Hepatocellular carcinoma (HCC) is the fifth (average
of men and women) most common tumor and the third leading cause of
cancer-related mortality worldwide (1). HCC is characterized by rapid growth,
early vascular invasion, high-grade malignant potential and
multidrug resistance (1–3). Although advances in treatment have
contributed to improved survival, the overall 5-year survival
remains less then 25% (4). Multiple
etiologies, such as aflatoxin (5)
as well as hepatitis B virus (HBV) (6) and hepatitis C virus (HCV) (7) infections, have been linked to HCC,
while no consistent genetic abnormalities have been attributed to
this disease. Numerous mutated proto-oncogenes and tumor-suppressor
genes, as well as signaling pathway abnormalities have been
detected in HCC such as p53, p73, Rb, adenomatous polyposis coli
(APC), DLC-1, DLC-2, PTEN, SOCS1, GSTP1, HCCS1, Smad2/4, AXIN1,
IGF-2, β-catenin, c-myc and cyclin D1 (8–10),
which has hindered the development of effective targeted therapies.
Therefore, identification of novel critical molecules that
contribute to the progression of HCC would be extremely beneficial,
not only for diagnostic/prognostic purposes but also for providing
significant targets for therapeutic intervention. Within the last
decade, overexpression of astrocyte-elevated gene-1 (AEG-1), a
novel oncogene, also known as metadherin (MTDH) and lysine-rich
CEACAM1 co-isolated (LYRIC), has been detected in the majority of
cancers studied to date (11–13),
and its elevated levels are associated with poor prognosis in
cancer patients (14,15). In HCC, AEG-1 has been described as
an essential gene involved in its progression (16).
AEG-1 was originally reported as a novel late
response gene induced in human fetal astrocytes after HIV-1
infection or treatment with viral glycoprotein gp120 or TNF-α
(17). Subsequently, in vivo
phage screening allowed the cloning of mouse AEG-1 as a protein
mediating the metastasis of breast cancer cells to the lung and was
named metadherin (MTDH) (18).
Mouse/rat AEG-1 was also cloned as a tight junction protein named
LYRIC (19) and by gene trapping
techniques and was named 3D3/LYRIC (20).
AEG-1 orthologues are found in most (over 90%)
vertebrate species but not in non-vertebrates. The human AEG-1 gene
is located on chromosome 8q22 having 12 exons/11 introns, and its
genomic amplification has been detected in HCC and breast cancer
(21). The human AEG-1 gene encodes
a 582-amino acid protein with a calculated molecular mass of 64
kDa, and is present in the cell membrane, cytoplasm, nucleus,
nucleolus and endoplasmic reticulum (20,22).
It contains a transmembrane domain and three putative nuclear
localization signals between amino acids 79–91, 432–451 and 561–580
(23). AEG-1 is ubiquitously lowly
expressed in all normal tissues, with higher expression detected in
the skeletal muscle and heart and in endocrine glands such as the
thyroid and adrenal gland (22).
AEG-1 is markedly upregulated in HCC (16,24),
breast (25), gastric (26), gallbladder (27), colorectal (28), prostate (23) and renal (29) cancer, esophageal squamous cell
carcinoma (ESSC) (30), non-small
cell lung cancer (NSCLC) (31),
pancreatic ductal adenocarcinoma (32), tongue carcinoma (33), melanoma (22), glioblastoma multiforme (GBM)
(34), acute myeloid leukemia
(35), neuroblastoma (36), oligodendroglioma (37) andosteosarcoma (38), cervical (39) and ovarian carcinoma (40).
Numerous studies have documented that AEG-1 is
overexpressed in HCC and is closely associated with the disease. In
the earliest study by Yoo et al (16), among 109 HCC samples, only 7 scored
negative for AEG-1 and the remaining 102 (93.58%) showed variable
overexpression levels of AEG-1. Based on the Barcelona Clinic Liver
Cancer (BCLC) staging system, the expression of AEG-1 is gradually
increased with stages from I to IV, and a statistically significant
correlation (P<0.0001) was obtained between the AEG-1 expression
level and the stage of HCC (16).
Our research team also found that AEG-1 was upregulated in HCC
tissues among 60 pairs of HCC samples (41).
In a subsequent study, AEG-1 expression was assessed
by immunohistochemistry in tissue microarrays of 323 HCC patients,
which demonstrated that the majority of the tumor tissues expressed
significantly higher levels of AEG-1 when compared with adjacent
non-tumor tissues; with AEG-1High present in 54.2% (175
of 323) of all the patients (42).
In addition, by Pearson χ2 test, AEG-1 expression was
found to be closely associated with microvascular invasion
(P<0.001), pathologic satellites (P= 0.007), tumor
differentiation (P=0.002) and TNM stage (P=0.001). Moreover,
according to a cohort study, the 1-, 3- and 5-year overall survival
(OS) rates in a high AEG-1-expressing group were significantly
lower than those in a low AEG-1-expressing group (83.0 vs. 89.7%,
52.0 vs. 75.3% and 37.4 vs. 66.9%, respectively); the 1-, 3- and
5-year cumulative recurrence rates were markedly higher in the high
AEG-1-expressing group than those in the low AEG-1-expressing group
(32.4 vs. 16.8%, 61.2 vs. 38.2% and 70.7 vs. 47.8%, respectively).
Furthermore, univariate and multivariate analyses revealed that
along with tumor diameter, encapsulation, microvascular invasion
and TNM stage, AEG-1 was an independent prognostic factor for both
OS (HR=1.870; P<0.001) and recurrence (HR=1.695; P<0.001).
Therefore, the overexpression of AEG-1 in HCC may predict shorter
OS and a higher recurrence rate, and further become a marker for
prognosis in HCC. In a more recent study in China in 89 human HCC
patients, Zheng et al (43)
also confirmed the above results.
In another separate study, AEG-1 expression levels
were identified to be elevated in HBV-related HCC tissues (n=73)
compared to normal liver tissues (n=11) or hepatitis samples
(n=45), and were found to be correlated with the American Joint
Committee on Cancer (AJCC, 7th edition) stage (P=0.020), T
classification (P= 0.007), N classification (P= 0.044), vascular
invasion (P=0.006) and histological differentiation (P=0.020) in
patients with HBV-associated HCC (44). Moreover, patients with high AEG-1
levels had shorter survival times compared to those with low AEG-1
expression (P=0.001) (44).
