Introduction
Hepatocellular carcinoma (HCC) is one of the most
common lethal cancers worldwide (1). Following the diagnosis of liver
cancer, approximately 20% patients benefit from curative surgical
therapies such as liver resection and transplantation. Accumulating
evidence has shown that insulin-like growth factors (IGF-1 and
IGF-2) and their receptors (IGF-1R) are involved in the progression
of cancers (2–5). The interaction between IGF-1 or IGF-2
and IGF-1R plays a pivotal role in tumorigenesis and the
proliferation of cancer cells due to promotion of the cell-cycle
progression (6). The mRNA
expression level of IGF-1 in human HCC tissues is much lower than
that in normal liver tissues, whereas, IGF-2 and IGF-1R reveal the
overexpression in animal models of hepatocarcinogenesis and in HCC
tissues (7). Mounting evidence
confirms that the IGFs/IGF-1R axis is crucial in the development of
HCC (7,8).
Daintain was first isolated from porcine intestine
(9) and allograft inflammatory
factor-1 (AIF-1) was first identified in heterotopic cardiac
allografts of rats (10). Daintain
has a high homology with AIF-1, thus this peptide has been
deisgnated as as daintain/AIF-1. Daintain/AIF-1 can be
constitutively expressed in monocytes and macrophages, and is
involved in macrophage activation (11,12).
It plays a crucial role in many autoimmune diseases (9,13–15).
In previous studies, daintain/AIF-1 was confirmed to promote the
proliferation and migration of breast cancer cells (16,17).
However, whether daintain/AIF-1 has any impact on the progression
of HCC remain unknown.
The association of inflammation with cancer has been
previously suggested (18). A clear
example of inflammation-associated cancer is HCC. Although mounting
evidence is gathered to explore its molecular mechanisms, an
accurate molecular connection between inflammation and HCC remains
elusive. It has been demonstrated that inflammatory factors such as
TNF-α, IL-6 and IL-1α contribute to the progression of HCC
(19,20). Daintain/AIF-1, as a novel
inflammatory factor, has been found to be expressed in activated
Kupffer cells lining the walls of liver sinusoids (21,22).
Therefore, we hypothesized that daintain/AIF-1 may be involved in
the development of HCC. Based on this hypothesis, the effects of
daintain/AIF-1 on the activation of IGF-1R and its downstream
signaling pathway in HepG2 cells were investigated. The results
revealed that daintain/AIF-1 accelerated the activation of
IGF-1-induced IGF-1R and its downstream signaling pathway.
Materials and methods
Reagents and antibodies
3-(4,5-dimethylthiazol-2-yl)-2,5-di-phenylterazolium
bromide (MTT), propidium iodide (PI), Triton X-100 and recombinant
human IGF-1 were all purchased from Sigma Chemical Co. (St. Louis,
MO, USA). Recombinant human daintain/AIF-1 was prepared by our
laboratory according to previous instructions (22). An Annexin V-FITC apoptosis detection
kit was obtained from th eBiyuntian Institute of Biotechnology
(Shanghai, China). Enzyme-linked immunosorbent assay (ELISA) kits
were purchased from R&D Systems (Minneapolis, MN, USA). Primary
antibodies were purchased from Santa Cruz Biotechnology, Inc.
(Santa Cruz, CA, USA).
Cell line and culture
HepG2 cells were purchased from the China Center for
Type Culture Collection (CCTCC) and cultured into Dulbecco’s
modified Eagle’s medium (DMEM; Gibco, Grand Island, NY, USA)
supplemented with 10% (v/v) fetal bovine serum (FBS), 100 U/ml
penicillin and 100 mg/l streptomycin at 37°C in an incubator
supplied with 5% CO2.
Secretion of IGF-1, IGF-2 and
IGFBP-3
HepG2 cells were plated in 6-well plates and
incubated with daintain/AIF-1 at various concentrations in
serum-free medium for 72 h. The cell supernatants were then
collected to examine the secretion of IGF-1, IGF-2 and IGFBP-3
using commercial ELISA kits.
Reverse transcription polymerase chain
reaction (RT-PCR) analysis
Total RNA was isolated using TRIzol reagent
(Invitrogen, Carlsbad, CA, USA) and reversely transcribed to cDNA
using a RevertAid™ cDNA Synthesis kit (Fermentas, Vilnius,
Lithuania) according to the manufacturer’s instructions. The primer
sequences and reaction conditions of IGF-1, IGF-2,
IGFBP-3, IGF-1R and GAPDH are shown in
Table I. The PCR products were
evaluated by 2% agarose gel electrophoresis.
 | Table IPrimer sequences and reaction
conditions of RT-PCR. |
Table I
Primer sequences and reaction
conditions of RT-PCR.
| Genes | Primers | Sequences
(5′-3′) | Annealing
temperature (°C) | Cycle no. |
|---|
| IGF-1 | Forward |
GCATTGTGGATGAGTGTTGC | 53 | 30 |
| Reverse |
GGCTCCTCCTACATTCTGTA | | |
| IGF-2 | Forward |
GAGCTCGAGGCGTTCAGG | 58 | 30 |
| Reverse |
GTCTTGGGTGGGTAGAGCAATC | | |
| IGFBP-3 | Forward |
ATATGGTCCCTGCCGTAGA | 55 | 30 |
| Reverse |
AAATCGAGGCTGAGCCAG | | |
| GAPDH | Forward |
CAAGGTCATCCATGACAACTTTG | 56 | 25 |
| Reverse |
GTCCACCACCCTGTTGCTGTAG | | |
Western blot analysis
Cell samples were lysed in 20 μl cell lysis
buffer containing 1 mM PMSF. The extracted proteins were separated
by 12% SDS-PAGE and transferred to PVDF membrane by 2-h
electroblotting. The blots were blocked in 5% non-fat dry milk for
1 h at room temperature, and then incubated overnight with
corresponding primary antibodies at 4°C. The membrane was washed
with TBS containing 0.05% Tween-20 (TBS-T) three times and
incubated with horseradish peroxidase-conjugated secondary
antibodies for 1 h at room temperature. The blots were then
developed with the enhanced chemiluminescence (ECL) kit (Pierce
Biotechnology, Rockford, IL, USA). The optical density of the
protein bands was assessed using Image J software.
