Introduction
Hepatocellular carcinoma (HCC) is one of the most
common types of cancer and the third cause of cancer-related
mortality worldwide. The survival expectancy is usually no more
than 6 months for HCC patients due to the lack of sensitive
detection method at the early stage of cancer. Although serum
α-fetoprotein (AFP) is the most commonly used biomarker to detect
HCC in clinical screening, its sensitivity and specificity are
limited. There are still ~40% of HCC patients that cannot be
identified using this approach. HCC patients with small tumors or
with well-to-moderately differentiated tumors may not have a high
level of serum AFP. Therefore, it is necessary to identify other
sensitive biomarkers to ensure a more accurate HCC diagnosis. In
the past decade, many studies have demonstrated that
tumor-associated antigens (TAAs) and anti-TAA antibodies may be
useful biomarkers for the diagnosis of certain types of cancer. The
identification of TAAs and anti-TAA antibodies may become a useful
tool for HCC diagnosis, focusing on early facets of the
disease.
Nucleophosmin (NPM1), also known as nucleolar
phosphoprotein B23 or numatrin, is a member of the nucleoplasmin
family, which is a ubiquitously expressed nucleolar protein that
localizes mainly to the nucleoli, yet also shuttles in and out of
the nucleolus, and also between the nucleus and the cytoplasm
(1,2). It is associated with nucleolar
ribonucleoprotein structures, forming complexes with
single-stranded nucleic acids. NPM1 plays multiple roles, including
genomic stability, tumorigenesis (3,4),
ribosome biogenesis (5,6), centriole replication (3,7,8), cell
aging, signal transduction, intracellular protein transport,
protein localization and oligomerization, endodeoxyribonuclease
activity, activation of the NF-κB transcription factor, DNA repair
(9), cell proliferation (10), ribosome assembly (11), response to nucleolar stress
(12) and apoptosis (13). Additionally, NPM1 functions as
regulator of the tumor-suppressor proteins p53 (14–18)
and p14ARF (19), and is also
required for the maintenance of genomic stability. NPM1 protein
expression increases rapidly in the early G1 phase during mitosis
(20). Mutations of the NPM1 gene
resulting in the expression of a cytoplasmic mutant protein,
NPMc+, are the most frequent genetic abnormalities found
in acute myeloid leukemia (21).
NPM1 is frequently overexpressed, mutated, rearranged and deleted
in human cancer cells, therefore it is regarded as a tumor marker
(22).
A previous study demonstrated that NPM1 is expressed
weakly in normal hepatocytes and is highly expressed in liver
cancer cells with a clear correlation between enhanced NPM1
expression and increased tumor grade and consequently poor
prognosis (23). Therefore, there
is the possibility for NPM1 to be a TAA biomarker for early HCC
diagnosis. In the present study, we used an ELISA immunoassay,
western blotting, an indirect immunofluorescence assay and
immunohistochemistry with a tissue array to evaluate and validate
whether the anti-NPM1 autoantibody in patient sera can be used as a
novel biomarker for the detection of HCC.
Materials and methods
Sera and general information
The sera comprised in the present study were
obtained from the serum bank of the Cancer Autoimmunity and
Epidemiology Research Laboratory at the University of Texas at El
Paso (UTEP), and their sources were as follows: 76 from patients
with HCC, 30 from patients with liver cirrhosis (LC), 30 from
patients with chronic hepatitis (CH), and 43 from patients with
systemic lupus erythematosus (SLE) as well as 89 normal human sera
(NHS). In addition, 18 sera from three HCC patients with serial
bleeding samples were also tested in this study. The present study
was approved by the Institutional Review Board of UTEP and
collaborating institutions.
All HCC patients were diagnosed according to the
criteria described in a previous study (24), and had not received treatment with
any chemotherapy or radiotherapy. Patients with CH and LC were
followed up at least 18 months after collecting blood to exclude
individuals with primary biliary cirrhosis and asymptomatic or
clinically undetectable HCC. Normal human sera were collected from
individuals at the same locality during annual health examinations,
who had no obvious evidence of malignancy. Of the 76 HCC patients,
50 (65.8%) were male, and 26 (34.2%) were female. Mean age was
57.0±11.2 years (range, 23–77 years). Fifty-two (68.4%) patients
were positive for hepatitis B virus (HBV), 6 (7.9%) patients for
hepatitis C virus (HCV) and 4 (5.3%) for both HBV and HCV.
Forty-eight (63.2%) had a previous history of CH, 13 (17.1%)
patients had a previous history of LC and 9 (11.8%) patients had no
previous history for either CH or LC. Seventy-one (93.4%) patients
were histologically confirmed. Based on the general diagnostic
rules for liver cancer, 23 (30.3%) patients were in clinical stage
I, 14 (18.4%) in stage II, 24 (31.6%) in stage III, 8 (10.3%) in
stage IV, respectively, and 5 (6.7%) patients had no available data
concerning clinical stage. In the present study, 62 sera were
available for AFP testing. The AFP test kit was provided by GenWay
Biotech (San Diego, CA, USA). The results showed that 61.3% (38/62)
of the sera had abnormal AFP levels (>100 ng/ml) whereas 24
(38.7%) had normal levels (<100 ng/ml).
Recombinant proteins and antibodies used
in the present study
NPM1 construct GFP-NPM1 WT (plasmid ID: 17578) was
purchased from Addgene Inc. (Cambridge, MA, USA), and then
subcloned into the pET28a vector to express the fusion protein with
an N-terminal 6X histidines and T7 epitope tags. The recombinant
protein expressed in Escherichia coli BL21 (DE3) was
purified using nickel column chromatography. Polyclonal anti-NPM1
rabbit antibody and monoclonal anti-β-actin mouse antibody were
obtained from commercial sources (Cell Signaling Technology, Inc.,
Danvers, MA, USA). Horseradish peroxidase (HRP)-conjugated goat
anti-human IgG, HRP-conjugated goat anti-rabbit IgG, HRP-conjugated
goat anti-mouse IgG and FITC-conjugated goat anti-human IgG were
purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA,
USA). Anti-rabbit IgG Fab2 (Alexa Fluor 488) was purchased from
Cell Signaling Technology, Inc.
