Introduction
As one of the most common malignant diseases of the
respiratory system, lung cancer is associated with a 5-year overall
survival rate of 15% and is among the leading causes of
cancer-related deaths worldwide (1). Treatment failure is mainly caused by
the high invasive and metastatic potential, which characterizes its
high malignant potential (2). It
was reported that ~90% of malignant tumor-related deaths are due to
invasion and metastasis, which is also true for lung cancer
(3). The mechanisms of invasion of
lung cancer are complicated since multiple biological processes
such as alterations in gene expression and changes in several
signaling pathways are usually involved. Thus, identifying the
therapeutic drugs specifically targeting these processes to inhibit
the invasiveness of lung cancer is of significant importance.
A zinc-dependent proteinase family, the matrix
metalloproteinases (MMPs), are believed to participate in numerous
pathological processes due to their activity in extracellular
matrix (ECM) degradation (4). MMPs
are associated with non-neoplastic diseases including ischemia
(5), trauma (6), and neoplastic diseases including
glioma (7), and breast (8) and liver cancer (9). It is currently generally accepted that
cancer cells invade through tissue by secreting these enzymes. As
an important member of the MMPs, MMP-9 overexpression is often
detected in various human invasive cancers (10). The regulation of expression of MMP-9
is mediated by multiple activators such as NF-κB (11). The most direct induction of MMP-9
expression is through the activating transcription factor-2 (ATF-2)
(12) which further binds to the
AP-1 site of the MMP-9 gene promoter to enhance MMP-9 transcription
(13). A previous study found that
ATF-2 was activated by phosphorylation in a reactive oxygen species
(ROS)-dependent manner in fibroblasts and myocytes (14).
1,7-Bis(4-hydroxy-3-methoxyphenol)-1,6-heptadiene-3,5-dione, also
known as curcumin, is extracted and obtained from the roots of a
plant named turmeric (Curcuma longa L.). Curcumin’s
hydrophobic polyphenol molecular structure contributes to its
multiple biological functions including its anticancer activity
(15). Studies have revealed that
curcumin inhibits malignant cell migration, invasion and metastasis
by repressing expression of multiple proteins including MMPs
(16,17); however, the exact mechanisms require
further investigation.
Curcumin reduces excessive production of ROS in
mammalian cells which may lead to multiple biological effects
(18). A previous study suggested
that cellular ROS were generated in an enzymatic reaction catalyzed
by NADPH oxidase-2 (Nox-2) under external stimuli (19). Nox-2 is activated by phosphorylation
of its subunit P47phox which was suggested to be
phosphorylated by protein kinase Cα (PKCα) (20). A recent study pointed out that
curcumin inhibits phosphorylation of Nox subunits by reducing PKC
expression (21).
In this context, we hypothesized a possible
mechanism that curcumin inhibits MMP-9 expression in lung cancer
cells by inhibiting activation of PKCα/Nox-2/ATF-2 signaling. The
results in the present study broaden the knowledge of the
mechanisms of the anticancer activity of curcumin, and also provide
new clues for the molecular-targeted treatment of malignant
diseases.
Materials and methods
Cell line, culturing and treatment
Human lung cancer cell line A549 was acquired from
the Cell Resource Center of the Chinese Academy of Sciences. The
cells were cultured in cell culturing flasks (Corning) and
maintained in RPMI-1640 culture medium (HyClone) supplemented with
fetal bovine serum (FBS, 10%; HyClone), glutamine (2 mmol/l; Sigma)
and antibiotics including 100 µg/ml streptomycin and 100
U/ml penicillin (both from Sigma). Cells were cultured in a cell
culture incubator (Thermo) providing a humidified environment with
95% fresh air and 5% carbon dioxide. Cells were treated with
serially diluted curcumin (Sigma) at various concentrations (0, 15,
30 and 60 µmol/l) for 24 h.
Cell viability evaluation
Cell viability of the A549 cells was assessed by
colorimetric 3-(4,5-dimethylthiazol-2-yl) 2,5-diphenyltetrazolium
bromide (MTT) assay in accordance with standard methods. After an
equal number of cells (2×104) were plated in a 96-well
plate (Corning), the treated cells were washed with sterilized
phosphate-buffered saline (PBS; Pioneer) twice, and then incubated
with MTT (Sigma) at a final concentration of 5 mg/ml for 4 h in an
incubator. After PBS washing, the cells were dissolved in dimethyl
sulfoxide (DMSO; Sigma). The 450-nm absorbance value was read by a
plate reader (Bio-Rad). The cell viability was presented as the
inhibition rate, which was expressed as a ratio of the number of
the non-viable cells in the experimental wells (cells treated by
curcumin) compared to the control wells.
DNA and RNA transfection
Six-well plates were seeded with 5×104
cells/well in 2 ml media 24 h before transfection; the cells were
80–90% confluent. Cells were transfected with siRNA (100 pmol/well)
or plasmid DNA (4 µg/well) using Lipofectamine 2000 reagent
(Life Technologies, Grand Island, NY, USA) according to the
manufacturer’s instructions. After 48 h of transfection, the cells
were collected for subsquent experiments. For establishing stable
cell lines, the cells were selected using puromycin for 2 weeks.
Stable transductants were pooled. All siRNAs and shRNA were
purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). The
pHACE-PKCα WT plasmid (22) was
provided by Addgene (Addgene plasmid, 21232).
Cell invasion assay
The invasive ability of the A549 cells was assessed
by Transwell assays using the BioCoat Matrigel invasion assay
system (BD Biosciences) according to standard protocols. After
suspension in serum-free medium, the cells were seeded. The
Matrigel was used to coat the upper surface of the Transwell
chambers for invasion assay. After a 24-h incubation, the cells
that invaded through the membrane were fixed with methanol. The
invasion was then observed and evaluated after crystal violet
staining under an optical microscopy.
Intracellular ROS detection
The intracellular ROS level in the A549 cells was
detected by 2′,7′-dichlorofluorescein diacetate (DCFH-DA). The
cultured A549 cells were incubated with serum-free RPMI-1640 medium
supplemented with DCFH-DA (Beyotime) at a final concentration of 10
µmol/l at 47°C for 40 min. Then the fluorescence of DCFH-DA
at 530 nm was detected using the FACSCalibur flow cytometer (BD
Biosciences).
