche Diagnostics GmbH, Germany).
Briefly, the cells were cultured on coverslides in 6-well plates,
incubated overnight at 37°C, and then treated with various
concentrations of MY-1 for 24 h. After an additional 24-h
incubation, the cells were fixed in freshly prepared 4%
paraformaldehyde-PBS (pH 7.4) for 20 min at room temperature. The
cells were rinsed three times in the wells with 1 ml of cold PBS on
a shaker and blocked with 1 ml of blocking solution (3%
H2O2 in methanol) in each well at room
temperature for 10 min. The cells were rinsed twice with PBS for 10
min. Then, 1 ml of permeabilization solution (0.1% Triton X-100 in
0.1% sodium citrate, freshly prepared) was added to each well of
the monolayer of cells for 2 min at 4°C. The cells were rinsed
twice with PBS for 10 min. Then, 50 μl of the TUNEL reaction
mixture was homogeneously spread across the cell monolayer, and the
cells were incubated for 60 min at 37°C in a humidified atmosphere
in the dark. After the cells were washed three times in PBS, 50 μl
of 4′,6-diamidino-2-phenylindole (DAPI) solution was added for 5
min at room temperature. The samples were analyzed in a drop of PBS
under a fluorescence microscope (Nikon TE2000).
Analysis of mitochondrial membrane
potential (ΔΨm)
The ΔΨm was estimated by JC-1 staining (Beyotime
Institute of Biotechnology). JC-1 is one of the fluorescence probes
that is most commonly used to detect ΔΨm. JC-1 is able to cross the
intact mitochondrial membrane to form aggregates in the
mitochondrial matrix and give a red fluorescence when ΔΨm is high,
whereas the ungathered monomeric JC-1 produces a green fluorescence
at lower ΔΨm. After treatment with various concentrations of MY-1
for 24 h, the cells were harvested, suspended and then incubated
with JC-1 dye according to the manufacturer’s instructions.
Subsequently, the cells were examined on a flow cytometer (BD
FACSAria; Becton-Dickinson, Franklin Lanes, NJ, USA).
Cell cycle analysis
PC-3 cells were seeded in 6-well plates and treated
with various concentrations of MY-1 for 24 h. After treatment, both
floating and adherent cells were collected, washed with cold PBS,
and fixed overnight in 70% cold ethanol at 4°C. After removing the
ethanol, cells were washed twice with cold PBS and stained with 25
μl PI and 10 μl RNase A in the dark for 30 min at 37°C. The DNA
content of cells was analyzed by FACS flow cytometer
(Becton-Dickinson).
Western blot analysis
PC-3 cells were grown for 24 h in the presence or
absence of MY-1 (9.6 or 14.4 μM) at 37°C. The total cell lysates
were prepared after the various treatments. An equal amount of
protein (10 μl) was separated by sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and then
electroblotted onto a nitrocellulose membrane. The membranes were
incubated with TBS-Tween and 5% fat-free dried milk for at least 1
h at room temperature to block any non-specific binding sites and
then incubated with primary antibodies against Bcl-2, Bax,
caspase-3 and p53, all of which were diluted in 5% BSA, overnight
at 4°C. All of the primary antibodies were obtained from Boster
Bio-Engineering Ltd. Co. (Hubei, China), whereas the secondary
antibodies, namely horseradish peroxidase-conjugated anti-rabbit
antibody and horseradish peroxidase-conjugated anti-mouse antibody,
were obtained from Santa Cruz Biotechnology (Berkeley, CA, USA).
β-actin was used as an internal control to verify loading of
similar amounts of the cell lysates.
Statistical analysis
Data are expressed as means ± standard deviation
(SD). The data were statistically analyzed by one-way ANOVA using
the GraphPad software package (GraphPad Software, Inc., USA), and
p<0.05 was considered to indicate a statistically significant
result.
Results
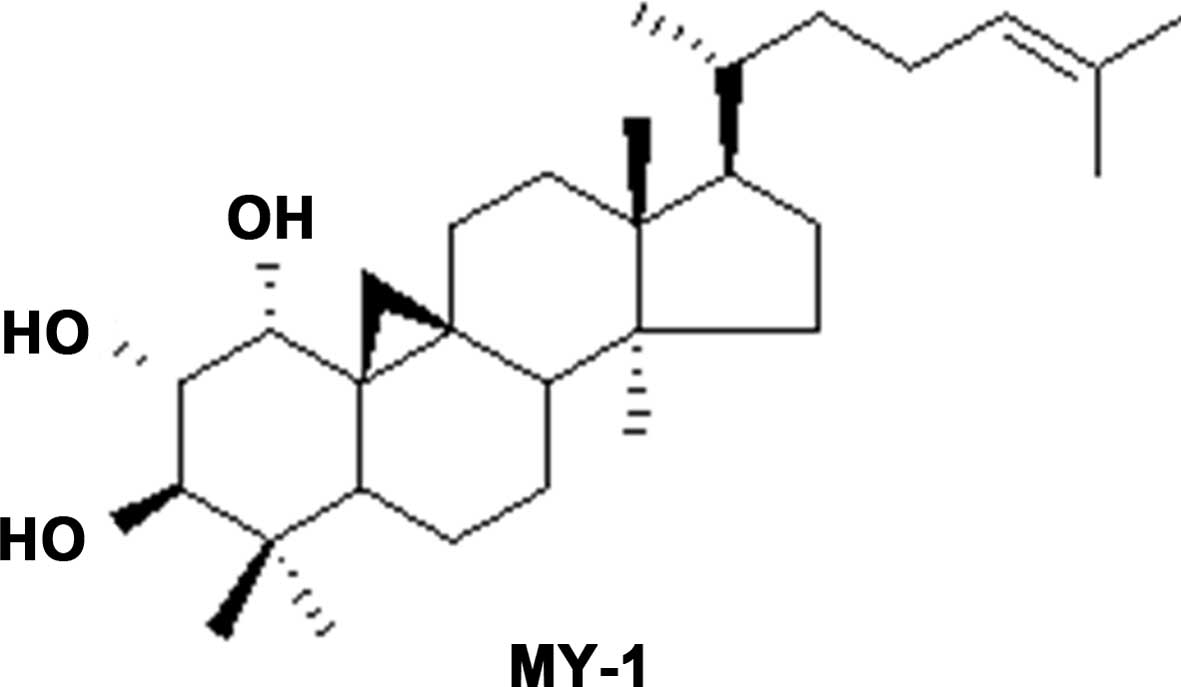
Identification of
cycloartan-24-ene-1α,2α,3β-triol (MY-1)
MY-1 was isolated as white plates, which showed a
molecular ion peak [M]+ at 458.6214 in the
high-resolution electrospray ionization mass spectrometry. A total
of 30 carbons were resolved in the 13C NMR spectrum. The
NMR displayed a pair of doublets at δH 0.52 and 0.73
(J=4.3 Hz), characteristic of the cyclopro-pane ring of a
cycloartane triterpene. By comparison of the NMR data with those of
cycloartan-24-ene-1α,2α,3β-triol (21), MY-1 was deduced to be
cycloartan-24-ene-1α,2α,3β-triol (Fig.
