Introduction
Epstein-Barr virus (EBV) is a type of human herpes
virus with a genome of double-stranded DNA and its natural host is
only the human (1,2). EBV has been reported to be closely
associated with a variety of malignant lymphoma and epithelial
cancers, particularly Burkitt’s lymphoma and nasopharyngeal
carcinoma (NPC). The tumorigenesis of NPC is a multifactor,
multistep and multiple gene process, in which the relationship
between EBV and NPC has been confirmed in most studies.
Furthermore, in most poorly differentiated and undifferentiated
squamous cell carcinoma the existence of EBV can be detected
(3). By detecting the EBV genome in
NPC tissues, it was shown that clusters of EBV tended to densely
distribute in carcinoma nests, particularly in the center and
periphery of tumor tissues, which suggests that NPC may originate
from a mass of EBV-infected cancer cells (4). Due to the lack of the CD21 receptor,
EBV cannot directly infect epithelial cells (5,6), which
is the main reason that EBV-stable transformational epithelial cell
models and its tumorigenic animal models are scarce. Consequently,
the mechanism of the tumorigenesis of EBV-associated NPC remains
unclear.
The short palate, lung and nasal epithelium clone 1
(SPLUNC1), also called nasopharynx associated-specific gene (NASG),
is a tissue-specific gene of the nasopharyngeal epithelium that has
been cloned by suppression subtractive hybridization and cDNA
microarray in our laboratory (7,8). As a
secretory protein, SPLUNC1 is specifically expressed in respiratory
mucosal cells of the upper respiratory tract. Moreover, SPLUNC1
plays a vital role in mediating the defense response in the host
respiratory tract to maintain normal physiological activity and
exert antimicrobial and anti-inflammatory activity and tumor
suppression. In recent years, some studies point out that SPLUNC1
has a close relationship with the inflammatory response. For
example, SPLUNC1 was able to regulate Pseudomonas
aeruginosa-induced lung inflammation in mice (9), and was found to be markedly
downregulated in patients with chronic sinusitis (10). SPLUNC1 was found to modulate the
inflammatory response to resist pulmonary inhaled particles by
increasing leukocyte recruitment and phagocytic activity (11). However, the mechanism of how SPLUNC1
effects EBV-infected NPC has not been reported to date. In the
present study, on the basis of two established types of cell
infection, we confirmed that EBV infection of NPC cells activated
the Toll-like receptor (TLR)9/NF-κB signaling pathway, promoted
inflammatory cytokine release and consequently enhanced the
inflammatory response; while SPLUNC1 weakened the inflammatory
response induced by EBV infection in NPC cells through regulation
of the TLR9/NF-κB signaling pathway and control of the tumor
inflammatory microenvironment thus playing a regulatory role in
tumorigenesis. Thus, the regulatory chain of EBV-SPLUNC1-TLR9/NF-κB
inflammation was designed to illustrate the inflammatory regulation
mechanism of NPC tumorigenesis.
Materials and methods
Ethics
The experiments were undertaken following the
understanding and written consent of each subject. The study
methodologies were approved by the Medical Ethics Committee of The
Affiliated Tumor Hospital of Xinjiang Medical University (no.
W201327) in accordance with the Code of Ethics of the World Medical
Association (Declaration of Helsinki) for experiments involving
humans.
Immunohistochemical assessment
Eight paraffin-embedded pathological biopsy samples
of NPC were obtained from patients presenting with no other
malignancies and inflammatory diseases (stage III, 48–73 years of
age, females/males 3/5, no preoperative chemoradiotherapy) and 8
paraffin-embedded pathological biopsy samples of normal
nasopharyngeal tissues (healthy individuals).
The protein expression levels of SPLUNC1 and TLR9
were determined by EliVision™ two-step immunohistochemical method
in the normal nasopharyngeal and NPC tissues. Briefly,
paraffin-embedded sections (4-μm) of the tissues were
dewaxed at 60°C for 1 h and rehydrated in gradient alcohol. For
antigen retrieval, the sections were incubated in sodium citrate
buffer (0.01 M, pH 6.0) for 15 min in a microwave oven (600 W).
After cooling to room temperature, endogenous peroxidase activity
was blocked with 3% H2O2 for 30 min at room
temperature. The slides were incubated with normal goat serum in
TBS (5%) for 30 min to reduce non-specific staining and then
incubated with mouse anti-human SPLUNC1 antibody (1:200) and mouse
anti-human TLR9 antibody (1:1,000) (both from Abcam, Cambridge, UK)
at 4°C overnight. After extensive washes with PBS, the slides were
then incubated with polymerized HRP-anti-mouse/rabbit IgG
(EliVision™ Plus/HRP kit; Maxin, Fuzhou, China) according to the
manufacturer’s instructions. Color reaction was developed using
liquid DAB (Maxin), and all slides were counterstained with
hematoxylin.
Cell cultures and plasmids
HEK-293 cells stably transfected with the p2089
plasmid (293-EBV) were cultured in Dulbecco’s modified Eagle’s
medium (DMEM) with 10% fetal calf serum (Therm, UK) (12). An immortalized normal nasopharyngeal
epithelial cell line NP69 was cultured in keratinocyte serum-free
medium (gifted by the Chinese University of Hong Kong) (13). The Raji cell line of human Burkitt’s
lymphoma is a EBV-infected and immortalized lymphocyte cell line
(provided by the GSF-National Research Center for Environment and
Health, Germany). Highly differentiated CNE1 cells, poorly
differentiated HNE2 cells and undifferentiated 5–8F cells were all
purchased from the Cancer Research Institute, Sun Yat-Sen
University. HNE2 cells were stably transfected with SPLUNC1
(HNE2/SPLUNC1 cells) and or transfected with the vector alone
(HNE2/Vector cells), which were cultured in RPMI-1640 media
supplemented with 10% fetal bovine serum (FBS) (both from
Invitrogen, Carlsbad, CA, USA). All of the cells were placed in a
5% CO2 atmosphere at 37°C.
The coding region of the SPLUNC1 gene was inserted
into the pEGFP vector as previously described (14). The plasmid p2089 (Maxi-EBV), which
contained the complete EBV genome of the B95-8 strain was kindly
provided by Professor W. Hammerschmidt (GSF-National Research
Center for Environment and Health, Germany). Two eukaryotic
expression vectors with the EBV-related BZLF1 and BALF4 genes
(pcDNA3.1(+)/BZLF1 and pcDNA3.1(+)/BALF4) were constructed as
previously described (15).
The established model of EBV-infected
epithelial cells
Collection of green fluorescence protein-labeled EBV
(GFP-EBV) was as follows. The viral genes BZLF1 and BALF4 in the
pcDNA3.1(+) vector were transiently transfected into the 293-EBV
cells (12,16), and the GFP-EBV was released from the
transfected cells into the supernatant due to expression of BZLF1
and BALF4 which was induced by lytic replication of the 293-EBV
cells (17). After 72 h, the
supernatant was collected and filtered through a 0.45-mm pore-size
filter. Finally, the GFP-EBV was stored at −80°C.
EBV transfer infection
Through the cell-to-cell contact method in our
laboratory as previously described (12,18),
three human NPC cell lines (highly differentiated CNE1, poorly
differentiated HNE2 and undifferentiated 5–8F cells) were infected
with recombinant GFP-EBV produced from the 293-EBV cells. Briefly,
Raji cells were exposed to EBV produced from 293-EBV cells that
were induced with the expression plasmids BZLF1 and BALF4 for 3 h
at 4°C. However, in the control group, Raji cells were directly
treated at 4°C for 3 h. The acceptor epithelial cells had been
previously seeded in 2-ml wells 24 h prior to infection with
3×105 cells. After co-culture for up to 24 h, the Raji
cells were removed from the acceptors by washing. After co-culture
for 48 h, GFP-positive cells were observed using green fluorescence
microscopy, and the total RNA and protein were extracted from
acceptor epithelial cells for further detection.