Quantitative real-time PCR
Total RNA from the A549 cells was isolated by using
the RNeasy Mini kit (Invitrogen) according to the manufacturer’s
instructions. Reverse transcription was performed, and cDNA was
synthesized using SuperScript III Reverse Transcriptase
(Invitrogen). Quantitative real-time PCR was then performed using
SYBR-Green II according to the PrimeScript RT-PCR kit protocol
(Takara). The specific primers for each gene (PKCα, Nox-2, MMP-9
and β-actin) were designed and synthesized by Takara. β-actin was
introduced as the internal control. The 2−ΔΔCt method
was used to analyze the relative expression levels for PKCα, Nox-2
and MMP-9.
Western blot analysis
The A549 cells in each group were harvested and
lysed in RIPA lysis buffering system (pH, 7.5; 40 mmol/l Tris-HCI,
150 mmol/l KCl, 100 mmol/l NaVO3, 1 mmol/l EDTA, 1%
Triton X-100 and 1 mmol/l PMSF). The concentration of the extracted
proteins was determined by the BCA kit (Thermo). The same amount of
protein (50 µg) from each group was separated by
SDS-polyacrylamide gel (8 or 10%) vertical electrophoresis and then
transferred to PVDF membranes. Defatted milk (5% in TBST buffer)
was used to block the non-specific binding by incubation with the
membranes at 37°C for 1 h. After washing, the membranes were
incubated with specific antibodies against PKCα, Nox-2 and
P47phox (all from Abcam), p-ATF-2 and ATF-2 (both from
Cell Signaling Technology) and MMP-9 and β-actin (both from Santa
Cruz Biotechnology) at 4°C for 12 h. After washing, the membranes
were then incubated by corresponding horseradish peroxidase
secondary antibodies at room temperature for 1 h. Then an enhanced
chemiluminescence kit (Amersham) was used to detect the bands.
Software Image J was used to perform the densitometric
analysis.
Statistical analysis
Data collected in the present study are expressed as
the (mean ± SD). Differences between groups were analyzed using the
Student’s t-test. P<0.05 was considered to be indicative of a
statistically significant result.
Results
Curcumin inhibits the proliferation and
invasion of A549 cells
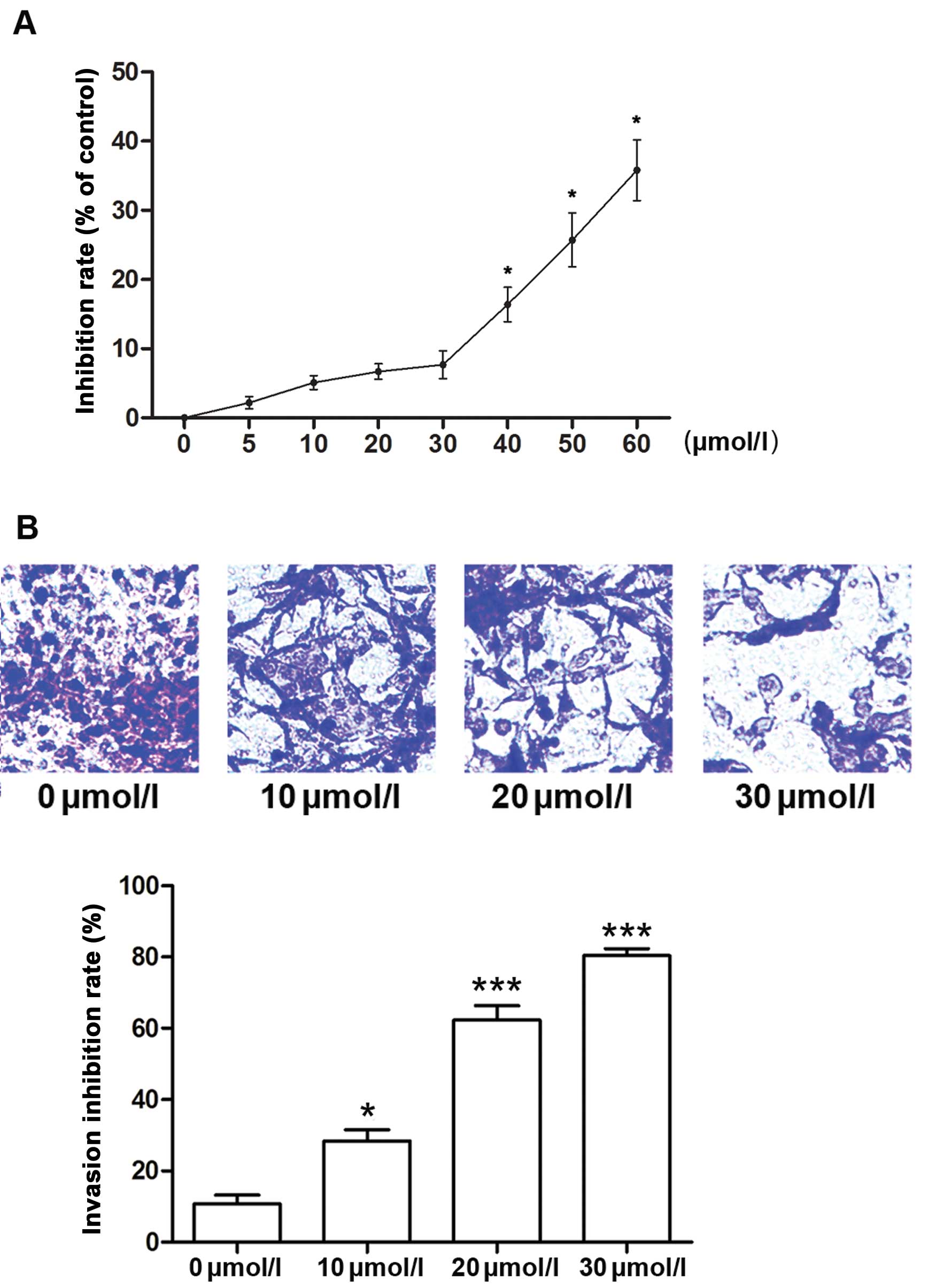
We first evaluated the effects of curcumin on cancer
cell proliferation and invasion. The MTT results showed that
curcumin inhibited the proliferation of A549 cells in a
dose-dependent manner with a significant anti-proliferative effect
>40 µmol/l (Fig. 1A). As
it was considered that the inhibition of metastasis may be
attributed to the tumor growth inhibition, the concentrations which
inhibit cell growth were excluded to avoid the cytotoxic effect of
curcumin on cell invasion. Thus, a concentration range <40
µmol/l (10, 20 and 30 µmol/l particularly in this
study) was chosen for subsequent invasion experiments. Transwell
assays demonstrated that curcumin inhibited the invasion of the
A549 cells at the above concentrations (Fig. 1B).