Additionally, expression of AEG-1 in HCV-related HCC was also
significantly increased in comparison with the expression level in
normal liver and cirrhotic tissue (16).
Taken together, AEG-1 overexpression is consistently
observed in HCC, and its level appears to be correlated with the
stage and grade as well as OS and the recurrence rate of HCC
cases.
In parallel with the evaluation of the
overexpression of AEG-1 in HCC, a substantial body of research has
also highlighted the functions of AEG-1 in mediating the growth,
metastasis and chemoresistance of the disease.
The most fundamental trait of cancer cells involves
their ability to sustain proliferation (45). Previous studies as well as our study
manipulating AEG-1 expression in HCC cells showed that
overexpression of AEG-1 promotes proliferation and increases
anchorage-independent growth in soft agar (16,46);
knockdown of AEG-1 was found to suppress proliferation and inhibit
colony formation as well as induce apoptosis through suppression of
IL-6 secretion (47,48). Further studies also demonstrated
that enhanced AEG-1 expression in HCC cells generated large and
highly vascular subcutaneous tumors compared to the control;
correspondingly, downregulation of the expression of AEG-1
decreased the tumor formation rate and the growth of subcutaneous
tumors in nude mice and the tumor volumes were found to be smaller
than the control (16,47). Additionally, transgenic mice with
hepatocyte-specific expression of AEG-1 (Alb/AEG-1) were treated
with N-nitrosodiethylamine, a hepatocarcinogen. A significant
increase in the ratio of liver weight to body weight and the
presence of more nodules of different sizes were noted in the
Alb/AEG-1 mice when compared to these parameters in the WT mice
(49). Based on the above studies,
we regard AEG-1 as an accelerator of the growth of HCC.
Metastasis is not only the major cause of death from
HCC, but is also the main obstacle to improving the prognosis of
HCC (50,51). In a study using cell lines and a
nude mouse model, downregulation of AEG-1 resulted in the reduced
migratory capacity of HCC cell lines, as well as a reduction in
pulmonary and abdominal metastases in mice (42). It was further demonstrated that the
expression level of AEG-1 was correlated with four
epithelial-to-mesenchymal transition (EMT) markers. Knockdown of
AEG-1 expression in HCC cell lines resulted in downregulation of
N-cadherin and Snail, upregulation of E-cadherin and translocation
of β-catenin (42). Another study
using Huaier polysaccharide also confirmed that downregulation of
AEG-1 inhibited the metastatic potential of HCC cells through EMT
(52). Apart from EMT, anoikis
resistance is an another important capacity to assess tumor
metastatic potential, and is a prerequisite for the survival of
circulating tumor cells in tumor metastasis (45,53).
Our laboratory also demonstrated that AEG-1 enhanced the anoikis
resistance in HCC cells through activation of the
phosphatidylinositol 3-kinase (PI3K)/Akt pathway and Bcl family
proteins to facilitate the metastasis of HCC (54).
Metastasis is a multistep biological process. In
addition to EMT and anoikis resistance, there are other steps
facilitated by AEG-1. Our laboratory found that AEG-1 conferred
orientation chemotaxis to human pulmonary microvascular endothelial
cells (HPMECs) mediated by CXCR4/CXCL12 in HCC cells (54). Moreover, overexpression of AEG-1 was
found to lead to increased production of angiogenic factors, such
as vascular endothelial growth factor (VEGF), placental growth
factor (PIGF) and fibroblast growth factor-α (FGFα) in human HCC
cells, which are essential for angiogenesis and metastasis
(16). In addition, Alb/AEG-1 mice
treated with N-nitrosodiethylamine, presented with multinodular HCC
with steatotic features and associated modulation of expression of
genes regulating invasion and metastasis (TSPAN8 and Lcn2)
(49). In concludion, AEG-1
promotes the metastasis of HCC by multiple steps, and plays a
pivotal role in the poor prognosis of HCC. Thus, suppression of
AEG-1 may be used as a candidate target therapy for HCC.
Chemo-resistance is an important hallmark of HCC.
Recent studies have documented that AEG-1 contributes to
broad-spectrum resistance to various chemotherapeutics including
5-fluorouracil (5-Fu), doxorubicin, paclitaxel, cisplatin and
4-hydroxycyclophosphamide (4-HC) (16,21,55–57).
Hepatocytes isolated from Alb/AEG-1 mice also displayed profound
resistance to chemotherapeutics (49). Furthermore, microarray analysis of
HCC revealed that AEG-1 upregulated several genes implicated in
chemoresistance including drug-metabolizing enzymes, such as
dihydropryimidine dehydrogenase (DPYD), cytochrome P450B6 (CYP2B6)
and dyhydrodiol dehydrogenase (ARK1C2), ATP-binding cassette
transporter ABCC11/MRP8 and the transcription factor LSF/TFCP2
(16). Moreover, AEG-1 was also
found to facilitate the association of multidrug resistance gene
(MDR) 1 mRNA to polysomes resulting in increased translation and
inhibition of ubiquitination and subsequent proteasome-mediated
degradation of MDR1 protein (56).
Therefore, AEG-1 may promote chemoresistance through facilitating
expression of drug-resistant genes at the transcription and
translation levels and the attenuation of the drugs.
Over the past several decades, a large body of
knowledge has been collected regarding Wnt/β-catenin,
mitogen-actived protein kinase (MAPK), NF-κB and PI3K/Akt signaling
pathways as the major signaling pathways activated in HCC (58–61).