Human-specific type B CpG oligodeoxynucleotides
(ODN) 2006 (5′-tcgtcgttttgtcgttttgtcgtt-3′) were resuspended in
endotoxin-free water and used as a positive control for TLR9
activation at the concentration as described in the literature
(lowercase letters are phosphorothioate linkage) (42).
RNA extraction and real-time quantitative
PCR (qRT-PCR)
Total RNA was isolated from the cells using TRIzol
reagent (Invitrogen). Reverse transcription reaction was performed
using a Fermentas Revert Aid First Strand cDNA Synthesis kit
(Fermentas, Burlington, ON, Canada), according to the
manufacturer’s instructions. qRT-PCR was performed using an iQ5
Multicolor Detection System (Bio-Rad, Hercules, CA, USA). The
following program was used for qPCR: 95°C for 30 sec followed by 40
cycles of 95°C for 5 sec and then 60°C for 30 sec. The relative
changes in expression were calculated using the 2−ΔΔCt
(where Ct is threshold cycle) method. Three parallel repeats were
performed for each sample in each experiment, and the results are
expressed as the mean of three independent experiments. All of the
qRT-PCR primers were designed by the online software Primer3 and
synthesized by Sangon Biotech, China. Primer sequences for the qPCR
were: TLR9 forward, 5′-CTGCCTTCCTACCCTGTGAG-3′ and reverse,
5′-GGATGCGGTTGGAGGACAA-3; LMP1 forward, 5′-TGAACACCACCACGATGACT-3′
and reverse, 5′-GTG CGCCTAGGTTTTGAGAG-3′; SPLUNC1 forward, 5′-CCC
ATTCAAGGTCTTCTGGA-3′ and reverse, 5′-CTGTAGTCC GTGGATCAGCA-3′; IL-6
forward, 5′-gaactccttctccacaa gcg-3′ and reverse,
5′-tctgaagaggtgagtggctg-3′; IL-8 forward,
5′-tccaaacctttccaccccaa-3′ and reverse, 5′-acttctccacaaccc
tctgc-3′; IL-1β forward, 5′-AGCCTCGTGCTGTCGGACCC-3′ and reverse,
5′-TCCAGCTGCAGGGTGGGTGT-3′; TNF-α forward,
5′-AGGGGCCACCACGCTCTTCT-3′ and reverse, 5′-CATGCCGTTGGCCAGGAGGG-3′;
GAPDH forward, 5′-ATCAAGATCATTGCTCCTCCTGAG-3′ and reverse,
5′-CTGCTTGCTGATCCACATCTG-3′.
Western blotting
Firstly, the cells were resuspended in lysis buffer
[1% Nonidet P-40, 50 mM Tris-HCl, pH 7.5, 50 mM NaF, 2 mM EDTA, 400
mM NaCl, 10% glycerol plus Complete protease inhibitor mixture
(Merck)]. Protein concentrations were determined using the
bicinchoninic acid protein assay kit (Pierce Chemical Co.,
Rockford, IL, USA) by a Bradford assay, and cell lysates (50
μg) were separated by 10% SDS-PAGE and transferred onto
nitrocellulose membranes (all from Bio-Rad). After the membranes
were blocked in Tris-buffered saline/Tween-20 (25 mM Tris-HCl, 150
mM NaCl, pH 7.5 and 0.05% Tween-20) with 5% defatted milk for 1 h
at 37°C, the membranes were incubated overnight at 4°C with the
primary antibodies in TBST with 5% defatted milk. After washing
with TBST, the membranes were incubated with horseradish
peroxidase-conjugated secondary antibodies (Santa Cruz
Biotechnology) and visualized using the ECL detection system.
TLR9 mouse anti-human (h) and LMP1 mouse anti-h
(Abcam); SPLUNC1 rabbit anti-h (Santa Cruz Biotechnology); CD14
rabbit anti-h/mouse (m) (Cell Signaling Technology, Danvers, MA,
USA); MyD88 rabbit anti-h/m (Santa Cruz Biotechnology); IKK rabbit
anti-h/m (Cell Signaling Technology); IKβα mouse anti-h/m, NF-κB
rabbit anti-h/m and p-NF-κB p65 (Ser 276) rabbit anti-h/m (all from
Santa Cruz Biotechnology); GAPDH rabbit anti-h/m and β-actin mouse
anti-h (Cell Signaling Technology) were used. HRP-labeled goat
anti-rabbit, mouse and rabbit anti-goat secondary antibodies were
purchased from Santa Cruz Biotechnology.
Virus purification and viral DNA
isolation
The EBV concentrated particles were collected via
‘the method of hunger’. After 7 days without replacing the medium,
the B95-8 cells were completely splitting and EBV particles were
fully released into the medium. Then, the virus particles were
precipitated under the condition of 12,000 rpm centrifugation for
2.5 h at 4°C and then it was dissolved in 1 ml fresh serum-free
medium. The concentrated EBV-B95-8 suspension was collected and
filtered through a 0.45-mm pore-size filter, and then stored at
−80°C. Viral dsDNA was purified from concentrated EBV-B95-8
suspension with a virus DNA extraction kit (Qiagen, Valencia, CA,
USA) and the concentration was determined by spectrophotometry (260
nm). The EBV DNA or CpG was transfected into the cells (NP69, HNE2,
HNE2/SPLUNC1 and HNE2/Vector) using Escort (Sigma-Aldrich, St.
Louis, MO, USA) according to the manufacturer’s instructions, and
the total RNA was extracted for further measurement.
Statistical analysis
The differences between groups were tested using the
Student’s t-test or one-way analysis of variance (ANOVA).
Calculations were performed using the SPSS 13.0 statistical
software. P<0.05 was considered to indicate a statistically
significant result.
Results
Tissue immunohistochemical results and
screening of high-efficient NPC cell lines infected by EBV
SPLUNC1 protein expression showed low or negative
expression in the NPC epithelial samples, while it demonstrated
positive expression in the normal nasopharyngeal epithelial
tissues. TLR9 protein expression exhibited positive expression in
the NPC epithelial samples, while it demonstrated low or negative
expression in the normal nasopharyngeal epithelial tissues
(Fig. 1A). In the 293-EBV cells
that stably expressed the plasmid p2089 (Maxi-EBV), green
fluorescence (GFP) was observed by inverted fluorescence microscopy
(Fig. 1B). After EBV infection of
the NPC cell lines for 48 h, the results showed that the poorly
differentiated HNE2 NPC cell line showed the strongest fluorescence
intensity, the undifferentiated NPC 5–8F cell line was second, and
the highly differentiated CNE1 NPC cell line showed the weakest
fluorescence intensity (Fig. 1C and
D). LMP1 mRNA expression gradually increased at different
time-points of 0, 12, 24 and 48 h after EBV infection of the HNE2
cells, with statistically significant differences between adjacent
time groups (P<0.001). After a 48-h EBV infection, the protein
expression of LMP1 in the HNE2 cell group was much higher than that
in the uninfected group (Fig. 1E).
These results revealed that EBV infection efficiency of the HNE2
cells gradually increased over time.