Curcumin inhibits the invasion of A549
cells by suppressing the expression of MMP-9
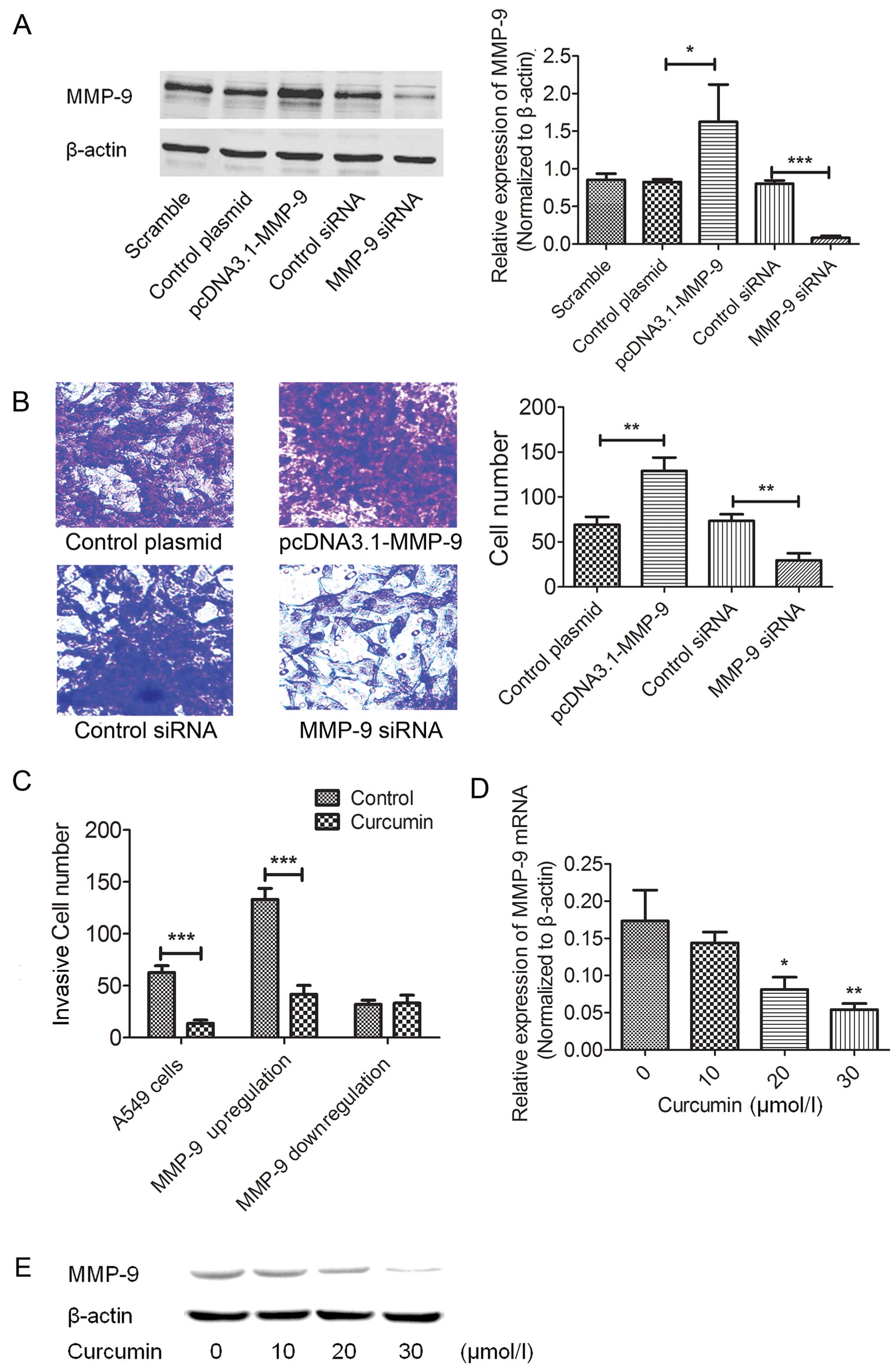
The potential involvement of MMP-9 in cellular
invasion was evaluated. MMP-9 was effectively upregulated or
downregulated by the pcDNA3.1-MMP-9 cDNA plasmid or siRNA (Fig. 2A). As shown in Fig. 2B, cell invasion was significantly
increased in the MMP-9-overexpressed cells and reduced in the
MMP-9-downregulated cells. We further tested the inhibitory effects
of curcumin on both the MMP-9 overexpressed and downregulated A549
cells. As expected, curcumin did not inhibit the invasion of the
MMP-9-downregulated A549 cells when compared with the controls
(Fig. 2C). However, in the
MMP-9-overexpressed A549 cells, curcumin still exhibited an
inhibitory effect on cell invasion (Fig. 2C). Furthermore, we also demonstrated
that curcumin dose-dependently suppressed the expression of MMP-9
in the A549 cells at both the mRNA and protein levels (Fig. 2D and E), supporting the functional
role of MMP-9 in curcumin-inhibited lung cancer cell invasion.
