Introduction
Osteosarcoma, the most common primary bone
malignancy, is most prevalent in children and young adults
(1). Due to its high propensity for
local invasion, early metastasis and recurrence, osteosarcoma leads
to a high mortality rate. It has been documented that the 3-year
disease-free survival of osteo sarcoma patients remains 60–70%
despite combined therapy including surgery, chemotherapy and
radiotherapy (2). The molecular
mechanism underlying osteosarcoma development and metastasis has
yet to be fully elucidated, which has a negative impact on the
improvement of diagnosis and treatment. Therefore, an improved
understanding of the molecular biology and carcinogenic mechanism
underlying the development and progression of osteosarcoma may be
useful in the treatment of this malignancy.
The forkhead box (FOX) superfamily comprises a
number of transcription factors which share a highly conserved
DNA-binding domain (forkhead/winged helix-box) that plays a role in
the regulation of various processes, including metabolism, cell
proliferation and gene expression during ontogenesis (3). The role of FOX proteins in various
types of cancer has been comprehensively investigated considering
that these proteins have vital functions on biological processes.
Certain Fox members including FOXP1, FOXM1, FOXR1 and FOXA1, show a
diversity of activities on tumorigenesis and progression of cancer
(4–8), while having dual roles in cancer
biology. For instance, on the one hand, FOXP1 overexpression by
chromosome translocations is associated with poor prognosis in
lymphomas, suggesting it acts as an oncogene (4). On the other hand, loss of FOXP1
expression is associated with poor prognosis in breast cancer,
suggesting FOXP1 also acts as a tumor suppressor in other types of
cancer (5). As tumor suppressors or
oncogenes, Fox proteins may function as direct or indirect targets
of therapeutic intervention.
The present study focused on FOXL1 which belongs to
the FOXL subfamily. As a novel identified tumor suppressor, the
re-expression or overexpression of FOXL1 inhibits growth and
induces apoptosis in colonic, pancreatic and gallbladder cancer
cells (9–11). However, the role of FOXL1 in
osteosarcoma remains to be investigated. In the present study, the
association of FOXL1 expression with prognosis of osteosarcoma
patients was assessed. In addition, the biological functions of
FOXL1 in proliferation, cell cycle and apoptosis of osteosarcoma
cells were evaluated in vitro or in vivo.
Materials and methods
Patients and tissue collections
The study was approved by the Ethics Committee of
The First Affiliated Hospital of Bengbu Medical College and written
informed consent was obtained from all the patients. Thirty-seven
of the patients with osteosarcoma enrolled in the present study
were treated with radical resection (without prior radiotherapy or
chemotherapy) between 2007 and 2009 at the Department of
Orthopedics, The First Affiliated Hospital of Bengbu Medical
College. Specimens of tumor tissues (n=37) and adjacent non-tumor
tissues (n=18) were collected immediately following surgery, fixed
with formalin and then embedded with paraffin. The specimens were
diagnosed by two pathologists. Following treatment with the same
chemotherapy regimens, a follow-up was performed for all the
patients.
Immunohistochemistry
Serial sections (5-µm) were cut from
formalin-fixed, paraffin-embedded tumor or non-tumor blocks. The
sections were dewaxed in xylene and rehydrated with concentration
gradients. Heat-induced antigen retrieval was performed in a
conventional pressure cooker, followed by elimination of endogenous
peroxidase activity with H2O2. To decrease
non-specific binding and background staining, the sections were
incubated with 10% goat serum in phosphate-buffered saline (PBS).
The sections were then incubated with primary antibodies against
FOXL1 (1:100 dilution) overnight at 4°C and then incubated with a
secondary antibody (1:500 dilution) (both from Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA) at room temperature for 2
h. The horseradish peroxidase-conjugated antibody was detected and
visualized by 3,3′-diaminobenzidine tetrachloride (DAB).
Immunostained sections were blindly reviewed by two independent
pathologists. The staining intensity was scored using a 3-scale
system (0, − or negative; 1, + or weak; 2, ++ or moderate; and 3,
+++ or strong) and osteosarcoma patients were classified into two
groups according to the scores of FOXL1 immunostaining: low
expression (− or +) and high expression (++ or +++).
Cell lines and culture conditions
Osteosarcoma cell lines U-2 OS, MG-63 and Saos-2
were all purchased from the Cell Bank of the Chinese Academy of
Sciences, Shanghai, China. All cell lines were cultured in
Dulbecco’s modified Eagle’s medium (DMEM; Invitrogen, Carlsbad, CA,
USA) supplemented with 10% (v/v) fetal bovine serum (FBS; HyClone,
Logan, UT, USA) and 1% penicillin/streptomycin (Invitrogen) at 37°C
in a humidified 5% CO2 incubator.
Transfections
The pcDNA-FOXL1 carrying full-length of FOXL1 cDNA
and the pcDNA3.1 empty vector were generous gifts from Dr Qin
(Department of General Surgery, Xinhua Hospital, Affiliated to
School of Medicine, Shanghai Jiaotong University, Shanghai, China).
pcDNA-FOXL1 was transfected into osteosarcoma cells to restore or
upregulate the FOXL1 expression. Transfected cells were cultured
with medium supplemented with geneticin (G418, 200–300
µg/ml) for 10–14 days to generate stable transfectants. The
expression of FOXL1 in transfected cells was examined by RT-PCR and
western blot analysis.