Cell proliferation assays
Cell proliferation was evaluated by an MTT assay.
For the MTT assay, the cells were plated in 96-well plates at a
density of 2.5×103 cells/well and incubated overnight
with standard culture medium. The cells were then starved for 24 h
in serum-free medium. After 24-h cell culture, the original medium
was replaced with fresh serum-free medium containing 40 μg/l
IGF-1 in the presence or absence of daintain/AIF-1. After
incubation for another 24 h, the medium was replaced with 100
μl MTT (0.5 mg/ml) and incubated at 37°C for 4 h. After 4 h,
the MTT solution was removed and 100 μl DMSO was added into
each well. After agitation for 10 min, the absorbance of each well
was measured by a micro-plate reader (Tecan Sunrise, Salzburg,
Austria) at a wavelength of 570 nm.
Cell cycle analysis
In order to investigate the distribution of the cell
cycle, HepG2 cells were seeded in 6-well plates at a density of
1×106cells/well. After adhesion, the cells were starved
for 24 h in serum-free DMEM. The original medium was then replaced
with fresh serum-free medium containing 40 μg/l IGF-1 in the
presence or absence of daintain/AIF-1. After incubation for 24 h,
the cells were collected and analyzed by flow cytometer (FC500;
Beckman Coulter, La Brea, CA, USA).
Statistical analysis
The experiments were repeated at least three times.
The data were presented as mean ± SD and calculated using the
Student’s t-test using GraphPad Prism version 5.0 (GraphPad
Software, San Diego, CA, USA). A statistically significant
difference was considered at P<0.05.
Results
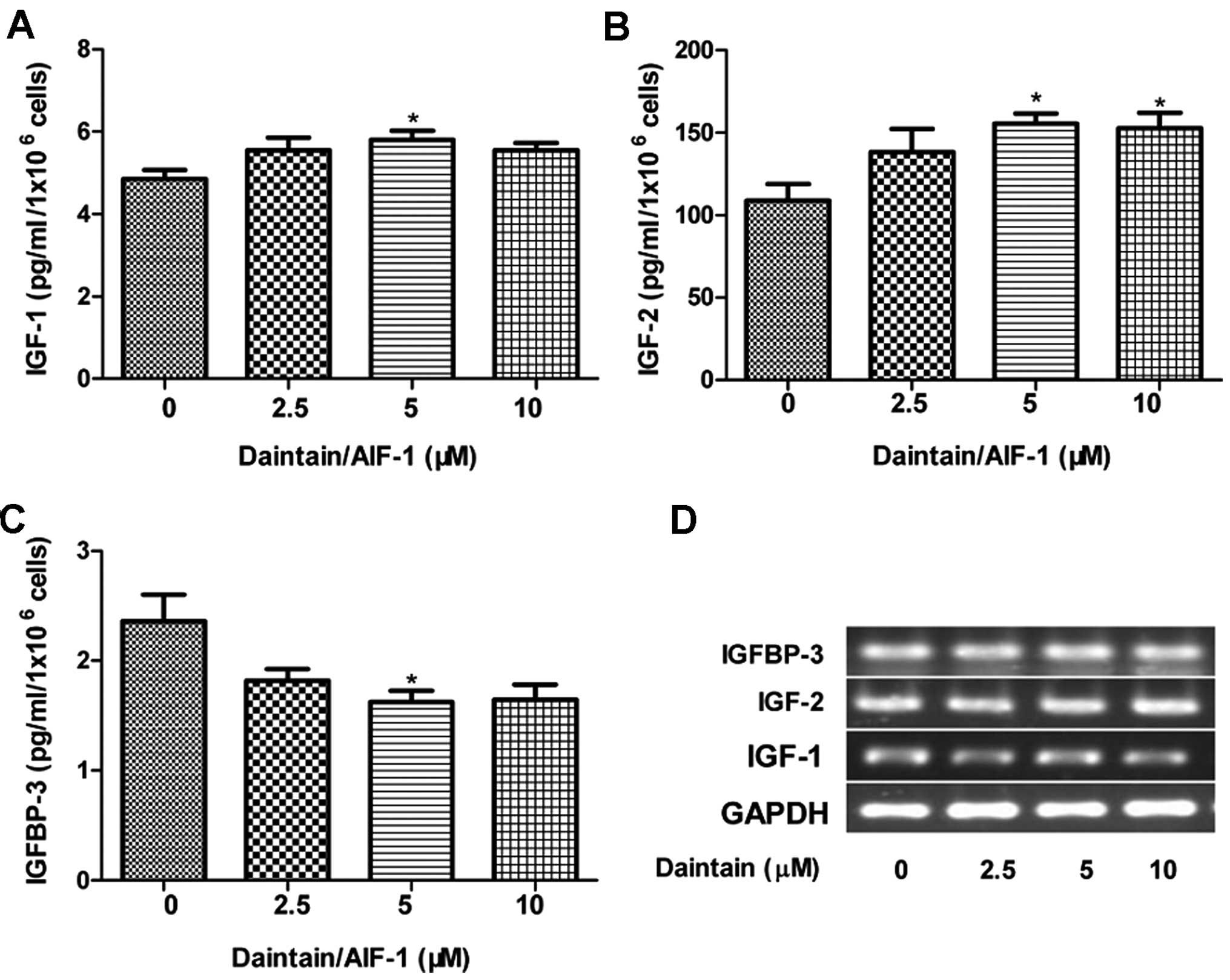
Daintain/AIF-1 promotes the secretion of
IGF-1, IGF-2 and IGFBP-3 in HepG2 cells, but fails to modulate
their gene expression
Accumulating data have demonstrated that alteration
of the autocrine/paracrine loops involving IGFs and IGFBP-3 is
associated with the proliferation of HCC cells (8,23–25).
