Introduction
Ovarian cancer incidence ranks third among all
gynecological tumors after cervical and uterine cancer, while it is
the most lethal (1). Early symptoms
of ovarian cancer are occult, and it is quite difficult to identify
the tissue types and whether the tumor is benign or malignant.
Moreover, more than 70% of ovarian cancer patients are diagnosed at
an advanced stage limiting treatment options (2). Statistics show that the 5-year
survival rate of FIGO I or II ovarian cancer patients is 70–90%,
when many patients can be cured only by surgery, while the 5-year
survival rate of FIGO III or IV patients is only 20% (3). Ovarian cancer is a malignant tumor
that seriously threaten the health of women, and the incidence of
ovarian cancer is increasing yearly.
MicroRNAs (miRs) are a type of small-molecule
endogenous RNAs, which play an important regulatory role in the
growth process and the development of higher organisms (4). miR-21 is a member of the miRs and is
highly expressed in various human tumors where it participates in
cell proliferation, differentiation and apoptosis, and is closely
related to the growth, invasion and metastasis of tumors (5). Chan et al found that
suppression of miR-21 accelerated the apoptosis and increased the
chemosensitivity of ovarian cancer (6). Liu et al reported that
berberine sensitized ovarian cancer cells to cisplatin through
inhibiting miR-21 expression (7).
PTEN/MMAC1/TEP1 is a tumor-suppressor gene similar
to pRb and p53, and has attracted much attention. It has been
demonstrated that deficiency of the PTEN gene and protein
expression abnormalities appear in a variety of malignant tumor
tissues and tumor cell lines (8).
Wu et al demonstrated that overexpression of PTEN improved
the cisplatin-resistance of human ovarian cancer cells (9). Lau et al reported that
E-cadherin inhibited ovarian cancer cell growth via
β-catenin-Egr1-mediated PTEN expression (10). Lou et al demonstrated that
miR-21 promoted the cell proliferation, invasion and migration
abilities of ovarian epithelial cancer cells by inhibiting PTEN
protein expression (11).
RECK is a newly discovered tumor-suppressor gene.
RECK protein has a unique inhibitory effect on the expression and
activity of MMPs, as a matrix metalloproteinase inhibitor, and RECK
gene expression is closely related to tumor invasion, metastasis
and angiogenesis (12). Studies
have shown that RECK gene expression in liver (13), pancreatic (14), breast (15) and lung cancer (16) is negatively correlated with tumor
invasiveness, and patients with high RECK gene expression have a
significantly more favorable prognosis than those with low
expression. Further research reported that upregulation of RECK
reduced ovarian cancer amplification (17). Histone deacetylase inhibitor
inhibited human ovarian cancer cell migration through upregulation
of RECK and downregulation of matrix metalloproteinase-2 (MMP-2) in
HDAC4-blocked SKOV-3 cells (18).
The relationship between apoptosis and tumors is one
of the ‘hotspots’ in recent years. Uncontrolled apoptosis is an
important feature leading to tumor occurrence and development. The
Bcl-2 family has attracted increased attention, and Bcl-2 and Bax
are important members of this family. They are mainly involved in
the regulation of apoptosis through the mitochondrial pathway
(19). Xie et al found that
estrogen and progesterone significantly inhibited the cell survival
and promoted the cell apoptosis of ovarian cancer cells through
inhibition of the expression of Bcl-2 (20). Ma et al showed that a
specific cell-penetrating peptide induced apoptosis by
down-regulation of Bcl-2 in human ovarian cancer SKOV3 cells
(21).
Icariin, a plant flavonoid glycoside, is an
effective pharmacological ingredient found in the traditional
Chinese medicinal plant Epimedium (22). Due to its estrogen-like structure,
icariin has a wide range of pharmacological effects on the heart
and brain vascular system, bone metabolism, immune and nervous
system, sexual function, and exhibits anti-inflammatory and
antitumor effects (23). Wang et
al found that icariin decreased the viability and induced the
apoptosis of MLTC-1 cells through regulation of the expression of
Bcl-2/Bax (24). It is vital to
explore new prospective effective anticancer drugs and develop
therapies against ovarian cancer. The present study aimed to
investigate the anticancer effect of icariin against ovarian cancer
cells. We assessed whether icariin regulates the proliferation and
apoptosis of human ovarian cancer cells and further explored the
mechanism underlying its activity.
Materials and methods
Chemicals and reagents
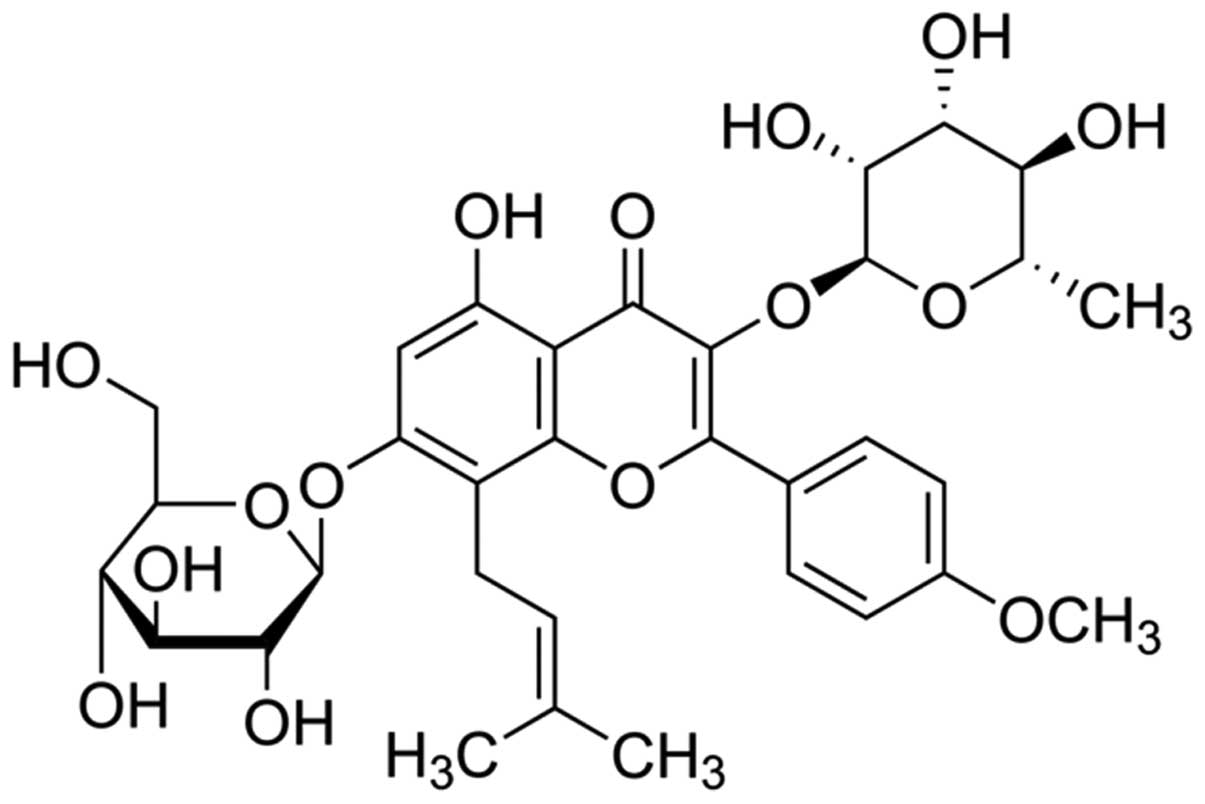
The chemical structure of icariin (with a purity
≥94%; Sigma) is shown in Fig. 1.
Roswell Park Memorial Institute (RPMI)-1640 and fetal calf serum
(FBS) were purchased from Gibco HyClone (Invitrogen Co., South
America) and Invitrogen Co., BRL, respectively.