RT-PCR assay
Reverse transcription-polymerase chain reaction
(RT-PCR) was used to examine the FOXL1 mRNA expression in Saos-2,
MG-63 and U-2 OS cells. Total RNA was extracted from the cell lines
using a TRIzol reagent kit (Gibco-BRL, Gaithersburg, MD, USA)
according to the manufacturer’s instructions. After quantification,
a certain amount of RNAs was reverse transcribed into cDNA using a
PrimeScript™ RT-PCR kit (Takara, Bio, Shiga, Japan). The obtained
cDNA was subjected to PCR using the following procedure: 1 cycle of
5 min at 94°C, then 30 cycles of 45 sec at 94°C, of 45 sec at 59°C
and of 30 sec at 72°C. The primer sequences for FOXL1 were designed
as described by Qin et al (11). The GAPDH expression was used as an
endogenous control.
Cell viability
Cell viability was measured using the
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay. Briefly, 5×103 of transfected cells were
collected and cultured in each well of 96-well plates for 24, 48,
72 or 96 h. The cells were then incubated with 20 µl of MTT
solutions (5 mg/ml) for 4 h at 37°C. The supernatant was removed
and crystals were dissolved in dimethylsulfoxide (DMSO). The
absorbance of each well was measured at 570 nm with the use of an
automated microplate reader. The cell viability was calculated
using the formula: (OD of treated groups/OD of control group)
×100%.
Xenograft tumor experiments
To assess the effects of FOXL1 expression on cell
proliferation in vivo, a xenograft model in nude mice was
established using U-2 OS cells. Twenty-four six-week-old specific
pathogen-free female athymic (nu/nu) nude mice were purchased from
the Shanghai Laboratory Animal Center of the Chinese Academy of
Sciences. The animals were kept in cages and provided with clean
water and food. The U-2 OS cells that lacked the FOXL1 expression
were stably transfected with pcDNA-FOXL1 or pcDNA3.1 and subjected
to the established xenograft model. Briefly, 1×106 of
U-2 OS cells were injected into the right flanks of the nude mice.
When xenograft tumors were visible and the volume reached ~0.5
cm3, tumor volumes were measured every three days and
calculated using the formula: volume = length × width2 ×
π/6. The mice were sacrificed after two weeks of observation and
the tumor weight was measured. The animal experiments were approved
by the Ethics Committee of The First Affiliated Hospital of Bengbu
Medical College and efforts were made to minimize suffering.
Cell-cycle analysis
The distribution of the cell cycle was examined and
analyzed using flow cytometry. Following transfection with
pcDNA-FOXL1, the U-2 OS cells were collected and washed with cold
PBS, and fixed with cold 70% ethanol overnight at 4°C. The fixed
cells were then washed twice with cold PBS and centrifuged at 500 g
for 5 min. The cells were incubated with 10 µg/ml RNase A
and 50 µg/ml PI at 37°C for 30 min in the dark. The cells
were then analyzed by flow cytometry and the cell cycle was
analyzed using FlowJo software.
Flow cytometric analysis of
apoptosis
Annexin V-FITC staining was performed to detect
apoptosis of the U-2 OS cells. Briefly, the transfected cells were
collected at a specific time point (24, 48 and 72 h), washed with
PBS and then stained with Annexin V-FITC and PI at room temperature
for 15 min. The percentage of apoptotic cells was then analyzed by
flow cytometry.
Flow cytometric analysis of the
mitochondrial membrane potential
The fluorescent probe JC-1
(5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethyl-benzimidazolo-carbocyanine
iodide) was used to measure the depolarization of the mitochondrial
membrane potential in the U-2 OS cells. Briefly, the transfected
cells were seeded in 6-well plates and were harvested at 24, 48 and
72 h. Approximately 1×106 cells were incubated with 1
µl of JC-1 (2 mg/ml) at 37°C for 20 min. The cells were then
washed with PBS twice, followed by FACS analysis (Becton-Dickinson,
Franklin Lakes, NJ, USA).
Western blot analysis
The expression of cell cycle- and apoptosis-related
proteins was examined using western blot analysis. Briefly, 48 h
after transfection, total proteins were extracted from the U-2 OS
cells. The protein (30 µg) was subjected to 10–12% sodium
dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and
electrotransferred to polyvinylidene difluoride (PVDF) membranes
(Millipore, Billerica, MA, USA). The membranes were incubated in
blocking buffer and probed with appropriate primary antibodies
(Santa Cruz Biotechnology, Inc.) overnight at 4°C, followed by
incubation with horseradish peroxidase-conjugated secondary
antibodies (Santa Cruz Biotechnology, Inc.). The blots were
incubated with an enhanced chemiluminescence solution (ECL) kit
(Pierce, Rockford, IL, USA) and exposed to X-OMAT Blue film (Kodak,
Rochester, NY, USA). Cytochrome c oxidase subunit IV (Cox
IV) and β-actin were used as controls for mitochondrial and
cytosolic proteins, respectively.