Therefore, we examined the effects of daintain/AIF-1 on the
production of IGFs and IGFBP-3 in the present study. Enhanced
secretion of IGF-1 and IGF-2 was observed due to the stimulation of
daintain/AIF-1 (Fig. 1A and B). By
contrast, daintain/AIF-1 at the concentration of 5 μM
obviously decreased the secretion of IGFBP-3 (Fig. 1C). However, semi-quantitative RT-PCR
analysis clearly showed that daintain/AIF-1 had no obvious effect
on the gene expression of IGF-1, IGF-2 and IGFBP-3 (Fig. 1D).
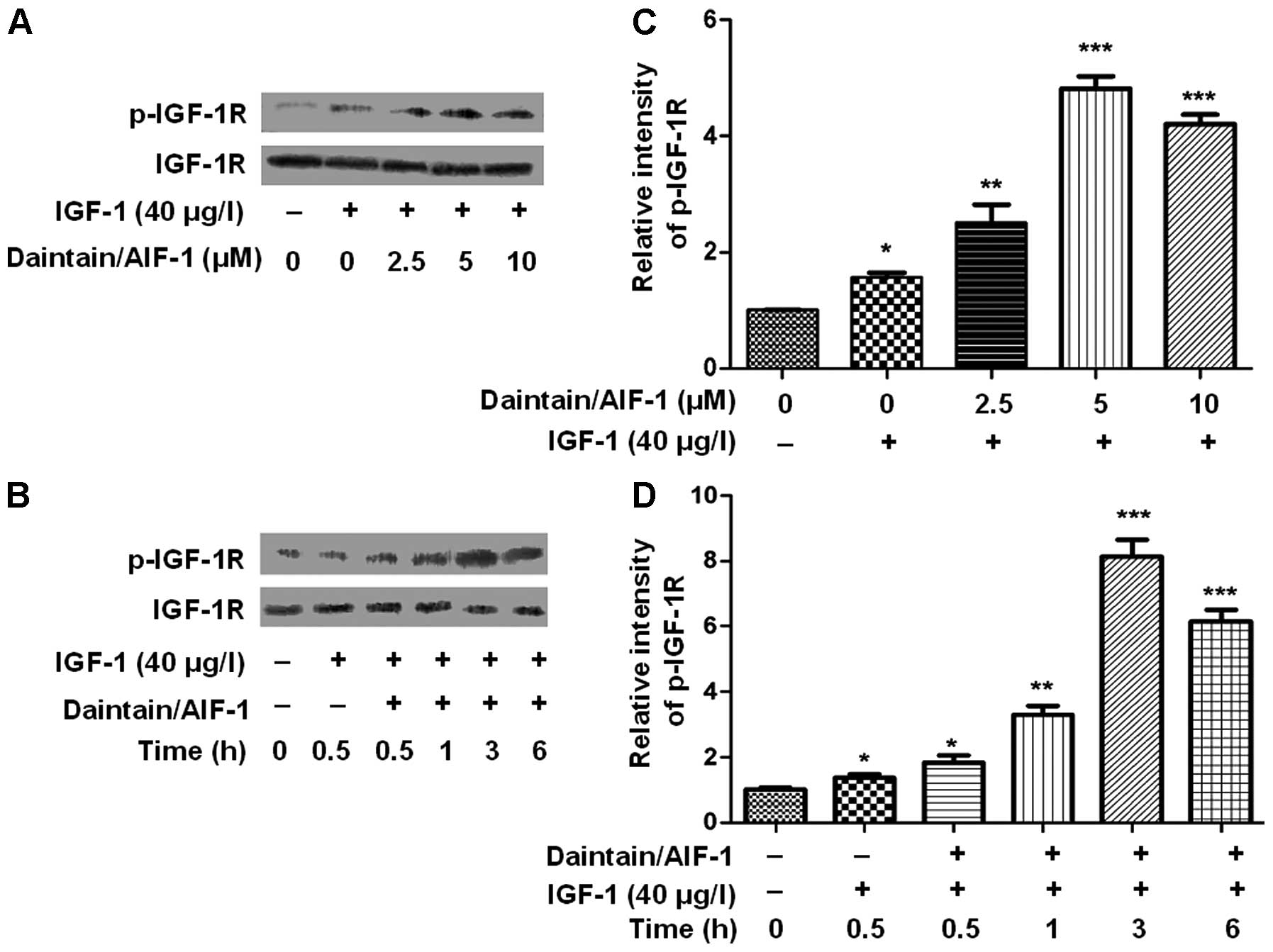
Daintain/AIF-1 accelerates the activation
of IGF-1-induced IGF-1R in HepG2 cells
Western blot analysis was conducted to examine the
effect of daintain/AIF-1 on the IGF-1-induced activation of IGF-1R.
As shown in Fig. 2A and C, IGF-1
induced an obvious phosphorylation of IGF-1R and daintain/AIF-1
accelerated IGF-1-induced activation of IGF-1R in a dose-dependent
manner (Fig. 2B and D). Similarly,
the increased level of phospho-IGF-1R was observed due to the
co-incubation of IGF-1 and daintain/AIF-1. HepG2 cells treated with
IGF-1 (40 μg/l) for 3 h in the presence of 5 μM
daintain/AIF-1 resulted in the maximum increase in
phospho-IGF-1R.