1).
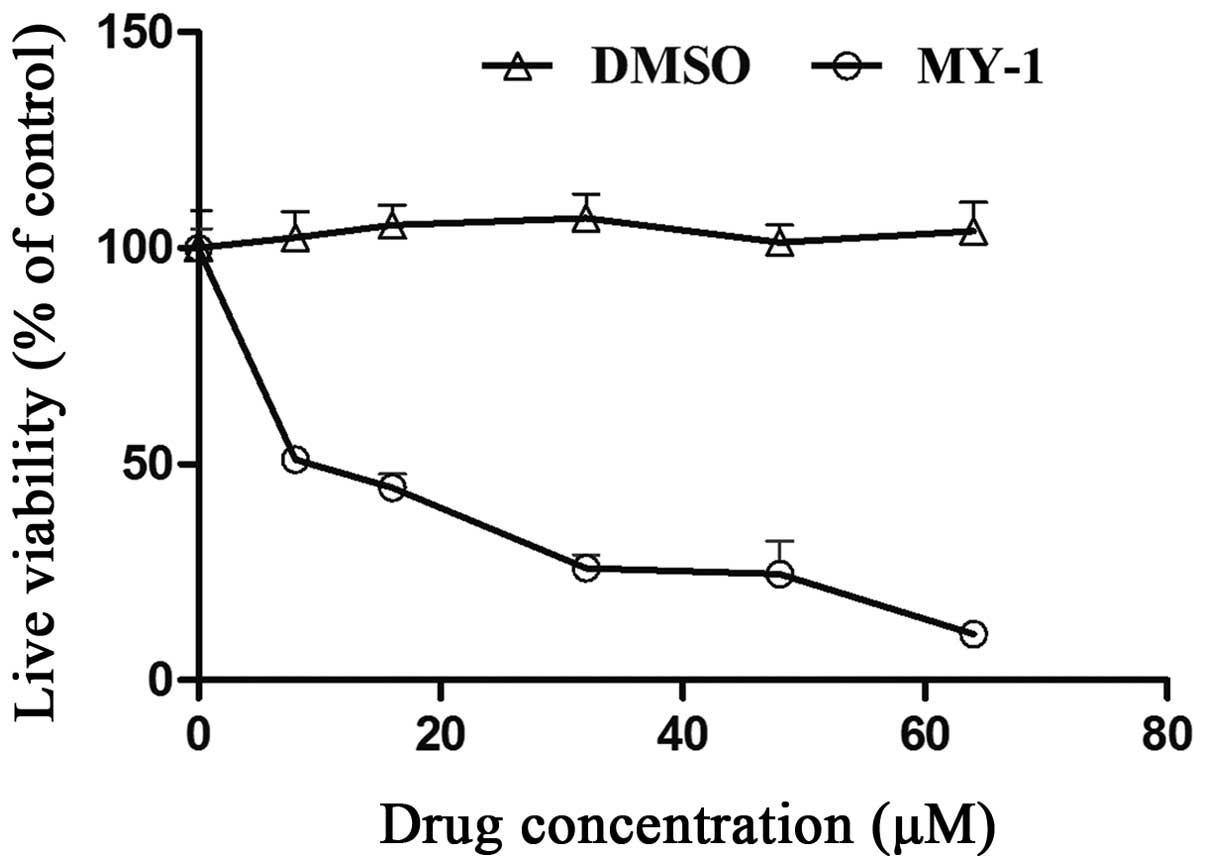
Cytotoxic effect of MY-1 on PC-3
cells
The cytotoxic effect of MY-1 on PC-3 cells was
assayed. As shown in Fig. 2, after
treatment for 24 h, the cell viability of PC-3 cells was decreased
in a concentration-dependent manner. The IC50 value (50%
inhibitory concentration) of MY-1 was 9.6±1.3 μM.