Enzyme-linked immunosorbent assay
(ELISA)
Standard protocol for ELISA was used as described in
our previous studies (25,26). In brief, a 96-well microtiter plate
(LLC; Immunochemistry Technologies, Bloomington, MN, USA) was
coated overnight at 4°C with recombinant NPM1 protein at a final
concentration of 0.5 μg/ml in phosphate-buffered saline
(PBS). The antigen-coated wells were blocked with gelatin
post-coating solution at room temperature for 2 h. Human sera
diluted at 1:100 with were added to the antigen-coated wells and
incubated for 2 h at room temperature, washed and then incubated
with HRP-conjugated goat anti-human IgG (Caltag Laboratories, San
Francisco, CA, USA) at a 1:4,000 dilution. The substrate
2,2-azino-bis-3-ethylbenzothiazoline-6-sulfonic acid (ABTS;
Sigma-Aldrich, St. Louis, MO, USA) was used to detect the immune
complexes. The average optical density (OD) value at a wavelength
of 405 nm was used for data analysis. The cut-off value designating
a positive reaction was the mean optical density of 90 normal human
sera plus 3 standard deviations (SD).
Western blotting
Denatured recombinant NPM1 protein was
electrophoresed on 10% SDS-PAGE and transferred to a nitrocellulose
membrane. After blocking in PBS with 5% non-fat milk and 0.05%
Tween-20 for 1 h at room temperature, the nitrocellulose membranes
were incubated overnight at 4°C with a 1:200 dilution of human
sera, a 1:500 dilution of polyclonal anti-NPM1 antibody or a 1:500
dilution of monoclonal anti-β-actin mouse antibody, separately.
HRP-conjugated goat anti-human IgG, HRP-conjugated goat anti-rabbit
IgG and HRP-conjugated goat anti-mouse IgG were subsequently
applied as secondary antibody at a 1:10,000 dilution. The ECL kit
was used to detect immunoreactive bands according to the
manufacturer's instructions (Thermo Scientific, Waltham, MA,
USA).
Indirect immunofluorescence assay (IIFA)
and confocal microscopy
An indirect immunofluorescence assay was performed
on Hep2 antinuclear antigen tissue slides (Bion Enterprises, Des
Plaines, IL, USA). The human sera were diluted at 1:80 in PBS, pH
7.4 and incubated with the slides for 30 min at room temperature.
After extensive washing, the slides were incubated with fluorescein
isothiocyanate (FITC)-conjugated goat anti-human IgG secondary
antibody (Santa Cruz Biotechnology, Inc.) or anti-rabbit IgG Fab2
(Alexa Fluor 488) as secondary antibody diluted 1:100 in PBS for 1
h at room temperature. The slides were washed two times with PBS
before adding a drop of mounting media containing 1.5 μg/ml
4',6'-diamidino-2-phenylindole (DAPI) (Vector Laboratories, Inc.,
Burlingame, CA, USA). To prevent photobleaching, the latest steps
were performed in the dark. The slides were then examined under
fluorescence microscopy (LSM 700 confocal microscope; Zeiss) at
x400 magnification, and Zen 2009 software was used for image
capture and analysis.
Absorption of the antibodies with
recombinant protein
The diluted human sera (1:80) were incubated
overnight at 4°C with recombinant NPM1 protein, at a final