3,3-(4,5-Dimethylthiazol-2yl)-2,5-diphenyltetrazolium bromide (MTT)
was purchased from Sangon Biotech (Shanghai, China). Annexin
V-FITC/propidium iodide (PI) apoptosis detection kit was obtained
from Takara Bio, Inc. (Dalian, China). Caspase-3 colorimetric assay
kits were obtained from Beyotime (Nanjing, China). TRIzol reagent,
cDNA synthesis kit and SYBR-Green kit were obtained from Tiangen
(Beijing, China). Lipofectamine 2000 and Opti-MEM I were obtained
from Invitrogen Life Technologies. The BSA kit was obtained from
Thermo Scientific, USA.
Cell lines and culture
The human ovarian cancer A2780 cell line was
acquired from the Animal Laboratory of Shengjing Hospital
Affiliated to China Medical University. The A2780 cells were
cultured in RPMI-1640 containing 10% FBS with 100 U/ml penicillin
and 100 U/ml streptomycin and were maintained at 37°C in a
humidified atmosphere of 5% CO2. A2780 cells were
passaged every 2–3 days.
Cell viability assay
A2780 cells (5×103/well) were seeded into
96-well culture plates and treated with icariin (3, 6, 13, 25, 50
and 100 μM), or vehicle for 24, 48 or 72 h. MTT (10
μl) was added to the A2780 cells and incubation was carried
out for 4 h at room temperature. The supernatant was discarded, and
dimethyl sulfoxide (DMSO) was added to each well and was shaken for
20 min. Then, A2780 cell viability was determined using the MTT
assay.
Determination of cell apoptosis by flow
cytometry
A2780 cells (2×106/well) were seeded into
6-well culture plates and treated with icariin (13, 25 and 50
μM), or vehicle for 24, 48 or 72 h. A2780 cells were
collected and washed twice with ice-cold phosphate-buffered saline
(PBS). A2780 cells (1×106/ml) were resuspended with
Annexin V binding buffer. Annexin V-FITC (5 μl) was added to
the A2780 cells and stained for 30 min at darkness. Then, 10
μl PI was added to the A2780 cells and stained for 10 min
according to the manufacture’s instructions (Takara Bio, Inc.).
Cell apoptosis of A2780 cells was immediately detected using flow
cytometry (EPICS® Altra™; Olympus).
Detection of caspase-3 activity
A2780 cells (2×106/well) were seeded into
6-well culture plates and treated with icariin (13, 25 and 50
μM), or vehicle for 24, 48 or 72 h. The caspase-3 activity
in fluorescence was detected at the wavelength of 405 nm with the
caspase-3 colorimetric assay kits.
qPCR analysis of miR-21 expression
A2780 cells (2×106/well) were seeded into
6-well culture plates and treated with icariin (13, 25 and 50
μM), or vehicle for 24, 48 or 72 h. According to the
manufacturer’s instructions (Tiangen), total RNA was isolated from
the A2780 cell samples using TRIzol reagent. cDNAs were synthesized
and detected using cDNA synthesis kit, according to the
manufacturer’s instructions (Tiangen). The expression of miR-130b
was detected using the SYBR-Green kit.
Immunoblot analysis
A2780 cells (2×106/well) were seeded into
6-well culture plates and treated with icariin (13, 25 and 50
μM), or vehicle for 24, 48 or 72 h. A2780 cells were
harvested and washed with ice-cold PBS. The harvested cells were
suspended in lysis buffer (10 mM Tris, 150 mM NaCl, 1% Triton
X-100, 1% Na-deoxycholate, 5 mM EDTA) and placed on ice for 30 min.
A2780 cells were harvested and centrifuged at 12,000 × g for 10 min
at 4°C. The total proteins of the cell lysates were determined
using the BSA kit (Thermo Scientific). Equal protein was separated
by SDS-PAGE, and then transferred onto PVDF membranes (0.22 mm;
Millipore, Billerica, MA, USA). The membranes were blocked and
incubated overnight with anti-PTEN (1:1,000), anti-PTEN (1:1,000),
anti-Bcl-2 (1:1,500) and anti-β-actin (1:3,000) (all from Santa
Cruz Biotechnology, Inc., Santa Cruz, CA, USA) overnight at 4°C.
The membranes were developed using ECL reagent (1:1,000; Santa Cruz
Biotechnology, Inc.) and imaged using a polaroid imaging system
(Amersham).
Transfection of miR-21 and
anti-miR-21
miR-21 inhibitors and miR-21 mimics were chemically
synthesized by BeastBio Co., Ltd. (Shanghai, China). A2780 cells
(2×106/well) were seeded into 6-well culture plates.
miR-21 mimics (100 pmol) and miR-21 inhibitors were transfected
into the A2780 cells with Lipofectamine 2000 and Opti-MEM I reduced
serum medium (Invitrogen Life Technologies). The expression of
miR-21 was detected using the SYBR-Green kit. The primers for
miR-21 were: 5′-GCCCGCTAGCTTATCAGACTGATG-3′ and
5′-GCCCGCTAGCTTATCAGACTGATG-3′, respectively. The primers for U6
were: 5′-GCGCGTCGTGAAGCG TTC-3′ and 5′-GTGCAGGGTCCGAGGT-3′,
respectively.
Statistical analysis
Each experiment was repeated at least three times.
Data analysis was performed with SPSS 17.0 software (SPSS, Inc.,
Chicago, IL, USA), and data are shown as the mean ± SD. P<0.05
was considered to indicate a statistically significant
difference.
Results
Icariin suppresses the proliferation of
ovarian cancer cells
To explore the potential anticancer effect of
icariin on the proliferation of ovarian cancer A2780 cells, MTT
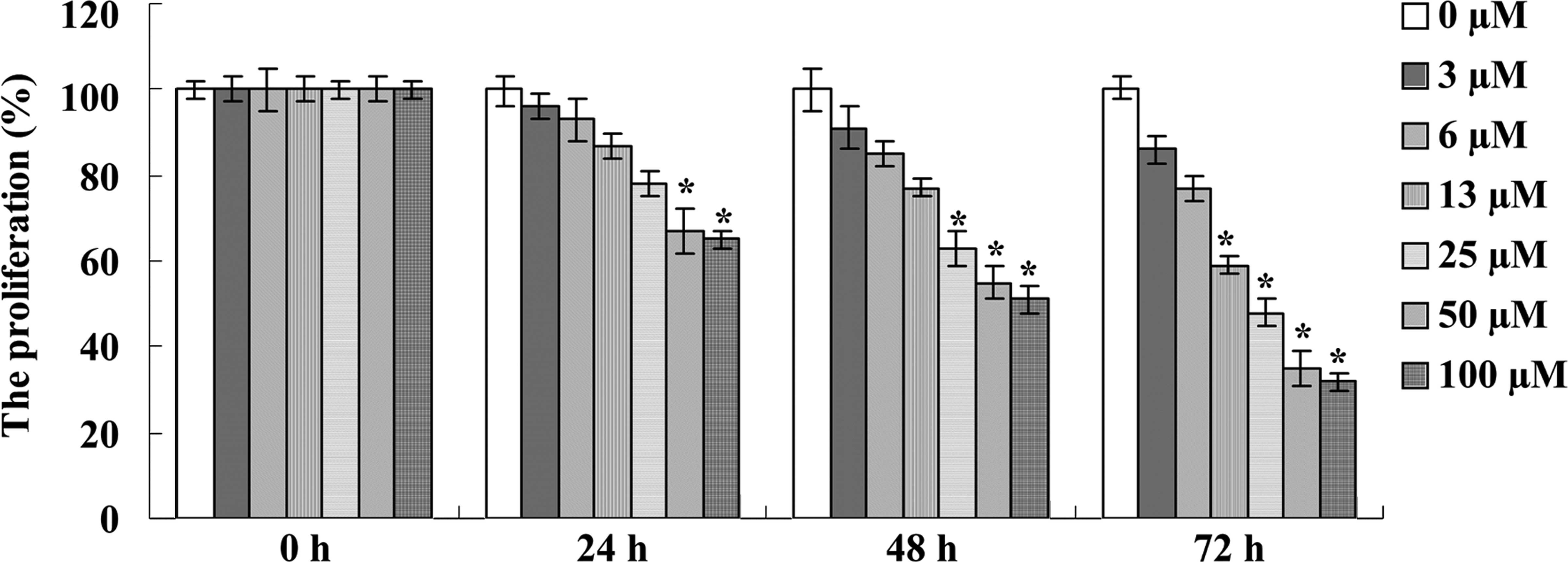
assays were performed. As expected, icariin treatment clearly
reduced the cell proliferation of the A2780 cells in a dose- and
time-dependent manner in comparison with the 0 μM icariin
treatment group (Fig. 2).