Statistical analysis
Statistical analysis was performed with SPSS 17.0
(SPSS, Inc., Chicago, IL, USA). The Student’s t-test, one-way
ANOVA, Kaplan-Meier, log-rank and Chi-square tests were used to
analyze the differences between the groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
FOXL1 expression is downregulated in the
osteosarcoma tissues and cell lines
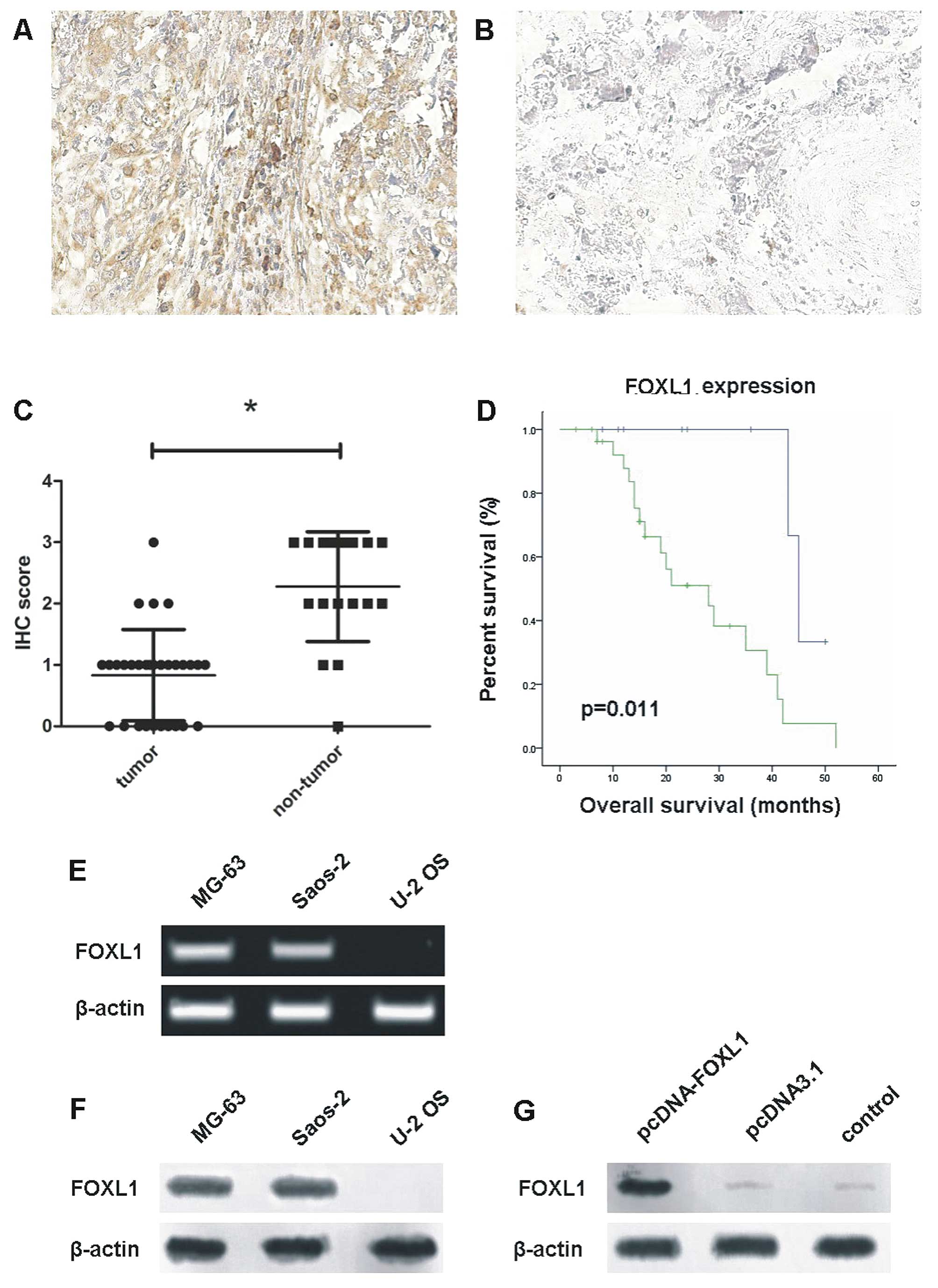
Immunohistochemical staining revealed that FOXL1
expression was downregulated in the osteosarcoma tissues. Brown in
the nucleus (FOXL1 staining) indicated that the FOXL1 protein was
located in the nucleus of normal and cancer cells (Fig. 1A and B). The staining scores of
FOXL1 expression in osteosarcoma cases mainly ranged between 0 and
1, while those in the corresponding normal tissues mainly range
from 2 to 3. The mean score of FOXL1 expression in osteosarcoma
tissues was reduced as compared to that in the normal tissues
(P<0.05, Fig. 1C). In addition,
the patients with a high expression of FOXL1 had a longer survival
rate, than those with a low expression of FOXL1 (P<0.05,
Fig. 1D).
RT-PCR and western blot analysis revealed that the
MG-63 and Saos-2 cell lines had endogenous FOXL1 mRNA and protein,
while the U-2 OS cell line lacked endogenous FOXL1 mRNA and protein
(Fig. 1E and F). Thus, the U-2 OS
cell line was selected for subsequent studies. Additionally, FOXL1
expression in the U-2 OS cell line was restored by transfection
with pcDNA-FOXL1 (Fig. 1G).