concentration of 0.03 μg/μl, and then centrifuged at
10,000 × g for 10 min. The antigen absorbed supernatant was used
for the immunofluorescence assay.
Immunohistochemistry (IHC) with tissue
array slides
Liver cancer tissue array slides with normal tissue
controls (38 cases/80 cores, including pathological diagnosis and
pathological grades, 10 normal liver tissues as controls) were
purchased (US Biomax, Inc., Rockville, MD, USA), and used to detect
the expression of the NPM1 protein. Tissue array slides were
deparaffinized with xylene and dehydrated with ethanol. Antigen
retrieval was performed by microwave-heating methods in Trilogy™
pretreatment solution for 20 min. Avidin/biotin blocking solution
was used to prevent nonspecific binding of the antibodies. The
tissue sections were incubated with polyclonal anti-NPM antibody
(1:50 dilution) overnight at 4°C. HRP detection system (HRP
streptavidin label and polyvalent biotinylated link) and
diaminobenzidine (DAB) substrate kit were used as detecting
reagents. After counterstaining with hematoxylin, the sections were
dehydrated and mounted. The slides were observed by light
microscopy (Leica DM1000, Germany).
Statistical analysis
The mean OD value of each group of patient sera was
compared using the Mann-Whitney U test; the frequency of
autoantibody to TAAs in each group of patient sera and the
expression profile of NPM1 in the liver cancer and normal tissue
groups were compared using the Chi-square (χ2) test with
Fisher's exact test, and two levels of significance (0.05 and 0.01)
were used.
Results
Frequency and titer of autoantibodies
against NPM1 in HCC
The full-length recombinant NPM1 protein was used as
a coating antigen in ELISA to screen autoantibodies against NPM1 in
sera from patients with HCC, LC, CH, SLE and as well as NHS. In
total, 76 sera from patients with HCC, 30 from LC, 30 from CH and
89 sera from normal human individuals were used in the present
study. As shown in Table I, the
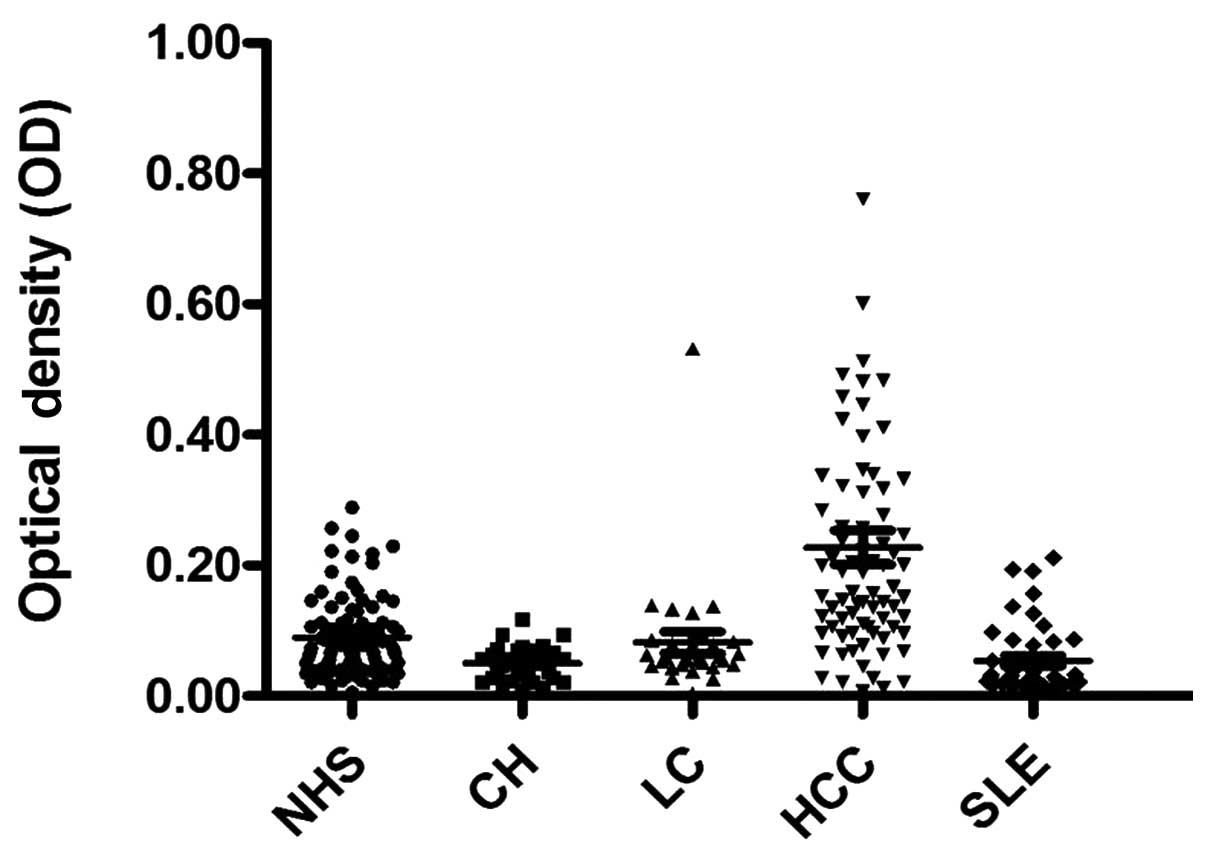
prevalence of the autoantibody against NPM1 was 22.4% (17/76) in
HCC, which was significantly higher than that in LC, CH, SLE and
NHS (P<0.01). The titer of the anti-NPM1 antibodies in human
sera is shown in Fig. 1. The
average titer of the autoantibody against NPM1 in HCC sera was
higher than that in LC, CH, SLE and NHS (P<0.01). The ELISA
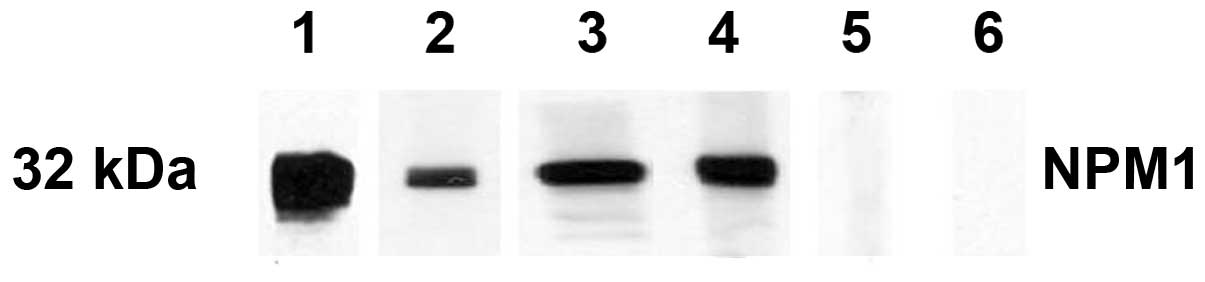
results were also confirmed by western blot analysis. Fig. 2 shows that representative HCC sera
with a positive reaction to NPM1 in ELISA also had strong
reactivity in the western blotting compared to the normal sera. The
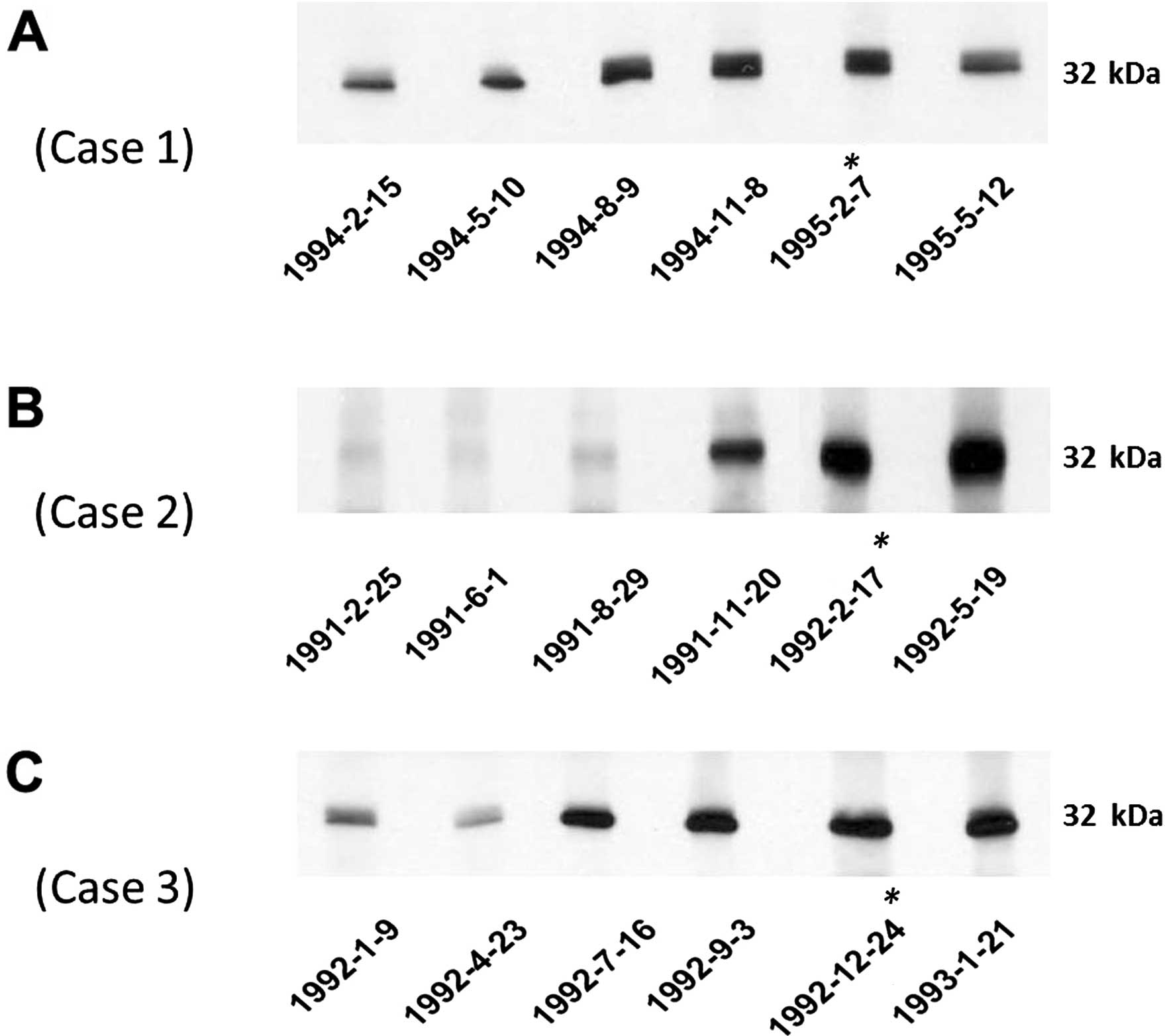
autoantibody to NPM1 in serial serum samples from three HCC
patients (case nos. 1–3) was also tested. The western blotting
results are shown in Fig. 3. In HCC
case 1 and HCC case 3, the anti-NPM1 autoantibody was stronger at 6
months before HCC was detected. In HCC case 2, the anti-NPM1
autoantibody appeared at 3 months before the date of the HCC
diagnosis.