Particularly, treatment with icariin (50 and 100 μM)
significantly inhibited the proliferation of A2780 cells at 24, 48
and 72 h, with statistical significance (Fig. 2). Meanwhile, treatment with 25
μM of icariin for 48 and 72 h significantly reduced the
proliferation of A2780 cells and treatment with 13 μM of
icariin for 72 h also significantly reduced the proliferation of
A2780 cells (Fig. 2). Based on the
MTT assay results, icariin treatments at final concentrations of
13, 25 and 50 μM at 48 h were chosen for future study.
Icariin promotes the cell apoptosis of
ovarian cancer cells
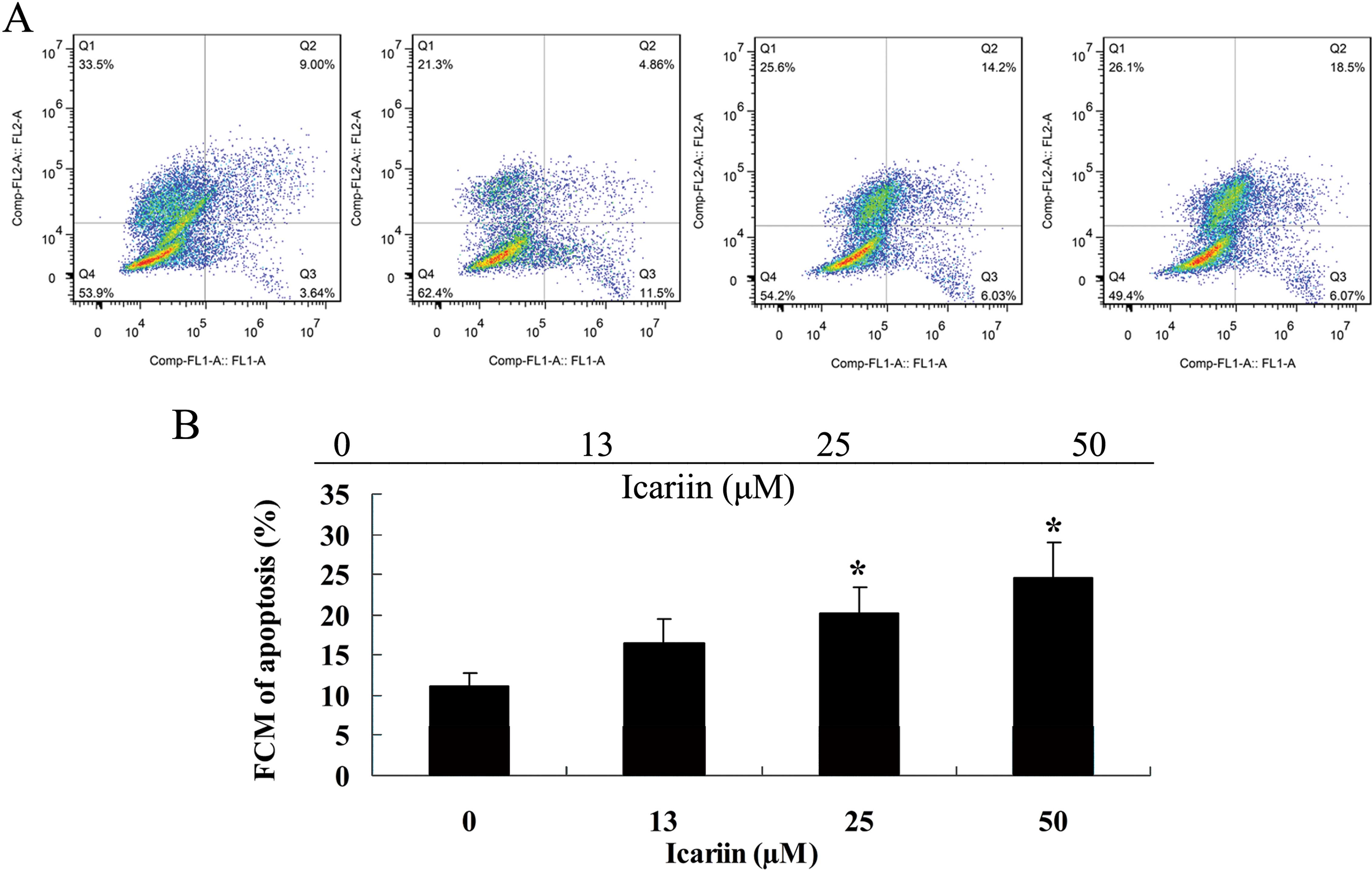
In our study, we determined that icariin promoted
the cell apoptosis of ovarian cancer A2780 cells. Thus, flow
cytometry was used to measure the cell apoptosis of A2780 cells.
Notably, a significant increase in cell apoptosis was observed
following the icariin treatment at the final concentrations of 25
and 50 μM for 48 h (Fig. 3A and
B).
Icariin induces caspase-3 activity in
ovarian cancer cells
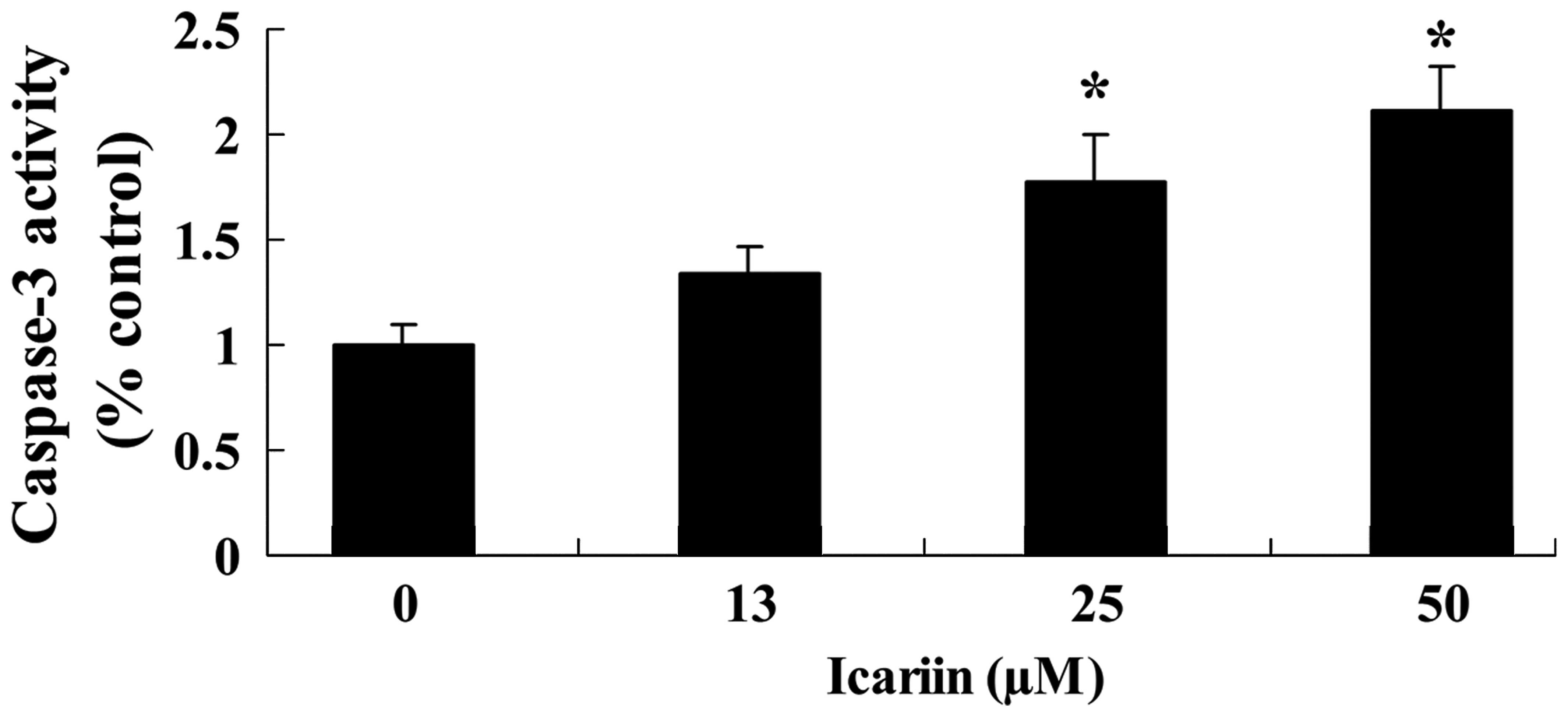
To further investigate whether icariin induces
caspase-3 activity of ovarian cancer A2780 cells, caspase-3
colorimetric assays were employed. Following treatment with icariin
at concentrations of 25 and 50 μM for 48 h, the
dose-dependent caspase-3 activity was obviously increased, when
compared with the activity in the 0 μM icariin treatment
group (Fig. 4). These results
clearly indicated that icariin efficiently induced apoptosis of the
A2780 cells, and this apoptotic effect was exerted in a
dose-dependent manner (Figs. 3 and
4).