Restoration of FOXL1 expression inhibits
U-2 OS cell growth in vitro and in vivo
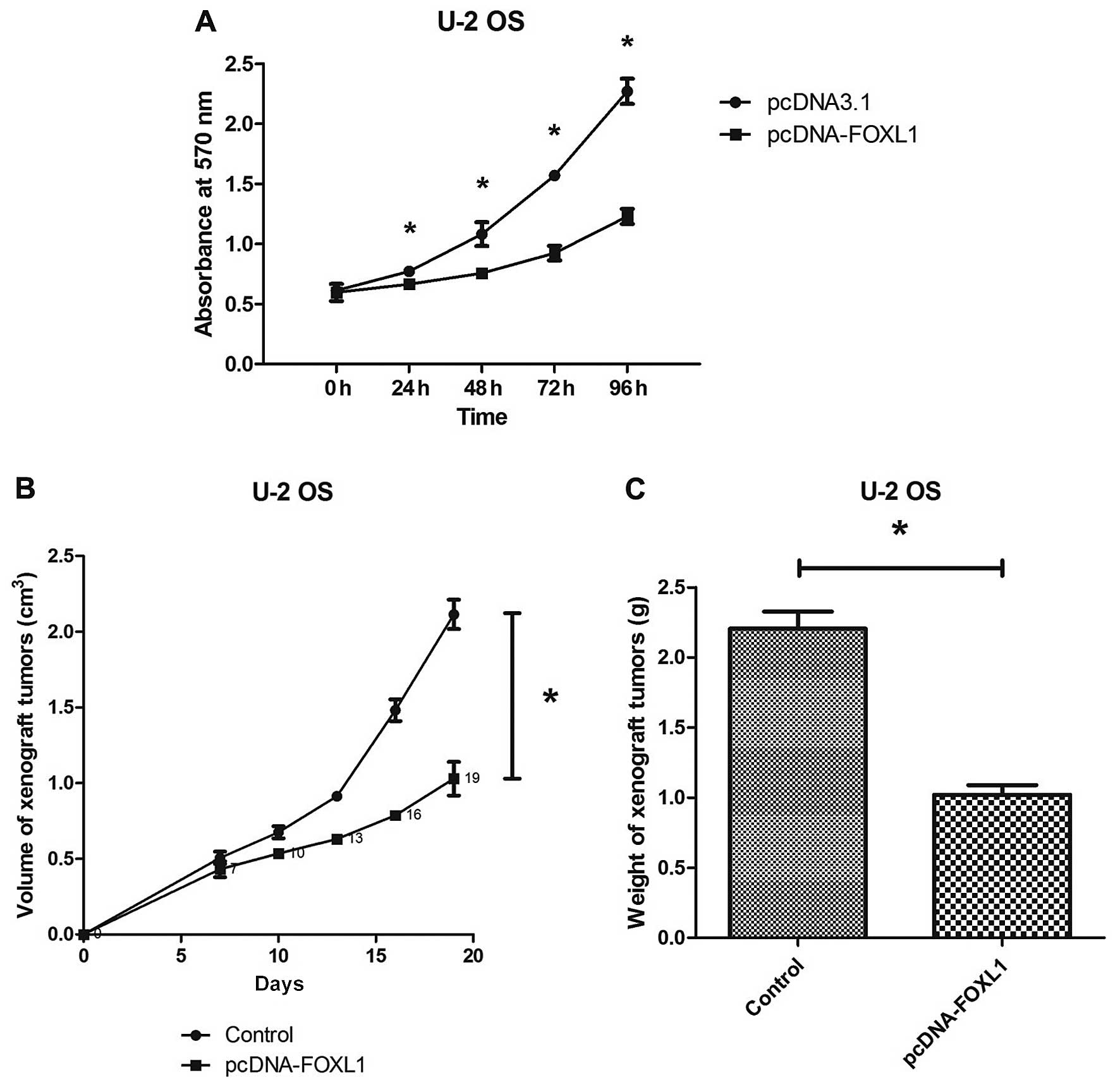
The MTT assay showed that transfection with
pcDNA-FOXL1 significantly inhibited the U-2 OS cell growth in
vitro and this growth inhibition was time-dependent (Fig. 2A). The in vivo inhibitory
effect of FOXL1 against the U-2 OS cells was evaluated by a
xenograft tumor model. It was observed that the ectopic FOXL1
expression significantly inhibited the growth of xenograft tumors
on nude mice. The volume and weight of the FOXL1-overexpressing
xenograft tumors were much less than those of the control xenograft
tumors (Fig. 2B and C). The in
vitro and in vivo studies demonstrated that restoration
of the FOXL1 expression inhibited the proliferation of the U-2 OS
cells, which lacked the endogenous FOXL1 expression.
Restoration of the FOXL1 expression