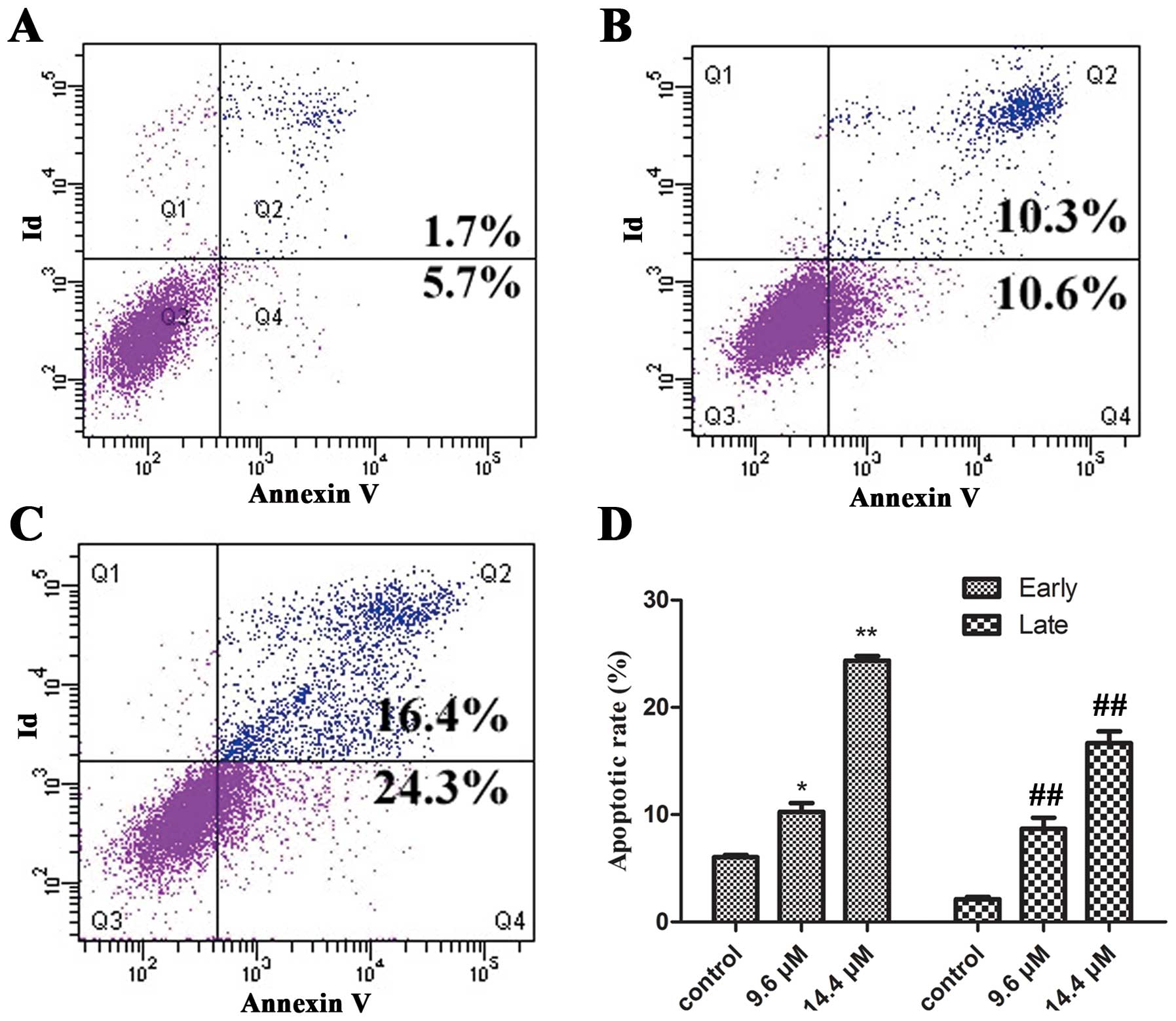
Effect of MY-1 on the apoptosis of PC-3
cells
To confirm and quantify the apoptosis induced by
MY-1 in PC3 cells, the cell groups were either untreated or treated
with MY-1 (9.6 and 14.4 μM) for 24 h. The cells were then stained
with Annexin V-FITC/PI and analyzed by flow cytometry. In the
dual-parameter fluorescent dot plots, the cells in early apoptosis
(Annexin V+/PI−, Q4) and in late
apoptosis (Annexin V+/PI+, Q2)
were counted. As observed in Fig.
3, 5.7±0.4% of the untreated cells were Annexin
V+/PI−, and 1.7±0.35% of the untreated cells
were Annexin V+/PI+. The percentages of
Annexin V+/PI− cells after treatment with 9.6
and 14.4 μM MY-1 were 10.6±1.77 and 24.3±1.43% (p<0.05),
respectively, and the percentages of Annexin
V+/PI+ cells were 10.3±1.91 and 16.4±0.74%
(p<0.01), respectively. These results demonstrated that MY-1
exerts its antitumor activity through the induction of apoptotic
cell death.