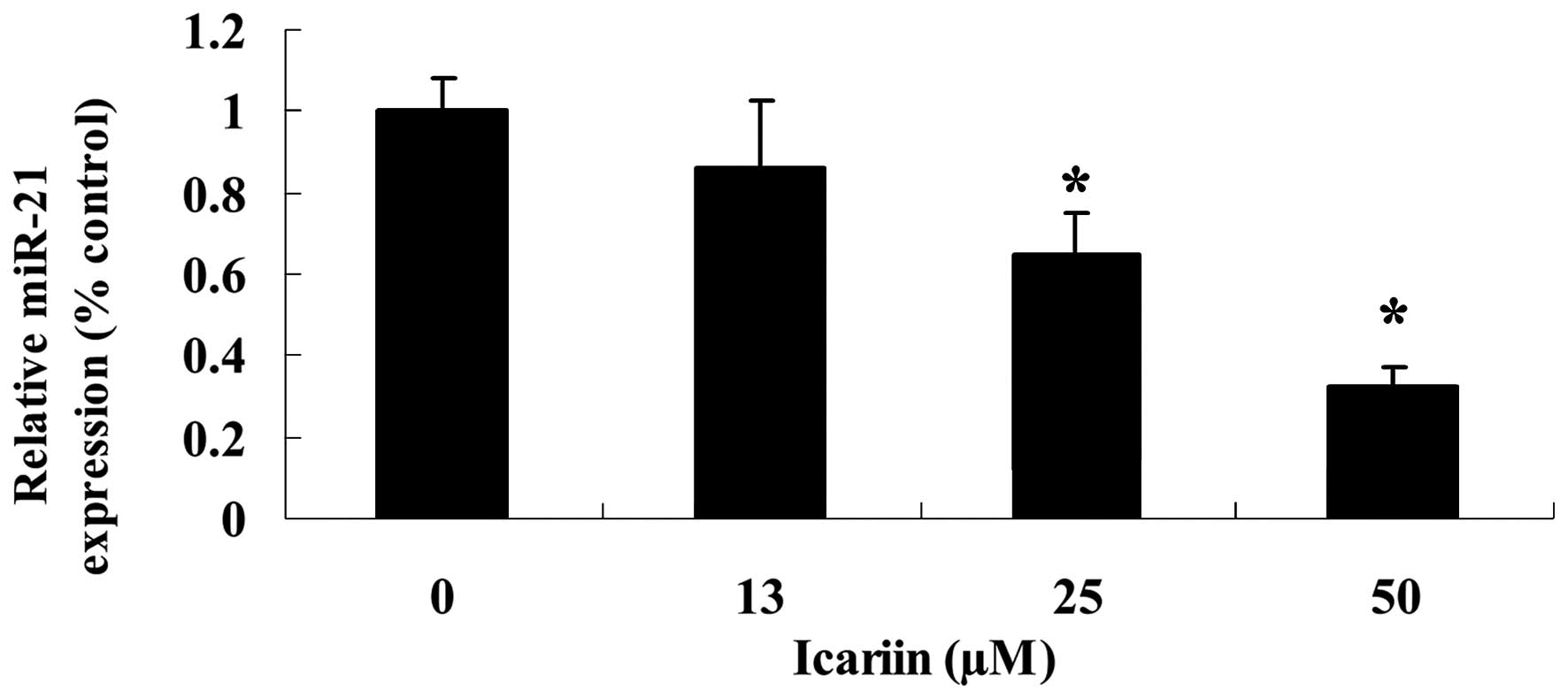
Icariin suppresses miR-21 expression in
ovarian cancer cells
To determine whether the expression of miR-21 is
correlated with icariin treatment, we evaluated the expression
level of miR-21 in the A2780 cells. As shown in Fig. 5, the miR-21 expression level in the
A2780 cells was significantly decreased following treatment with
icariin at concentrations of 25 and 50 μM for 48 h, compared
with the 0 μM icariin treatment group.
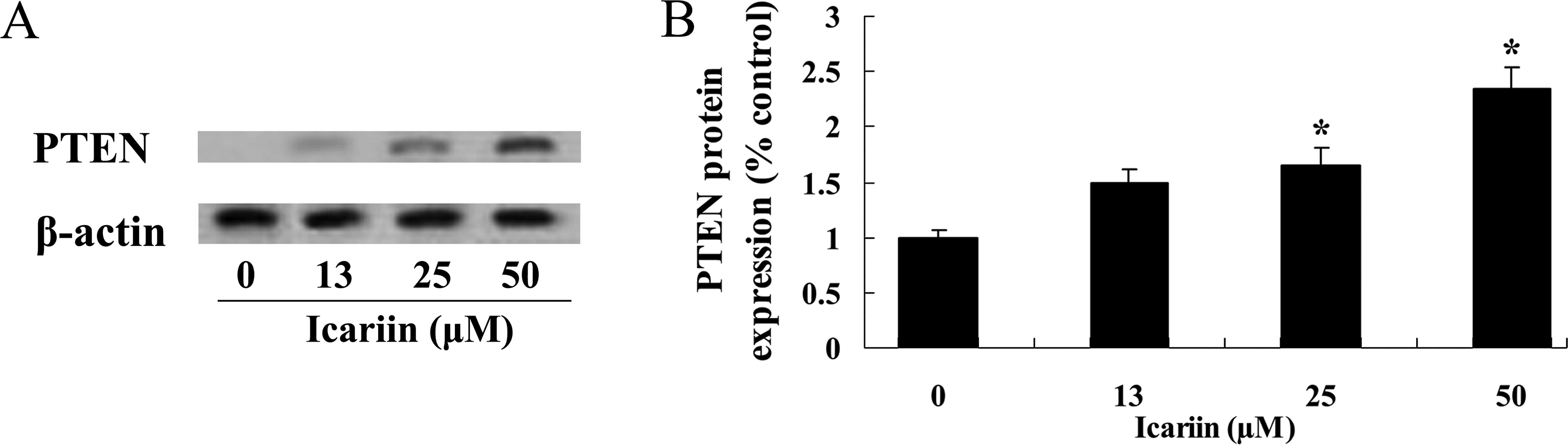
Icariin increases PTEN protein expression
in ovarian cancer cells
We determined the expression of PTEN in the
potential anticancer effect of icariin to study the role of miR-21
in ovarian cancer A2780 cells. As a result, icariin (25 and 50
μM) effectively increased PTEN protein expression in the
A2780 cells at 48 h, compared with that in the 0 μM icariin
treatment group (Fig. 6A and
B).