induces G1 arrest in the U-2 OS cell line
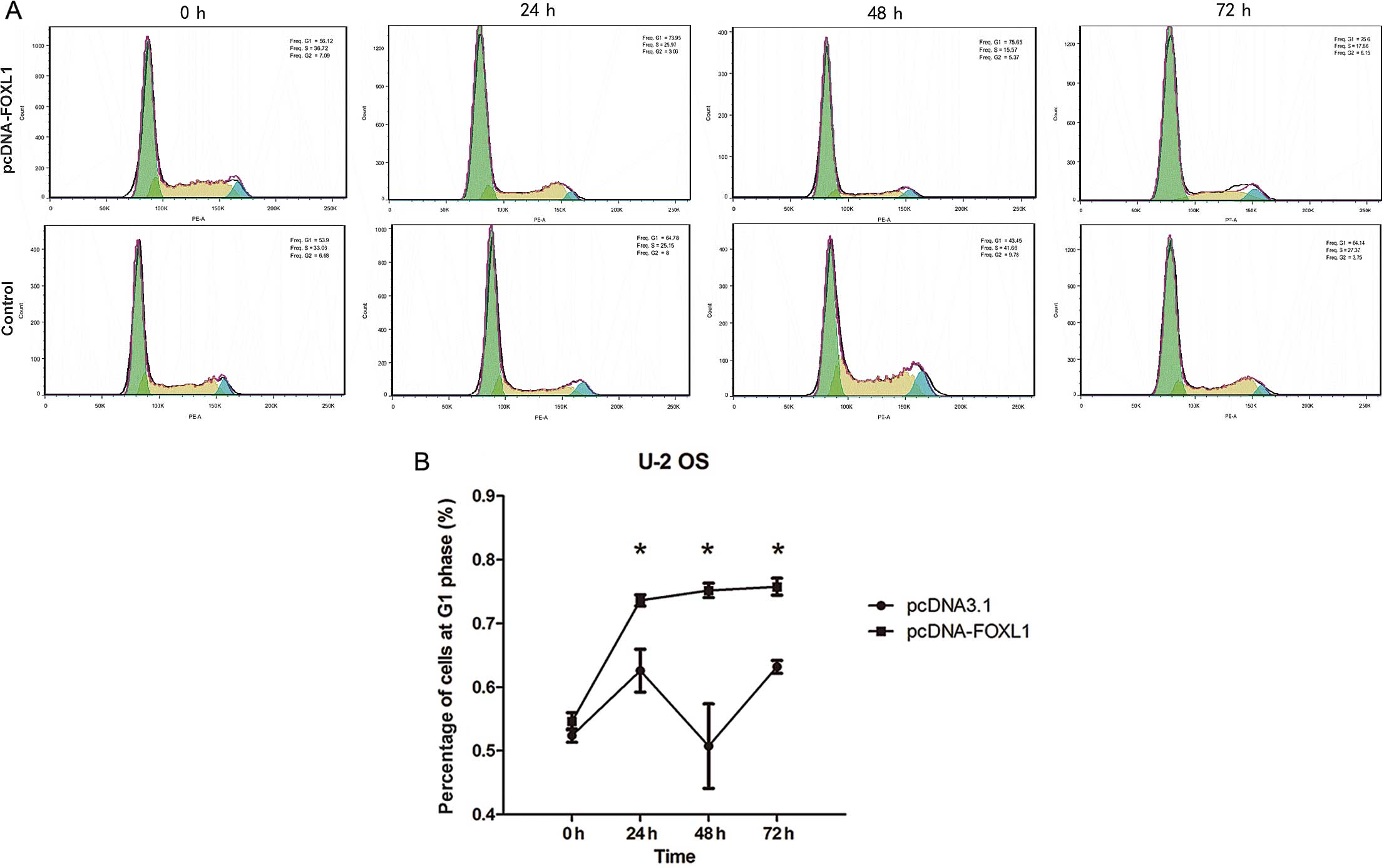
The distribution of the cell cycle was analyzed by
FACS. It was noted that the proportion of the U-2 OS cells in G1
phase was significantly increased following transfection with
pcDNA-FOXL1 (P<0.05 between 24 and 72 h) (Fig. 3).
Restoration of FOXL1 expression induces
apoptosis in the U-2 OS cell line
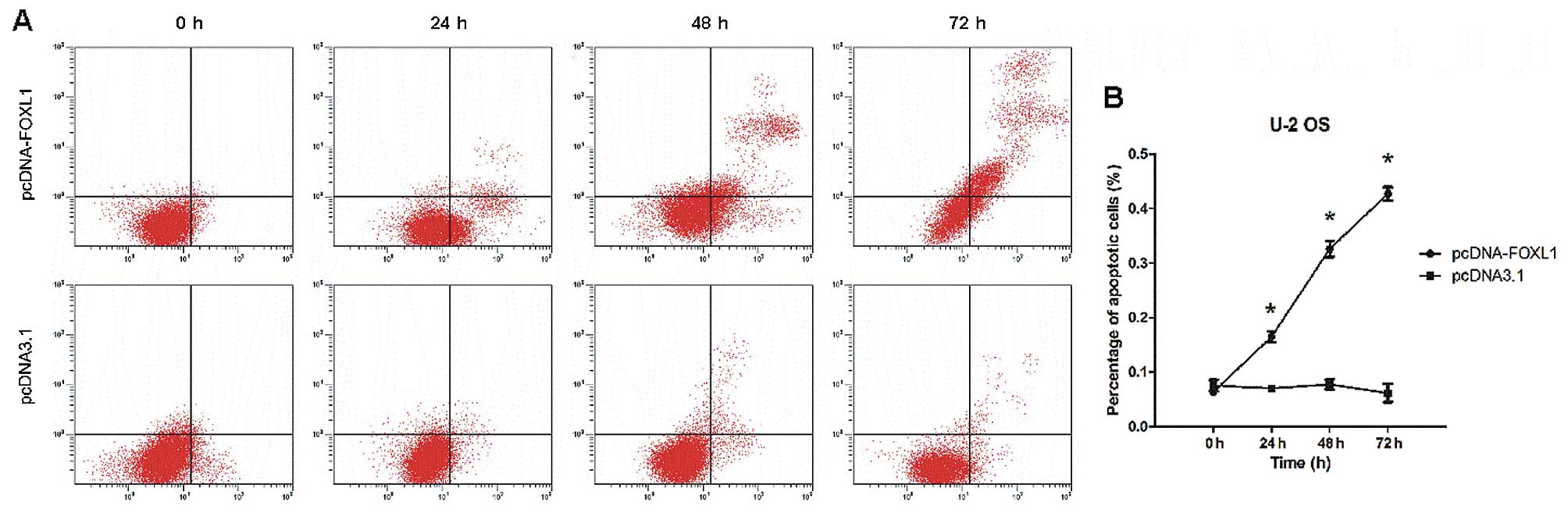
Annexin V/PI staining showed that restoration of
FOXL1 expression induced the apoptosis of U-2 OS cells in
vitro. As shown in Fig. 4A and
B, a significant increase in the proportion of apoptotic U-2 OS
cells (early plus late apoptosis) was observed following
transfection with pcDNA-FOXL1 (P<0.05 24-72 h). The proportion
of late apoptotic or necrotic cells increased in a time-dependent
manner (starting from 24 h and peaking at 72 h).