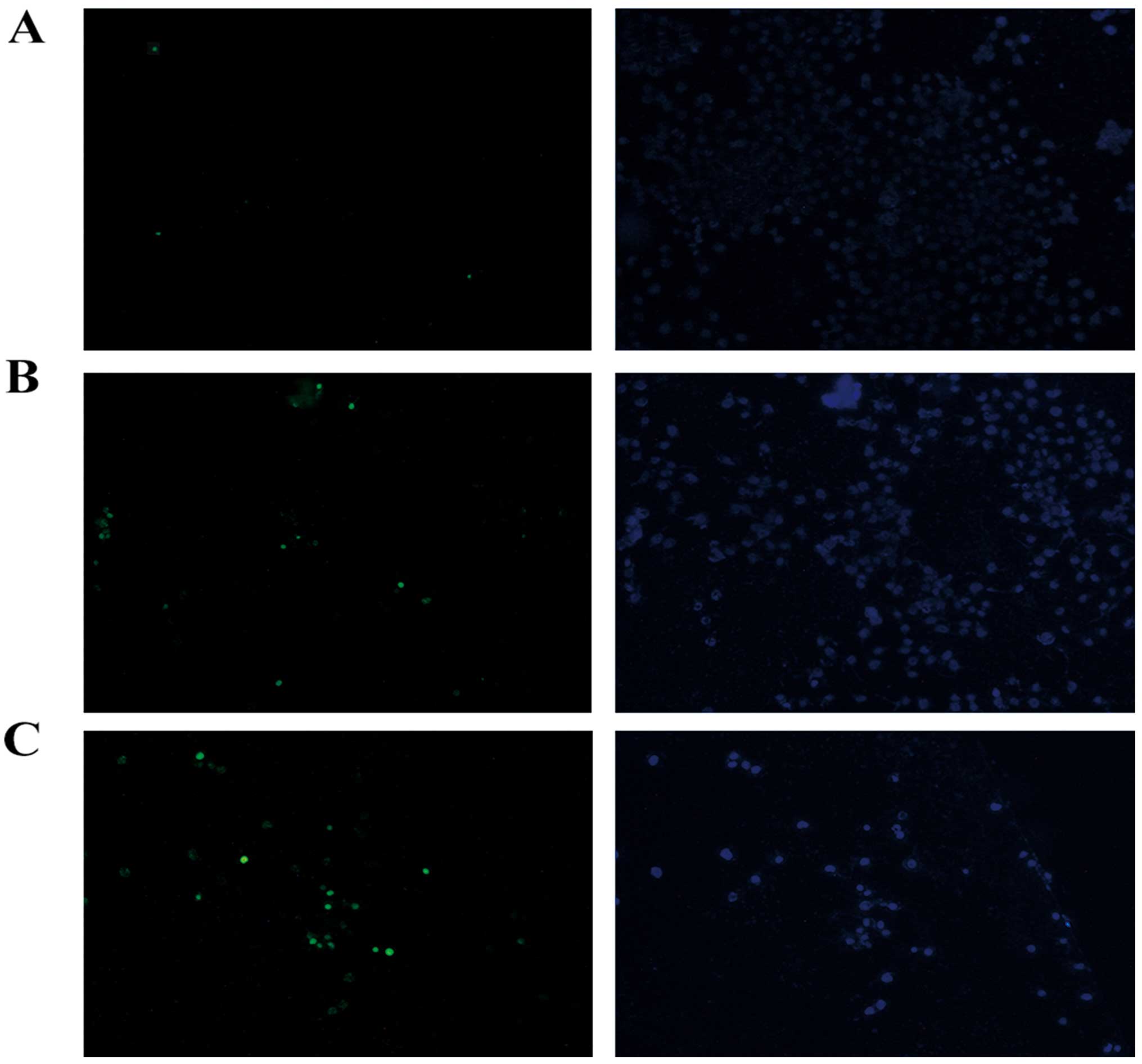
Observation of the cell morphology
To further confirm the results obtained by flow
cytometric analysis, TUNEL assay was used to analyze the apoptotic
morphological changes in the PC-3 cells treated with MY-1. After
staining, the apoptotic dead cells, which showed a green
fluorescence (TUNEL-positive), were distinguished from normal
cells, and these changes were observed to be
concentration-dependent (Fig.
4).
Loss of mitochondrial membrane potential
(ΔΨm)
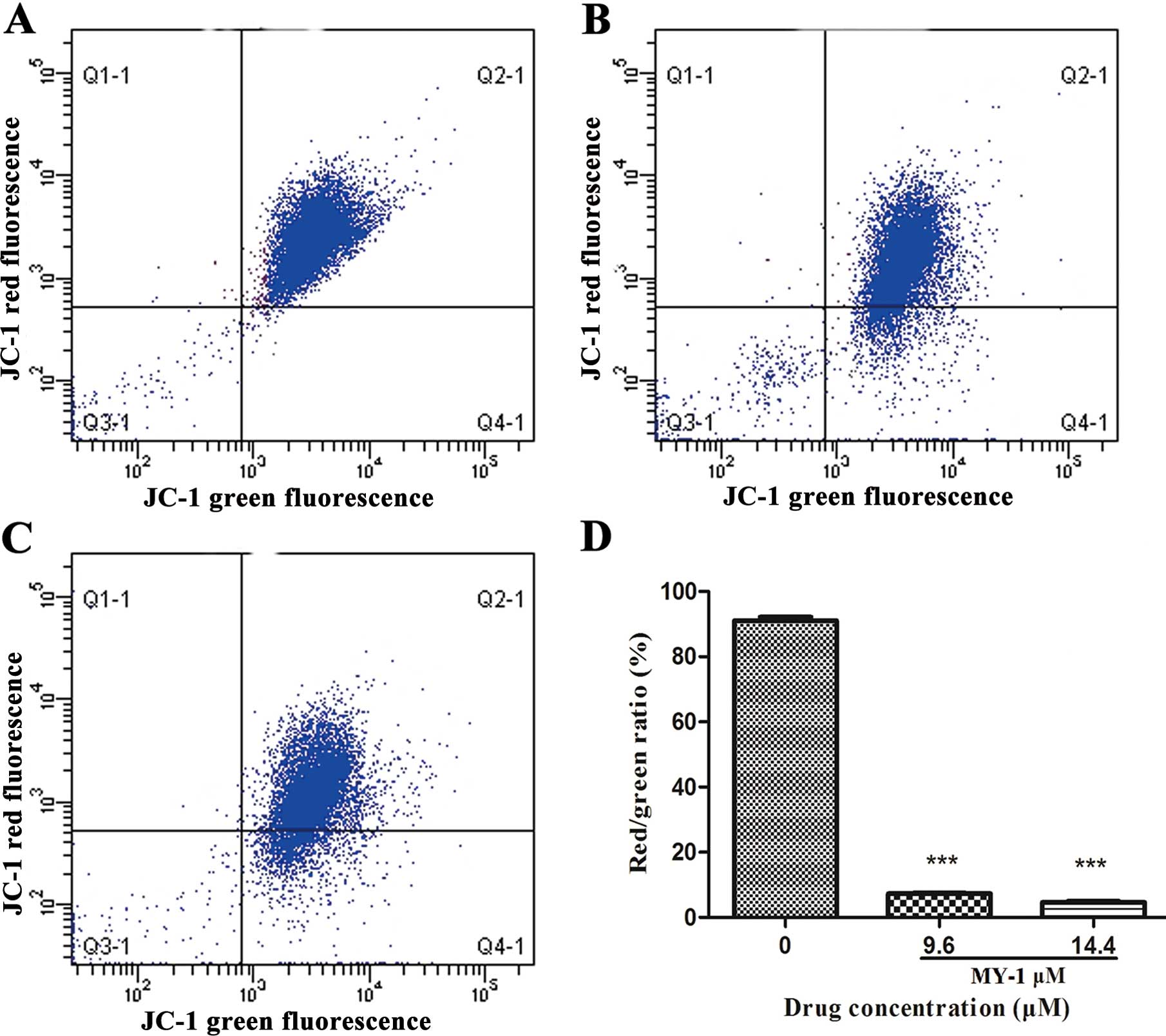
To explore whether the MY-1-induced apoptosis is
triggered by the mitochondrial apoptotic pathway, JC-1 staining was
used to explore the changes in the ΔΨm in MY-1-treated PC-3 cells.
As observed in Fig. 5, after
treatment with 9.6 and 14.4 μM MY-1 for 24 h, the percentages of
cells with green fluorescence were 11 and 19%, respectively,