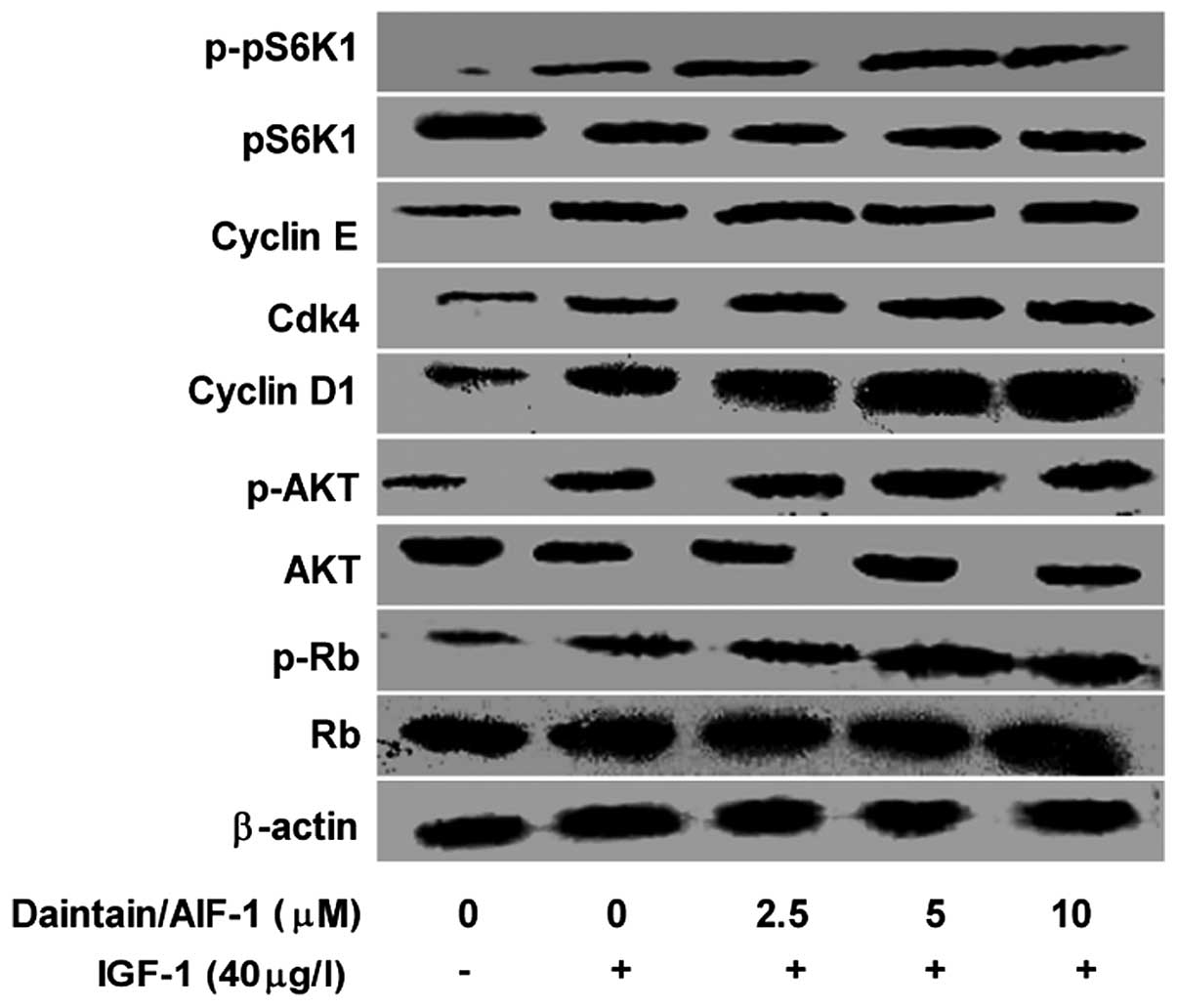
Daintain/AIF-1 enhances the activation of
IGF-1-induced IGF-1R downstream signaling pathway in HepG2
cells
To determine the role of daintain/AIF-1 in the
activation of IGF-1-induced IGF-1R, activation of the downstream
signaling pathway of IGF-1R was also examined by western blot
analysis. Consistent with the results shown in Fig. 3, IGF-1 enhanced the expression of
phospho-AKT, which was further promoted by daintain/AIF-1 in a
dose-dependent manner. Similarly, daintain/AIF-1 at the increasing
concentration resulted in an elevated expression of phospho-pS6K1.
Moreover, in the presence of IGF-1, daintain/AIF-1 stimulation
enhanced the expression of cyclin D1, CDK4 and phosphorylated Rb,
but had no effect on the expression of cyclin E although IGF-1
alone resulted in an increased expression of cyclin E.