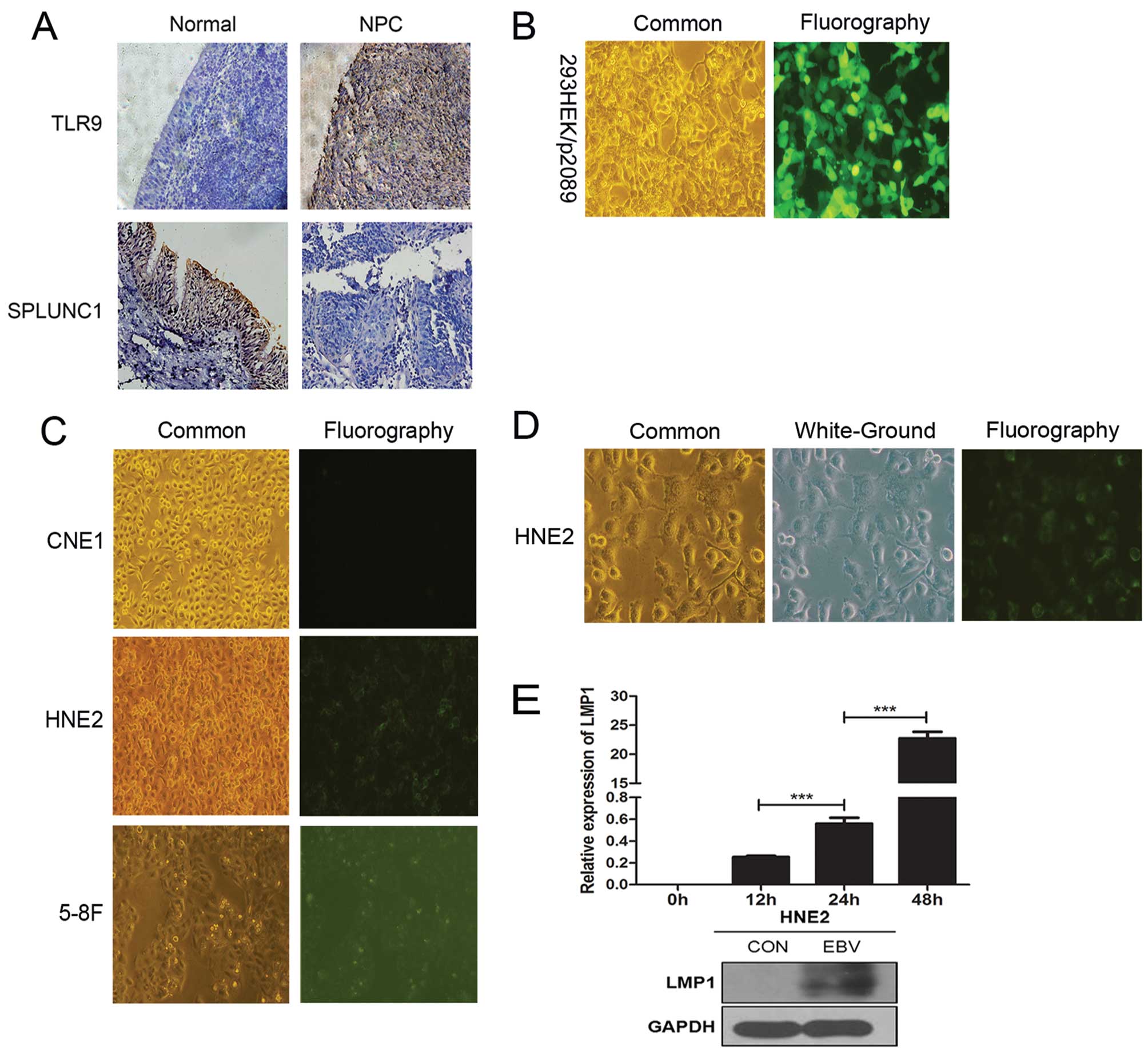 | Figure 1Tissue immunohistochemical analysis
(magnification, ×200) and establishment of the HNE2 cell line model
of high-efficient transfer infection with EBV by ‘cell-to-cell’
method. (A) SPLUNC1 protein expression was weak or absent in the
NPC epithelial samples, while it was positive in the normal
nasopharyngeal epithelial tissues. TLR9 protein expression was
positive in the NPC epithelial samples, while it was weak or absent
in the normal nasopharyngeal epithelial tissues. (B) Fluorescence
intensity of 293HEK/p2089 cells stably transfected with the BAC-EBV
plasmid (p2089) which carries GFP (both, magnification, ×100). (C)
After a 48-h GFP-EBV infection, the appearance of the fluorescence
intensity of the different cell lines is shown (magnification,
×100). The poorly differentiated HNE2 NPC cell line showed the
strongest fluorescence intensity, the undifferentiated
nasopharyngeal carcinoma 5-8F cell line was second, and the highly
differentiated CNE1 NPC cell line showed weakest fluorescence
intensity. (D) Common, white-ground and fluorography presentation
of the HNE2 cell line. (E) LMP1 mRNA expression in the HNE2 cells
was gradually increased at different time-points of 0, 12, 24 and
48 h after EBV infection. After a 48-h EBV infection, the protein
expression of LMP1 in the HNE2 cell group was higher than that in
the control group. Data represent means ± SEM and are
representative of three independent experiments;
***P<0.001 vs. adjacent time period. NPC,
nasopharyngeal carcinoma; SPLUNC1, short palate, lung and nasal
epithelium clone 1; EBV, Epstein-Barr virus; TLR, Toll-like
receptor. |
EBV infection of HNE2 cells affects the
TLR9/NF-κB signaling pathway
TLR3 is the natural ligand of EBV dsRNA. After a
48-h infection, the expression of TLR3 mRNA in the HNE2 cell group
was higher than that in the uninfected group (P<0.001, Fig. 2A). Expression of both TLR9 mRNA and
SPLUNC1 mRNA in the HNE2 cells was gradually increased at different
time-points of 0, 12, 24 and 48 h after EBV infection (adjacent
group, P<0.05). After a 48-h infection, protein expression of
TLR9 and SPLUNC1 in the HNE2 cell group was higher than that in the
uninfected group (Fig. 2B). After a
48-h infection, the protein expression levels of TLR9/NF-κB
signaling pathway-associated cytoines (TLR9, CD14, MyD88, IKK,
P-IKα, IKβα, P-NF-κB and NF-κB) in the HNE2 cell group were higher
than these levels in the uninfected group (Fig. 2C). Meanwhile, after a 48-h
infection, mRNA expression levels of inflammatory cytokines IL-6,
IL-8, IL-1β and TNF-α in the HNE2 cell group were higher than these
levels in the uninfected group (P<0.05, Fig. 2D). These results revealed that EBV
infection of NPC cells can activate the TLR9/NF-κB signaling
pathway, promote the release of inflammatory cytokines and
consequently enhance the inflammatory response.