Restoration of FOXL1 expression
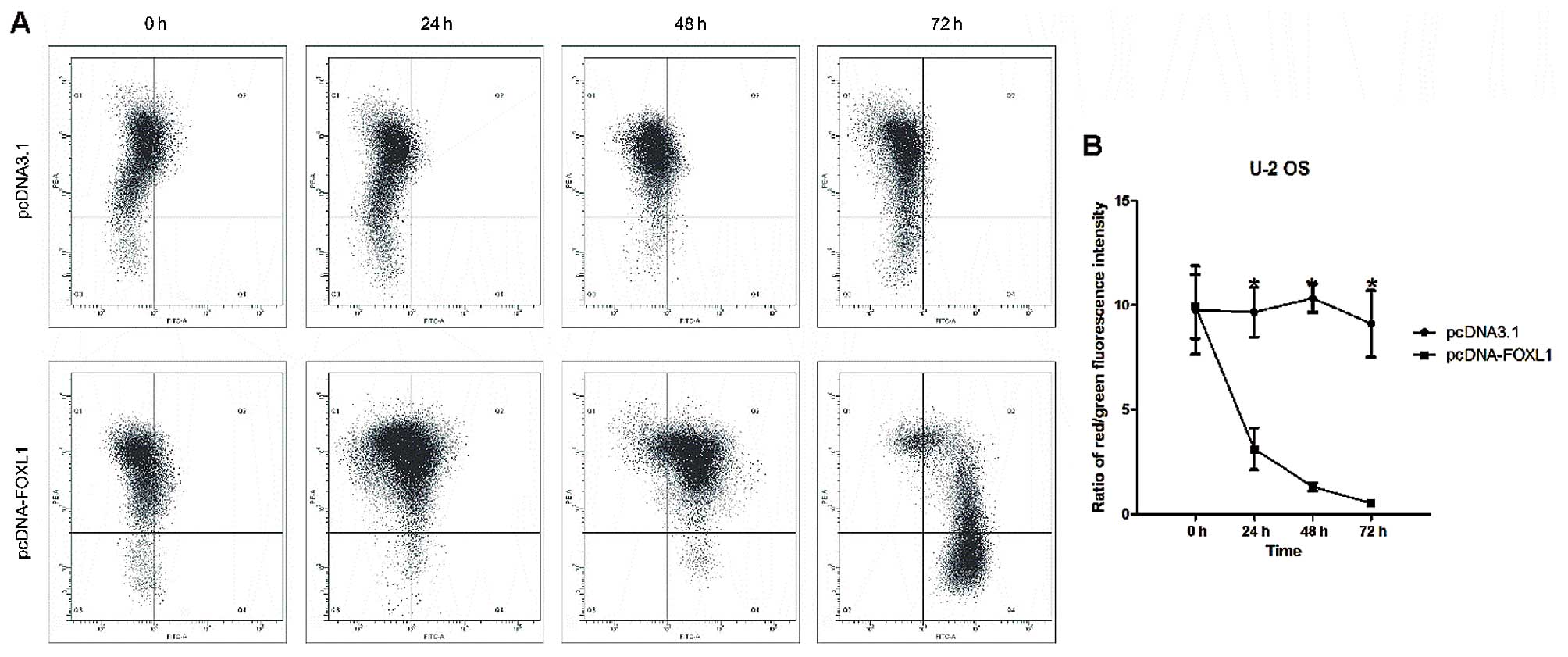
depolarizes transmembrane potential depolarization
Disruption of the mitochondrial inner transmembrane
potential (ΔΨm) is often associated with early apoptosis (20). In the present study, loss of ΔΨm was
determined by the JC-1 staining assay. The shift of JC-1
fluorescence from red to green indicated a collapse of ΔΨ. As shown
in Fig. 5A and B, the percentage of
low ΔΨ cells with JC-1 (green fluorescence-positive) was increased
while that of low ΔΨ cells with JC-1 (red fluorescence-positive)
was decreased, suggesting ectopic FOXL1 expression-induced
disruption of the mitochondrial membrane potential of U-2 OS
cells.
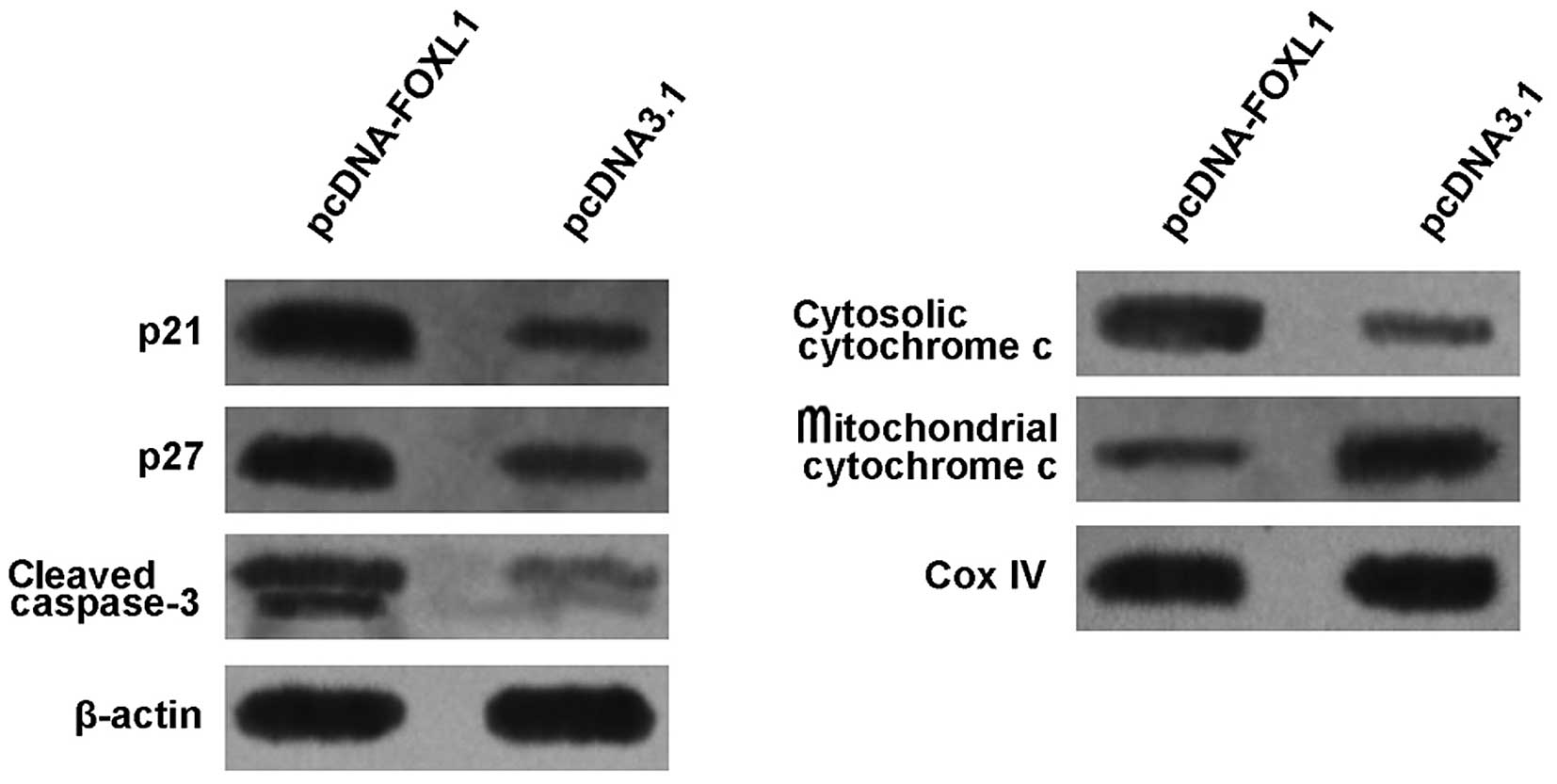
Restoration of the FOXL1 expression
increases the expression of p21, p27, cytosolic cytochrome c and
cleaved caspase-3
To preliminarily study the molecular mechanisms for
the induction of cycle arrest and apoptosis of the U-2 OS cells
following transfection with pcDNA-FOXL1, the expression of p21,
p27, cytochrome c and caspase-3 was examined by the western
blot analysis. As shown in Fig. 6,
the level of cytosolic cytochrome c was markedly increased,
while that of mitochondrial cytochrome c was markedly
decreased 48 h after transfection with pcDNA-FOXL1, suggesting that
FOXL1 promoted cytochrome c release from mitochondria to
cytosol. Furthermore, it was observed that pcDNA-FOXL1 induced an
increase in the expression of p21, p27 and cleaved caspase-3.
Discussion
Gene therapy has been considered as potential
strategy for the treatment of osteosarcoma (12). However, the molecular pathogenesis
of osteosarcoma-regulating growth and progression remains to be
fully elucidated, which may hinder the development of gene therapy.
In the present study, the role of FOXL1 in the regulation of
growth, cell cycle and apoptosis in osteosarcoma cells was
preliminarily investigated.
The levels of FOXL1 mRNA and protein were initially
examined in osteosarcoma tissues and cell lines. It was
demonstrated that the FOXL1 expression was significantly decreased