These signaling pathways have been demonstrated as being directly
downstream of AEG-1. In addition, AEG-1 is also subtly regulated by
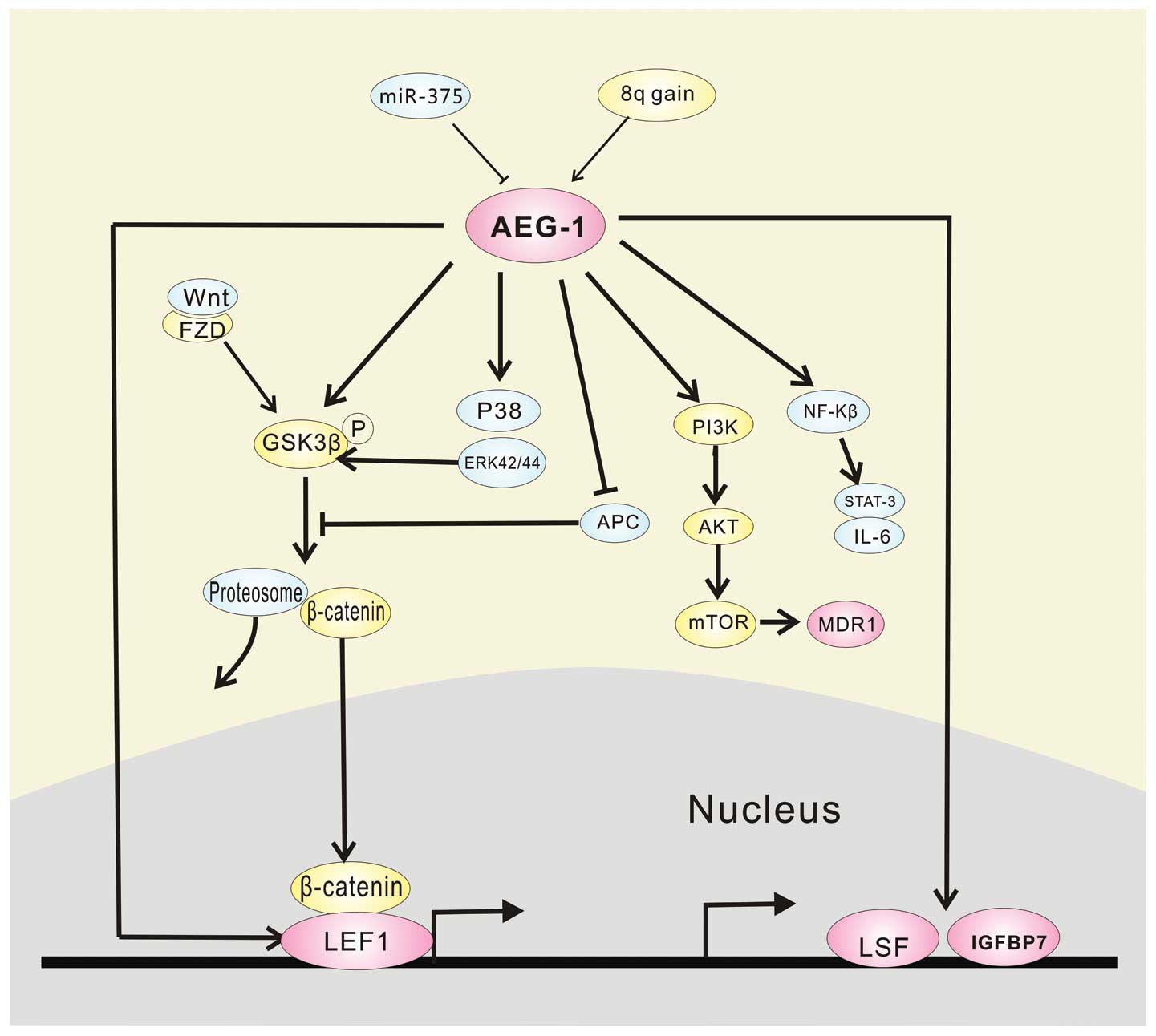
its upstream, such as Haras and miR-375.
AEG-1 is reported as a downstream target of Ha-ras,
and Ha-ras increases the binding of c-Myc to the E-box elements in
the AEG-1 promoter through the PI3K/Akt/GSK3β/c-Myc pathway, which
contributes to Haras-mediated oncogenesis through AEG-1 (62). AEG-1 overexpression is also
associated with elevated copy numbers of it, predominantly due to
gains of large regions of chromosome 8q in HCC (16). We also demonstrated that miR-375
suppressed AEG-1 expression by binding directly to the 3′-UTR of
AEG-1, and the negative regulation of AEG-1 by miR-375 may
contribute partially to the antitumor effects of miR-375 involved
in HCC. Thus, miR-375 is an important regulator of AEG-1 (41).
Increasing evidence indicates that activation of the
WNT/β-catenin-mediated signaling cascade plays a key role in
hepatic oncogenesis (59,63). The transcription factor LEF-1, the
ultimate executor of the Wnt pathway, heterodimerizes with
β-catenin for its action. In the absence of Wnts, β-catenin is
phosphorylated by GSK3β. Conversely, when Wnts are secreted, they
can bind FZD and LRP5/6, which leads to inactivation of GSK3β by
phosphorylation, therefore increasing nuclear translocation of
β-catenin to activate gene transcription such as c-Myc, cyclin D1,
and members of the WISP family (64), faciliting the development of HCC.
The Wnt pathway is activated by AEG-1 in the following ways
(16). i) AEG-1 directly induces
expression of LEF-1 itself as well as LEF-1-induced genes. ii) By
indirectly activating ERK42/44, AEG-1 leads to GSK3β
phosphorylation and inactivation resulting in nuclear translocation
of β-catenin. (iii) AEG-1 downregulates negative regulators of the
Wnt pathway, such as APC and CTBP2.
Aberrant activation of the MAPK pathway also plays a
critical role in the development and progression of HCC (65,66).
Analysis of signal transduction pathways revealed activation of
ERK42/44 and p38 MAPK in Hep-AEG-1 clones compared to control
Hep-pc-4 clones. Inhibition of ERK42/44 and p38 MAPK pathways by
their specific inhibitors PD98059 and SB203580, respectively,
significantly inhibited AEG-1-induced Matrigel inhibition and
anchorage-independent growth, but did not significantly affect
increased proliferation, which indicates that the MEK/ERK and p38
MAPK pathways might mediate a more aggressive phenotype conferred
by AEG-1 (16). Furthermore,
activation of ERK42/44 above through interacting with GSK3β,
crosstalks with the Wnt signaling pathway (16). That is to say, through two different
manners, AEG-1 phosphorylates GSK3β to activate gene transcription.
Additionally, AEG-1 also induces phosphorylation and inactivation
of retinoid X receptor by activating ERK and p38MAPK signaling,
which is indispensable to drive the oncogenic functions of HCC such
as proliferation and apoptosis (67).
Finally, AEG-1 regulates these major signaling
pathways and plays a crucial role in the development of HCC.