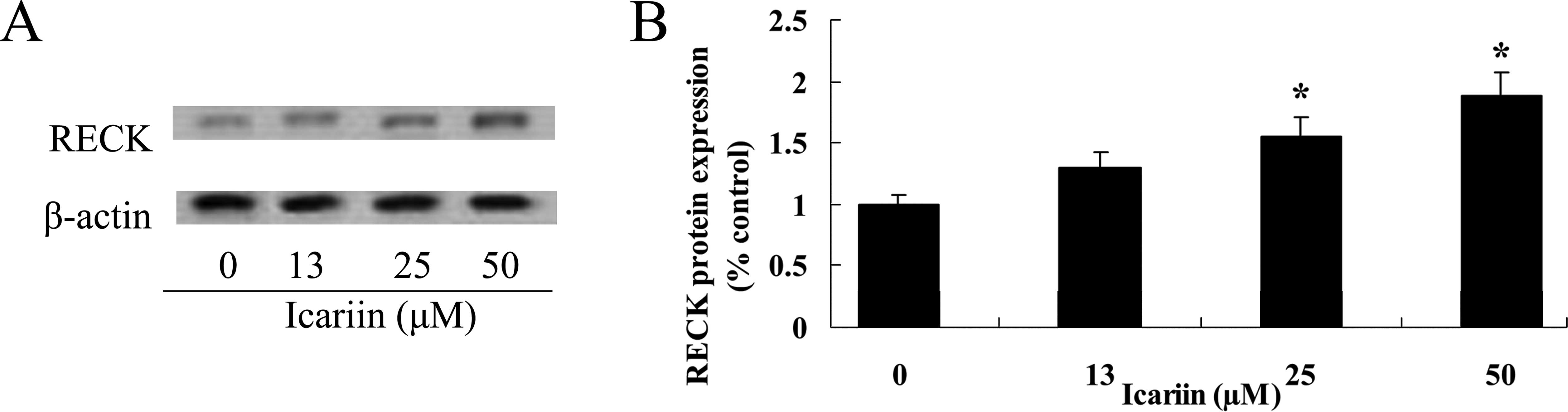
Icariin increases RECK protein expression
in ovarian cancer cells
To evaluate the potential efficacy of icariin
treatment on RECK protein in the ovarian cancer A2780 cells, we
detected RECK protein expression levels. Following treatment with
icariin at concentrations of 25 and 50 μM for 48 h, the RECK
protein expression level was markedly increased in comparison to
the level in the 0 μM icariin treatment group (Fig. 7A and B).
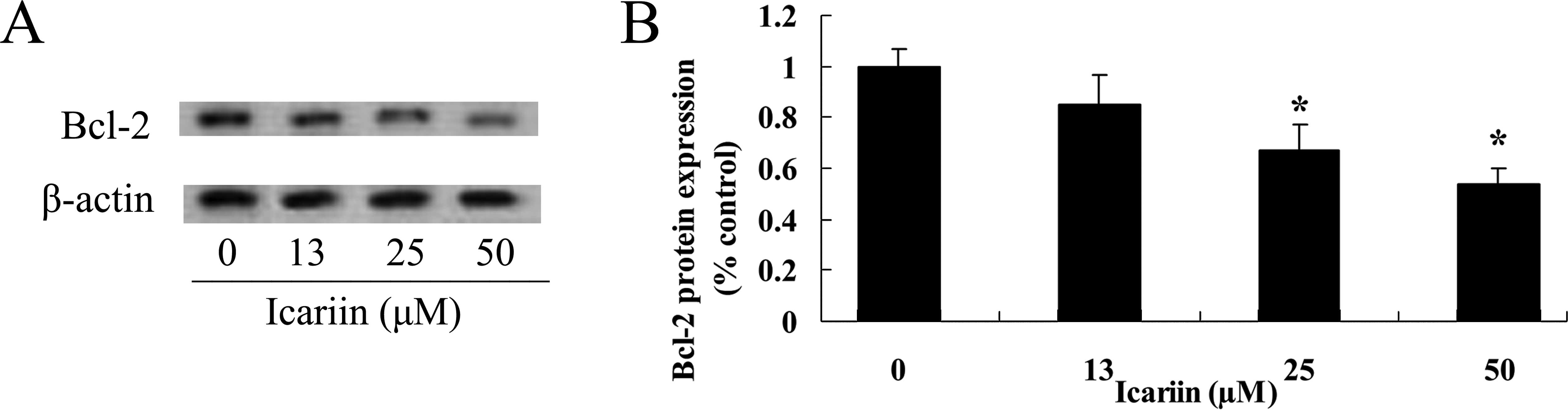
Icariin suppresses Bcl-2 protein
expression in the ovarian cancer cells
To further investigate the potential effect of
icariin on Bcl-2 protein expression in the ovarian cancer A2780
cells, the Bcl-2 protein expression level in the A2780 cells was
measured. As shown in Fig. 8A and
B, the Bcl-2 protein expression level was decreased following
treatment with icariin at concentrations of 25 and 50 μM for
48 h, compared with that in the 0 μM icariin treatment
group.
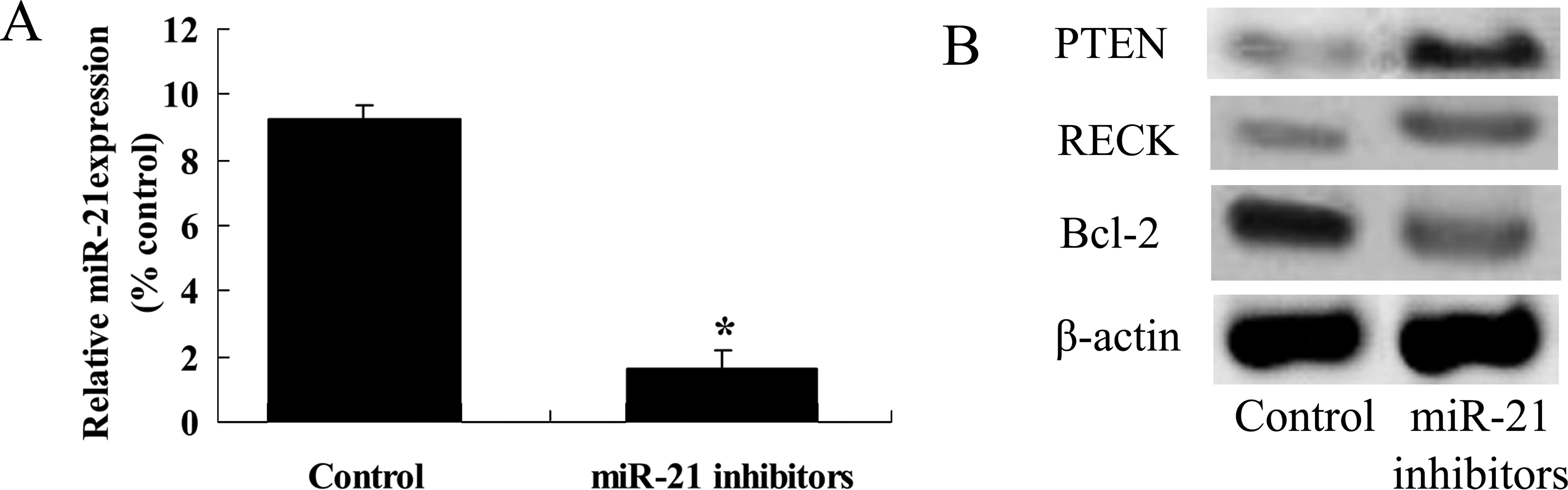
Anti-miR-21 influences the expression
levels of PTEN, RECK and Bcl-2 protein in the ovarian cancer
cells
We selected miR-21 and its potential target proteins
PTEN, RECK and Bcl-2 to study the effect of icariin on ovarian
cancer A2780 cells. The miR-21 inhibitor was transfected with
Lipofectamine 2000 into the A2780 cells. We found that miR-21
expression was effectively silenced in the A2780 cells, compared
with the control group (Fig. 9A).