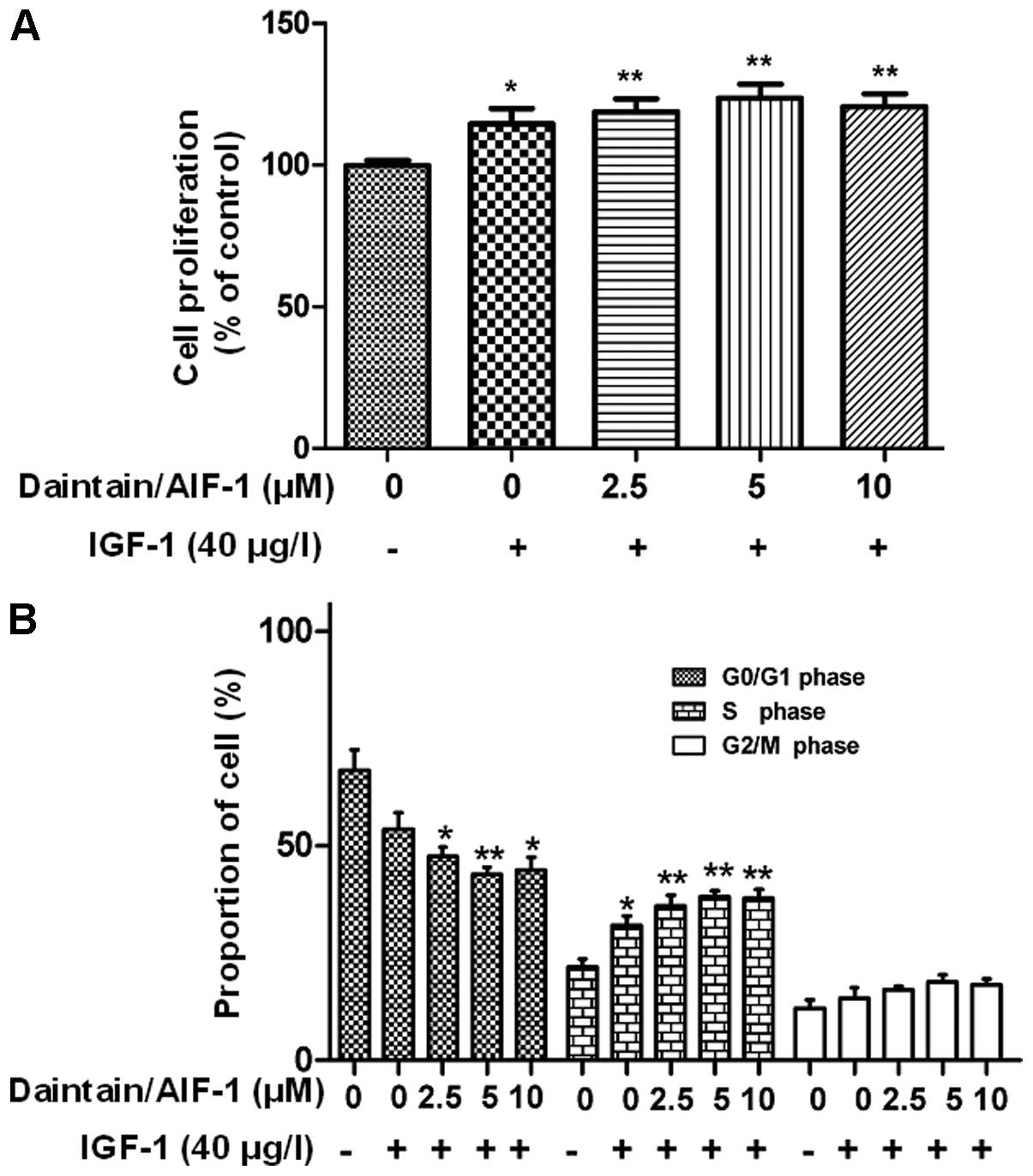
Daintain/AIF-1 promotes IGF-1-induced
proliferation of HepG2 cells
As shown in Fig. 4A,
IGF-1 significantly accelerated the cell growth (115% of the
control) and daintain/AIF-1 promoted IGF-1-induced cell
proliferation. The flow cytometric analysis revealed that IGF-1
stimulation decreased the proportion of HepG2 cells in the G0/G1
phase (from 67.7±5.4 to 54.0±5.1%), but increased the proportion of
HepG2 cells in S phase (from 21.9±1.4 to 31.5±2.8%) and G2/M phase
(from 12.1±2.4 to 14.3±3.0%) (Fig.
4B). More significant changes in the cell cycle were observed
due to the combinatorial application of daintain/AIF-1 and IGF-1.
In the presence of IGF-1 (40 μg/l), daintain/AIF-1 reduced
the ratio of HepG2 cells in the G0/G1 phase and enhanced the
proportion of HepG2 cells in the S and G2/M phase in a
dose-dependent manner (Fig.
4B).
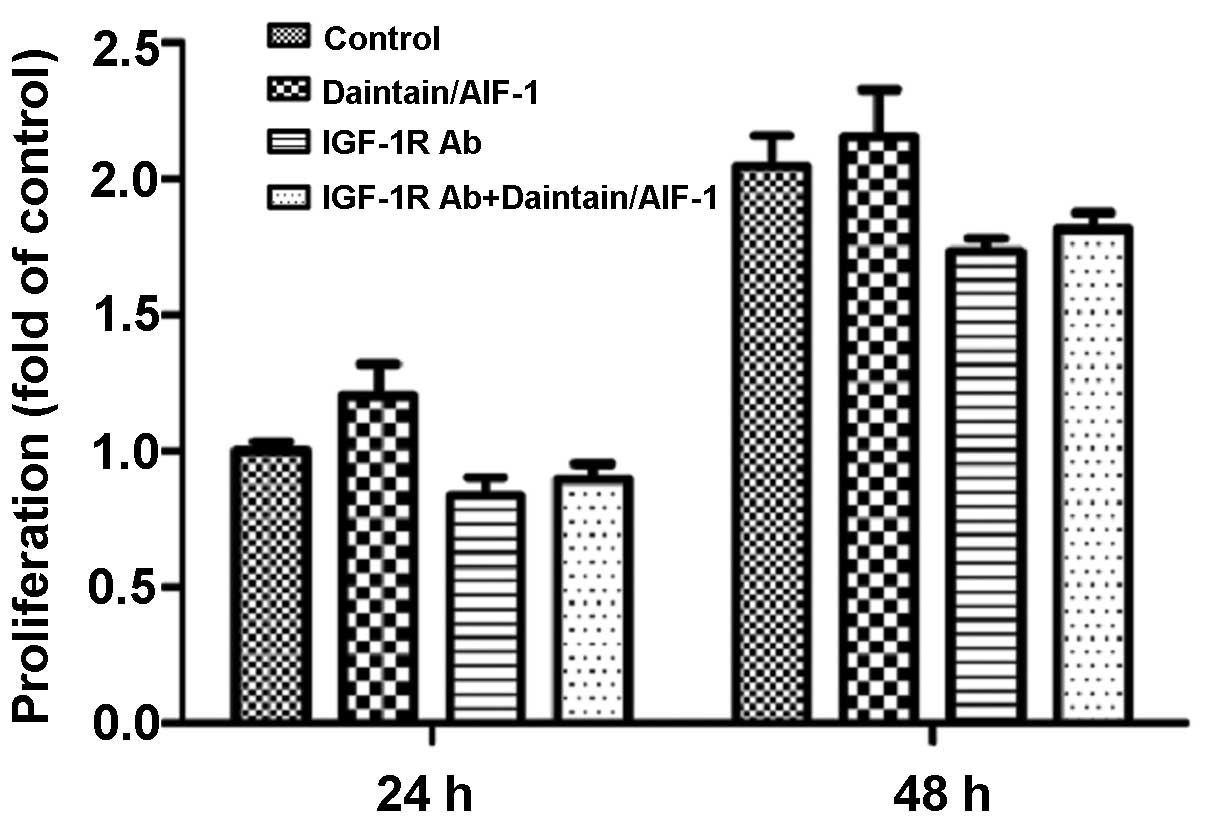
IGF-1R is involved in the promotion of
daintain/AIF-1 on IGF-1-induced HepG2 cell proliferation
To confirm whether IGF-1R plays a role in the
promotion of daintain/AIF-1 on the IGF-1-induced proliferation of
HepG2 cells, the anti-IGF-1R antibody was used to suppress the
expression of IGF-1R. The results showed that the anti-IGF-1R
neutralizing antibody decreased the proliferation of HepG2 cells
under the induction of IGF-1. Furthermore, the promotion effect of
daintain/AIF-1 on IGF-1-induced cell proliferation was eliminated
due to the blockage of IGF-1R. Of note, after the blockage of
IGF-1R, treatment of HepG2 cells with daintain/AIF-1 only caused a
slight increase in the cell number (Fig. 5).
Discussion
Although previous findings have demonstrated that
daintain/AIF-1 promotes breast cancer cell proliferation via the
activation of the NF-κB/cyclin D1 pathway, (16) the precise mechanisms of
daintain/AIF-1 for promoting cancer cell proliferation are not
completely understood. In the present study, we employed HepG2 cell
lines to investigate the molecular mechanism of daintain/AIF-1 on
promoting cell proliferation. Our results showed that
daintain/AIF-1 is closely associated with the activation of
IGF-1-induced IGF-1R and its downstream signaling pathway.
The IGF/IGF-1R axis has been previously found to
play a critical role in the development of HCC (3,7,8).
IGF-1R is predominantly activated by its ligands, such as IGF-1 and
IGF-2. The bioactivity of IGFs can be modulated by IGFBPs due to