However, there is still a need to explore the network of AEG-1 in
HCC, and AEG-1 also regulates the expression of many downstream
genes such as late SV40 factor (LSF), insulin-like growth factor
binding protein-7 (IGFBP7) and participates in RISC by interacting
with SND1, which acts to accelerate HCC.
LSF, an ubiquitous transcription factor, has been
demonstrated to function as an oncogene in HCC (75). It is highly expressed in HCC, and
transcriptionally modulates specific genes, such as
metalloproteinase-9 (MMP-9), c-Met and osteopontin (OPN), resulting
in the regulation of proliferation, invasion, angiogenesis and
chemoresistance of HCC (76,77).
LSF has been identified as an AEG-1 downstream gene by Affymetrix
microarray comparing global gene expression profiles between
AEG-1-overexpressed clones of HepG3 cells and controls, which was
also confirmed by TaqMan quantitative PCR (16). A subsequent study reported that LSF
mRNA expression was ~15-fold higher in AEG-1-overexpressed clones
compared to a control, which also indcates a potential role of LSF
in mediating the oncogenic functions of AEG-1 (75).
IGFBP7, a secreted protein belonging to the IGFBP
family, functions as a potential tumor suppressor in HCC (78). Multiple studies have documented that
IGFBP7 expression is significantly decreased in HCC (79,80),
and it profoundly decreases the viability and induces apoptosis in
multiple human HCC cell lines and inhibits primary tumor growth and
intrahepatic metastasis in orthotopic xenograft models (78). Notably, IGFBP7 has been identified
as the most robustly downregulated gene by AEG-1 in HCC (16). Another study also demonstrated that
stable IGFBP7-overexpressing clones were established in
Hep-AEG-1–14 background, and forced overexpression of IGFBP7 in
AEG-1-overexpressing HCC cells inhibited in vitro growth and
induced senescence, and profoundly suppressed in vivo growth
in nude mice (79). Thus, mediated
by LSF and IGFBP7, AEG-1 plays an important role in HCC
progression.
Staphylococcal nuclease domain-containing 1 (SND1)
is a multifunctional protein modulating a variety of cellular
processes such as transcription (81,82),
RNA splicing (83) and RNA
metabolism (84). SND1 is
overexpressed in HCC (85);
knockdown of SND1 leads to reduced HCC cell proliferation, clone
formation and tumor formation in nude mice (86); and it also regulates HCC
angiogenesis by activation of NF-κB and miR-221 inducing angiogenic
factors such as angiogenin and CXCL16 (87). The identification of SND1 as an
AEG-1-interacting protein was found using two independent
approaches, including yeast two hybrid screening using a human
liver cDNA library and isolation of AEG-1 interacting proteins by
co-immunoprecipitation followed by mass spectrometry. Moreover,
immunofluorescence and co-immunoprecipitation analyses further
demonstrated that AEG-1 interacts with SND1 via the region 101–205
a.a. in the cytoplasm (85). It was
also documented that both AEG-1 and SND1 are required for optimum
RNA-induced silencing complex (RISC) activity (85). Moreover, increased RISC activity,
conferred by AEG-1 or SND1, was found to result in increased
degradation of tumor-suppressor mRNAs, which are the target of
oncomiRs, including PTEN (target of miR-221 and miR-21), CDKN1C
(target of miR-221), CDKN1A (target of miR-106b), SPRY2 (target of
miR-21) and TGFBR2 (target of miR-93) (85).
Since AEG-1 is markedly overexpressed in HCC tissues
and its levels are tightly correlated with the stage and grade as
well as the OS and recurrence rate of the disease, it might serve
as a potential diagnostic/prognostic marker for HCC. In addition to
HCC, in breast cancer, prostate cancer, ESSC, NSCLC, some subtypes
of brain cancer such as GBM, and colorectal carcinoma, AEG-1
expression is also correlated with the stage or outcome of these
diseases (23,25,30,31,34,88).
Thus, AEG-1 may be a universal diagnostic/prognostic marker for
cancer including HCC.
As known, HCC is a progressive and highly
chemoresistant cancer, and there is no effective therapy for
advanced HCC. The only FDA-approved targeted drug, the multikinase
inhibitor sorafenib, provides a survival benefit of only 2.8 months
in non-resectable HCC patients (89). AEG-1 is a key molecule involved in
several important signaling pathways which mediate the progression
of HCC and is markedly over-expressed in HCC. Thus, specific
inhibition of AEG-1 may be a strategy to counteract the progression
of HCC. Moreover, as mentioned above, AEG-1 overexpression
contributes to HCC drug resistance at multiple levels. Therefore,
specific inhibition of AEG-1 not only blocks HCC progression, but
also enhances the effect of anti-HCC drugs such as 5-Fu. A
combination of AEG-1 inhibitors with chemotherapeutics may be an
effective treatment for HCC. A lentivirus delivering AEG-1 siRNA in
combination with 5-Fu was found to markedly inhibit the growth of
QGY-7703 HCC cell xenografts in athymic nude mice when compared to
either agent alone, and the combination treatment reduced the tumor
volume and tumor weight ~70% compared to the control (55).
To date, it has been established that AEG-1 is
frequently upregulated and functions as an oncogene by regulating
several major signaling pathways in HCC as summarized in Fig. 1. Given the importance of AEG-1 in
HCC carcinogenesis, it is not surprising that the potential
clinical application of AEG-1 in HCC diagnosis and therapy warrants
further investigation. In addition to tissue AEG-1, whether AEG-1
in blood, urine or other secretions is also associated with the
stage and grade of HCC needs to be determined. Moreover, methods to
detect these levels efficiently are vitally needed. Therefore,
further studies using large cohorts of patients are warranted to
resolve these issues. Moreover, there are still challenges in
regards to the means of transport of AEG-1 inhibitors in the
clinical therapy of HCC. A safe and effective carrier to deliver
inhibitors of AEG-1 into HCC cells is needed. Recently, gold
nanoparticles have demonstrated increasingly wide applications in
drug delivery due to their unique physicochemical and optical
properties as well as their low toxicity when compared to organic
nanocarriers (90,91). These may be helpful as a new means
of transport of AEG-1 inhibitors in clinical application.
This study was financially supported by the National
Natural Science Foundation of China (nos. 81372663 and 81472832)
and the Outstanding Youth Science Fundation of Tongji Hospital (no.