as compared with corresponding non-tumor tissues. Additionally,
FOXL1 expression was associated with an improved prognosis in
patients undergoing surgery for osteosarcoma. These data indicate
the possibility that FOXL1 plays a tumor-suppressor role in
osteosarcoma. Similar results were observed in other types of
cancer, such as pancreatic and gallbladder cancer (10,11).
The regulation of FOXL1 expression remains to be determined, while
future studies should be focused on genetic and epigenetic
mechanisms for the deregulated FOXL1 expression in tumors.
To identify its role in osteosarcoma, the effects of
pcDNA-FOXL1 on the proliferation of the U2-OS osteosarcoma cell
line lacking the endogenous expression of FOXL1 was subsequently
studied. The ectopic expression of FOXL1 inhibited the
proliferation of the U-2 OS cells as well as the tumorigenicity and
growth of xenograft tumors in vivo. Based on the results of
the cell cycle and apoptosis analysis, it was concluded that the
antitumor activities of pcDNA-FOXL1 may be attributable to its
ability to induce cell cycle arrest and apoptosis.
It has been documented that certain forkhead
transcription factors have been shown to play an intrinsic role in
controlling cell-cycle progression (13,14).
In the present study, it was found that FOXL1 plays a role in
controlling G1/S transition. The G1/S checkpoint controls the entry
of cells from the first preparation phase (G1) into the DNA
synthesis phase (S). A number of cell cycle regulators participate
in controlling this checkpoint. p21CIP1 and
p27Kip1, which belong to cyclin-dependent kinase
inhibitors (CKIs), interact with cyclin-dependent kinases,
including CDK2, to inhibit kinase activity and reduce the
phosphorylation of the retinoblastoma tumor-suppressor protein (Rb)
(15,16). The Rb complex binds to the E2F-DP1
transcription factors, inhibiting the downstream transcription
(17). In the present study,
transfection with pcDNA-FOXL1 caused the accumulation of the
cyclin-dependent kinase inhibitors p21 and p27, which may inhibit
cdk2 kinase activity and the phosphorylation of Rb, leading to G1
arrest.