the high affinity to IGF-1 and IGF-2 (26). IGFBPs can sequester IGF from IGF-1R,
thereby attenuating the bioactivity of these growth factors.
IGFBP-3 accounts for the majority of circulating IGF (3,27,28).
Treatment of HCC cells with recombinant IGFBP-3 can lead to a
significant reduction in cell proliferation by inhibiting the
activation of IGF-1-induced IGF-1R, ERK and AKT signaling pathways
(23). In our study, daintain/AIF-1
enhanced the secretion of IGF-1 and IGF-2, reduced the secretion of
IGFBP-3, but had no impact on their gene expression. On the basis
of these results, daintain/AIF-1 may be involved in the activation
of IGF-1R signaling pathway in HepG2 cells through the elevated
IGF-1 and IGF-2.
The activation of IGF-1R is a vital process in the
promotion of cell proliferation. Therefore, we monitored
IGF-1-induced activation of IGF-1R in the presence of
daintain/AIF-1 by western blot analysis. Treatment of HepG2 cells
with daintain/AIF-1 for 24 h significantly enhanced IGF-1-induced
tyrosine phosphorylation of IGF-1R in a dose- and time-dependent
manner.
When IGF-1R is stimulated, the tyrosine kinase
activates its downstream signaling pathways, including the Ras/MAPK
and PI3K/AKT pathways. The phosphorylation of MAPK can induce a
subsequent increase in cell proliferation, and the activation of
PI3K/AKT can lead to the modulation of mammalian target of
rapamycin (mTOR), thus resulting in translational adaptation
(29–31). mTOR can regulate cyclin D1
expression and Rb phosphorylation, and the inhibition of mTOR can
arrest cells in the G0/G1 phase of the cell cycle (32,33).
Based on our study, daintain/AIF-1 revealed an obvious enhancement
on the IGF-1-induced phosphorylation of PI3K/AKT and its downstream
target pS6K1. pS6K1 is a key downstream element of the AKT
signaling pathway, and the de-phosphorylation of pS6K1 can block
the G1 cell cycle progression (34). We have also demonstrated that the
incubation of HepG2 cells with daintain/AIF-1 upregulated the
expression of cyclin D1, CDK4 and phosphorylated Rb in the presence
of IGF-1. Cyclin D1 is important in the transition from G1 to S
phase (35,36). Cyclin D1 can couple with CDK4 or
CDK6 to form cyclin D1/CDK complexes, thus correspondingly
resulting in the activation of CDKs and the phosphorylation of Rb
(37,38). Hypophosphorylated Rb can abrogate
the inhibitory effect of Rb on the cell-cycle progression and
accelerate cell entry into the S phase (39).