YXQN005).
|
1
|
El Serag HB: Hepatocellular carcinoma. N
Engl J Med. 365:1118–1127. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pang RW, Joh JW, Johnson PJ, Monden M,
Pawlik TM and Poon RT: Biology of hepatocellular carcinoma. Ann
Surg Oncol. 15:962–971. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
El Serag HB and Rudolph KL: Hepatocellular
carcinoma: Epidemiology and molecular carcinogenesis.
Gastroenterology. 132:2557–2576. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Altekruse SF, McGlynn KA and Reichman ME:
Hepatocellular carcinoma incidence, mortality, and survival trends
in the United States from 1975 to 2005. J Clin Oncol. 27:1485–1491.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hagymási K and Tulassay Z: Epidemiology,
risk factors and molecular pathogenesis of primary liver cancer.
Orv Hetil. 149:541–548. 2008.In Hungarian. View Article : Google Scholar
|
|
6
|
Feitelson MA and Duan LX: Hepatitis B
virus X antigen in the pathogenesis of chronic infections and the
development of hepatocellular carcinoma. Am J Pathol.
150:1141–1157. 1997.PubMed/NCBI
|
|
7
|
Majumder M, Ghosh AK, Steele R, Ray R and
Ray RB: Hepatitis C virus NS5A physically associates with p53 and
regulates p21/waf1 gene expression in a p53-dependent manner. J
Virol. 75:1401–1407. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Boyault S, Rickman DS, de Reyniès A,
Balabaud C, Rebouissou S, Jeannot E, Hérault A, Saric J, Belghiti
J, Franco D, et al: Transcriptome classification of HCC is related
to gene alterations and to new therapeutic targets. Hepatology.
45:42–52. 2007. View Article : Google Scholar
|
|
9
|
Katoh H, Ojima H, Kokubu A, Saito S, Kondo
T, Kosuge T, Hosoda F, Imoto I, Inazawa J, Hirohashi S, et al:
Genetically distinct and clinically relevant classification of
hepatocellular carcinoma: Putative therapeutic targets.
Gastroenterology. 133:1475–1486. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mann CD, Neal CP, Garcea G, Manson MM,
Dennison AR and Berry DP: Prognostic molecular markers in
hepatocellular carcinoma: A systematic review. Eur J Cancer.
43:979–992. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sarkar D and Fisher PB: AEG-1/MTDH/LYRIC:
Clinical significance. Adv Cancer Res. 120:39–74. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yoo BK, Emdad L, Lee SG, Su ZZ,
Santhekadur P, Chen D, Gredler R, Fisher PB and Sarkar D: Astrocyte
elevated gene-1 (AEG-1): A multifunctional regulator of normal and
abnormal physiology. Pharmacol Ther. 130:1–8. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lee SG, Kang DC, DeSalle R, Sarkar D and
Fisher PB: AEG-1/MTDH/LYRIC, the beginning: Initial cloning,
structure, expression profile, and regulation of expression. Adv
Cancer Res. 120:1–38. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wan L, Lu X, Yuan S, Wei Y, Guo F, Shen M,
Yuan M, Chakrabarti R, Hua Y, Smith HA, et al: MTDH-SND1
interaction is crucial for expansion and activity of
tumor-initiating cells in diverse oncogene- and carcinogen-induced
mammary tumors. Cancer Cell. 26:92–105. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ying Z, Li J and Li M: Astrocyte elevated
gene 1: Biological functions and molecular mechanism in cancer and
beyond. Cell Biosci. 1:362011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yoo BK, Emdad L, Su ZZ, Villanueva A,
Chiang DY, Mukhopadhyay ND, Mills AS, Waxman S, Fisher RA, Llovet
JM, et al: Astrocyte elevated gene-1 regulates hepatocellular
carcinoma development and progression. J Clin Invest. 119:465–477.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Su ZZ, Kang DC, Chen Y, Pekarskaya O, Chao
W, Volsky DJ and Fisher PB: Identification and cloning of human
astrocyte genes displaying elevated expression after infection with
HIV-1 or exposure to HIV-1 envelope glycoprotein by rapid
subtraction hybridization, RaSH. Oncogene. 21:3592–3602. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Brown DM and Ruoslahti E: Metadherin, a
cell surface protein in breast tumors that mediates lung
metastasis. Cancer Cell. 5:365–374. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Britt DE, Yang DF, Yang DQ, Flanagan D,
Callanan H, Lim YP, Lin SH and Hixson DC: Identification of a novel
protein, LYRIC, localized to tight junctions of polarized
epithelial cells. Exp Cell Res. 300:134–148. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sutherland HG, Lam YW, Briers S, Lamond AI
and Bickmore WA: 3D3/lyric: A novel transmembrane protein of the
endoplasmic reticulum and nuclear envelope, which is also present
in the nucleolus. Exp Cell Res. 294:94–105. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hu G, Chong RA, Yang Q, Wei Y, Blanco MA,
Li F, Reiss M, Au JL, Haffty BG and Kang Y: MTDH activation by 8q22
genomic gain promotes chemoresistance and metastasis of
poor-prognosis breast cancer. Cancer Cell. 15:9–20. 2009.
View Article : Google Scholar :
|
|
22
|
Kang DC, Su ZZ, Sarkar D, Emdad L, Volsky
DJ and Fisher PB: Cloning and characterization of HIV-1-inducible
astrocyte elevated gene-1, AEG-1. Gene. 353:8–15. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Thirkettle HJ, Girling J, Warren AY, Mills
IG, Sahadevan K, Leung H, Hamdy F, Whitaker HC and Neal DE:
LYRIC/AEG-1 is targeted to different subcellular compartments by
ubiquitinylation and intrinsic nuclear localization signals. Clin
Cancer Res. 15:3003–3013. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sarkar D: AEG-1/MTDH/LYRIC in liver
cancer. Adv Cancer Res. 120:193–221. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li J, Zhang N, Song LB, Liao WT, Jiang LL,
Gong LY, Wu J, Yuan J, Zhang HZ, Zeng MS, et al: Astrocyte elevated
gene-1 is a novel prognostic marker for breast cancer progression
and overall patient survival. Clin Cancer Res. 14:3319–3326. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jianbo X, Hui W, Yulong H, Changhua Z,
Longjuan Z, Shirong C and Wenhua Z: Astrocyte-elevated gene-1
overexpression is associated with poor prognosis in gastric cancer.