compared with 1% in the untreated cells. These results further
suggest that the mitochondrial-mediated signaling mechanisms
induced by MY-1 triggered the induction of apoptosis in prostate
cancer PC-3 cells.
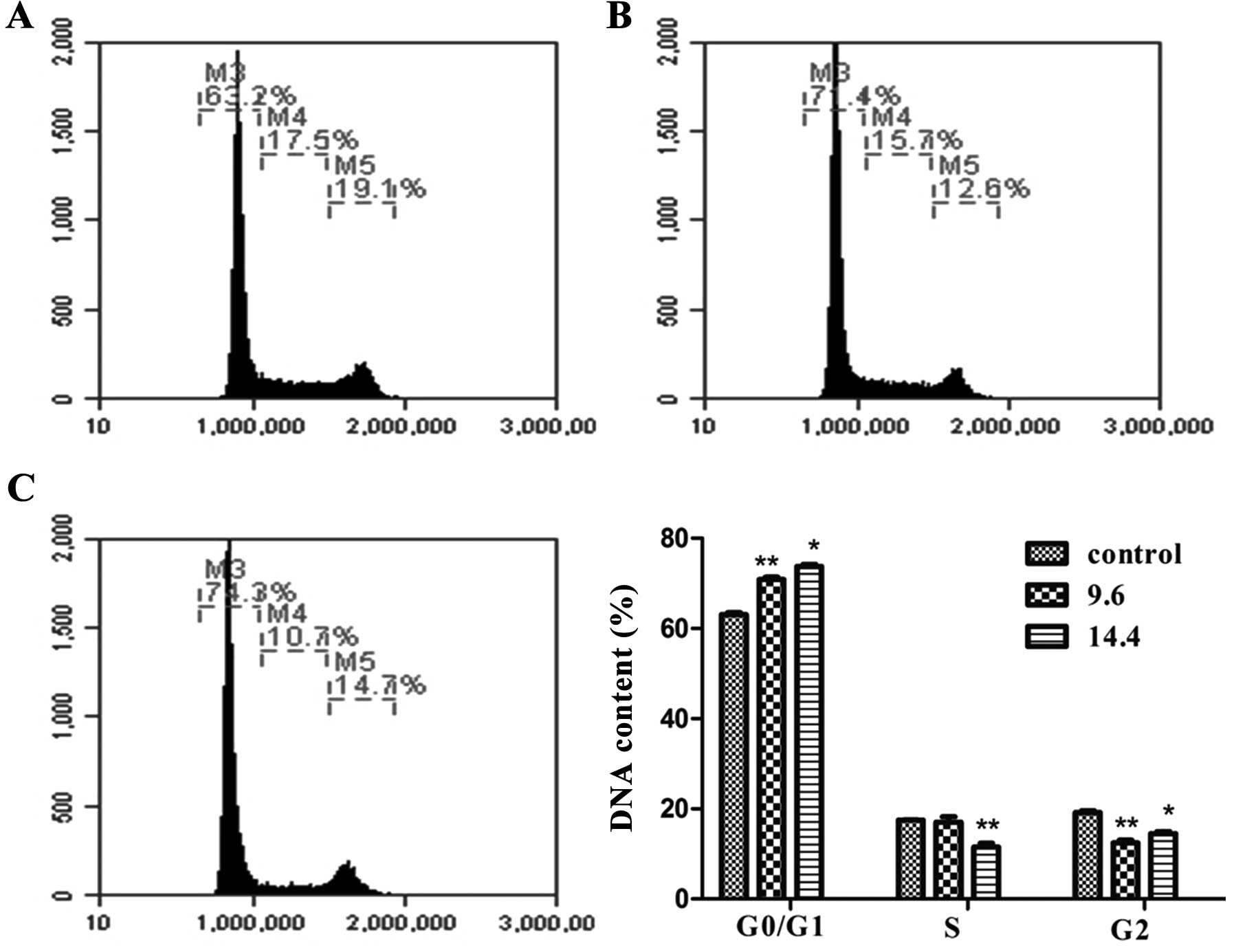
Effect of MY-1 on cell cycle
distribution
To examine the effect of MY-1 on cell cycle
regulation, PC-3 cells were incubated with different concentrations
of MY-1 (9.6 and 14.4 μM) for 24 h. After being stained with PI,
the cells were analyzed for DNA content by flow cytometry. After
exposure to MY-1, the percentage of cells in the G0/G1 phase
increased from 63.2 (control) to 71.4% (9.6 μM), and 74.3% (14.4
μM), respectively. The S phase fraction decreased from 17.5 to
15.7%, and 10.7% with a concomitant decrease in the G2/M phase
(Fig. 6). This result suggests that
MY-1 treatment interfered with cell cycle progression and cell
division in PC-3 cells.