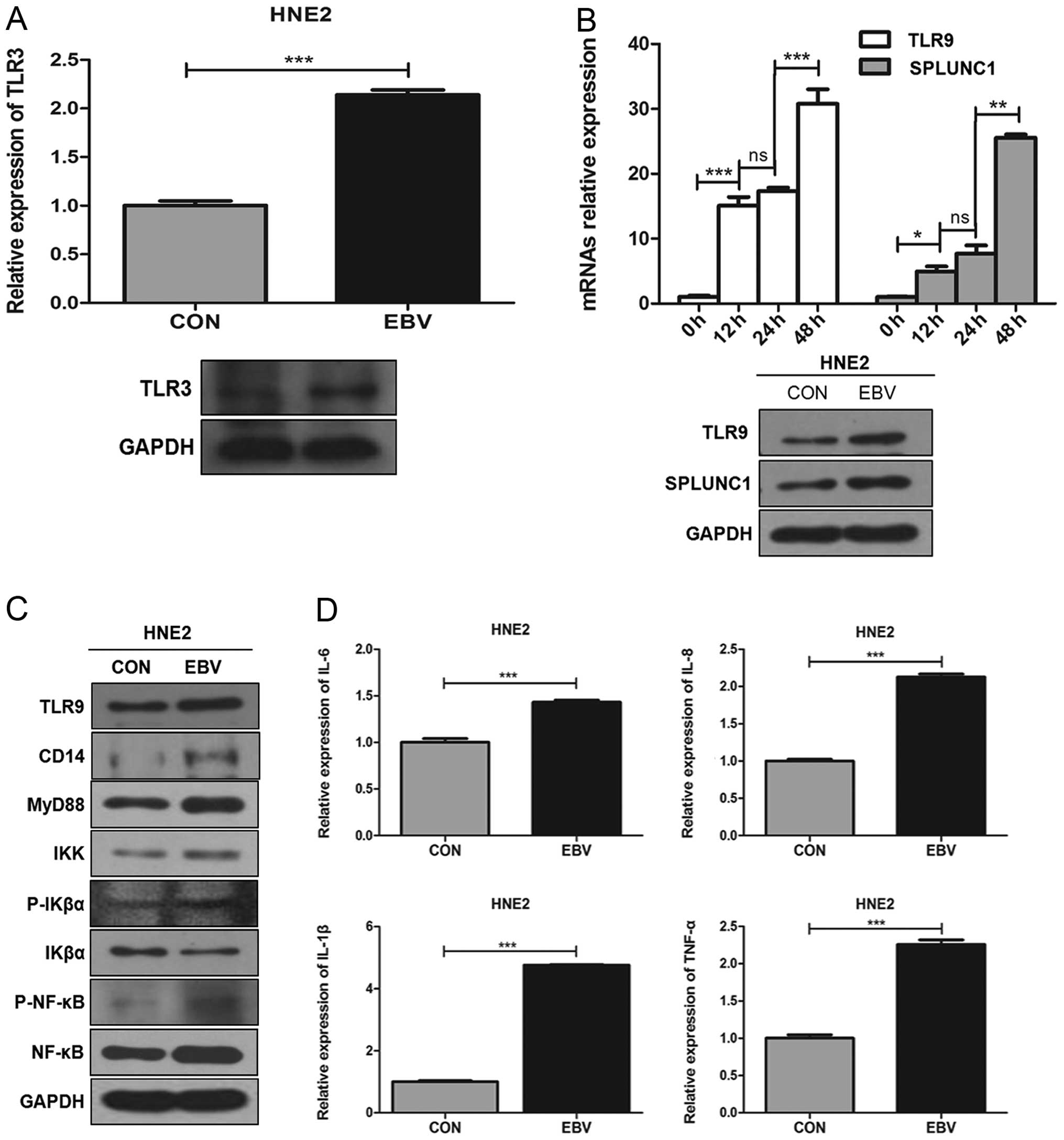 | Figure 2The effect of EBV infection of HNE2
cells on the TLR9/NF-κB signaling pathway. (A) After a 48-h
infection, the expression of TLR3 mRNA in the HNE2 cell group was
higher than that in the uninfected group. ***P<0.001
vs. control. (B) Expression of TLR9 mRNA and SPLUNC1 mRNA was
gradually increased in the HNE2 cells at different time-points of
0, 12, 24 and 48 h after EBV infection. After a 48-h infection, the
protein expression levels of TLR9 and SPLUNC1 were higher in the
HNE2 cell group than these levels in the uninfected group.
***P<0.001, **P<0.01,
*P<0.05 vs. adjacent time-points; ns, not
significant. (C) After a 48-h infection, the protein expression
levels of TLR9/NF-κB signaling pathway-associated cytokines (TLR9,
CD14, MyD88, IKK, P-IKα, IKβα, P-NF-κB and NF-κB) were higher in
the HNE2 cell group than these levels in the uninfected group. (D)
After a 48-h infection, mRNA expression levels of inflammatory
cytokines IL-6, IL-8, IL-1β and TNF-α were higher in the HNE2 cell
group than these levels in the uninfected group.
***P<0.001 vs. control. Data represent means ± SEM
and are representative of three independent experiments. SPLUNC1,
short palate, lung and nasal epithelium clone 1; EBV, Epstein-Barr
virus; TLR, Toll-like receptor. |
SPLUNC1 regulatory effect on the
inflammatory reaction induced by EBV transferred infection of HNE2
cells
After a 48-h infection, the protein expression of
not only SPLUNC1 but also all TLR9/NF-κB signaling
pathway-associated cytokines in the HNE2/SPLUNC1 cell group
(transient transfection) were higher than these levels in the
HNE2/Vector cell group (Fig. 3A).
SPLUNC1 mRNA and protein expression levels in the HNE2/SPLUNC1 cell
group (stable transfection) were significantly higher than these
levels in the HNE2/Vector cell group. In addition, TLR9 mRNA and
protein expression levels in the HNE2/SPLUNC1 cell group (stable
transfection) were lower than these levels in the HNE2/Vector cell
group (Fig. 3B). After a 48-h
infection, the protein expression levels of all TLR9/NF-κB
signaling pathway-associated cytokines but also SPLUNC1 were higher
in the HNE2/SPLUNC1 cell group than these levels in the uninfected
cell group. On the contrary IKβα protein expression had a
contrasting trend (Fig. 3C). After
a 48-h infection, the protein expression levels of all TLR9/NF-κB
signaling pathway-associated cytokines were lower in the
HNE2/SPLUNC1 cell group than these levels in the HNE2/Vector cell
group; the protein expression of IKβα showed a contrasting trend
(Fig. 3D). The mRNA expression
levels of inflammatory cytokines IL-6, IL-8, IL-1β and TNF-α after
a 48-h infection were significantly higher in the HNE2 cell group
than these levels in the uninfected cell group. Meanwhile, the mRNA
expression levels of these inflammatory cytokines in the
HNE2/SPLUNC1 cell group were lower than these levels in the
HNE2/Vector cell group (P<0.05, Fig.
3E).
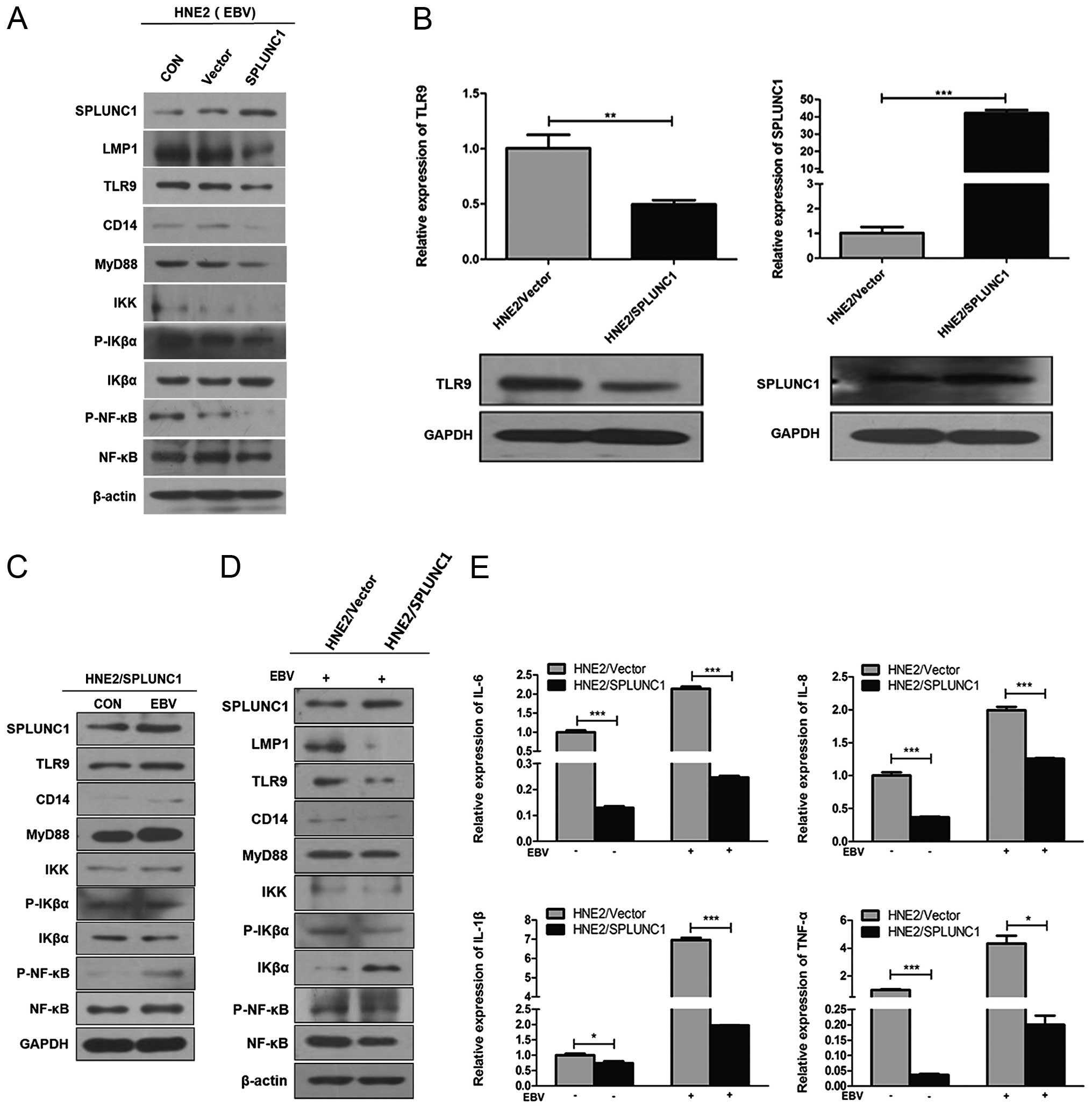 | Figure 3The regulatory effect of SPLUNC1 on
the inflammatory reaction induced by EBV transferred infection of
HNE2 cells. (A) After a 48-h infection, the protein expression
levels of SPLUNC1 and all TLR9/NF-κB signaling pathway-associated
cytokines were higher in the HNE2/SPLUNC1 cell group (transient
transfection) than these levels in the HNE2/Vector cell group. (B)
SPLUNC1 mRNA and protein expression levels were significantly
higher in the HNE2/SPLUNC1 cell group (stable transfection) than
these levels in the HNE2/Vector cell group. TLR9 mRNA and protein
expression levels were lower in the HNE2/SPLUNC1 cell group (stable
transfection) than these levels in the HNE2/Vector cell group.