Med Oncol. 28:455–462. 2011. View Article : Google Scholar
|
|
27
|
Sun W, Fan YZ, Xi H, Lu XS, Ye C and Zhang
JT: Astrocyte elevated gene-1 overexpression in human primary
gallbladder carcinomas: An unfavorable and independent prognostic
factor. Oncol Rep. 26:1133–1142. 2011.PubMed/NCBI
|
|
28
|
Song HT, Qin Y, Yao GD, Tian ZN, Fu SB and
Geng JS: Astrocyte elevated gene-1 mediates glycolysis and
tumorigenesis in colorectal carcinoma cells via AMPK signaling.
Mediators Inflamm. 2014:2873812014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chen W, Ke Z, Shi H, Yang S and Wang L:
Overexpression of AEG-1 in renal cell carcinoma and its correlation
with tumor nuclear grade and progression. Neoplasma. 57:522–529.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yu C, Chen K, Zheng H, Guo X, Jia W, Li M,
Zeng M, Li J and Song L: Overexpression of astrocyte elevated
gene-1 (AEG-1) is associated with esophageal squamous cell
carcinoma (ESCC) progression and pathogenesis. Carcinogenesis.
30:894–901. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Song L, Li W, Zhang H, Liao W, Dai T, Yu
C, Ding X, Zhang L and Li J: Over-expression of AEG-1 significantly
associates with tumour aggressiveness and poor prognosis in human
non-small cell lung cancer. J Pathol. 219:317–326. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Huang Y, Ren GP, Xu C, Dong SF, Wang Y,
Gan Y, Zhu L and Feng TY: Expression of astrocyte elevated gene-1
(AEG-1) as a biomarker for aggressive pancreatic ductal
adenocarcinoma. BMC Cancer. 14:4792014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ke ZF, He S, Li S, Luo D, Feng C and Zhou
W: Expression characteristics of astrocyte elevated gene-1 (AEG-1)
in tongue carcinoma and its correlation with poor prognosis. Cancer
Epidemiol. 37:179–185. 2013. View Article : Google Scholar
|
|
34
|
Emdad L, Sarkar D, Lee SG, Su ZZ, Yoo BK,
Dash R, Yacoub A, Fuller CE, Shah K, Dent P, et al: Astrocyte
elevated gene-1: A novel target for human glioma therapy. Mol
Cancer Ther. 9:79–88. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Long M, Hao M, Dong K, Shen J, Wang X, Lin
F, Liu L, Wei J, Liang Y, Yang J, et al: AEG-1 overexpression is
essential for maintenance of malignant state in human AML cells via
up-regulation of Akt1 mediated by AuRKA activation. Cell Signal.
25:1438–1446. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lee SG, Jeon HY, Su ZZ, Richards JE,
Vozhilla N, Sarkar D, Van Maerken T and Fisher PB: Astrocyte
elevated gene-1 contributes to the pathogenesis of neuroblastoma.
Oncogene. 28:2476–2484. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Xia Z, Zhang N, Jin H, Yu Z, Xu G and
Huang Z: Clinical significance of astrocyte elevated gene-1
expression in human oligodendrogliomas. Clin Neurol Neurosurg.
112:413–419. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang F, Ke ZF, Sun SJ, Chen WF, Yang SC,
Li SH, Mao XP and Wang LT: Oncogenic roles of astrocyte elevated
gene-1 (AEG-1) in osteosarcoma progression and prognosis. Cancer
Biol Ther. 12:539–548. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Long M, Dong K, Gao P, Wang X, Liu L, Yang
S, Lin F, Wei J and Zhang H: Overexpression of astrocyte-elevated
gene-1 is associated with cervical carcinoma progression and
angiogenesis. Oncol Rep. 30:1414–1422. 2013.PubMed/NCBI
|
|
40
|
Li C, Chen K, Cai J, Shi QT, Li Y, Li L,
Song H, Qiu H, Qin Y and Geng JS: Astrocyte elevated gene-1: A
novel independent prognostic biomarker for metastatic ovarian
tumors. Tumour Biol. 35:3079–3085. 2014. View Article : Google Scholar
|
|
41
|
He XX, Chang Y, Meng FY, Wang MY, Xie QH,
Tang F, Li PY, Song YH and Lin JS: MicroRNA-375 targets AEG-1 in
hepatocellular carcinoma and suppresses liver cancer cell growth in
vitro and in vivo. Oncogene. 31:3357–3369. 2012. View Article : Google Scholar
|
|
42
|
Zhu K, Dai Z, Pan Q, Wang Z, Yang GH, Yu
L, Ding ZB, Shi GM, Ke AW, Yang XR, et al: Metadherin promotes
hepatocellular carcinoma metastasis through induction of
epithelial-mesenchymal transition. Clin Cancer Res. 17:7294–7302.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zheng J, Li C, Wu X, Yang Y, Hao M, Sheng
S, Sun Y, Zhang H, Long J and Hu C: Astrocyte elevated gene-1 is a
novel biomarker of epithelial-mesenchymal transition and
progression of hepatocellular carcinoma in two China regions.
Tumour Biol. 35:2265–2269. 2014. View Article : Google Scholar
|
|
44
|
Gong Z, Liu W, You N, Wang T, Wang X, Lu
P, Zhao G, Yang P, Wang D and Dou K: Prognostic significance of
metadherin over-expression in hepatitis B virus-related
hepatocellular carcinoma. Oncol Rep. 27:2073–2079. 2012.PubMed/NCBI
|
|
45
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Srivastava J, Siddiq A, Gredler R, Shen
XN, Rajasekaran D, Robertson CL, Subler MA, Windle JJ, Dumur CI,
Mukhopadhyay ND, et al: Astrocyte elevated gene-1 (AEG-1) and c-Myc
cooperate to promote hepatocarcinogenesis. Hepatology. 61:915–929.
2014. View Article : Google Scholar
|
|
47
|
Deng H, Zhou Z, Tu W, Xia Y, Huang H and
Tian D: Knockdown of astrocyte elevated gene-1 inhibits growth
through suppression of IL-6 secretion in HepG2 human hepatoma
cells. Oncol Lett. 7:101–106. 2014.
|
|
48
|
Ma J, Xie SL, Geng YJ, Jin S, Wang GY and
Lv GY: In vitro regulation of hepatocellular carcinoma cell
viability, apoptosis, invasion, and AEG-1 expression by LY294002.