The physiological function of daintain/AIF-1 on
promoting IGF-1-induced activation of the IGF-1R signaling pathway
was confirmed by the MTT assay. The results showed that
daintain/AIF-1 markedly promoted IGF-1-induced HepG2 cell
proliferation. Data from the flow cytometric analysis also revealed
that daintain/AIF-1 accelerated HepG2 cell entry into the S and
G2/M phase, which is consistent with our previous observation that
daintain/AIF-1 promoted cyclin D1 expression and Rb
phosphorylation.
In our study, the blockage of IGF-1R using the
anti-IGF-1R antibody inhibited IGF-1-induced HepG2 cell
proliferation and eliminated the promotion effect of daintain/AIF-1
on IGF-1-induced HepG2 cell proliferation. These results suggest
that the IGF/IGF-1R signaling pathway plays a crucial role in the
promotion function of daintain/AIF-1 on IGF-1-induced HepG2 cell
proliferation. However, daintain/AIF-1 may have other pathways to
promote HepG2 cell proliferation besides the activation of IGF-1R
signaling pathway. Moreover, we observed the accelerated migration
of HepG2 cells in the presence of IGF-1. However, daintain/AIF-1
failed to modulate IGF-1-induced cell migration (data not
shown).
Taken together, according to our current studies and
knowledge, daintain/AIF-1 induces activation of the IGF/IGF-1R axis
and its downstream signaling pathways by regulating IGF-1, IGF-2
and IGFBP-3 secretion and enhancing IGF-1R sensitivity in the
presence of IGF-1 stimulation, thereby resulting in the
proliferation of HepG2 cells. However, more studies are required to
elucidate the exact mechanisms of daintain/AIF-1 involved in the
activation of the IGF-1R signaling pathway and its downstream
signaling pathways.
Acknowledgments
This study was supported by the National Science
Foundation of China (grant no. 31370773 and 81172511) and the
Chutian Scholar Program to N.C. from Education Department of Hubei
Province.
References
|
1
|
Bosch FX, Ribes J, Díaz M and Cléries R:
Primary liver cancer: worldwide incidence and trends.
Gastroenterology. 127(Suppl 1): S5–S16. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
LeRoith D and Roberts CT Jr: The
insulin-like growth factor system and cancer. Cancer Lett.
195:127–137. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pollak MN, Schernhammer ES and Hankinson
SE: Insulin-like growth factors and neoplasia. Nat Rev Cancer.
4:505–518. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Khandwala HM, McCutcheon IE, Flyvbjerg A
and Friend KE: The effects of insulin-like growth factors on
tumorigenesis and neoplastic growth. Endocr Rev. 21:215–244. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Baserga R: Oncogenes and the strategy of
growth factors. Cell. 79:927–930. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Reynolds AR and Kyprianou N: Growth factor
signalling in prostatic growth: significance in tumour development
and therapeutic targeting. Br J Pharmacol. 147(Suppl 2): S144–S152.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Scharf JG and Braulke T: The role of the
IGF axis in hepatocar-cinogenesis. Horm Metab Res. 35:685–693.
2003. View Article : Google Scholar
|
|
8
|
Alexia C, Fallot G, Lasfer M,
Schweizer-Groyer G and Groyer A: An evaluation of the role of
insulin-like growth factors (IGF) and of type-I IGF receptor
signalling in hepatocarcinogenesis and in the resistance of
hepatocarcinoma cells against drug-induced apoptosis. Biochem
Pharmacol. 68:1003–1015. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen ZW, Ahren B, Ostenson CG, Cintra A,
Bergman T, Möller C, Fuxe K, Mutt V, Jörnvall H and Efendic S:
Identification, isolation, and characterization of daintain
(allograft inflammatory factor 1), a macrophage polypeptide with
effects on insulin secretion and abundantly present in the pancreas
of prediabetic BB rats. Proc Natl Acad Sci USA. 94:13879–13884.
1997. View Article : Google Scholar
|
|
10
|
Utans U, Arceci RJ, Yamashita Y and
Russell ME: Cloning and characterization of allograft inflammatory
factor-1: a novel macrophage factor identified in rat cardiac
allografts with chronic rejection. J Clin Invest. 95:2954–2962.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tian Y, Kelemen SE and Autieri MV:
Inhibition of AIF-1 expression by constitutive siRNA expression
reduces macrophage migration, proliferation, and signal
transduction initiated by atherogenic stimuli. Am J Physiol Cell
Physiol. 290:C1083–C1091. 2006. View Article : Google Scholar
|
|
12
|
Yang ZF, Ho DW, Lau CK, Lam CT, Lum CT,
Poon RT and Fan ST: Allograft inflammatory factor-1 (AIF-1) is
crucial for the survival and pro-inflammatory activity of
macrophages. Int Immunol. 17:1391–1397. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kimura M, Kawahito Y, Obayashi H, Ohta M,
Hara H, Adachi T, Tokunaga D, Hojo T, Hamaguchi M, Omoto A, et al:
A critical role for allograft inflammatory factor-1 in the
pathogenesis of rheumatoid arthritis. J Immunol. 178:3316–3322.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Del Galdo F, Maul GG, Jiménez SA and
Artlett CM: Expression of allograft inflammatory factor 1 in
tissues from patients with systemic sclerosis and in vitro
differential expression of its isoforms in response to transforming
growth factor beta. Arthritis Rheum. 54:2616–2625. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Pashenkov M, Efendic S, Zhu J, Zou LP,
Ostenson CG and Mustafa M: Augmented expression of
daintain/allograft inflammatory factor-1 is associated with
clinical disease: dynamics of daintain/allograft inflammatory
factor-1 expression in spleen, peripheral nerves and sera during
experimental autoimmune neuritis. Scand J Immunol. 52:117–122.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu S, Tan WY, Chen QR, Chen XP, Fu K,
Zhao YY and Chen ZW: Daintain/AIF-1 promotes breast cancer
proliferation via activation of the NF-kappaB/cyclin D1 pathway and
facilitates tumor growth. Cancer Sci. 99:952–957. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li T, Feng Z, Jia S, Wang W, Du Z, Chen N
and Chen Z: Daintain/AIF-1 promotes breast cancer cell migration by
up-regulated TNF-α via activate p38 MAPK signaling pathway. Breast
Cancer Res Treat. 131:891–898. 2012. View Article : Google Scholar
|
|
18
|
Balkwill F and Mantovani A: Inflammation
and cancer: back to Virchow? Lancet. 357:539–545. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Naugler WE, Sakurai T, Kim S, Maeda S, Kim
K, Elsharkawy AM and Karin M: Gender disparity in liver cancer due
to sex differences in MyD88-dependent IL-6 production. Science.