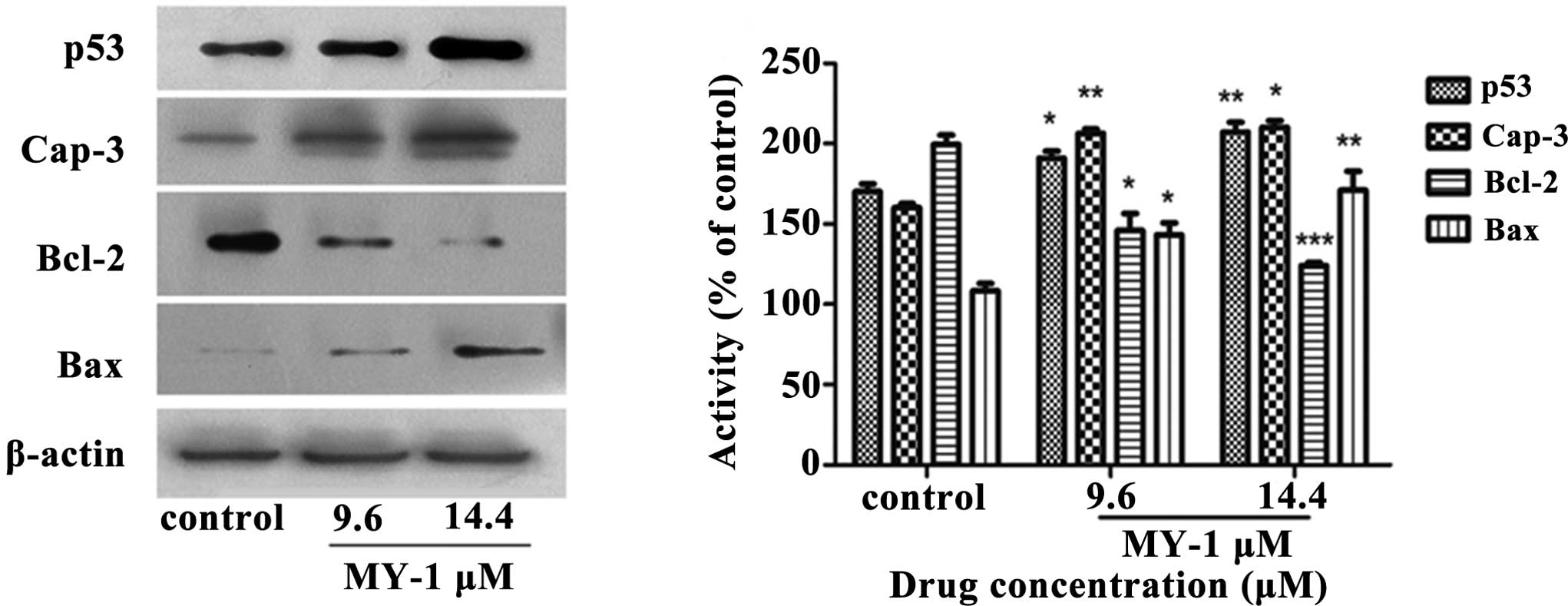
Effect of MY-1 on the expression of p53,
caspase-3, Bcl-2 and Bax proteins
Exposure to cellular and genotoxic stresses can
trigger the tumor suppressor protein p53, which is a
sequence-specific transcription factor, to induce apoptosis via the
regulation of its various downstream proteins. As shown in Fig. 7, the expression of p53 was activated
by MY-1 treatment in a dose-dependent manner.
Bcl-2 and Bax are two proteins that play vital roles
in the apoptotic process (39). The
expression of the proapoptotic protein Bax was significantly
upregulated by MY-1 treatment, whereas the expression of the
anti-apoptotic protein Bcl-2 was downregulated. Based on these
results, MY-1 induced apoptosis through the regulation of
apoptosis-associated proteins.
Discussion
Some studies have reported that tetracyclic
triterpenoids isolated from C. opobalsamum exert
antiproliferative effects on various cancer cells (21,22).
The purpose of the present study was to evaluate the cytotoxic
effect of one tetracyclic triterpenoid (MY-1) from C. myrrha
and investigate whether MY-1 induces apoptosis in human prostate
cancer through the mitochondrial pathway.
In the present study, the chemical structure of MY-1
was found to be similar to other reported cycloartane-type
triterpenoids (21,22) however, a thorough examination of
MY-1 revealed that the locations of the C-24 and C-25 are
different. We assume that the mechanisms of action of
cycloartane-type triterpenoids on human cancer cells are similar,
yet the occurrence of certain structures at C-24 and C-25 may make
the bioactivity of these cycloartane-type triterpenoids different
from each other. Although the pro-apoptotic effects of MY-1 on the
growth of PC-3 cells have been reported (22), the possible mechanism of action
through which it induces apop-tosis in PC-3 cells is still
unknown.