In addition, PTEN and RECK protein expression levels were increased
in the A2780 cells, compared with these levels in the control group
(Fig. 9B). However, Bcl-2 protein
expression level in the A2780 cells was reduced in comparison with
the level in the control group (Fig.
9B).
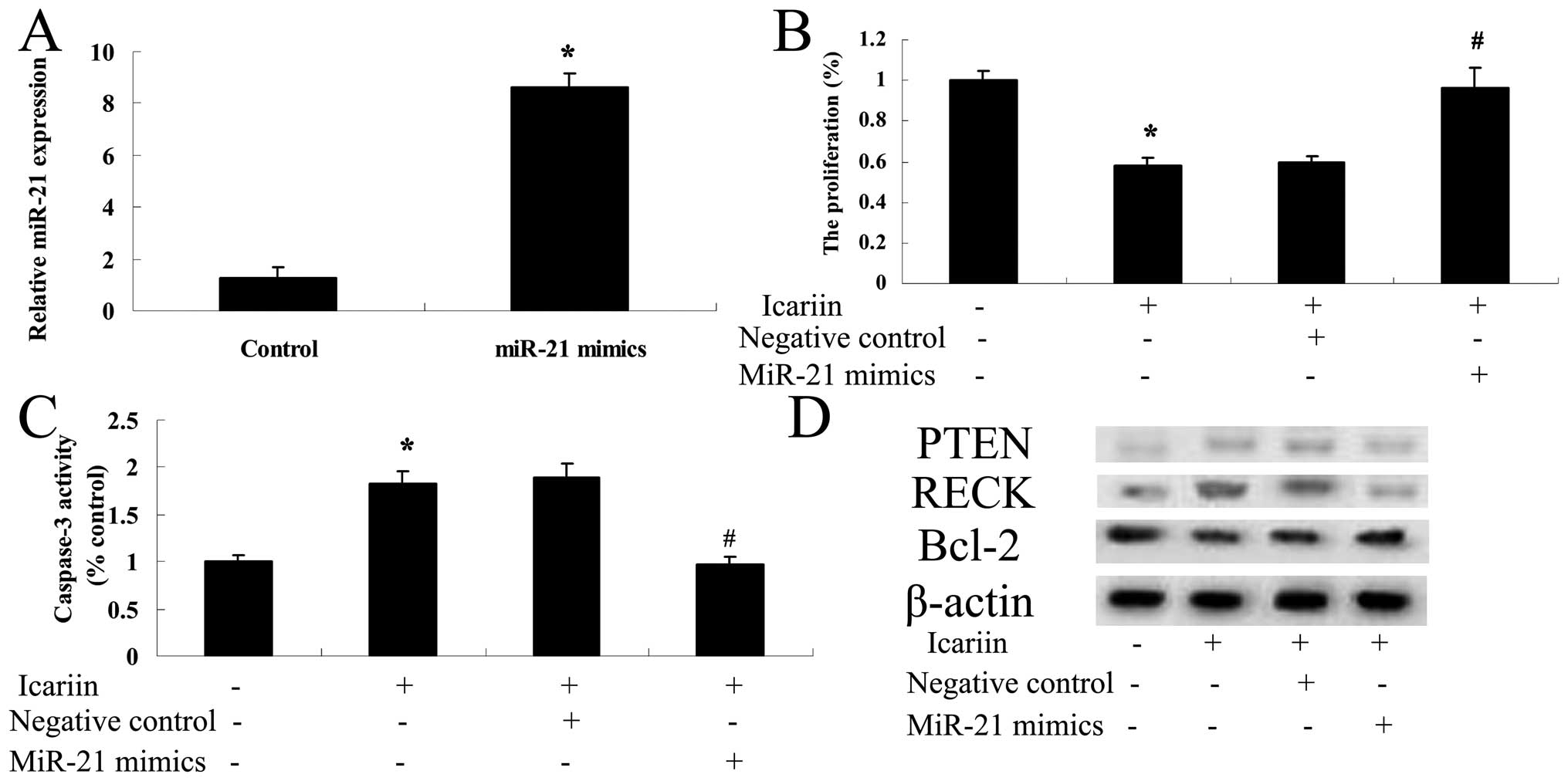
Overexpression of miR-21 influences the
effect of icariin on ovarian cancer cells
To further investigate the potential connection
between overexpression of miR-21 and the potential effect of
icariin on ovarian cancer A2780 cells, miR-21 mimics were
transfected with Lipofectamine 2000 into the A2780 cells. Our
results indicated that transfection of miR-21 significantly
increased the expression level of miR-21 in the A2780 cells
(Fig. 10A). miR-21 mimics reversed
the potential effect of icariin on A2780 cell proliferation and
reduced activation of caspase-3 (Fig.
10B and C). Moreover, PTEN and RECK protein expression levels
were decreased in the A2780 cells (Fig. 10D). In contrast, the Bcl-2 protein
expression level in the A2780 cells was increased (Fig. 10D).
Discussion
Malignant ovarian tumor is one of the three most
common female genital cancers. During the past two decades, the
application of effective chemotherapy has significantly improved
the treatment of malignant ovarian germ cell tumors, with a decline
in the mortality rate from 90 to 10% (25). However, the therapeutic effect of
malignant ovarian epithelial tumors has not improved. Malignant
ovarian epithelial cancer has become one of the major gynecological
cancers, seriously threatening the health and lives of females
(26). In the present study,
icariin reduced the cell proliferation of human ovarian cancer
A2780 cells in a dose- and time-dependent manner. Meanwhile,
icariin induced cell apoptosis and enhanced caspase-3 activity in
the A2780 cells. Our results were consistent with those of other
studies. For example, Li et al found that icariin induced
apoptosis of human hepatoma cells via an ROS/JNK-dependent
mitochondrial pathway (27). Yang
et al found that icariin exhibited antitumor activity in
glioblastoma by inhibiting NF-κB activity (22). Zhang et al reported that
icariin suppressed cell proliferation, induced apoptosis, and
enhanced caspase-3 activity of gallbladder cancer through
suppression of the expression of Bcl-2 (28).
Recent studies have shown that miRs play an
important role in the occurrence and development of tumors through
interaction with mRNAs, involved in proliferation, differentiation,
apoptosis and metastasis processes of tumor cells (29). miR-21 has been identified as an
oncogene, and is abnormally highly expressed in a variety of tumor
tissues through the regulation of tumor-suppressor genes such as
NFIB, PDCD4, PTEN and SPRY2 (30).
The present study showed that icariin increased the miR-21
expression level in ovarian cancer A2780 cells. Zhang et al
reported that icariin adjusted miR-21 in mouse preimplantation
embryos (31). The present study
for the first time investigated the role of miR-21 in the
anticancer effects of icariin on ovarian cancer cells. However, the
particular mechanisms of how icariin influences miR-21 expression
are still unclear, and further clarification is need.