317:121–124. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fausto N, Campbell JS and Riehle KJ: Liver
regeneration. Hepatology. 43(Suppl 1): S45–S53. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nagakawa Y, Nomoto S, Kato Y, Montgomery
RA, Williams GM, Klein AS and Sun Z: over-expression of AIF-1 in
liver allografts and peripheral blood correlates with acute
rejection after transplantation in rats. Am J Transplant.
4:1949–1957. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hirsch J, Hansen KC, Choi S, Noh J, Hirose
R, Roberts JP, Matthay MA, Burlingame AL, Maher JJ and Niemann CU:
Warm ischemia-induced alterations in oxidative and inflammatory
proteins in hepatic Kupffer cells in rats. Mol Cell Proteomics.
5:979–986. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Huynh H, Chow PK, Ooi LL and Soo KC: A
possible role for insulin-like growth factor-binding protein-3
autocrine/paracrine loops in controlling hepatocellular carcinoma
cell proliferation. Cell Growth Differ. 13:115–122. 2002.PubMed/NCBI
|
|
24
|
Aishima S, Basaki Y, Oda Y, Kuroda Y,
Nishihara Y, Taguchi K, Taketomi A, Maehara Y, Hosoi F, Maruyama Y,
et al: High expression of insulin-like growth factor binding
protein-3 is correlated with lower portal invasion and better
prognosis in human hepatocellular carcinoma. Cancer Sci.
97:1182–1190. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lund P, Schubert D, Niketeghad F and
Schirmacher P: Autocrine inhibition of chemotherapy response in
human liver tumor cells by insulin-like growth factor-II. Cancer
Lett. 206:85–96. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Firth SM and Baxter RC: Cellular actions
of the insulin-like growth factor binding proteins. Endocr Rev.
23:824–854. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yakar S, Wu Y, Setser J and Rosen CJ: The
role of circulating IGF-I: lessons from human and animal models.
Endocrine. 19:239–248. 2002. View Article : Google Scholar
|
|
28
|
Bach LA, Headey SJ and Norton RS:
IGF-binding proteins - the pieces are falling into place. Trends
Endocrinol Metab. 16:228–234. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Adams TE, Epa VC, Garrett TP and Ward CW:
Structure and function of the type 1 insulin-like growth factor
receptor. Cell Mol Life Sci. 57:1050–1093. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Butler AA, Yakar S, Gewolb IH, Karas M,
Okubo Y and LeRoith D: Insulin-like growth factor-I receptor signal
transduction: at the interface between physiology and cell biology.
Comp Biochem Physiol B Biochem Mol Biol. 121:19–26. 1998.
View Article : Google Scholar
|
|
31
|
Dearth RK, Cui X, Kim HJ, Hadsell DL and
Lee AV: Oncogenic transformation by the signaling adaptor proteins
insulin receptor substrate (IRS)-1 and IRS-2. Cell Cycle.
6:705–713. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hashemolhosseini S, Nagamine Y, Morley SJ,
Desrivières S, Mercep L and Ferrari S: Rapamycin inhibition of the
G1 to S transition is mediated by effects on cyclin D1 mRNA and
protein stability. J Biol Chem. 273:14424–14429. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chen Y, Knudsen ES and Wang JY: The
RB/p107/p130 phosphorylation pathway is not inhibited in
rapamycin-induced G1-prolongation of NIH3T3 cells. Oncogene.
13:1765–1771. 1996.PubMed/NCBI
|
|
34
|
Gao N, Flynn DC, Zhang Z, Zhong XS, Walker
V, Liu KJ, Shi X and Jiang BH: G1 cell cycle progression and the
expression of G1 cyclins are regulated by PI3K/AKT/mTOR/p70S6K1
signaling in human ovarian cancer cells. Am J Physiol Cell Physiol.
287:C281–C291. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Burkhart DL and Sage J: Cellular
mechanisms of tumour suppression by the retinoblastoma gene. Nat
Rev Cancer. 8:671–682. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Santamaria D and Ortega S: Cyclins and
CDKS in development and cancer: lessons from genetically modified
mice. Front Biosci. 11:1164–1188. 2006. View Article : Google Scholar
|
|
37
|
Malumbres M and Barbacid M: Cell cycle,
CDKs and cancer: a changing paradigm. Nat Rev Cancer. 9:153–166.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lindqvist A, Rodríguez-Bravo V and Medema
RH: The decision to enter mitosis: feedback and redundancy in the
mitotic entry network. J Cell Biol. 185:193–202. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Schuuring E, Verhoeven E, Mooi WJ and
Michalides RJ: Identification and cloning of two overexpressed
genes, U21B31/PRAD1 and EMS1, within the amplified chromosome 11q13
region in human carcinomas. Oncogene. 7:355–361. 1992.PubMed/NCBI
|