 | Table IFrequency of the autoantibody against
NPM1 in human sera by ELISA. |
Table I
Frequency of the autoantibody against
NPM1 in human sera by ELISA.
| Type of sera | No. tested | Autoantibody to
NPM1 (%) |
|---|
| HCC | 76 | 17 (22.4)a |
| LC | 30 | 1 (3.3) |
| CH | 30 | 0 |
| SLE | 43 | 0 |
| NHS | 89 | 1 (1.1) |
Of the 76 HCC sera, 62 were tested for the presence
of both the anti-NPM1 autoantibody and AFP; 38 sera (61.3%) had an
AFP level >100 ng/ml, and 15 sera (24.2%) were positive for the
anti-NPM1 autoantibody. When both the anti-NPM1 autoantibody and
AFP (>100 ng/ml) were simultaneously used as diagnostic markers,
43 (69.4%, AFP >100 ng/ml) of the 62 HCC sera were positive. Out
of the 17 sera with a normal level of AFP, 5 (29.4%) were positive
for the anti-NPM1 autoantibody.
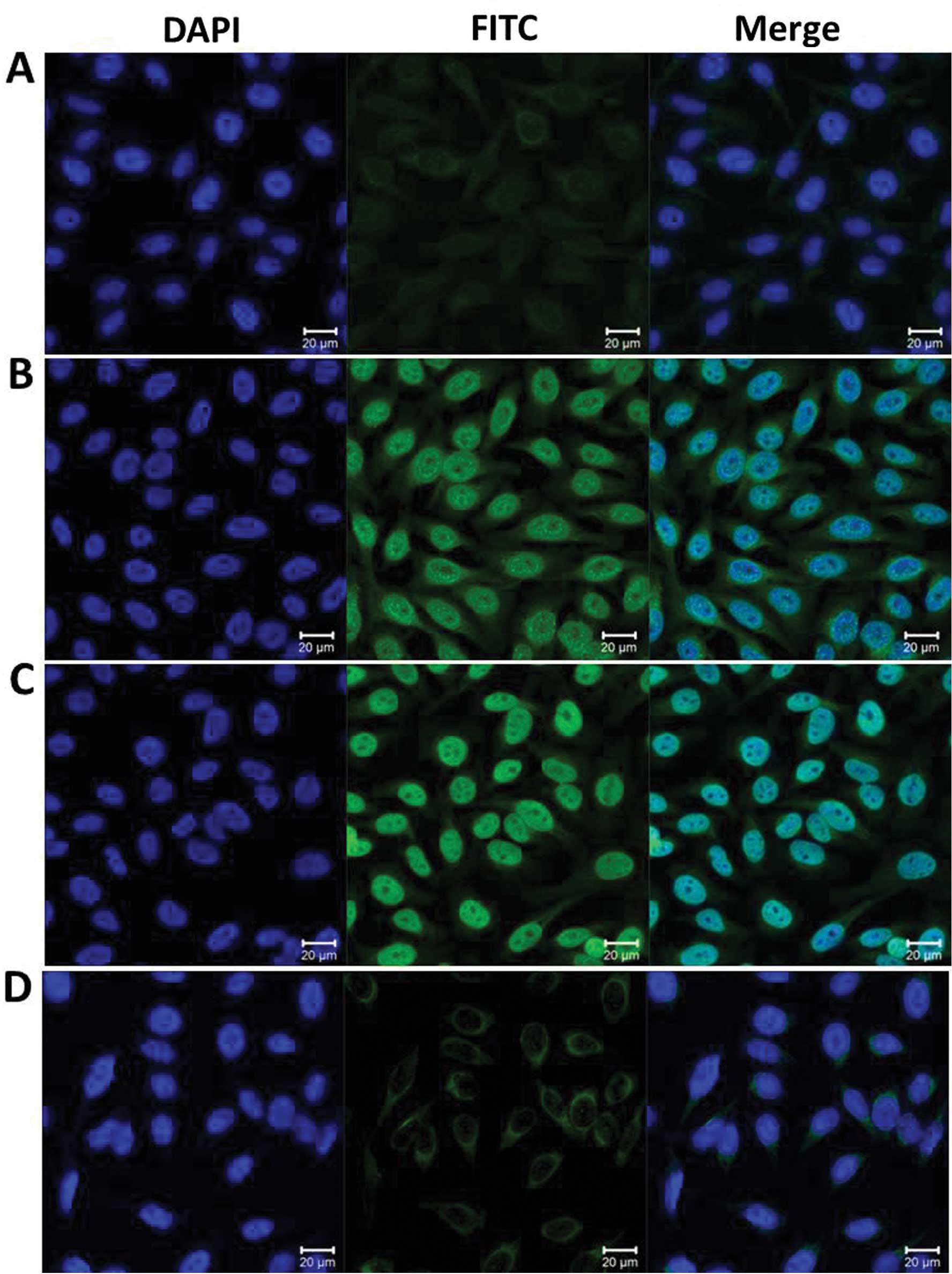
Detection of an intense nuclear staining
pattern in Hep2 cells by indirect immunofluorescence assay with
representative positive HCC sera
To further confirm the reactivity of the auto
antibody in HCC sera to NPM1 and the intracellular location of
NPM1, commercially purchased Hep2 cell slides were used in an
indirect immunofluorescence assay to detect HCC sera with anti-NPM1
positivity in ELISA. As shown in Fig.
4, a representative anti-NPM1-positive HCC serum had an intense
nuclear staining pattern, which was similar in fluorescent staining
pattern and cellular location to that shown by the polyclonal
anti-NPM1 antibody. This fluorescence signal was significantly
reduced when the same HCC serum was pre-absorbed with recombinant
NPM1 protein.