In the present study, the MTT results showed that
MY-1 had a moderate cytotoxic effect against PC-3 cells. The flow
cytometric analyses demonstrated cell cycle arrest at G0/G1 phase
and the characteristic morphological changes further confirmed that
apoptosis was induced by MY-1 (Figs.
3, 4 and 6). It has been demonstrated that the
activation of the anti-oncogene p53 plays an important role in the
suppression of the formation of cancer. Increased expression of the
p53 protein may lead to DNA damage in PC-3 cells after treatment
with MY-1. Further experiments in the present study also showed
that MY-1 treatment notably decreased the expression of the
anti-apoptotic Bcl-2 protein and increased the expression of the
pro-apoptotic Bax protein. These findings confirm the hypothesis
that the MY-1-induced apoptosis in PC-3 cells is triggered by the
downregulation of Bcl-2 and the upregulation of Bax. In addition,
the experimental results also revealed that MY-1 treatment induced
a concentration-dependent activation of caspase-3, which is a key
executioner of apoptosis (Fig. 7).
The observed loss of ΔΨm (Fig. 5)
clarified that the mitochondrial-mediated pathway may be at least
one of the mechanisms responsible for the induction of
apoptosis.
This study has some limitations. It is preliminary
in nature and as such we did not compare the action of MY-1 with
other similar compounds that have also been shown to have
anti-cancer potential. The action of MY-1 on other cell lines would
be of interest, as we can provide no conclusions concerning whether
the apoptotic action on PC-3 cells is unique or could be replicated
in cells that for example express the prostate-specific antigen or
on non-cancerous cell lines.
In conclusion, a tetracyclic triterpenoid (MY-1) was
isolated from C. myrrha. The MY-1 compound has potential
anticancer activity through its induction of apoptosis in PC-3
cells. The mechanism of the apoptosis induction is probably through
the mitochondrial-initiated pathway. As a potential candidate in
the field of anticancer drug discovery against human prostatic
cancer, the in vivo activity of MY-1 in an animal model of
prostate cancer warrants investigation.
Acknowledgements
This study was supported by the Action Plan Project
for the Enterprise of the Ministry of Science and Technology of
China (2009CB522808), the Natural Science Foundation of Jiangsu
Province (BK2008194), the Jiangsu Overseas Research and Training
Program for University Prominent Young and Middle-Aged Teachers and
Presidents, and the Science and Technology Project of the
Department of Traditional Chinese Medicines in Jiangsu Province
(LZ11163).
References
|
1
|
Kerr JF, Wyllie AH and Currie AR:
Apoptosis: a basic biological phenomenon with wide-ranging
implications in tissue kinetics. Br J Cancer. 26:239–257. 1972.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Luo KW, Sun JG, Chan JY, et al: Anticancer
effects of impera-torin isolated from Angelica dahurica: induction
of apoptosis in HepG2 cells through both death-receptor- and
mitochondria-mediated pathways. Chemotherapy. 57:449–459. 2011.
View Article : Google Scholar
|
|
3
|
Gottlieb E, Armour SM, Harris MH and
Thompson CB: Mitochondrial membrane potential regulates matrix
configuration and cytochrome c release during apoptosis. Cell Death
Differ. 10:709–717. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Creagh EM and Martin SJ: Caspases:
cellular demolition experts. Biochem Soc Trans. 29:696–702. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Porter AG and Jänicke RU: Emerging roles
of caspase-3 in apoptosis. Cell Death Differ. 6:99–104. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Amundson SA, Myers TG and Fornace AJ Jr:
Roles for p53 in growth arrest and apoptosis: putting on the brakes
after genotoxic stress. Oncogene. 17:3287–3299. 1998. View Article : Google Scholar
|
|
7
|
Los M, Wesselborg S and Schulze-Osthoff K:
The role of caspases in development, immunity, and apoptotic signal
transduction: lessons from knockout mice. Immunity. 10:629–639.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wijesekara I, Zhang C, Van Ta Q, Vo TS, Li
YX and Kim SK: Physcion from marine-derived fungus Microsporum sp
induces apoptosis in human cervical carcinoma HeLa cells. Microbiol
Res. 169:255–261. 2014. View Article : Google Scholar
|
|
9
|
Wang M, Huang T, Zeng F, et al: Effect of
Smac on TRAIL-induced apoptosis of prostate cancer cell line PC-3
and the molecular mechanism. J Huazhong Univ Sci Technolog Med Sci.
32:233–236. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tian Z, Liu YM, Chen SB, et al:
Cytotoxicity of two triterpenoids from Nigella glandulifera.
Molecules. 11:693–699. 2006. View
Article : Google Scholar
|
|
11
|
da Rocha AB, Lopes RM and Schwartsmann G:
Natural products in anticancer therapy. Curr Opin Pharmacol.
1:364–369. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cheng X, Xiao Y, Wang P, et al: The ethyl
acetate fraction of Polytrichum commune L.ex Hedw induced cell
apoptosis via reactive oxygen species in L1210 cells. J
Ethnopharmacol. 148:926–933. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cragg GM and Newman DJ: Plants as a source
of anti-cancer agents. J Ethnopharmacol. 100:72–79. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gordaliza M: Natural products as leads to
anticancer drugs. Clin Transl Oncol. 9:767–776. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nobili S, Lippi D, Witort E, et al:
Natural compounds for cancer treatment and prevention. Pharmacol
Res. 59:365–378. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Su SL, Duan JA, Tang YP, et al: Isolation
and biological activities of neomyrrhaol and other terpenes from
the resin of Commiphora myrrha. Planta Med. 75:351–355. 2009.