Involvement of PKCα in expression of
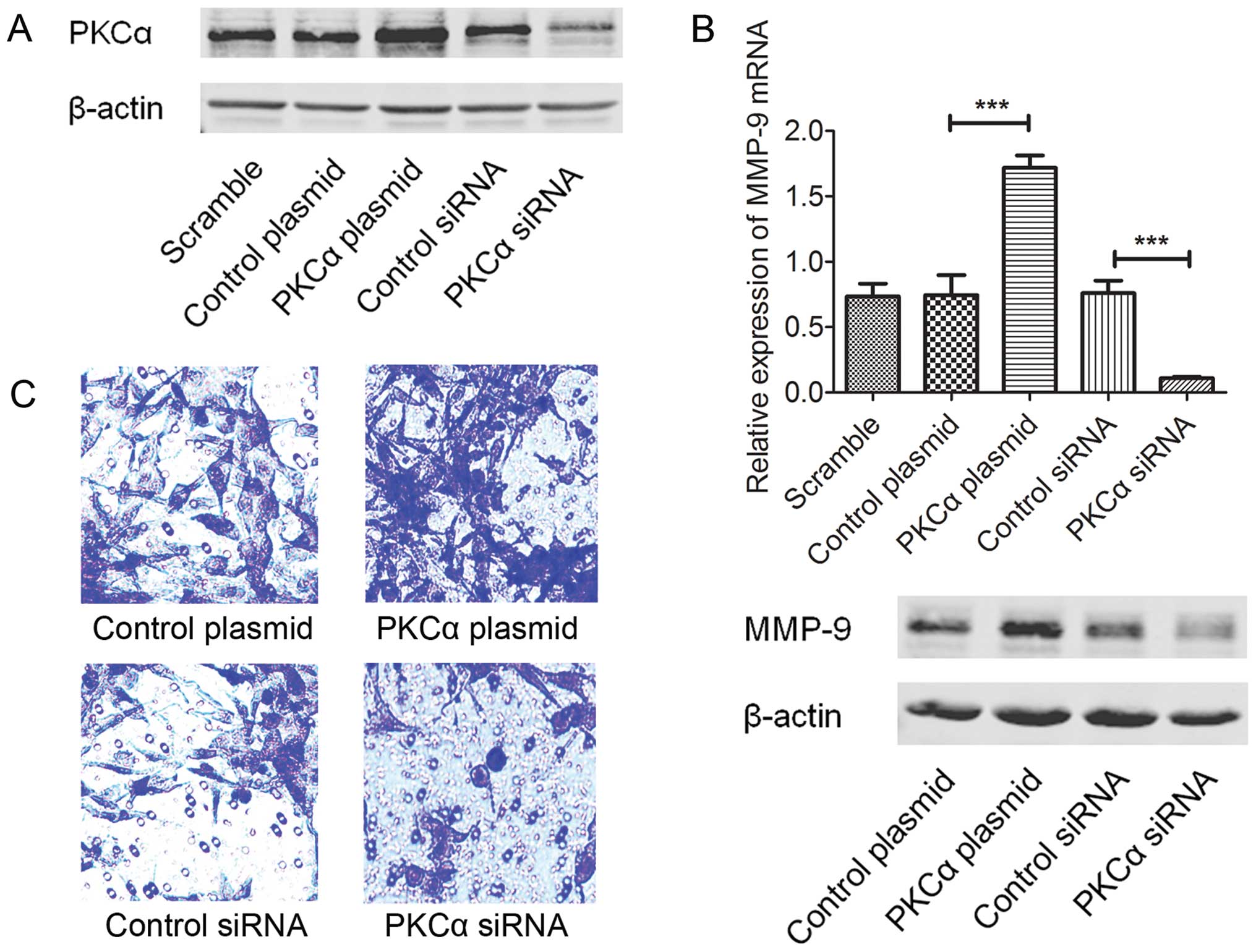
MMP-9 and cell invasion of A549 cells
The PKCα cDNA plasmid (pHACE-PKCα WT) and siRNA were
used to establish the PKCα upregulated and downregulated A549 cells
(Fig. 3A). Expression of MMP-9 was
then evaluated by real-time PCR and western blot analyses. As shown
in Fig. 3B, upregulation of PKCα
induced expression of MMP-9 and inhibition of PKCα by siRNA
suppressed the MMP-9 protein expression. In addition, we assessed
the potential roles of PKCα in the cell invasion of A549 cells.
When expression of PKCα was inhibited by siRNA, the number of
invasive A549 cells was reduced (Fig.
3C). In contrast, over-expression of PKCα increased the