Expression of NPM1 in liver cancer and
normal hepatic tissues by immunohistochemistry
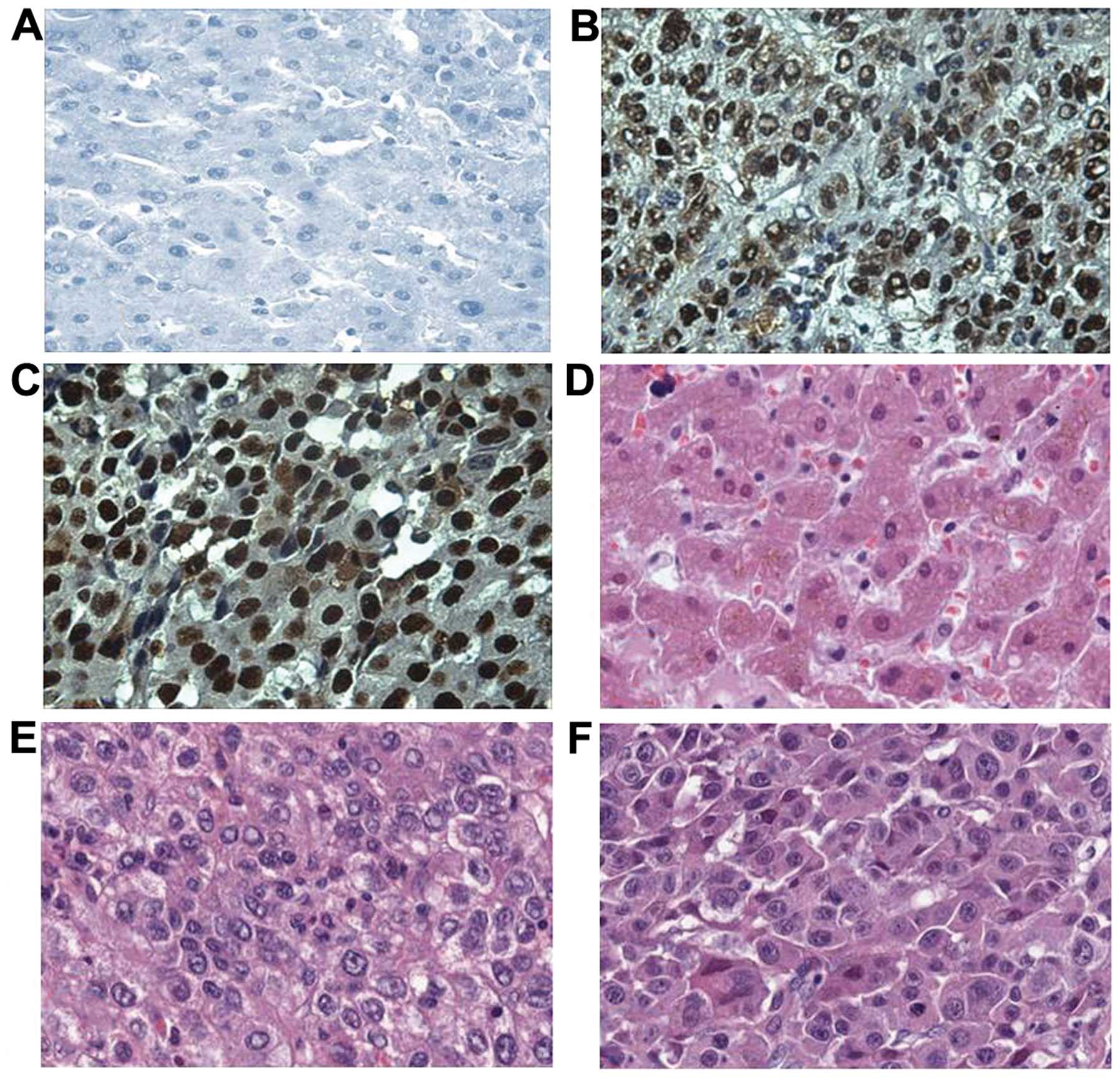
In the present study, the expression profile of NPM1
in liver cancer and normal liver tissues was examined by
immunohistochemistry with tissue array slides. Tissue array slides
were commercially available for the present study; including 30
liver tissues from HCC patients and 10 from normal hepatic tissue
donors. The polyclonal anti-NPM1 antibody was used as the primary
antibody to detect the expression of NPM1 in both liver cancer and
normal tissues. As a result, 22 of the 30 HCC tissues were
positively stained (73.3%). Among these 22 positively stained
tissues, 5 were strongly stained, whereas 17 were moderately
stained. In contrast, one of the 10 normal hepatic tissues
exhibited a positive staining pattern (10%). The characteristics of
patients and NPM1 expression in liver cancer are shown in Table II. The expression of NPM1 in liver
cancer and that in normal tissues are shown in Fig. 5.
| Table IICharacteristics of the patients and
NPM1 expression in liver cancer. |
Table II
Characteristics of the patients and
NPM1 expression in liver cancer.
| Variable | Frequency | % |
|---|
| Age (years) |
| ≥60 | 7 | 23.3 |
| <60 | 23 | 76.7 |
| Gender |
| Male | 20 | 66.7 |
| Female | 10 | 33.3 |
| Grade |
| 2 | 15 | 50.0 |
| 3 | 15 | 50.0 |
| Normal liver
tissue |
| Negative | 9 | 90.0 |
| Positive | 1 | 10.0 |
| Liver cancer |
| Negative | 8 | 26.7 |
| Positive | 22 | 73.3a |
Discussion
Antigenic changes in cancer cells can be recognized
by the immune system of patients themselves as autoantibody
responses to proteins involved in malignant transformation. These
autoantibodies, which have been called ‘reporters’ from the immune
system, can be used as probes in immunoscreening cDNA expression
libraries and immunoproteomics to isolate, identify and
characterize potential TAAs (27,28).
Many studies have demonstrated that serological screening of
autoantibodies to TAAs is useful in the diagnosis of certain types
of cancer in an early stage of their development. In recent years,
a large number of TAAs in cancer have been identified using these
approaches. Some of them have the possibility to be used as
biomarkers in cancer immunodiagnosis (29–31).
It is well demonstrated that cancer has long been recognized as a
multistep process, which involves not only genetic changes
conferring growth advantage but also factors that disrupt the
regulation of growth and differentiation (32,33).
It is possible that some of these factors could be identified and
their functions evaluated with the aid of autoantibodies arising
during tumorigenesis. In our previous study, certain proteins such
as p62, c-Myc, p53, cyclin B1, survivin, p16, RalA, Koc, IMP-1,
Sui1, HCC1, GRP78 and p90, were evaluated and validated as TAAs in
HCC, and autoantibodies against these TAAs have been detected in
sera from patients with HCC (34,35).