***P<0.001, **P<0.01 vs. the
HNE2/Vector cell group. (C) After a 48-h infection, the protein
expression of all TLR9/NF-κB signaling pathway-associated cytokines
but also SPLUNC1 were higher in the HNE2/NASG cell group than these
levels in the uninfected cell group. IKβα protein expression had a
contrasting trend. (D) After a 48-h infection, the protein
expression levels of all TLR9/NF-κB signaling pathway-associated
cytokines were lower in the HNE2/SPLUNC1 cell group than these
levels in the HNE2/vector cell group. IKβα protein expression had a
contrasting trend. (E) After 48-h infection, the mRNA expression
levels of inflammatory cytokines IL-6, IL-8, IL-1β and TNF-α were
significantly higher in the HNE2 cell group than these levels in
the uninfected cell group, while the mRNA expression levels of
these inflammatory cytokines were lower in the HNE2/SPLUNC1 cell
group than these levels in the HNE2/Vector cell group.
***P<0.001, *P<0.05. Data represent
means ± SEM and are representative of three independent
experiments. SPLUNC1, short palate, lung and nasal epithelium clone
1; EBV, Epstein-Barr virus; TLR, Toll-like receptor. |
SPLUNC1 regulatory effect on the
inflammatory reaction induced by EBV-DNA direct infection of HNE2
cells
SPLUNC1 mRNA expression in the HNE2 cell group was
lower than that in the NP69 cell group (P<0.05), while TLR9 mRNA
expression was slightly higher than that in the NP69 cell group
(P>0.05); After a direct 48-h transfection of EBV-DNA, TLR9 mRNA
expression in both the HNE2 and NP69 cell groups was increased, but
it was increased more obviously in the HNE2 cell group (Fig. 4A). After a direct 48-h transfection,
the mRNA expression levels of inflammatory cytokines IL-6, IL-8,
IL-1β and TNF-α in the HNE2 cell group were increased more
significantly than these levels in the NP69 cell group (Fig. 4B). After a direct 48-h transfection
in both groups, TLR9 mRNA expression in the HNE2/SPLUNC1 cell group
was not significantly changed (P>0.05), while its mRNA
expression in the HNE2/Vector cell group was significantly
increased (P<0.05, Fig. 4C).
After a direct 48-h transfection in both groups, the mRNA
expression levels of inflammatory cytokines IL-6, IL-8, IL-1β and
TNF-α were increased less significantly in the HNE2/SPLUNC1 cell
group than the expression levels in the HNE2/Vector cell group
(Fig. 4D).
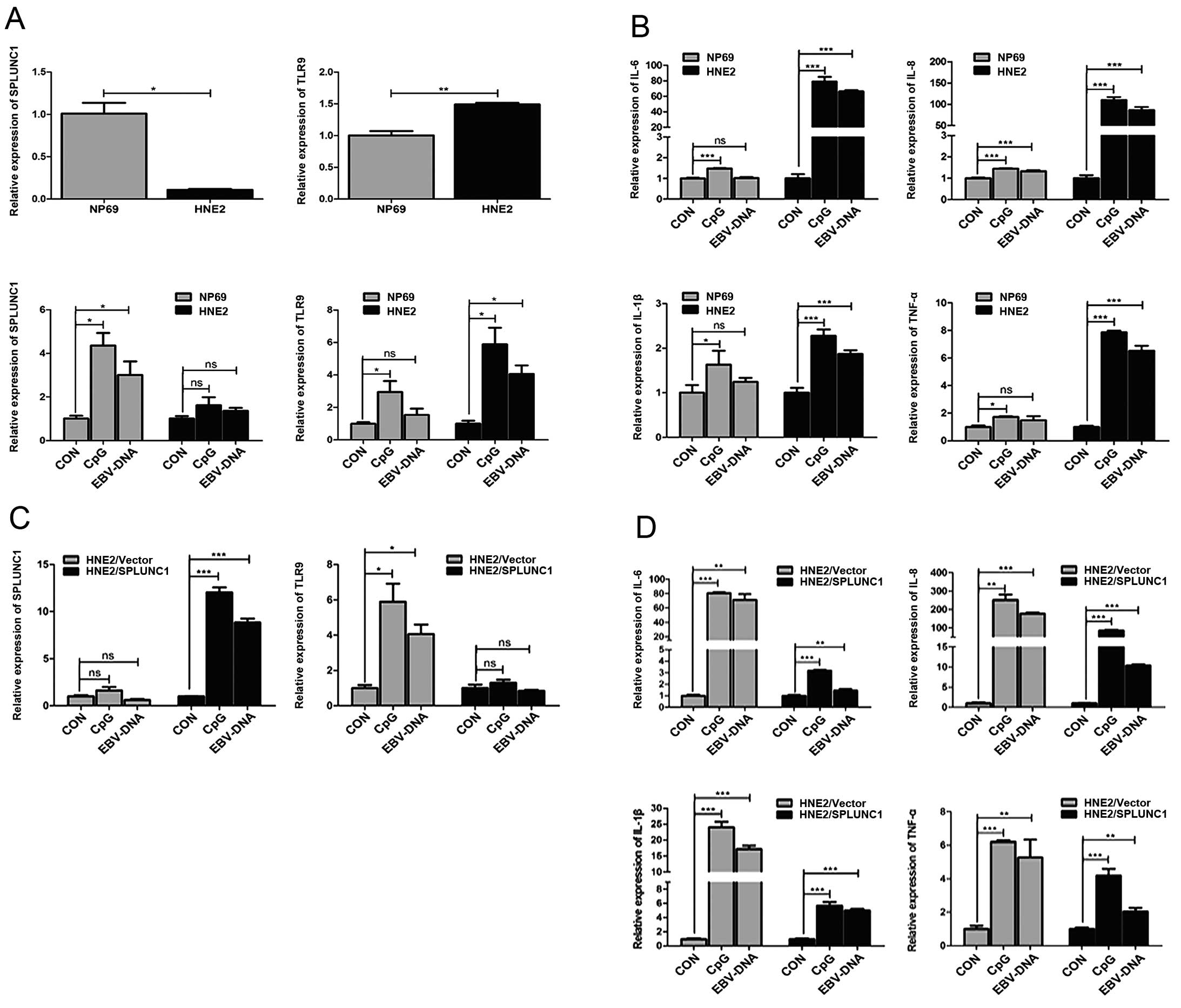 | Figure 4Regulatory effect of SPLUNC1 on the
inflammatory reaction induced by EBV-DNA direct infection of HNE2
cells. (A) SPLUNC1 mRNA expression was lower in the HNE2 cell group
than that in the NP69 cell group. TLR9 mRNA expression was slightly
higher in the HNE2 cell group than that in the NP69 cell group.