invasive ability of the A549 cells (Fig. 3C).
PKCα-induced expression of MMP-9 is
Nox-2-dependent
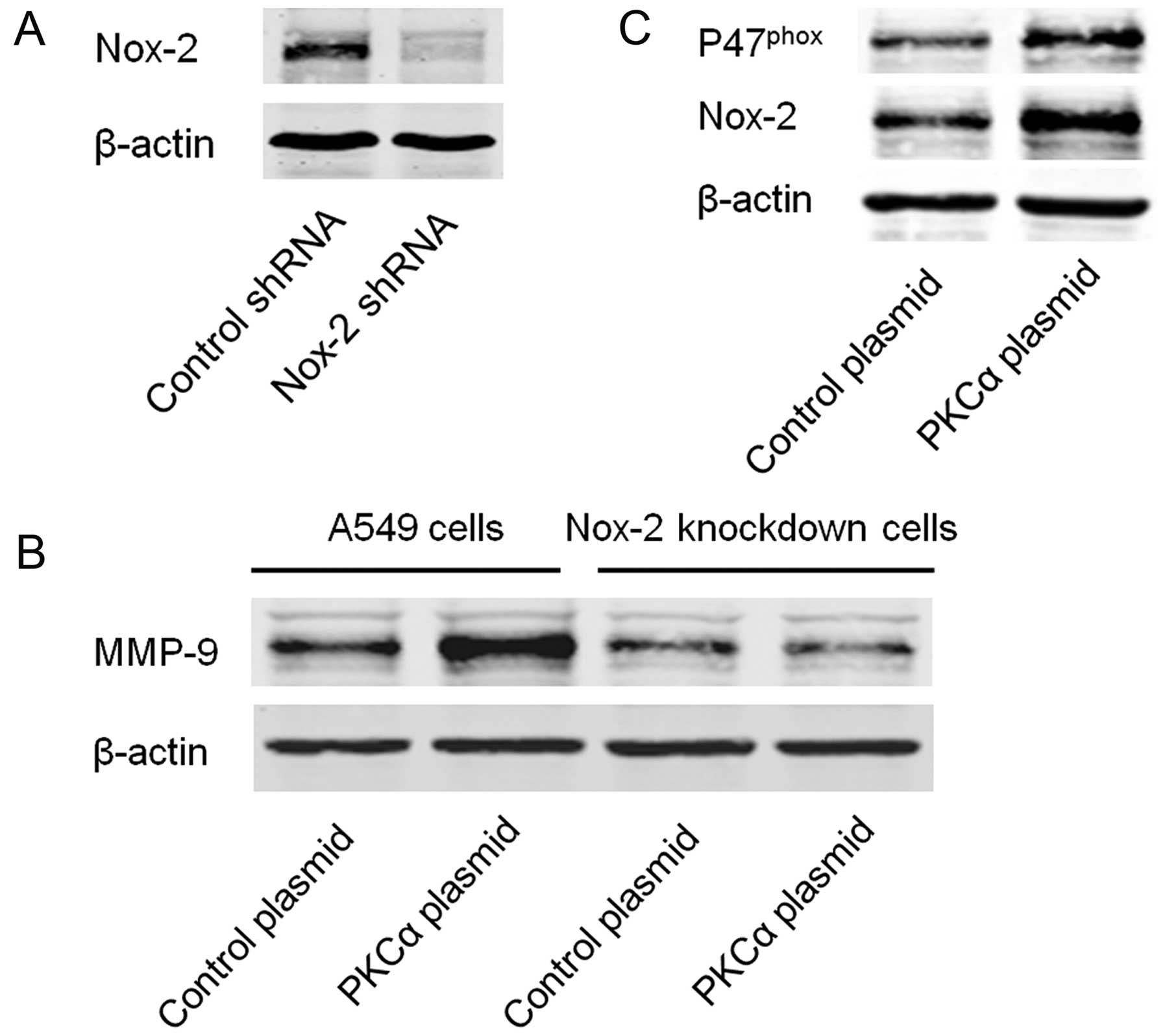
Since Nox-2 is one of the downstream molecules of
PKCα (20,21), we aimed to ascertain whether Nox-2
is involved in the PKCα/MMP-9 signaling pathway. Specific shRNA was
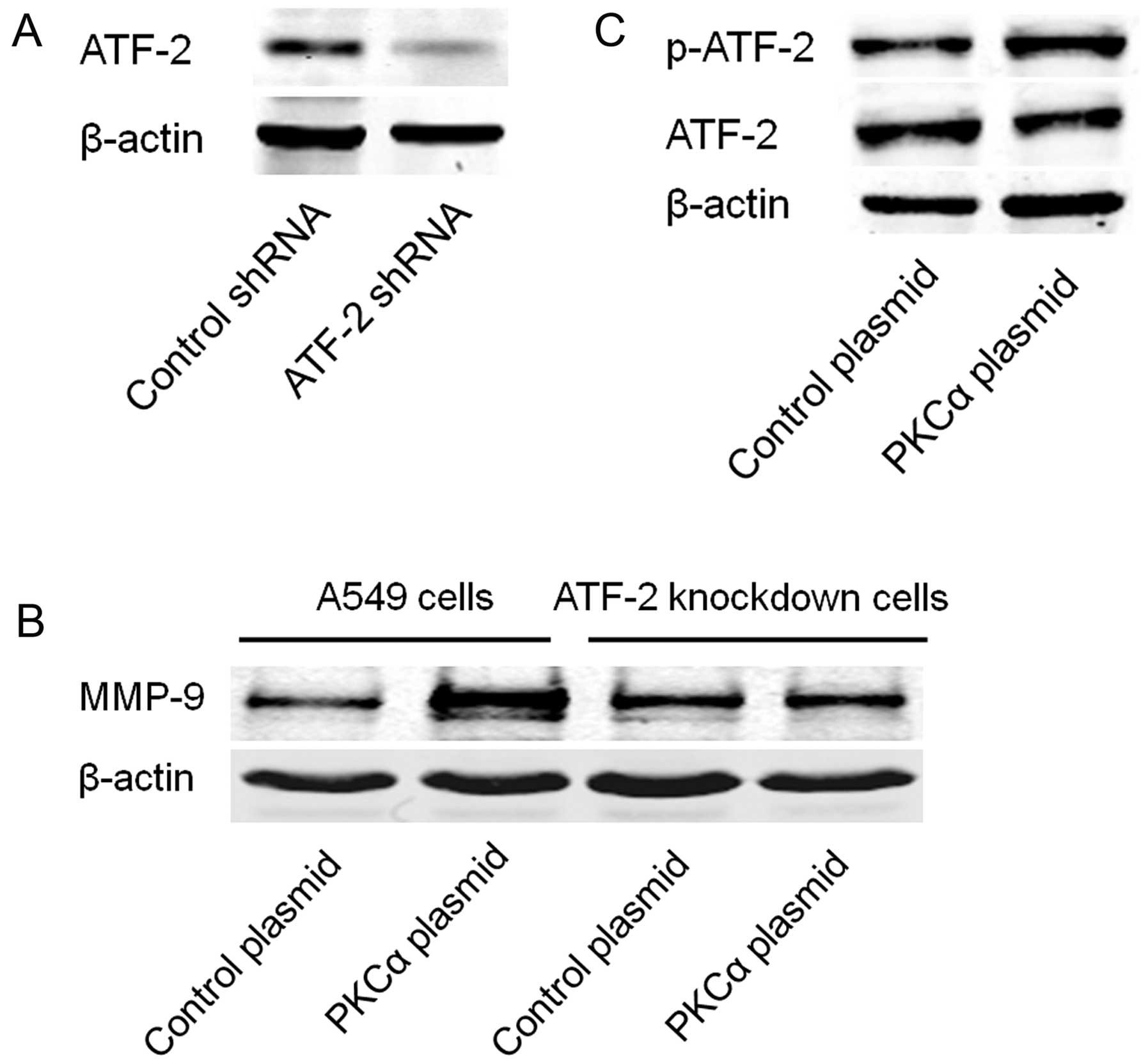
used to establish the stable Nox-2-knockdown A549 cells (Fig. 4A). Expression of MMP-9 was then
detected in the Nox-2-knockdown and control cells with or without
transfection of the PKCα cDNA plasmid. The results showed that
knockdown of Nox-2 markedly inhibited PKCα-induced expression of
MMP-9 (Fig. 4B). Moreover, we found
that over-expression of PKCα increased the expression of
P47phox and Nox-2 (Fig.