View Article : Google Scholar
|
|
17
|
Abbas FA, Al-Massarany SM, Khan S,
Al-Howiriny TA, Mossa JS and Abourashed EA: Phytochemical and
biological studies on Saudi Commiphora opobalsamum L. Nat Prod Res.
21:383–391. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shen T, Li GH, Wang XN and Lou HX: The
genus Commiphora: a review of its traditional uses, phytochemistry
and pharmacology. J Ethnopharmacol. 142:319–330. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shoemaker M, Hamilton B, Dairkee SH, Cohen
I and Campbell MJ: In vitro anticancer activity of twelve Chinese
medicinal herbs. Phytother Res. 19:649–651. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yuan H, Pan Y and Young CY: Overexpression
of c-Jun induced by quercetin and resverol inhibits the expression
and function of the androgen receptor in human prostate cancer
cells. Cancer Lett. 213:155–163. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shen T, Wan W, Yuan H, et al: Secondary
metabolites from Commiphora opobalsamum and their antiproliferative
effect on human prostate cancer cells. Phytochemistry.
68:1331–1337. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shen T, Yuan HQ, Wan WZ, et al:
Cycloartane-type triterpenoids from the resinous exudates of
Commiphora opobalsamum. J Nat Prod. 71:81–86. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xu R, Fazio GC and Matsuda SP: On the
origins of triterpenoid skeletal diversity. Phytochemistry.
65:261–291. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yang JL and Shi YP: Cycloartane-type
triterpenoids and sesquiterpenoids from the resinous exudates of
Commiphora opobalsamum. Phytochemistry. 76:124–132. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Qiu J, Gao Z and Shima H: Growth of human
prostate cancer cells is significantly suppressed in vitro with
sodium butyrate through apoptosis. Oncol Rep. 27:160–167. 2012.
|
|
26
|
Liu YQ, Hu XY, Lu T, et al: Retigeric acid
B exhibits antitumor activity through suppression of nuclear
factor-κB signaling in prostate cancer cells in vitro and in vivo.
PLoS One. 7:e380002012. View Article : Google Scholar
|
|
27
|
Ghosh J and Myers CE: Inhibition of
arachidonate 5-lipoxy-genase triggers massive apoptosis in human
prostate cancer cells. Proc Natl Acad Sci USA. 95:13182–13187.
1998. View Article : Google Scholar
|
|
28
|
Tan C, Cai LQ, Wu W, et al: NSC606985, a
novel camptothecin analog, induces apoptosis and growth arrest in
prostate tumor cells. Cancer Chemother Pharmacol. 63:303–312. 2009.
View Article : Google Scholar
|
|
29
|
Marcelli M, Marani M, Li X, et al:
Heterogeneous apoptotic responses of prostate cancer cell lines
identify an association between sensitivity to
staurosporine-induced apoptosis, expression of Bcl-2 family
members, and caspase activation. Prostate. 42:260–273. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ahmad KA, Harris NH, Johnson AD, Lindvall
HC, Wang G and Ahmed K: Protein kinase CK2 modulates apoptosis
induced by resveratrol and epigallocatechin-3-gallate in prostate
cancer cells. Mol Cancer Ther. 6:1006–1012. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hahm ER, Arlotti JA, Marynowski SW and
Singh SV: Honokiol, a constituent of Oriental medicinal herb
Magnolia officinalis, inhibits growth of PC-3 xenografts in vivo in
association with apoptosis induction. Clin Cancer Res.
14:1248–1257. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lee DH, Szczepanski MJ and Lee YJ:
Magnolol induces apoptosis via inhibiting the EGFR/PI3K/Akt
signaling pathway in human prostate cancer cells. J Cell Biochem.
106:1113–1122. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Park EJ, Kim SH, Kim BJ, Kim SY, So I and
Jeon JH: Menthol enhances an antiproliferative activity of
1α,25-dihydroxyvitamin D3 in LNCaP Cells. J Clin Biochem
Nutr. 44:125–130. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chung BH, Lee HY, Lee JS and Young CY:
Perillyl alcohol inhibits the expression and function of the
androgen receptor in human prostate cancer cells. Cancer Lett.
236:222–228. 2006. View Article : Google Scholar
|
|
35
|
Obasaju C and Hudes GR: Paclitaxel and
docetaxel in prostate cancer. Hematol Oncol Clin North Am.
15:525–545. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kiviharju TM, Lecane PS, Sellers RG and
Peehl DM: Antiproliferative and proapoptotic activities of
triptolide (PG490), a natural product entering clinical trials, on
primary cultures of human prostatic epithelial cells. Clin Cancer
Res. 8:2666–2674. 2002.PubMed/NCBI
|
|
37
|
Yang H, Landis-Piwowar KR, Lu D, et al:
Pristimerin induces apoptosis by targeting the proteasome in
prostate cancer cells. J Cell Biochem. 103:234–244. 2008.
View Article : Google Scholar
|
|
38
|
Zhang Y, Kong C, Zeng Y, et al: Ursolic
acid induces PC-3 cell apoptosis via activation of JNK and
inhibition of Akt pathways in vitro. Mol Carcinog. 49:374–385.
2010.PubMed/NCBI
|
|
39
|
Sun JG, Chen CY, Luo KW, et al:
3,5-Dimethyl-7H-furo[3,2-g] chromen-7-one as a potential
anticancer drug by inducing p53-dependent apoptosis in human
hepatoma HepG2 cells. Chemotherapy. 57:162–172. 2011. View Article : Google Scholar
|