After a 48-h EBV-DNA direct transfection, TLR9 mRNA expression in
the HNE2 and NP69 cell groups was increased, but it was increased
more obviously in the HNE2 cell group. **P<0.01,
*P<0.05. ns, not significant. (B) After a 48-h direct
transfection, the mRNA expression levels of inflammatory cytokines
IL-6, IL-8, IL-1β and TNF-α were increased more significantly in
the HNE2 cell group than these levels in the NP69 cell group.
***P<0.001, *P<0.05. ns, not
significant. (C) After a 48-h direct transfection in both groups,
TLR9 mRNA expression in the HNE2/SPLUNC1 cell group was not
significantly changed, while its mRNA expression in the HNE2/Vector
cell group was significantly increased. ***P<0.001,
*P<0.05. ns, not significant. (D) After a 48-h direct
transfection in both groups, the mRNA expression levels of
inflammatory cytokines IL-6, IL-8, IL-1β and TNF-α were increased
less significantly in the HNE2/SPLUNC1 cell group than these levels
in HNE2/Vector cell group. ***P<0.001,
**P<0.01. Data represent means ± SEM and are
representative of three independent experiments. SPLUNC1, short
palate, lung and nasal epithelium clone 1; EBV, Epstein-Barr virus;
TLR, Toll-like receptor. |
Discussion
The present study was the first to screen the NPC
cell line with susceptibility to EBV and demonstrate that a poorly
differentiated HNE2 cell line was more susceptible to EBV than the
other cell lines. The results were consistent with a study by Wang
et al (19) who found that
the vast majority of poorly differentiated or undifferentiated NPC
samples exhibited EBV-DNA infection. One reason may be that poorly
differentiated NPC cell lines have a more rapid proliferative
ability with the status of cell division activity, which causes EBV
to be embedded more easily into the DNA of NPC cells through a
series of processes and leads to EBV infection phenomenon in these
cells.
A sign of effective EBV infection is that EBV-DNA
transfers into the host cells and promotes genome replication and
transcription (20). In addition,
EBV infection may occur in the nasopharyngeal inflammatory stage
and result in enhanced membrane permeability of nasopharyngeal
epithelial cells. In the present study, after HNE2 cells were
infected, the natural ligand TLR9 with the combination ability of
EBV non-methylated nucleotide sequence was activated, its
expression quantity increased, the levels of cytokines of the
TLR9/NF-κB pathway were upregulated and expression levels of
inflammatory factors IL-6, IL-8, IL-1β and TNF-α were increased. We
believe that when EBV infects HNE2 cells, a series of cascade
reactions in molecules such as CD14 and MyD88 occurs, and cytokine
IKβα, an NF-κB inhibitory factor, is phosphorylated by the IKK
kinase complex, which relieves the inhibition of the NF-κB pathway.
Consequently, the TLR9/NF-κB signaling pathway is activated and
inflammatory cytokines IL-6, IL-8, IL-1β and TNF-α are upregulated.
Studies (21–23) have reported that cytokine NF-κB is a
key node protein of the inflammatory response and resistance to
apoptosis and its high expression and overactivity are one of the
main characteristics of inflammation-associated cancer. Moreover,
cytokine NF-κB transfers into the nucleus after activation, leading
to the induction of the secretion of numerous inflammatory
cytokines such as IL-6, IL-8, IL-1β and TNF-α. Furthermore, the
immunosuppression induced by accumulation of these inflammatory
factors greatly increases the risk of inflammation-associated
carcinogenesis (24,25).
TLRs are a family of receptors that play a critical
role in mounting an immune response against microbial pathogens.
TLRs are the pathogen-associated molecular pattern recognition
receptors linking innate and specific immunity (26–29).
The interaction of these motifs with the TLRs triggers a cascade of
signaling events which stimulate the products of pro-inflammatory
cytokines and chemokines to respond to the invading pathogen. As a
member of the TLR family, TLR9 expression is noted in many
malignancies, such as lung and cervical cancer and NPC (30–33).
TLR9 activation can induce high expression of cytokines and
chemokines, cause tumor-inflammatory microenvironment formation and
promote the occurrence and development of malignancies (34,35).
Baumann et al (36) found
that the CD14 molecule plays an essential role in induction of the
inflammatory response following TLR9/NF-κB pathway activation. In
the present study, in the EBV-infected HNE2 cells, the expression
of TLR9/NF-κB pathway-associated factors was significantly
upregulated. Meanwhile, IKβα molecule expression was downregulated.
Inflammatory cytokine expression levels of IL-6, IL-8, IL-1β and
TNF-α were correspondingly and highly increased. The phenomenon
suggests that the TLR9/NF-κB pathway plays an important role in the
mediated inflammatory reaction induced by the EBV infection of HNE2
cells.
SPLUNC1 belonging to innate immune molecules, is a
secreted protein with specific expression in the respiratory mucosa
and is one of the key proteins involved in the host defense in the
respiratory tract. Therefore, SPLUNC1 plays a significant role in
maintaining normal physiological activities of the upper
respiratory tract, inflammatory inhibition, sterilization and tumor
suppression. Our previous studies found that SPLUNC1 removes and
inhibits gram-negative bacteria proliferation (37), and also plays an important role in
resisting EBV infection (38,39).
By assessing the levels in clinical specimens, we observed that
SPLUNC1 expression in nasopharyngeal epithelial tissues was weaker
than that in normal nasopharyngeal epithelial tissues, while TLR9
expression exhibited a contrasting trend. The phenomenon was found
mostly consistent with the two expression in corresponding cells.
Meanwhile, the expression of TLR9 and SPLUNC1 exhibited an
increasing trend following EBV infection in the HNE2 cells. We
considered that the increase in SPLUNC1 in the tumor-inflammatory
microenvironment was not sufficient enough to inhibit the
inflammatory responses mediated by TLR9. In addition, SPLUNC1 had
low expression in HNE2 cells. This phenomenon was found not only in
EBV cell-to-cell transferred infection of HNE2 cells, but also in
EBV-DNA direct transfection of HNE2 cells. Therefore, after EBV-DNA
direct transfer, inflammatory cytokine expression of TLR 9, IL-6,
IL-8, IL-1β and TNF-α was weaker in the NP69 cells with higher
SPLUNC1 expression than these levels in the HNE2 cells with lower
SPLUNC1 expression. The results suggest that SPLUNC1 had a
protective effect on the natural resistance to inflammatory
reaction in the tumor-inflammatory microenvironment induced by EBV
infection and both SPLUNC1 and TLR9 are involved in the
tumorigenesis of NPC.
Zhou et al (38) demonstrated that after SPLUNC1
treatment, defects appeared in EBV particle capsid; EBV-infected
lymphocytes were interfered by SPLUNC1 and demonstrated apoptosis.