4C).
PKCα-induced expression of MMP-9 is
ATF-2-dependent
We further assessed the potential role of ATF-2 in
the PKCα/MMP-9 signaling pathway. Specific shRNA was used to
establish stable ATF-2-knockdown A549 cells (Fig. 5A). Expression of MMP-9 was then
detected in the ATF-2-knockdown and control cells with or without
transfection of the PKCα cDNA plasmid. As shown in Fig. 5B, knockdown of ATF-2 markedly
inhibited PKCα-induced expression of MMP-9. Meanwhile,
overexpression of PKCα induced phosphorylation of ATF-2 without
affecting the protein level of ATF-2 (Fig. 5C).
Curcumin inhibits activation of the
PKCα/Nox-2/ROS/ATF-2 pathway in A549 cells
Western blot analyses were used to assess the
expression of PKCα, Nox-2 and ATF-2 in the A549 cells after
treatment with curcumin. As shown in Fig. 6A, curcumin significantly inhibited
the expression of PKCα, P47phox and Nox-2 in a
dose-dependent manner without affecting the protein levels of
ATF-2. However, phosphorylated ATF-2 was markedly reduced by
curcumin, suggesting the potential role of curcumin in suppressing
the activation of ATF-2. We also detected the intracellular ROS
generation in the A549 cells by DCFH-DA fluorescence which was
determined by flow cytometry. As shown in Fig. 6B, after incubation with curcumin,
the ROS production was significantly decreased. Similar results of
decreased ROS generation were also observed in the PKCα and Nox-2
siRNA-treated A549 cells.
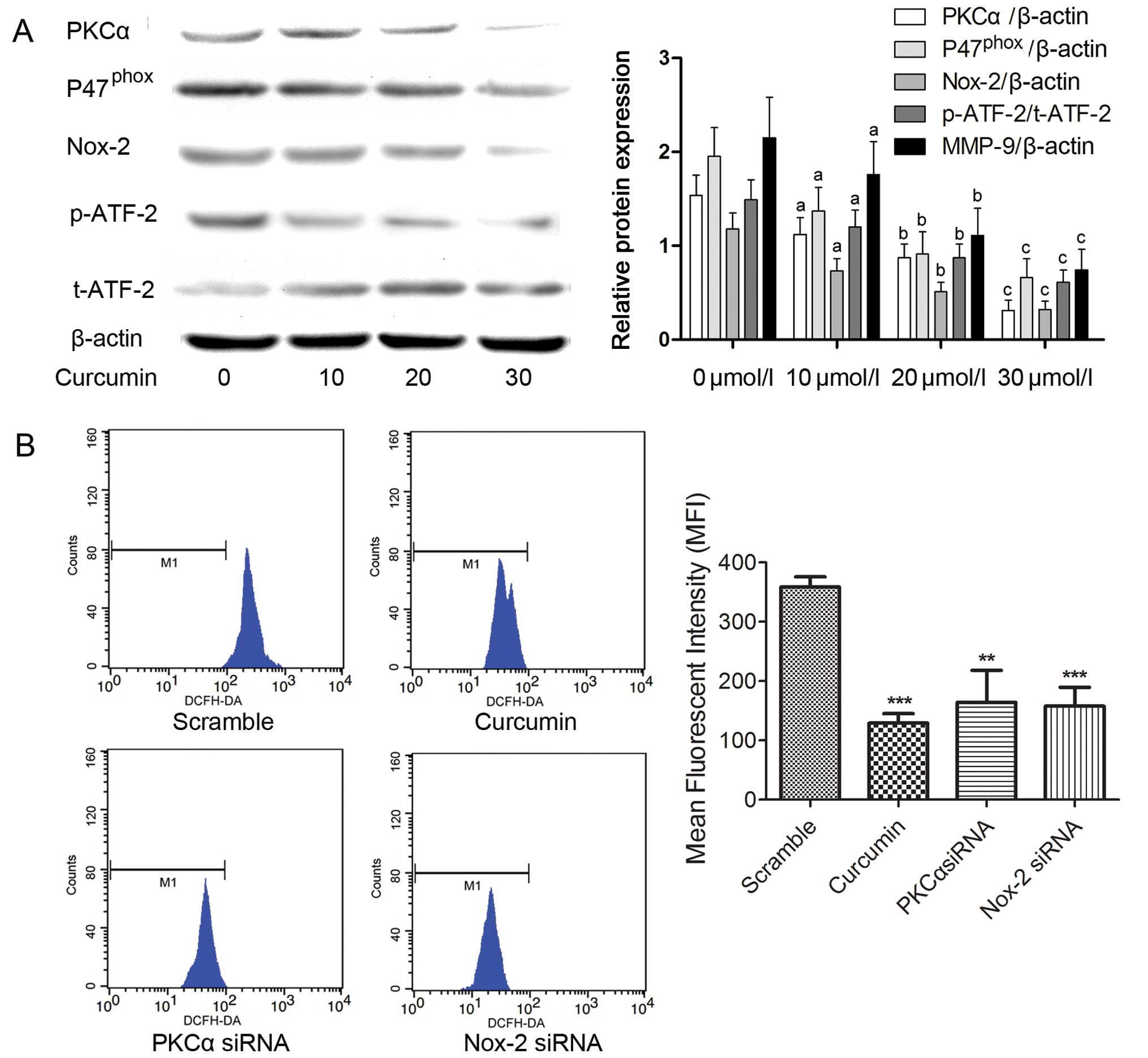 | Figure 6Curcumin inhibits the activation of
the PKCα/Nox-2/ROS/ATF-2 signaling pathway in the A549 cells. (A)
A549 cells were treated with the indicated concentrations of
curcumin. The expression of PKCα, P47phox, Nox-2,
phosphorylated (p)-ATF-2 and total (t)-ATF-2 were detected by
western blot analysis. The columns indicate the relative protein
expression of PKCα, P47phox, (p)-ATF-2 and (t)-ATF-2 in
the A549 cells treated with curcumin at the indicated
concentrations, respectively. (B) The effects of curcumin (30
µmol/l), PKCα siRNA and Nox-2 siRNA on intracellular ROS
levels in the A549 cells were assessed by detection of DCFH-DA
using a flow cytometer. The columns indicate the mean fluorescent
intensity of detected DCFH-DA for each treatment expressed as mean
± SD of three independent experiments; **P<0.01,
***P<0.001. PKCα, protein kinase Cα; ATF-2,
activating transcription factor-2; Nox-2, NADPH oxidase-2; ROS,
reactive oxygen species. |
Discussion
Various types of natural therapeutic drugs such as
ginsen-oside, baicalein and sophocarpine exhibit anticancer effects
in many human types of cancer such as hepatic, breast, cervical and
lung cancer (23–25). The therapeutic mechanisms of these
natural drugs are extremely complicated, and involve effects on
apoptotic-related genes, adhesive molecules and transcription
factors (26). In the present
study, we demonstrated that the overexpression of MMP-9 which is
associated with the invasive ability of lung cancer A549 cells was
mediated by the PKCα/Nox-2/ATF-2 signaling pathway. Furthermore, we
revealed that the anti-invasive therapeutic effect of curcumin was
based on modulating this pathway.