In another study, it was suggested that autoantibodies against
these TAAs appear to be supplementary serological markers for the
diagnosis of HCC in AFP-negative cases (36). Our previous studies revealed that
the sensitivity and specificity of autoantibodies to a single TAA
as a diagnostic marker in HCC are currently not high enough for the
diagnosis of HCC (34). Therefore,
it is necessary to continue the study to identify more TAAs for
HCC. Another technique to solve this problem is to combine all the
known HCC-related TAAs to develop a mini-array of multiple TAAs to
detect autoantibodies simultaneously in order to enhance the
sensitivity and specificity (37).
One of our previous studies showed that the final cumulative
prevalence of autoantibodies to 10 TAAs can reach a sensitivity of
66.2% in sera from patients with HCC (34).
NPM1 is a multifunctional protein involved in a
complex network of interactions, yet its actual role in oncogenesis
is controversial (22). The NPM1
gene is mutated or rearranged in a number of hematological
disorders, and it was considered as the most frequently mutated
gene in acute myeloid leukemia (38). Yet, its role in leukemogenesis
remains unclear. A knock-in NPM1 mutation in mice was found to
result in myeloproliferation and denoted a disruption in the
hematopoietic microenvironment (39). Knockdown of NPM1 by RNA interference
inhibited cell proliferation and induced apoptosis in a leukemic
cell line (40). Yet, such changes
have not been detected in solid cancers. However, overexpression of
NPM1 is often found in many types of solid tumors (41–43).
Another study uncovered the critical role of NPM1 in the regulation
of colon cancer cell migration and invasion, and NPM1 may serve as
a potential marker for the prognosis of colon cancer patients
(44). Some results support a
tumor-suppressive role for NPM1 in breast cancer (45). Experiments with NPM1-knockout cells
indicate a tumor-suppressor function for NPM1, both through its
role in the maintenance of genomic stability and in the regulation
of the alternative reading frame (ARF) tumor-suppressing pathway.
Although genetic analysis implicates NPM1 in tumorigenesis, it is
still not clear whether NPM1 may operate either as an oncogene or a
tumor-suppressor gene, perhaps with dual functions (22). Our present study indicated that more
than 20% of HCC sera showed an immune response to NPM1 recombinant
protein. The mean titer of autoantibodies against NPM1 in sera from
patients with HCC was significantly higher than that in LC, CH, SLE
and normal individuals. It is known that LC and CH are common
precursor conditions of HCC. Clinical surveillance for high-risk
individuals, such as CH and LC patients, is important for detecting
early-stage HCC. During transition to malignancy, some HCC patients
develop autoantibodies that are not present during the preceding
chronic liver disease phase. These types of autoantibodies may have
value for the diagnosis of HCC patients at an early stage of
tumorigenesis. Anti-NPM1 autoantibodies are potential biomarkers
for the immunodiagnosis of HCC due to the low positive rate in LC,
CH, SLE and normal individuals and the higher positive rate in HCC
patients. As a control group, sera from patients with SLE which is
an autoimmune disease were also detected for the presence of
anti-NPM1 antibodies. The data suggested that anti-NPM1
autoantibodies are not related to autoimmune diseases such as SLE
and other chronic liver diseases, and these antibodies may have
close relevance to HCC.
The results in the present study also indicated that
when both the autoantibody against NPM1 and AFP are used as
diagnostic markers simultaneously, sensitivity can reach 69.4%,
which is much higher than that when using either anti-NPM1 or AFP
as a marker. It is more important to note that, in HCC patients
with AFP negativity, there were still 29.4% of patients who could
be positively detected with the anti-NPM1 autoantibody. Our data
indicate that the anti-NPM1 autoantibody may be a good supplemental
marker of AFP in HCC diagnosis. In order to test dynamic changes in
anti-NPM1 autoantibodies during malignant transformation from
chronic liver diseases to liver cancer, the sera from three HCC
cases with serial bleeding serum samples were available in our
laboratory, and were tested in the present study. Notably, six
serum samples collected from each patient showed a gradually
increased reactivity to NPM1. Western blot analysis showed that
stronger reactive bands were observed in the serum at 3–6 months
before the clinical diagnosis of HCC. These results indicated that
the titer of the autoantibodies against NPM1 in HCC sera increased
at 3–6 months before the diagnosis of HCC, and the autoantibodies
against NPM1 in HCC sera may be potential biomarkers for
early-stage HCC screening and diagnosis.
Taken together, much evidence suggests that NPM1 is
highly expressed in various tumors and is correlated with the stage
of tumor progression and poor prognosis (23). Overexpression of NPM1 mRNA is
independently associated with the recurrence of bladder carcinoma
and progression to a more advanced stage of disease. A higher NPM1
level was linked to more advanced tumor stages, grades, poor
prognosis and likelihood of recurrence (41,46).
In a study of human breast cancer, NPM1 was identified as an
estrogen-regulated protein associated with acquired estrogen
independence by two-dimensional gel electrophoresis analyses
(47). Yun et al found that
NPM expression was significantly higher in HCC than in
non-malignant hepatocytes, while it was weakly expressed in
hepatocytes from a 5-month-old embryo and in stationary hepatocytes
of healthy adults (48). In the
present study, we also found that NPM1 expression was markedly
increased in liver cancer tissues. Grade III HCC tissues exhibited
a stronger positive signal than HCC tissues with grade II cancer,
when using immunohistochemistry approach with HCC tissue array
slides.
Biological effects such as increased cell growth and
proliferation, and inhibition of differentiation and apoptosis are
characteristics of neoplastic transformation. All of these
biological effects have been demonstrated to be correlated with
NPM1 overexpression in tumor cells. Due to the diversity of
cellular activities exhibited, NPM1 is a key player with dual
functions of either a potential oncogene or a potential tumor
suppressor. Deregulation of NPM1 expression and/or localization
could therefore contribute to tumorigenesis through different
mechanisms (22). The functions of
overexpression of NPM1 in HCC are not clear, yet our data support
the conclusion that NPM1 has a tight relationship with the
occurrence of HCC. Many studies have demonstrated that NPM1 is a
ubiquitously expressed nucleolar protein that localizes mainly to
the nucleoli, but also shuttles in and out of the nucleolus, and
also between the nucleus and the cytoplasm. This complex mechanism
of NPM1 may be the reason why NPM1 is involved in multiple cellular
functions. The shuttling activity of NPM1 and its proper
subcellular localization may be crucial for cellular homeostasis.
In the present study, NPM1 was found in the nucleus of cells. An
IHC study with an HCC tissue array further verified that the
cellular localization of NPM1 is also in the nucleus. In contrast,
in a study on acute myelogenous leukemia, a mutant NPM1 counterpart
(NPMc+) was aberrantly localized in the cytoplasm of
leukemic blasts (49). The
different locations of NPM1 in cancer cells may indicate a
different role of NPM1 in the tumorigenesis pathway. Although much
evidence demonstrates that NPM1 expression correlates with clinical
parameters of cancer patients, the detailed role of NPM1 in cancer
progression is largely unknown.