SPLUNC1 promoted the cell lysis and apoptosis of B95-8 cells which
can produce EBV. Chen et al (39) showed that SPLUNC1 inhibits EBV from
transferring into peripheral blood lymphocyte which were pretreated
with SPLUNC1 protein, and reduced EBV-related gene expression of
EBER, BZLF1 and LMP1 after EBV integration into cells. In the
present study, we found after EBV infection, inflammatory cytokine
expression levels of IL-6, IL-8, IL-1β and TNF-α were significantly
lower in the HNE2/SPLUNC1 cell group than those in the HNE2/Vector
cell group. The results suggest that SPLUNC1 participates in
regulating the inflammatory reaction induced by EBV infection of
HNE2 cells. Several studies have indicated that SPLUNC1 exhibited a
host defense function through interaction with TLRs. Chu et
al (40) reported that airway
epithelial TLR2 signaling was pivotal in mycoplasma-induced SPLUNC1
production. When the TLR2 signaling pathway was treated with RNAi
of Pam3CSK4 which is a TLR-2 ligand, SPLUNC1 expression in normal
human bronchial epithelial cells was obviously decreased.
Thaikoottathil and Chu (41) found
that TLR2-mediated MAPK/AP-1 activation upregulated lung epithelial
SPLUNC1 expression at the transcriptional level. However, to date,
there are rare reports concerning the interaction of SPLUNC1 and
TLR9 in the inflammatory response induced by EBV infection of NPC.
In the present study, the TLR9 level was significantly lower in the
HNE2/SPLUNC1 cell group than that in the HNE2/Vector cell group.
Obviously, this indicates that TLR9 plays a certain role in
regulating the inflammatory response of SPLUNC1.
In the present study, we found that exogenous
SPLUNC1 molecules were transiently transfected to HNE2 cells which
had been infected with EBV for 48 h. They could obviously promote
NF-κB inhibitor IKβα, reduce TLR9/NF-κB pathway-associated cytokine
expression (TLR9, CD14, MyD88, IKK, P-IKβα, P-NF-κB and NF-κB)
which was induced by EBV infection of HNE2 cells, and consequently
increase inflammatory cytokine expression of IL-6, IL-8, IL-1β and
TNF-α. Meanwhile, we investigated endogenous SPLUNC1 regulatory
effect on EBV infection in HNE2/SPLUNC1 stably transfected cell
lines. The results showed that the expression levels of the
TLR9/NF-κB pathway-associated cytokines were greatly decreased
after EBV infection of HNE2/SPLUNC1 cells, and on the contrary the
expression of NF-κB inhibitor IKβα had a conflicting pattern, which
was similar to transient transfection above. Regardless of the
transfection type, the inflammation cytokine expression levels of
IL-6, IL-8, IL-1β and TNF-α in the HNE2/SPLUNC1 cell group were
markedly lower than these levels in the HNE2/Vector cell group.
These results suggest that SPLUNC1 plays a significant role in the
regulation of the TLR9/NF-κB signal pathway-mediated inflammatory
response induced by EBV infection in NPC.
In conclusion, in the present study, on the basis of
two established modes of cell infection, we demonstrated that EBV
infection of NPC cells activates the TLR9/NF-κB signaling pathway,
promotes release of inflammatory cytokines and consequently
enhances the inflammatory response; SPLUNC1 has the ability to
weaken this inflammatory response induced by EBV infection of NPC
cells through the regulation of the TLR9/NF-κB signaling pathway,
controlling the tumor-inflammatory microenvironment and playing a
regulatory role in tumorigenesis. Thereby, the regulatory chain of
EBV-SPLUNC1-TLR9/NF-κB inflammation was designed to illustrate the
inflammatory regulation mechanism of NPC tumorigenesis and provide
an innovative theoretical foundation for potential therapeutic
targets for NPC.
Acknowledgments
This study was supported by grants from the National
High Technology Research and Development Program of China (no.
2012AA02A206), the National Natural Science Foundation of China
(nos. 91229122, 81071644, 81101509, 81171934, 81172189, 81171930
and 81272298), the 111 Project (no. 111-2-12) and the Hunan
Province Natural Sciences Foundation of China (no. 10JJ7003). We
thank the GSF-National Research Center for Environment and Health
(Germany) and Professor Wolfgang Hammerschmidt for allowing us to
use the Maxi-EBV system.
References
|
1
|
Gu AD, Zeng MS and Qian CN: The criteria
to confirm the role of Epstein-Barr virus in nasopharyngeal
carcinoma initiation. Int J Mol Sci. 13:13737–13747. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Odumade OA, Hogquist KA and Balfour HH Jr:
Progress and problems in understanding and managing primary
Epstein-Barr virus infections. Clin Microbiol Rev. 24:193–209.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Thompson MP and Kurzrock R: Epstein-Barr
virus and cancer. Clin Cancer Res. 10:803–821. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Teramoto N, Gogolák P, Nagy N, Maeda A,
Kvarnung K, Björkholm T and Klein E: Epstein-Barr virus-infected
B-chronic lymphocyte leukemia cells express the virally encoded
nuclear proteins but they do not enter the cell cycle. J Hum Virol.
3:125–136. 2000.PubMed/NCBI
|
|
5
|
Imai S, Nishikawa J and Takada K:
Cell-to-cell contact as an efficient mode of Epstein-Barr virus
infection of diverse human epithelial cells. J Virol. 72:4371–4378.
1998.PubMed/NCBI
|
|
6
|
Li QX, Young LS, Niedobitek G, Dawson CW,
Birkenbach M, Wang F and Rickinson AB: Epstein-Barr virus infection
and replication in a human epithelial cell system. Nature.
356:347–350. 1992. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhang B, Nie X, Xiao B, Xiang J, Shen S,
Gong J, Zhou M, Zhu S, Zhou J, Qian J, et al: Identification of
tissue-specific genes in nasopharyngeal epithelial tissue and
differentially expressed genes in nasopharyngeal carcinoma by
suppression subtractive hybridization and cDNA microarray. Genes
Chromosomes Cancer. 38:80–90. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bingle CD and Craven CJ: Comparative
analysis of the PLUNC (palate, lung and nasal epithelium clone)
protein families. Biochem Soc Trans. 31:806–809. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jiang D, Persinger R, Wu Q, Gross A and
Chu HW: α1-antitrypsin promotes SPLUNC1-mediated lung defense
against Pseudomonas aeruginosa infection in mice. Respir Res.
14:1222013. View Article : Google Scholar
|
|
10
|
Seshadri S, Lin DC, Rosati M, Carter RG,
Norton JE, Suh L, Kato A, Chandra RK, Harris KE, Chu HW, et al:
Reduced expression of antimicrobial PLUNC proteins in nasal polyp
tissues of patients with chronic rhinosinusitis. Allergy.
67:920–928. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Di YP, Tkach AV, Yanamala N, Stanley S,
Gao S, Shurin MR, Kisin ER, Kagan VE and Shvedova A: Dual acute
proinflam-matory and antifibrotic pulmonary effects of short
palate, lung, and nasal epithelium clone-1 after exposure to carbon
nanotubes. Am J Respir Cell Mol Biol. 49:759–767. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lu JH, Tang YL, Yu HB, Zhou JH, Fu CY,
Zeng X, Yu ZY, Yin HL, Wu MH, Zhang JY, et al: Epstein-Barr virus
facilitates the malignant potential of immortalized epithelial
cells: from latent genome to viral production and maintenance. Lab
Invest. 90:196–209. 2010. View Article : Google Scholar
|
|
13
|
Tsao SW, Wang X, Liu Y, Cheung YC, Feng H,
Zheng Z, Wong N, Yuen PW, Lo AK, Wong YC, et al: Establishment of
two immor-talized nasopharyngeal epithelial cell lines using SV40
large T and HPV16E6/E7 viral oncogenes. Biochim Biophys Acta.