MMPs are a large family of proteases which play
crucial roles in the development and progression of cancer
(27). As a member of the MMPs,
MMP-9 is indispensible in tumori-genesis, invasion and metastasis
based on its collagenase activity in ECM degradation (28). Loss of ECM of blood and lymphatic
vessel walls prompts cancer cells to spread to organs by invading
into blood and lymphatic systems. According to a previous study,
the nuclear transcription factor AP-1 regulates the expression of
MMP-9 directly by binding to the AP-1 site of the promoter region
of the MMP-9 gene (29). This
binding process is believed to depend on ATF-2 which facilitates
AP-1 binding to the MMP-9 promoter after forming a heterodimer
structure based on its activation by phosphorylation (30). It was reported that the
phosphorylation of ATF-2 is induced by excessive intracellular ROS
generation (14).
Under many pathological conditions, Nox is the major
source of ROS production (31).
Nox-2, also regarded as gp91phox, is a prototype NADPH
oxidase. As a transmembrane protein, it transfers electrons through
a chain reaction to electron donors to form ROS inside cells. The
activation of Nox-2 is initiated by phosphorylation of
P47phox, the ̔organizer subunit’, which recruits
P67phox and P40phox to form the activated
Nox-2 complex (32,33). It has been suggested that the
phosphorylation of P47phox is conducted by PKCα in
neutrophils (34). Thus, the above
described molecules form a signaling pathway which could be
referred as PKCα-stimulated Nox-2-dependent ROS-activated
ATF-2-induced MMP-9 expression.
In the present study, RNA interference technique was
employed to confirm the existence of this pathway in lung cancer
cells. Knockdown of both PKCα and Nox-2 not only reduced the
production of intracellular production of ROS, but also markedly
impaired MMP-9 expression in the lung cancer A549 cells. As a
result, PKCα and Nox-2 knockdown suppressed the invasive ability of
A549 cells. These results indicate that PKCα/Nox-2/ROS/ATF-2/MMP-9
signaling is activated to induce invasion.
With a long history, from ancient times, curcumin
has been used as a herbal remedy in traditional Chinese and Indian
medicine in the treatment of numerous diseases (35). In recent decades, studies have
identified numerous therapeutic activities including antioxidant,
antidiabetic, anti-inflammatory and anticancer for this plant
polyphenol, which is extracted from the roots of the spice turmeric
(Curcuma longa L.) (36,37).
Curcumin exerts its inhibitory effects on the invasion and
metastasis of many human cancers which are responsible for
malignant tumor-related mortality. Several mechanisms of the
effects of curcumin have been described in previous studies by
investigating MMPs (38), adhesion
molecules (39), angiogenesis
(40) and the tumor
microenvironment (41). We observed
that the invasive ability of the A549 cells was significantly
inhibited by curcumin treatment via reduced MMP-9 expression. The
results also indicated that the MMP-9 reduction was due to
curcumin’s effect of downregulating the PKCα/Nox-2/ROS/ATF-2
signaling pathway in lung cancer A549 cells.
The PKCα/Nox-2/ROS/ATF-2/MMP-9 signaling pathway is
activated in lung cancer A549 cells to maintain invasiveness.
Curcumin impairs cell invasion by modulating the
PKCα/Nox-2/ROS/ATF-2 signaling pathway to reduce MMP-9 expression
in the A549 cells.
References
|
1
|
Minami-Shimmyo Y, Ohe Y, Yamamoto S, Sumi
M, Nokihara H, Horinouchi H, Yamamoto N, Sekine I, Kubota K and
Tamura T: Risk factors for treatment-related death associated with
chemotherapy and thoracic radiotherapy for lung cancer. J Thorac
Oncol. 7:177–182. 2012. View Article : Google Scholar
|
|
2
|
Hung JJ, Jeng WJ, Hsu WH, Chou TY, Huang
BS and Wu YC: Predictors of death, local recurrence, and distant
metastasis in completely resected pathological stage-I
non-small-cell lung cancer. J Thorac Oncol. 7:1115–1123. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Schiavina R, Borghesi M, Brunocilla E,
Manferrari F, Fiorentino M, Vagnoni V, Baccos A, Pultrone CV, Rocca
GC, Rizzi S, et al: Differing risk of cancer death among patients
with lymph node metastasis after radical prostatectomy and pelvic
lymph node dissection: Identification of risk categories according
to number of positive nodes and Gleason score. BJU Int.
111:1237–1244. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mizutani K, Kofuji K and Shirouzu K: The
significance of MMP-1 and MMP-2 in peritoneal disseminated
metastasis of gastric cancer. Surg Today. 30:614–621. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhang HM, Han YJ, Zhao WB, Hu LJ, Fei YT
and Tu Y: Effects of scalp acupuncture on expression of hippocampal
MMP-9 in cerebral ischemia injury rats. Zhen Ci Yan Jiu.
36:193–198. 2011.In Chinese. PubMed/NCBI
|
|
6
|
Kim HJ, Fillmore HL, Reeves TM and
Phillips LL: Elevation of hippocampal MMP-3 expression and activity
during trauma-induced synaptogenesis. Exp Neurol. 192:60–72. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sun C, Wang Q, Zhou H, Yu S, Simard AR,
Kang C, Li Y, Kong Y, An T, Wen Y, et al: Antisense MMP-9 RNA
inhibits malignant glioma cell growth in vitro and in vivo.