In summary, the data from the present study provide
further evidence that NMP1 can be used as a potential TAA, and the
anti-NMP1 autoantibody is a useful immunodiagnostic biomarker for
the early detection of HCC. The underlying mechanism of how NPM1
induces humoral immune response in HCC patients, and how it is
involved in the tumorigenesis of HCC still require
investigation.
Acknowledgments
This study was mainly supported by a grant
(SC1CA166016) from the National Institutes of Health (NIH), and was
also partially supported by grants from the National Science and
Technology Key Project of China on ‘Major Infectious Diseases such
as HIV/AIDS, Viral Hepatitis Prevention and Treatment’
(2012ZX10002004-006, 2012ZX10004904-003-001 and
2013ZX10002002-006), The High Technical Personnel Training Item in
Beijing Health System (2011-3-083), Clinical Scientific Research
Cooperation fund of Capital Medical University (15JL67), and the
Beijing Municipal Administration of Hospitals Clinical Medicine
Development of Special Funding Support (XM201308). We also thank
the Border Biological Research Center (BBRC) Core Facilities at The
University of Texas at El Paso (UTEP) for their support, which were
funded by RCMI-NIMHD-NIH grant (2G12MD007592).
References
|
1
|
Borer RA, Lehner CF, Eppenberger HM and
Nigg EA: Major nucleolar proteins shuttle between nucleus and
cytoplasm. Cell. 56:379–390. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wang D, Umekawa H and Olson MO: Expression
and subcellular locations of two forms of nucleolar protein B23 in
rat tissues and cells. Cell Mol Biol Res. 39:33–42. 1993.PubMed/NCBI
|
|
3
|
Grisendi S, Bernardi R, Rossi M, Cheng K,
Khandker L, Manova K and Pandolfi PP: Role of nucleophosmin in
embryonic development and tumorigenesis. Nature. 437:147–153. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wu MH and Yung BY: UV stimulation of
nucleophosmin/B23 expression is an immediate early gene response
induced by damaged DNA. J Biol Chem. 277:48234–48240. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Verheggen C, Almouzni G and
Hernandez-Verdun D: The ribosomal RNA processing machinery is
recruited to the nucleolar domain before RNA polymerase I during
Xenopus laevis sdevelopment. J Cell Biol. 149:293–306. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Huang N, Negi S, Szebeni A and Olson MO:
Protein NPM3 interacts with the multifunctional nucleolar protein
B23/nucleophosmin and inhibits ribosome biogenesis. J Biol Chem.
280:5496–5502. 2005. View Article : Google Scholar
|
|
7
|
Okuda M: The role of nucleophosmin in
centrosome duplication. Oncogene. 21:6170–6174. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Okuda M, Horn HF, Tarapore P, Tokuyama Y,
Smulian AG, Chan PK, Knudsen ES, et al: Nucleophosmin/B23 is a
target of CDK2/cyclin E in centrosome duplication. Cell.
103:127–140. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Okuwaki M, Matsumoto K, Tsujimoto M and
Nagata K: Function of nucleophosmin/B23, a nucleolar acidic
protein, as a histone chaperone. FEBS Lett. 506:272–276. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lindström MS and Zhang Y: Ribosomal
protein S9 is a novel B23/NPM-binding protein required for normal
cell proliferation. J Biol Chem. 283:15568–15576. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Maggi LB Jr, Kuchenruether M, Dadey DY,
Schwope RM, Grisendi S, Townsend RR, Pandolfi PP and Weber JD:
Nucleophosmin serves as a rate-limiting nuclear export chaperone
for the mammalian ribosome. Mol Cell Biol. 28:7050–7065. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Avitabile D, Bailey B, Cottage CT,
Sundararaman B, Joyo A, McGregor M, Gude N, et al: Nucleolar stress
is an early response to myocardial damage involving nucleolar
proteins nucleostemin and nucleophosmin. Proc Natl Acad Sci USA.
108:6145–6150. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Khandelwal N, Simpson J, Taylor G, Rafique
S, Whitehouse A, Hiscox J and Stark LA: Nucleolar NF-κB/RelA
mediates apoptosis by causing cytoplasmic relocalization of
nucleophosmin. Cell Death Differ. 18:1889–1903. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li J, Zhang X, Sejas DP, Bagby GC and Pang
Q: Hypoxia-induced nucleophosmin protects cell death through
inhibition of p53. J Biol Chem. 279:41275–41279. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Colombo E, Alcalay M and Pelicci PG:
Nucleophosmin and its complex network: a possible therapeutic
target in hematological diseases. Oncogene. 30:2595–2609. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Colombo E, Marine JC, Danovi D, Falini B
and Pelicci PG: Nucleophosmin regulates the stability and
transcriptional activity of p53. Nat Cell Biol. 4:529–533. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kurki S, Peltonen K, Latonen L, Kiviharju
TM, Ojala PM, Meek D and Laiho M: Nucleolar protein NPM interacts
with HDM2 and protects tumor suppressor protein p53 from
HDM2-mediated degradation. Cancer Cell. 5:465–475. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Maiguel DA, Jones L, Chakravarty D, Yang C
and Carrier F: Nucleophosmin sets a threshold for p53 response to
UV radiation. Mol Cell Biol. 24:3703–3711. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bertwistle D, Sugimoto M and Sherr CJ:
Physical and functional interactions of the Arf tumor suppressor
protein with nucleophosmin/B23. Mol Cell Biol. 24:985–996. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Falini B, Nicoletti I, Martelli MF and
Mecucci C: Acute myeloid leukemia carrying cytoplasmic/mutated
nucleophosmin (NPMc+ AML): biologic and clinical
features. Blood. 109:874–885. 2007. View Article : Google Scholar
|
|
21
|
Meani N and Alcalay M: Role of
nucleophosmin in acute myeloid leukemia. Expert Rev Anticancer
Ther. 9:1283–1294. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Grisendi S, Mecucci C, Falini B and
Pandolfi PP: Nucleophosmin and cancer. Nat Rev Cancer. 6:493–505.
2006. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu X, Liu D, Qian D, Dai J, An Y, Jiang
S, Stanley B, et al: Nucleophosmin (NPM1/B23) interacts with
activating transcription factor 5 (ATF5) protein and promotes
proteasome- and caspase-dependent ATF5 degradation in
hepatocellular carcinoma cells. J Biol Chem. 287:19599–19609. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Johnson PJ, Leung N, Cheng P, Welby C,
Leung WT, Lau WY, Yu S and Ho S: ‘Hepatoma-specific’
alphafetoprotein may permit preclinical diagnosis of malignant
change in patients with chronic liver disease. Br J Cancer.