Clin Res Hepatol Gastroenterol. 38:73–80. 2014. View Article : Google Scholar
|
|
49
|
Srivastava J, Siddiq A, Emdad L,
Santhekadur PK, Chen D, Gredler R, Shen XN, Robertson CL, Dumur CI,
Hylemon PB, et al: Astrocyte elevated gene-1 promotes
hepatocarcinogenesis: Novel insights from a mouse model.
Hepatology. 56:1782–1791. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Tang ZY, Ye SL, Liu YK, Qin LX, Sun HC, Ye
QH, Wang L, Zhou J, Qiu SJ, Li Y, et al: A decade’s studies on
metastasis of hepato cellular carcinoma. J Cancer Res Clin Oncol.
130:187–196. 2004. View Article : Google Scholar
|
|
51
|
Peng YF, Shi YH, Ding ZB, Ke AW, Gu CY,
Hui B, Zhou J, Qiu SJ, Dai Z and Fan J: Autophagy inhibition
suppresses pulmonary metastasis of HCC in mice via impairing
anoikis resistance and colonization of HCC cells. Autophagy.
9:2056–2068. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Zheng J, Li C, Wu X, Liu M, Sun X, Yang Y,
Hao M, Sheng S, Sun Y, Zhang H, et al: Huaier polysaccharides
suppresses hepatocarcinoma MHCC97-H cell metastasis via
inactivation of EMT and AEG-1 pathway. Int J Biol Macromol.
64:106–110. 2014. View Article : Google Scholar
|
|
53
|
Frisch SM and Screaton RA: Anoikis
mechanisms. Curr Opin Cell Biol. 13:555–562. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Zhou Z, Deng H, Yan W, Luo M, Tu W, Xia Y,
He J, Han P, Fu Y and Tian D: AEG-1 promotes anoikis resistance and
orientation chemotaxis in hepatocellular carcinoma cells. PLoS One.
9:e1003722014. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Yoo BK, Gredler R, Vozhilla N, Su ZZ, Chen
D, Forcier T, Shah K, Saxena U, Hansen U, Fisher PB, et al:
Identification of genes conferring resistance to 5-fluorouracil.
Proc Natl Acad Sci USA. 106:12938–12943. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Yoo BK, Chen D, Su ZZ, Gredler R, Yoo J,
Shah K, Fisher PB and Sarkar D: Molecular mechanism of
chemoresistance by astrocyte elevated gene-1. Cancer Res.
70:3249–3258. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Liu H, Song X, Liu C, Xie L, Wei L and Sun
R: Knockdown of astrocyte elevated gene-1 inhibits proliferation
and enhancing chemo-sensitivity to cisplatin or doxorubicin in
neuroblastoma cells. J Exp Clin Cancer Res. 28:192009. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Min L, He B and Hui L: Mitogen-activated
protein kinases in hepatocellular carcinoma development. Semin
Cancer Biol. 21:10–20. 2011. View Article : Google Scholar
|
|
59
|
Thompson MD and Monga SP: WNT/beta-catenin
signaling in liver health and disease. Hepatology. 45:1298–1305.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Villanueva A, Chiang DY, Newell P, Peix J,
Thung S, Alsinet C, Tovar V, Roayaie S, Minguez B, Sole M, et al:
Pivotal role of mTOR signaling in hepatocellular carcinoma.
Gastroenterology. 135:1972–1983. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Pikarsky E, Porat RM, Stein I, Abramovitch
R, Amit S, Kasem S, Gutkovich-Pyest E, Urieli-Shoval S, Galun E and
Ben-Neriah Y: NF-kappaB functions as a tumour promoter in
inflammation-associated cancer. Nature. 431:461–466. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Lee SG, Su ZZ, Emdad L, Sarkar D and
Fisher PB: Astrocyte elevated gene-1 (AEG-1) is a target gene of
oncogenic Ha-ras requiring phosphatidylinositol 3-kinase and c-Myc.
Proc Natl Acad Sci USA. 103:17390–17395. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Wands JR and Kim M: WNT/β-catenin
signaling and hepato-cellular carcinoma. Hepatology. 60:452–454.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Tanaka S, Sugimachi K, Kameyama T, Maehara
S, Shirabe K, Shimada M, Wands JR and Maehara Y: Human WISP1v, a
member of the CCN family, is associated with invasive
cholan-giocarcinoma. Hepatology. 37:1122–1129. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Wang S, Huang X, Li Y, Lao H, Zhang Y,
Dong H, Xu W, Li JL and Li M: RN181 suppresses hepatocellular
carcinoma growth by inhibition of the ERK/MAPK pathway. Hepatology.
53:1932–1942. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Guichard C, Amaddeo G, Imbeaud S, Ladeiro
Y, Pelletier L, Maad IB, Calderaro J, Bioulac-Sage P, Letexier M,
Degos F, et al: Integrated analysis of somatic mutations and focal
copy-number changes identifies key genes and pathways in
hepatocellular carcinoma. Nat Genet. 44:694–698. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Srivastava J, Robertson CL, Rajasekaran D,
Gredler R, Siddiq A, Emdad L, Mukhopadhyay ND, Ghosh S, Hylemon PB,
Gil G, et al: AEG-1 regulates retinoid X receptor and inhibits
retinoid signaling. Cancer Res. 74:4364–4377. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Emdad L, Sarkar D, Su ZZ, Randolph A,
Boukerche H, Valerie K and Fisher PB: Activation of the nuclear
factor κB pathway by astrocyte elevated gene-1: Implications for
tumor progression and metastasis. Cancer Res. 66:1509–1516. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Sarkar D, Park ES, Emdad L, Lee SG, Su ZZ
and Fisher PB: Molecular basis of nuclear factor-κB activation by
astrocyte elevated gene-1. Cancer Res. 68:1478–1484. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Hösel M, Quasdorff M, Wiegmann K, Webb D,
Zedler U, Broxtermann M, Tedjokusumo R, Esser K, Arzberger S,
Kirschning CJ, et al: Not interferon, but interleukin-6 controls
early gene expression in hepatitis B virus infection. Hepatology.