Low expression of PTEN protein is involved in
evolutionary processes of multiple malignant tumors such as
prostate and endometrial cancer (32). Studies have shown that PTEN inhibits
the motility of tumor cells and angiogenesis by regulating matrix
metalloproteinases and vascular endothelial growth factors;
moreover, PTEN can dephosphorylate focal adhesion kinase, in which
the latter downregulates the MAPK pathway of mitogen-activated
protein kinase, thereby regulating cell adhesion (33). Consistently, we found that icariin
effectively increased PTEN protein expression in the ovarian cancer
A2780 cells. Our results were in keeping with those of other
studies. For example, Zhang et al reported that icariin
adjusted PTEN protein expression of mouse preimplantation embryos
(31). However, the detailed
mechanisms on how icariin induces PTEN protein expression in A2780
cells are quite unclear, and further explanation is needed.
RECK protein in patients with deep infiltrative
tongue cancer, and in patients with lymph node metastasis is
decreased. Thus, RECK protein is considered as an important
regulatory protein in inhibiting tumor cell invasion and
metastasis. A low level of expression increased the rates of tongue
cancer invasion and metastasis, and the regulation may be related
to an inhibitory mechanism of RECK on MMP-2 and MMP-9 expression
(34). RECK shows an obviously
negative correlation with MMP-2 and MMP-9 expression, suggesting
that in the process of tumor invasion and metastasis, the
expression of RECK is downregulated, limiting the inhibition of
RECK on MMP-2 and MMP-9, thus achieving invasion and metastasis of
tumor tissues (35). The present
study first demonstrated that icariin increased the RECK protein
expression level in ovarian cancer A2780 cells. This result
demonstrated the potential connection between the anticancer
effects of icariin on cell apoptosis and RECK protein expression.
Thus, the specific mechanisms of how icariin induces RECK protein
expression in A2780 cells remain unclear and further explanation is
needed.
Apoptosis is a form of programmed cell death
regulated by genes. Malignant tumors often exhibit increased
proliferation and decreased apoptosis. The Bcl-2 family is a group
of genes controlling apoptosis, and the Bcl-2 gene has an
anti-apoptotic effect (36). Under
normal circumstances, Bcl-2 shows cyclical low level expression;
yet overexpression of Bcl-2 can inhibit apoptosis. The Bax gene
belongs to the same family of Bcl-2 genes, with an opposite
function of Bcl-2. Bax promotes apoptosis (37). The dynamic equilibrium between Bcl-2
and Bax is the main mechanism in the regulation of normal
apoptosis, and recently many researchers have observed that the
level of Bcl-2 protein is related to the sensitivity and resistance
of ovarian tumor cells to chemotherapy (38). In the present study, we demonstrated
that the Bcl-2 protein expression level was decreased by icariin.
This result was in accordance with the findings of other studies.
For example, Wang et al reported that icariin induced
apoptosis through regulation of the expression of Bcl-2/Bax, and
activation of caspase-9 and -3 in mouse MLTC-10 Leydig tumor cells
(24). Li et al found that
icariin triggered the mitochondrial/caspase-mediated apoptosis of
human hepatoma SMMC-7721 cells by suppression of the Bcl-2/Bax
ratio (27).
To illuminate the mechanism involved in the
suppression of A2780 cell proliferation, the effect of icariin on
miR-21 and its target genes, including PTEN, RECK and Bcl-2 was
assessed in the A2780 cells. We first found that the miR-21
inhibitor increased PTEN and RECK protein expression levels and
decreased the Bcl-2 protein expression level in the ovarian cancer
A2780 cells. Then, overexpression of miR-21 suppressed the PTEN and
RECK protein expression levels in the A2780 cells. Notably, our
data revealed the positive regulation of miR-21, yet negative
regulation of the anticancer effect of icariin on cell
proliferation and apoptosis in the A2780 cells. These results were
in accord with that of other studies. For example, Xu et al
demonstrated that suppression of miR-21 decreased cell
proliferation and enhanced apoptosis of lung carcinoma by
increasing PTEN and RECK and decreasing Bcl-2 mRNA expression
(39). The accurate mechanism of
how it occurs needs to be further studied.
In summary, the results of the present study further
showed the potential anticancer curative effect of icariin on
ovarian cancer, and its regulatory mechanism of miR-21 subsequently
targeting PTEN, RECK and Bcl-2 in A2780 cells. In particular, our
findings revealed that upregulation of miR-21 promoted the
progression of ovarian cancer cells through negative regulation of
PTEN and RECK expression levels and positive regulation of Bcl-2
expression. We propose that the potential anticancer curative
effect of icariin is through the reduction in cell proliferation
and promotion of apoptosis through miR-21 and its target genes
(PTEN, RECK and Bcl-2) in A2780 cells. We need to further study the
molecular mechanisms of icariin on tumorigenesis and
progression.
References
|
1
|
Luo J, Zhou J, Cheng Q, Zhou C and Ding Z:
Role of microRNA-133a in epithelial ovarian cancer pathogenesis and
progression. Oncol Lett. 7:1043–1048. 2014.PubMed/NCBI
|
|
2
|
Tang AQ, Cao XC, Tian L, He L and Liu F:
Apigenin inhibits the self-renewal capacity of human ovarian cancer
SKOV3-derived sphere-forming cells. Mol Med Rep. 11:2221–2226.
2015.
|
|
3
|
Hays JL, Kim G, Walker A, Annunziata CM,
Lee JM, Squires J, Houston N, Steinberg SM and Kohn EC: A phase II
clinical trial of polyethylene glycol-conjugated L-asparaginase in
patients with advanced ovarian cancer: Early closure for safety.
Mol Clin Oncol. 1:565–569. 2013.
|
|
4
|
Chen WC, Lin MS, Ye YL, Gao HJ, Song ZY
and Shen XY: microRNA expression pattern and its alteration
following celecoxib intervention in human colorectal cancer. Exp
Ther Med. 3:1039–1048. 2012.PubMed/NCBI
|
|
5
|
Jiang JX, Zhang N, Liu ZM and Wang YY:
Detection of microRNA-21 expression as a potential screening
biomarker for colorectal cancer: A meta-analysis. Asian Pac J
Cancer Prev. 15:7583–7588. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chan JK, Blansit K, Kiet T, Sherman A,
Wong G, Earle C and Bourguignon LY: The inhibition of miR-21
promotes apoptosis and chemosensitivity in ovarian cancer. Gynecol
Oncol. 132:739–744. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liu S, Fang Y, Shen H, Xu W and Li H:
Berberine sensitizes ovarian cancer cells to cisplatin through
miR-21/PDCD4 axis. Acta Biochim Biophys Sin. 45:756–762. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yang L, Kuang LG, Zheng HC, Li JY, Wu DY,
Zhang SM and Xin Y: PTEN encoding product: A marker for
tumorigenesis and progression of gastric carcinoma. World J
Gastroenterol. 9:35–39. 2003.PubMed/NCBI
|
|
9
|
Wu H, Wang K, Liu W and Hao Q: PTEN
overexpression improves cisplatin-resistance of human ovarian
cancer cells through upregulating KRT10 expression. Biochem Biophys
Res Commun. 444:141–146. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lau MT, Klausen C and Leung PC: E-cadherin
inhibits tumor cell growth by suppressing PI3K/Akt signaling via
β-catenin-Egr1-mediated PTEN expression. Oncogene. 30:2753–2766.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lou Y, Yang X, Wang F, Cui Z and Huang Y:
MicroRNA-21 promotes the cell proliferation, invasion and migration
abilities in ovarian epithelial carcinomas through inhibiting the
expression of PTEN protein. Int J Mol Med. 26:819–827. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhou X, Huang S, Jiang L, Zhang S, Li W,
Chen Z and Zhang D: Expression of RECK and MMP-2 in salivary
adenoid cystic carcinoma: Correlation with tumor progression and
patient prognosis. Oncol Lett. 7:1549–1555. 2014.PubMed/NCBI
|
|
13
|
Xue G, Zou X, Zhou JY, Sun W, Wu J, Xu JL
and Wang RP: Raddeanin A induces human gastric cancer cells
apoptosis and inhibits their invasion in vitro. Biochem Biophys Res
Commun. 439:196–202. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Masui T, Doi R, Koshiba T, Fujimoto K,
Tsuji S, Nakajima S, Koizumi M, Toyoda E, Tulachan S, Ito D, et al:
RECK expression in pancreatic cancer: Its correlation with lower
invasiveness and better prognosis. Clin Cancer Res. 9:1779–1784.