75:236–240. 1997. View Article : Google Scholar
|
|
25
|
Zhang JY, Casiano CA, Peng XX, Koziol JA,
Chan EK and Tan EM: Enhancement of antibody detection in cancer
using panel of recombinant tumor-associated antigens. Cancer
Epidemiol Biomarkers Prev. 12:136–143. 2003.PubMed/NCBI
|
|
26
|
Zhang JY, Megliorino R, Peng XX, Tan EM,
Chen Y and Chan EK: Antibody detection using tumor-associated
antigen mini-array in diagnosing human hepatocellular carcinoma. J
Hepatol. 46:107–114. 2007. View Article : Google Scholar
|
|
27
|
Looi KS, Nakayasu ES, Diaz RA, Tan EM,
Almeida IC and Zhang JY: Using proteomic approach to identify
tumor-associated antigens as markers in hepatocellular carcinoma. J
Proteome Res. 7:4004–4012. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang J, Wang K, Zhang J, Liu SS, Dai L
and Zhang JY: Using proteomic approach to identify tumor-associated
proteins as biomarkers in human esophageal squamous cell carcinoma.
J Proteome Res. 10:2863–2872. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ersvaer E, Zhang JY, McCormack E, Olsnes
A, Anensen N, Tan EM, Gjertsen BT and Bruserud O: Cyclin B1 is
commonly expressed in the cytoplasm of primary human acute
myelogenous leukemia cells and serves as a leukemia-associated
antigen associated with autoantibody response in a subset of
patients. Eur J Haematol. 79:210–225. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Looi K, Megliorino R, Shi FD, Peng XX,
Chen Y and Zhang JY: Humoral immune response to p16, a
cyclin-dependent kinase inhibitor in human malignancies. Oncol Rep.
16:1105–1110. 2006.PubMed/NCBI
|
|
31
|
Liu W, Wang P, Li Z, Xu W, Dai L, Wang K
and Zhang J: Evaluation of tumour-associated antigen (TAA)
miniarray in immunodiagnosis of colon cancer. Scand J Immunol.
69:57–63. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Vogelstein B and Kinzler W: The multistep
nature of cancer. Trends Genet. 9:138–141. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: the next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chen Y, Zhou Y, Qiu S, Wang K, Liu S, Peng
XX, Li J, et al: Autoantibodies to tumor-associated antigens
combined with abnormal alpha-fetoprotein enhance immunodiagnosis of
hepatocellular carcinoma. Cancer Lett. 289:32–39. 2010. View Article : Google Scholar :
|
|
35
|
Zhang JY: Mini-array of multiple
tumor-associated antigens to enhance autoantibody detection for
immunodiagnosis of hepatocellular carcinoma. Autoimmun Rev.
6:143–148. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Himoto T, Kuriyama S, Zhang JY, Chan EK,
Nishioka M and Tan EM: Significance of autoantibodies against
insulin-like growth factor II mRNA-binding proteins in patients
with hepatocellular carcinoma. Int J Oncol. 26:311–317.
2005.PubMed/NCBI
|
|
37
|
Zhang JY and Tan EM: Autoantibodies to
tumor-associated antigens as diagnostic biomarkers in
hepatocellular carcinoma and other solid tumors. Expert Rev Mol
Diagn. 10:321–328. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chen W, Rassidakis GZ and Medeiros LJ:
Nucleophosmin gene mutations in acute myeloid leukemia. Arch Pathol
Lab Med. 130:1687–1692. 2006.PubMed/NCBI
|
|
39
|
Chou SH, Ko BS, Chiou JS, Hsu YC, Tsai MH,
Chiu YC, Yu IS, et al: A knock-in Npm1 mutation in mice results in
myeloproliferation and implies a perturbation in hematopoietic
microenvironment. PLoS One. 7:e497692012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Qin FX, Shao HY, Chen XC, Tan S, Zhang HJ,
Miao ZY, Wang L, et al: Knockdown of NPM1 by RNA interference
inhibits cells proliferation and induces apoptosis in leukemic cell
line. Int J Med Sci. 8:287–294. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Tsui KH, Juang HH, Lee TH, Chang PL, Chen
CL and Yung BY: Association of nucleophosmin/B23 with bladder
cancer recurrence based on immunohistochemical assessment in
clinical samples. Acta Pharmacol Sin. 29:364–370. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Montazeri A, Vahdaninia M, Harirchi I, et
al: Breast cancer in Iran: need for greater women awareness of
warning signs and effective screening methods. Asia Pac Fam Med.
7(6)2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Pianta A, Puppin C, Passon N, Franzoni A,
Romanello M, Tell G, Di Loreto C, et al: Nucleophosmin
delocalization in thyroid tumour cells. Endocr Pathol. 22:18–23.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Liu Y, Zhang F, Zhang XF, Qi LS, Yang L,
Guo H and Zhang N: Expression of nucleophosmin/NPM1 correlates with
migration and invasiveness of colon cancer cells. J Biomed Sci.
19(53)2012. View Article : Google Scholar
|
|
45
|
Karhemo PR, Rivinoja A, Lundin J, Hyvönen
M, Chernenko A, Lammi J, Sihto H, et al: An extensive tumor array
analysis supports tumor suppressive role for nucleophosmin in
breast cancer. Am J Pathol. 179:1004–1014. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Tsui KH, Cheng AJ, Chang Pe, Pan TL and
Yung BY: Association of nucleophosmin/B23 mRNA expression with
clinical outcome in patients with bladder carcinoma. Urology.
64:839–844. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Skaar TC, Prasad SC, Sharareh S, Lippman
ME, Brünner N and Clarke R: Two-dimensional gel electrophoresis
analyses identify nucleophosmin as an estrogen regulated protein
associated with acquired estrogen-independence in human breast
cancer cells. J Steroid Biochem Mol Biol. 67:391–402. 1998.
View Article : Google Scholar
|
|
48
|
Yun JP, Miao J, Chen GG, Tian QH, Zhang
CQ, Xiang J, Fu J and Lai PB: Increased expression of
nucleophosmin/B23 in hepatocellular carcinoma and correlation with
clinicopathological parameters. Br J Cancer. 96:477–484. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Falini B, Mecucci C, Tiacci E, Alcalay M,
Rosati R, Pasqualucci L, La Starza R, et al: ; GIMEMA Acute
Leukemia Working Party: Cytoplasmic nucleophosmin in acute
myelogenous leukemia with a normal karyotype. N Engl J Med.
352:254–266. 2005. View Article : Google Scholar : PubMed/NCBI
|