50:1773–1782. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Tai DI, Tsai SL, Chang YH, Huang SN, Chen
TC, Chang KS and Liaw YF: Constitutive activation of nuclear factor
κB in hepato-cellular carcinoma. Cancer. 89:2274–2281. 2000.
View Article : Google Scholar
|
|
72
|
Liu P, Kimmoun E, Legrand A, Sauvanet A,
Degott C, Lardeux B and Bernuau D: Activation of NF-κB, AP-1 and
STAT transcription factors is a frequent and early event in human
hepatocellular carcinomas. J Hepatol. 37:63–71. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Robertson CL, Srivastava J, Siddiq A,
Gredler R, Emdad L, Rajasekaran D, Akiel M, Shen XN, Guo C,
Giashuddin S, et al: Genetic deletion of AEG-1 prevents
hepatocarcinogenesis. Cancer Res. 74:6184–6193. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Grabinski N, Ewald F, Hofmann BT, Staufer
K, Schumacher U, Nashan B and Jücker M: Combined targeting of AKT
and mTOR synergistically inhibits proliferation of hepatocellular
carcinoma cells. Mol Cancer. 11:852012. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Yoo BK, Emdad L, Gredler R, Fuller C,
Dumur CI, Jones KH, Jackson-Cook C, Su ZZ, Chen D, Saxena UH, et
al: Transcription factor Late SV40 Factor (LSF) functions as an
oncogene in hepatocellular carcinoma. Proc Natl Acad Sci USA.
107:8357–8362. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Santhekadur PK, Gredler R, Chen D, Siddiq
A, Shen XN, Das SK, Emdad L, Fisher PB and Sarkar D: Late SV40
factor (LSF) enhances angiogenesis by transcriptionally
up-regulating matrix metalloproteinase-9 (MMP-9). J Biol Chem.
287:3425–3432. 2012. View Article : Google Scholar :
|
|
77
|
Yoo BK, Gredler R, Chen D, Santhekadur PK,
Fisher PB and Sarkar D: c-Met activation through a novel pathway
involving osteopontin mediates oncogenesis by the transcription
factor LSF. J Hepatol. 55:1317–1324. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Chen D, Siddiq A, Emdad L, Rajasekaran D,
Gredler R, Shen XN, Santhekadur PK, Srivastava J, Robertson CL,
Dmitriev I, et al: Insulin-like growth factor-binding protein-7
(IGFBP7): A promising gene therapeutic for hepatocellular carcinoma
(HCC). Mol Ther. 21:758–766. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Chen D, Yoo BK, Santhekadur PK, Gredler R,
Bhutia SK, Das SK, Fuller C, Su ZZ, Fisher PB and Sarkar D:
Insulin-like growth factor-binding protein-7 functions as a
potential tumor suppressor in hepatocellular carcinoma. Clin Cancer
Res. 17:6693–6701. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Tomimaru Y, Eguchi H, Wada H, Kobayashi S,
Marubashi S, Tanemura M, Umeshita K, Kim T, Wakasa K, Doki Y, et
al: IGFBP7 downregulation is associated with tumor progression and
clinical outcome in hepatocellular carcinoma. Int J Cancer.
130:319–327. 2012. View Article : Google Scholar
|
|
81
|
Leverson JD, Koskinen PJ, Orrico FC,
Rainio EM, Jalkanen KJ, Dash AB, Eisenman RN and Ness SA: Pim-1
kinase and p100 cooperate to enhance c-Myb activity. Mol Cell.
2:417–425. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Wang X, Liu X, Fang J, Lu Y, He J, Yao X,
Yao Z and Yang J: Coactivator P100 protein enhances STAT6-dependent
transcriptional activation but has no effect on STAT1-mediated gene
transcription. Anat Rec. 293:1010–1016. 2010. View Article : Google Scholar
|
|
83
|
Gao X, Zhao X, Zhu Y, He J, Shao J, Su C,
Zhang Y, Zhang W, Saarikettu J, Silvennoinen O, et al: Tudor
staphylococcal nuclease (Tudor-SN) participates in small
ribonucleoprotein (snRNP) assembly via interacting with
symmetrically dimethylated Sm proteins. J Biol Chem.
287:18130–18141. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Gao X, Ge L, Shao J, Su C, Zhao H,
Saarikettu J, Yao X, Yao Z, Silvennoinen O and Yang J: Tudor-SN
interacts with and co-localizes with G3BP in stress granules under
stress conditions. FEBS Lett. 584:3525–3532. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Yoo BK, Santhekadur PK, Gredler R, Chen D,
Emdad L, Bhutia S, Pannell L, Fisher PB and Sarkar D: Increased
RNA-induced silencing complex (RISC) activity contributes to
hepatocellular carcinoma. Hepatology. 53:1538–1548. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Yin J, Ding J, Huang L, Tian X, Shi X, Zhi
L, Song J, Zhang Y, Gao X, Yao Z, et al: SND1 affects proliferation
of hepato-cellular carcinoma cell line SMMC-7721 by regulating
IGFBP3 expression. Anat Rec. 296:1568–1575. 2013. View Article : Google Scholar
|
|
87
|
Santhekadur PK, Das SK, Gredler R, Chen D,
Srivastava J, Robertson C, Baldwin AS Jr, Fisher PB and Sarkar D:
Multifunction protein staphylococcal nuclease domain containing 1
(SND1) promotes tumor angiogenesis in human hepatocellular
carcinoma through novel pathway that involves nuclear factor κB and
miR-221. J Biol Chem. 287:13952–13958. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Song H, Li C, Li R and Geng J: Prognostic
significance of AEG-1 expression in colorectal carcinoma. Int J
Colorectal Dis. 25:1201–1209. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Llovet JM, Ricci S, Mazzaferro V, Hilgard
P, Gane E, Blanc JF, de Oliveira AC, Santoro A, Raoul JL, Forner A,
et al: SHARP Investigators Study Group: Sorafenib in advanced
hepatocellular carcinoma. N Engl J Med. 359:378–390. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Liang JJ, Zhou YY, Wu J and Ding Y: Gold
nanoparticle-based drug delivery platform for antineoplastic
chemotherapy. Curr Drug Metab. 15:620–631. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Dykman LA and Khlebtsov NG: Uptake of
engineered gold nanoparticles into mammalian cells. Chem Rev.
114:1258–1288. 2014. View Article : Google Scholar
|