2003.PubMed/NCBI
|
|
15
|
Zhang Y, Cheng S, Zhang G, Ma W, Liu Y,
Zhao R, Zhang Q and Pang D: Low expression of RECK indicates a
shorter survival for patients with invasive breast cancer. Cancer
Sci. 103:1084–1089. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Qi Q, Lu N, Li C, Zhao J, Liu W, You Q and
Guo Q: Involvement of RECK in gambogic acid induced anti-invasive
effect in A549 human lung carcinoma cells. Mol Carcinog. Feb
14–2014.Epub ahead of print. View
Article : Google Scholar
|
|
17
|
Fejzo MS, Ginther C, Dering J, Anderson L,
Venkatesan N, Konecny G, Karlan B and Slamon DJ: Knockdown of
ovarian cancer amplification target ADRM1 leads to downregulation
of GIPC1 and upregulation of RECK. Genes Chromosomes Cancer.
50:434–441. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ahn MY, Kang DO, Na YJ, Yoon S, Choi WS,
Kang KW, Chung HY, Jung JH, Min do S and Kim HS: Histone
deacetylase inhibitor, apicidin, inhibits human ovarian cancer cell
migration via class II histone deacetylase 4 silencing. Cancer
Lett. 325:189–199. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sadahira K, Sagawa M, Nakazato T, Uchida
H, Ikeda Y, Okamoto S, Nakajima H and Kizaki M: Gossypol induces
apoptosis in multiple myeloma cells by inhibition of interleukin-6
signaling and Bcl-2/Mcl-1 pathway. Int J Oncol. 45:2278–2286.
2014.PubMed/NCBI
|
|
20
|
Xie YL, Yang YJ, Tang C, Sheng HJ, Jiang
Y, Han K and Ding LJ: Estrogen combined with progesterone decreases
cell proliferation and inhibits the expression of Bcl-2 via
microRNA let-7a and miR-34b in ovarian cancer cells. Clin Transl
Oncol. 16:898–905. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ma C, Yin G, You F, Wei Y, Huang Z, Chen X
and Yan D: A specific cell-penetrating peptide induces apoptosis in
SKOV3 cells by down-regulation of Bcl-2. Biotechnol Lett.
35:1791–1797. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang L, Wang Y, Guo H and Guo M:
Synergistic anti-cancer effects of icariin and temozolomide in
glioblastoma. Cell Biochem Biophys. Nov 11–2014.Epub ahead of
print.
|
|
23
|
Zhou J, Wu J, Chen X, Fortenbery N,
Eksioglu E, Kodumudi KN, Pk EB, Dong J, Djeu JY and Wei S: Icariin
and its derivative, ICT, exert anti-inflammatory, anti-tumor
effects, and modulate myeloid derived suppressive cells (MDSCs)
functions. Int Immunopharmacol. 11:890–898. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang Q, Hao J, Pu J, Zhao L, Lü Z, Hu J,
Yu Q, Wang Y, Xie Y and Li G: Icariin induces apoptosis in mouse
MLTC-10 Leydig tumor cells through activation of the mitochondrial
pathway and down-regulation of the expression of piwil4. Int J
Oncol. 39:973–980. 2011.PubMed/NCBI
|
|
25
|
Nakamura K, Banno K, Yanokura M, Iida M,
Adachi M, Masuda K, Ueki A, Kobayashi Y, Nomura H, Hirasawa A, et
al: Features of ovarian cancer in Lynch syndrome (Review). Mol Clin
Oncol. 2:909–916. 2014.PubMed/NCBI
|
|
26
|
Li S, Li Y, Wen Z, Kong F, Guan X and Liu
W: microRNA-206 overexpression inhibits cellular proliferation and
invasion of estrogen receptor α-positive ovarian cancer cells. Mol
Med Rep. 9:1703–1708. 2014.PubMed/NCBI
|
|
27
|
Li S, Dong P, Wang J, Zhang J, Gu J, Wu X,
Wu W, Fei X, Zhang Z, Wang Y, et al: Icariin, a natural flavonol
glycoside, induces apoptosis in human hepatoma SMMC-7721 cells via
a ROS/JNK-dependent mitochondrial pathway. Cancer Lett.
298:222–230. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang DC, Liu JL, Ding YB, Xia JG and Chen
GY: Icariin potentiates the antitumor activity of gemcitabine in
gallbladder cancer by suppressing NF-κB. Acta Pharmacol Sin.
34:301–308. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang B and Zhang Q: The expression and
clinical significance of circulating microRNA-21 in serum of five
solid tumors. J Cancer Res Clin Oncol. 138:1659–1666. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu M, Tang Q, Qiu M, Lang N, Li M, Zheng
Y and Bi F: miR-21 targets the tumor suppressor RhoB and regulates
proliferation, invasion and apoptosis in colorectal cancer cells.
FEBS Lett. 585:2998–3005. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang C, Shi YR, Liu XR, Cao YC, Tian JL,
Jia ZY, Zhen D, Liu FH and Gao JM: The regulatory role of icariin
on apoptosis in mouse preimplantation embryos with reduced
microRNA-21. Theriogenology. 82:461–468. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Waite KA and Eng C: Protean PTEN: Form and
function. Am J Hum Genet. 70:829–844. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Gu J, Tamura M and Yamada KM: Tumor
suppressor PTEN inhibits integrin- and growth factor-mediated
mitogen-activated protein (MAP) kinase signaling pathways. J Cell
Biol. 143:1375–1383. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Alexius-Lindgren M, Andersson E, Lindstedt
I and Engström W: The RECK gene and biological malignancy - its
significance in angiogenesis and inhibition of matrix
metalloproteinases. Anticancer Res. 34:3867–3873. 2014.PubMed/NCBI
|
|
35
|
Prosdócimi FC, Rodini CO, Sogayar MC,
Sousa SC, Xavier FC and Paiva KB: Calcifying cystic odontogenic
tumour: Immunohistochemical expression of matrix
metalloproteinases, their inhibitors (TIMPs and RECK) and inducer
(EMMPRIN). J Oral Pathol Med. 43:545–553. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu N, Zheng Y, Zhu Y, Xiong S and Chu Y:
Selective impairment of
CD4+CD25+Foxp3+ regulatory T cells
by paclitaxel is explained by Bcl-2/Bax mediated apoptosis. Int
Immunopharmacol. 11:212–219. 2011. View Article : Google Scholar
|
|
37
|
Li J, Sun GZ, Lin HS, Pei YX, Qi X, An C,
Yu J and Hua BJ: The herb medicine formula ‘Yang Wei Kang Liu’
improves the survival of late stage gastric cancer patients and
induces the apoptosis of human gastric cancer cell line through
Fas/Fas ligand and Bax/Bcl-2 pathways. Int Immunopharmacol.
8:1196–1206. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yao Y, Huang C, Li ZF, Wang AY, Liu LY,
Zhao XG, Luo Y, Ni L, Zhang WG and Song TS: Exogenous
phosphatidylethanolamine induces apoptosis of human hepatoma HepG2
cells via the bcl-2/Bax pathway. World J Gastroenterol.
15:1751–1758. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Xu LF, Wu ZP, Chen Y, Zhu QS, Hamidi S and
Navab R: MicroRNA-21 (miR-21) regulates cellular proliferation,
invasion, migration, and apoptosis by targeting PTEN, RECK and
Bcl-2 in lung squamous carcinoma, Gejiu City, China. PLoS One.
9:e1036982014. View Article : Google Scholar : PubMed/NCBI
|