During apoptosis, cytochrome c is released
into the cytosol as the outer membrane of mitochondria becomes
permeable and the mitochondrial transmembrane potential is
collapsed, and these events trigger caspase activation and
initiation of apoptosis (18). In
the present study, it was observed that ΔΨm was disrupted and
cytochrome c was released from mitochondria to cytosol
following transfection with pcDNA-FOXL1. In addition, caspase-3,
the key executioner in the process of apoptosis (19) was activated. These findings suggest
that the ectopic FOXL1 expression activated the intrinsic
(mitochondrial) apoptotic pathway.
In conclusion, our results suggest that FOXL1 plays
a tumor-suppressor role in osteosarcoma and the restoration of
FOXL1 expression by gene therapy providing a potential treatment
for osteosarcoma patients lacking endogenous FOXL1 expression.
Acknowledgments
The present study was supported by the Natural
Science Foundation of Bengbu Medical College (No. Byky1340). We
thank Dr Qin for his support for the present study.
References
|
1
|
Kim HJ, Chalmers PN and Morris CD:
Pediatric osteogenic sarcoma. Curr Opin Pediatr. 22:61–66. 2010.
View Article : Google Scholar
|
|
2
|
Marina N, Gebhardt M, Teot L and Gorlick
R: Biology and therapeutic advances for pediatric osteosarcoma.
Oncologist. 9:422–441. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Coffer PJ and Burgering BM: Forkhead-box
transcription factors and their role in the immune system. Nat Rev
Immunol. 4:889–899. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hoeller S, Schneider A, Haralambieva E,
Dirnhofer S and Tzankov A: FOXP1 protein overexpression is
associated with inferior outcome in nodal diffuse large B-cell
lymphomas with non-germinal centre phenotype, independent of gains
and structural aberrations at 3p14.1. Histopathology. 57:73–80.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Koon HB, Ippolito GC, Banham AH and Tucker
PW: FOXP1: A potential therapeutic target in cancer. Expert Opin
Ther Targets. 11:955–965. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bergamaschi A, Madak-Erdogan Z, Kim YJ,
Choi YL, Lu H and Katzenellenbogen BS: The forkhead transcription
factor FOXM1 promotes endocrine resistance and invasiveness in
estrogen receptor-positive breast cancer by expansion of stem-like
cancer cells. Breast Cancer Res. 16:4362014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Santo EE, Ebus ME, Koster J, Schulte JH,
Lakeman A, van Sluis P, Vermeulen J, Gisselsson D, Øra I, Lindner
S, et al: Oncogenic activation of FOXR1 by 11q23 intrachromosomal
deletion-fusions in neuroblastoma. Oncogene. 31:1571–1581. 2012.
View Article : Google Scholar
|
|
8
|
Gong C, Fujino K, Monteiro LJ, Gomes AR,
Drost R, Davidson-Smith H, Takeda S, Khoo US, Jonkers J, Sproul D,
et al: FOXA1 repression is associated with loss of BRCA1 and
increased promoter methylation and chromatin silencing in breast
cancer. Oncogene. Dec 22–2014.Epub ahead of print. View Article : Google Scholar
|
|
9
|
Perreault N, Sackett SD, Katz JP, Furth EE
and Kaestner KH: Foxl1 is a mesenchymal Modifier of Min in
carcinogenesis of stomach and colon. Genes Dev. 19:311–315. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang G, He P, Gaedcke J, Ghadimi BM, Ried
T, Yfantis HG, Lee DH, Hanna N, Alexander HR and Hussain SP: FOXL1,
a novel candidate tumor suppressor, inhibits tumor aggressiveness
and predicts outcome in human pancreatic cancer. Cancer Res.
73:5416–5425. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Qin Y, Gong W, Zhang M, Wang J, Tang Z and
Quan Z: Forkhead box L1 is frequently downregulated in gallbladder
cancer and inhibits cell growth through apoptosis induction by
mitochondrial dysfunction. PLoS One. 9:e1020842014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tan ML, Choong PF and Dass CR:
Osteosarcoma: Conventional treatment vs. gene therapy. Cancer Biol
Ther. 8:106–117. 2009. View Article : Google Scholar
|
|
13
|
Furukawa-Hibi Y, Yoshida-Araki K, Ohta T,
Ikeda K and Motoyama N: FOXO forkhead transcription factors induce
G2-M checkpoint in response to oxidative stress. J Biol
Chem. 277:26729–26732. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kazantseva YA, Yarushkin AA and Pustylnyak
VO: CAR-mediated repression of Foxo1 transcriptional activity
regulates the cell cycle inhibitor p21 in mouse livers. Toxicology.
321:73–79. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
LaBaer J, Garrett MD, Stevenson LF,
Slingerland JM, Sandhu C, Chou HS, Fattaey A and Harlow E: New
functional activities for the p21 family of CDK inhibitors. Genes
Dev. 11:847–862. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Møller MB: P27 in cell cycle control and
cancer. Leuk Lymphoma. 39:19–27. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Giacinti C and Giordano A: RB and cell
cycle progression. Oncogene. 25:5220–5227. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jiang X and Wang X: Cytochrome c-mediated
apoptosis. Annu Rev Biochem. 73:87–106. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Porter AG and Jänicke RU: Emerging roles
of caspase-3 in apoptosis. Cell Death Differ. 6:99–104. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ly JD, Grubb DR and Lawen A: The
mitochondrial membrane potential (deltapsi(m)) in apoptosis; an
update. Apoptosis. 8:115–128. 2003. View Article : Google Scholar : PubMed/NCBI
|