1590:150–158. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhou HD, Fan SQ, Zhao J, Huang DH, Zhou M,
Liu HY, Zeng ZY, Yang YX, Huang H, Li XL, et al: Tissue
distribution of the secretory protein, SPLUNC1, in the human fetus.
Histochem Cell Biol. 125:315–324. 2006. View Article : Google Scholar
|
|
15
|
Tang YL, Lu JH, Cao L, Wu MH, Peng SP,
Zhou HD, Huang C, Yang YX, Zhou YH, Chen Q, et al: Genetic
variations of EBV-LMP1 from nasopharyngeal carcinoma biopsies:
Potential loss of T cell epitopes. Braz J Med Biol Res. 41:110–116.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lu J, Tang Y, Zhou M, Wu M, Ouyang J, Gao
J, Zhang L, Li D, Chen Q, Xiong W, et al: Gene modification in the
genome of Epstein-Barr virus cloned as a bacterial artificial
chromosome. Wei Sheng Wu Xue Bao. 48:385–390. 2008.In Chinese.
PubMed/NCBI
|
|
17
|
Chen L, Yin J, Chen Y and Zhong J:
Induction of Epstein-Barr virus lytic replication by recombinant
adenoviruses expressing the zebra gene with EBV specific promoters.
Acta Biochim Biophys Sin (Shanghai). 37:215–220. 2005. View Article : Google Scholar
|
|
18
|
Yu H, Lu J, Zuo L, Yan Q, Yu Z, Li X,
Huang J, Zhao L, Tang H, Luo Z, et al: Epstein-Barr virus
downregulates microRNA 203 through the oncoprotein latent membrane
protein 1: a contribution to increased tumor incidence in
epithelial cells. J Virol. 86:3088–3099. 2012. View Article : Google Scholar :
|
|
19
|
Wang WY, Chien YC, Jan JS, Chueh CM and
Lin JC: Consistent sequence variation of Epstein-Barr virus nuclear
antigen 1 in primary tumor and peripheral blood cells of patients
with naso-pharyngeal carcinoma. Clin Cancer Res. 8:2586–2590.
2002.PubMed/NCBI
|
|
20
|
Omerović J, Lev L and Longnecker R: The
amino terminus of Epstein-Barr virus glycoprotein gH is important
for fusion with epithelial and B cells. J Virol. 79:12408–12415.
2005. View Article : Google Scholar
|
|
21
|
Karin M: Nuclear factor-kappaB in cancer
development and progression. Nature. 441:431–436. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cavallo F, De Giovanni C, Nanni P, Forni G
and Lollini PL: 2011: the immune hallmarks of cancer. Cancer
Immunol Immunother. 60:319–326. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Oeckinghaus A, Hayden MS and Ghosh S:
Crosstalk in NF-κB signaling pathways. Nat Immunol. 12:695–708.
2011. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Mantovani A, Allavena P, Sica A and
Balkwill F: Cancer-related inflammation. Nature. 454:436–444. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Colotta F, Allavena P, Sica A, Garlanda C
and Mantovani A: Cancer-related inflammation, the seventh hallmark
of cancer: links to genetic instability. Carcinogenesis.
30:1073–1081. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Takeuchi O and Akira S: Pattern
recognition receptors and inflammation. Cell. 140:805–820. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Iwasaki A and Medzhitov R: Regulation of
adaptive immunity by the innate immune system. Science.
327:291–295. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Basith S, Manavalan B, Yoo TH, Kim SG and
Choi S: Roles of toll-like receptors in cancer: a double-edged
sword for defense and offense. Arch Pharm Res. 35:1297–1316. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Amirchaghmaghi E, Taghavi SA, Shapouri F,
Saeidi S, Rezaei A and Aflatoonian R: The role of toll like
receptors in pregnancy. Int J Fertil Steril. 7:147–154. 2013.
|
|
30
|
Droemann D, Albrecht D, Gerdes J, Ulmer
AJ, Branscheid D, Vollmer E, Dalhoff K, Zabel P and Goldmann T:
Human lung cancer cells express functionally active Toll-like
receptor 9. Respir Res. 6:12005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lee JW, Choi JJ, Seo ES, Kim MJ, Kim WY,
Choi CH, Kim TJ, Kim BG, Song SY and Bae DS: Increased toll-like
receptor 9 expression in cervical neoplasia. Mol Carcinog.
46:941–947. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Berger R, Fiegl H, Goebel G, Obexer P,
Ausserlechner M, Doppler W, Hauser-Kronberger C, Reitsamer R, Egle
D, Reimer D, et al: Toll-like receptor 9 expression in breast and
ovarian cancer is associated with poorly differentiated tumors.
Cancer Sci. 101:1059–1066. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ronkainen H, Hirvikoski P, Kauppila S,
Vuopala KS, Paavonen TK, Selander KS and Vaarala MH: Absent
Toll-like receptor-9 expression predicts poor prognosis in renal
cell carcinoma. J Exp Clin Cancer Res. 30:842011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Woods DC, White YA, Dau C and Johnson AL:
TLR4 activates NF-κB in human ovarian granulosa tumor cells.
Biochem Biophys Res Commun. 409:675–680. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ben Abdelwahed R, Cosette J, Donnou S,
Crozet L, Ouakrim H, Fridman WH, Sautès-Fridman C, Mahjoub A and
Fisson S: Lymphoma B-cell responsiveness to CpG-DNA depends on the
tumor microenvironment. J Exp Clin Cancer Res. 32:182013.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Baumann CL, Aspalter IM, Sharif O,
Pichlmair A, Blüml S, Grebien F, Bruckner M, Pasierbek P, Aumayr K,
Planyavsky M, et al: CD14 is a coreceptor of Toll-like receptors 7
and 9. J Exp Med. 207:2689–2701. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhou HD, Li GY, Yang YX, Li XL, Sheng SR,
Zhang WL and Zhao J: Intracellular co-localization of SPLUNC1
protein with nanobacteria in nasopharyngeal carcinoma epithelia
HNE1 cells depended on the bactericidal permeability increasing
protein domain. Mol Immunol. 43:1864–1871. 2006. View Article : Google Scholar
|
|
38
|
Zhou HD, Li XL, Li GY, Zhou M, Liu HY,
Yang YX, Deng T, Ma J and Sheng SR: Effect of SPLUNC1 protein on
the Pseudomonas aeruginosa and Epstein-Barr virus. Mol Cell
Biochem. 309:191–197. 2008. View Article : Google Scholar
|
|
39
|
Chen P, Guo X, Zhou H, Zhang W, Zeng Z,
Liao Q, Li X, Xiang B, Yang J, Ma J, et al: SPLUNC1 regulates cell
progression and apoptosis through the miR-141-PTEN/p27 pathway, but
is hindered by LMP1. PLoS One. 8:e569292013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chu HW, Gally F, Thaikoottathil J,
Janssen-Heininger YM, Wu Q, Zhang G, Reisdorph N, Case S, Minor M,
Smith S, et al: SPLUNC1 regulation in airway epithelial cells: role
of Toll-like receptor 2 signaling. Respir Res. 11:1552010.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Thaikoottathil J and Chu HW: MAPK/AP-1
activation mediates TLR2 agonist-induced SPLUNC1 expression in
human lung epithelial cells. Mol Immunol. 49:415–422. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zheng Y, Qin Z, Ye Q, Chen P, Wang Z, Yan
Q, Luo Z, Liu X, Zhou Y, Xiong W, et al: Lactoferrin suppresses the
Epstein-Barr virus-induced inflammatory response by interfering
with pattern recognition of TLR2 and TLR9. Lab Invest.
94:1188–1199. 2014. View Article : Google Scholar : PubMed/NCBI
|