Neurosci Bull. 29:83–93. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lin ZM, Zhao JX, Duan XN, Zhang LB, Ye JM,
Xu L and Liu YH: Effects of tissue factor, PAR-2 and MMP-9
expression on human breast cancer cell line MCF-7 invasion. Asian
Pac J Cancer Prev. 15:643–646. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Oshima T, Kunisaki C, Yoshihara K, Yamada
R, Yamamoto N, Sato T, Makino H, Yamagishi S, Nagano Y, Fujii S, et
al: Clinicopathological significance of the gene expression of
matrix metalloproteinases and reversion-inducing cysteine-rich
protein with Kazal motifs in patients with colorectal cancer: MMP-2
gene expression is a useful predictor of liver metastasis from
colorectal cancer. Oncol Rep. 19:1285–1291. 2008.PubMed/NCBI
|
|
10
|
Lee LY, Wu CM, Wang CC, Yu JS, Liang Y,
Huang KH, Lo CH and Hwang TL: Expression of matrix
metalloproteinases MMP-2 and MMP-9 in gastric cancer and their
relation to claudin-4 expression. Histol Histopathol. 23:515–521.
2008.PubMed/NCBI
|
|
11
|
Park SY, Kim YH, Kim Y and Lee SJ:
Frondoside A has an anti-invasive effect by inhibiting TPA-induced
MMP-9 activation via NF-κB and AP-1 signaling in human breast
cancer cells. Int J Oncol. 41:933–940. 2012.PubMed/NCBI
|
|
12
|
Kim ES, Sohn YW and Moon A:
TGF-beta-induced transcriptional activation of MMP-2 is mediated by
activating transcription factor (ATF)2 in human breast epithelial
cells. Cancer Lett. 252:147–156. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hsieh HL, Lin CC, Shih RH, Hsiao LD and
Yang CM: NADPH oxidase-mediated redox signal contributes to
lipoteichoic acid-induced MMP-9 upregulation in brain astrocytes. J
Neuroinflammation. 9:1102012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu WH and Chang LS: Arachidonic acid
induces Fas and FasL upregulation in human leukemia U937 cells via
Ca2+/ROS-mediated suppression of ERK/c-Fos pathway and
activation of p38 MAPK/ATF-2 pathway. Toxicol Lett. 191:140–148.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ji JL, Huang XF and Zhu HL: Curcumin and
its formulations: Potential anti-cancer agents. Anticancer Agents
Med Chem. 12:210–218. 2012. View Article : Google Scholar
|
|
16
|
Chen CC, Sureshbabul M, Chen HW, Lin YS,
Lee JY, Hong QS, Yang YC and Yu SL: Curcumin suppresses metastasis
via Sp-1, FAK inhibition, and E-cadherin upregulation in colorectal
cancer. Evid Based Complement Alternat Med. 2013:5416952013.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen HW, Lee JY, Huang JY, Wang CC, Chen
WJ, Su SF, Huang CW, Ho CC, Chen JJ, Tsai MF, et al: Curcumin
inhibits lung cancer cell invasion and metastasis through the tumor
suppressor HLJ1. Cancer Res. 68:7428–7438. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chan WH, Wu HJ and Hsuuw YD: Curcumin
inhibits ROS formation and apoptosis in methylglyoxal-treated human
hepatoma G2 cells. Ann NY Acad Sci. 1042:372–378. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Carbone F, Camillo Teixeira P,
Braunersreuther V, Mach F, Vuilleumier N and Montecucco F:
Pathophysiology and treatments of oxidative injury in ischemic
stroke: Focus on the phagocytic NADPH oxidase 2. Antioxid Redox
Signal. Apr 22–2014.Epub ahead of print. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Makino J, Kamiya T, Hara H and Adachi T:
TPA induces the expression of EC-SOD in human monocytic THP-1
cells: Involvement of PKC, MEK/ERK and NOX-derived ROS. Free Radic
Res. 46:637–644. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Soetikno V, Watanabe K, Sari FR, Harima M,
Thandavarayan RA, Veeraveedu PT, Arozal W, Sukumaran V, Lakshmanan
AP, Arumugam S, et al: Curcumin attenuates diabetic nephropathy by
inhibiting PKC-α and PKC-β1 activity in streptozotocin-induced type
I diabetic rats. Mol Nutr Food Res. 55:1655–1665. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Soh JW and Weinstein IB: Roles of specific
isoforms of protein kinase C in the transcriptional control of
cyclin D1 and related genes. J Biol Chem. 278:34709–34716. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen XP, Qian LL, Jiang H and Chen JH:
Ginsenoside Rg3 inhibits CXCR4 expression and related migrations in
a breast cancer cell line. Int J Clin Oncol. 16:519–523. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen J, Li Z, Chen AY, Ye X, Luo H, Rankin
GO and Chen YC: Inhibitory effect of baicalin and baicalein on
ovarian cancer cells. Int J Mol Sci. 14:6012–6025. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li LQ, Li XL, Wang L, Du WJ, Guo R, Liang
HH, Liu X, Liang DS, Lu YJ, Shan HL, et al: Matrine inhibits breast
cancer growth via miR-21/PTEN/Akt pathway in MCF-7 cells. Cell
Physiol Biochem. 30:631–641. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Saha SK and Khuda-Bukhsh AR: Molecular
approaches towards development of purified natural products and
their structurally known derivatives as efficient anti-cancer
drugs: Current trends. Eur J Pharmacol. 714:239–248. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Schmalfeldt B, Prechtel D, Härting K,
Späthe K, Rutke S, Konik E, Fridman R, Berger U, Schmitt M, Kuhn W,
et al: Increased expression of matrix metalloproteinases (MMP)-2,
MMP-9, and the urokinase-type plasminogen activator is associated
with progression from benign to advanced ovarian cancer. Clin
Cancer Res. 7:2396–2404. 2001.PubMed/NCBI
|
|
28
|
Mehner C, Hockla A, Miller E, Ran S,
Radisky DC and Radisky ES: Tumor cell-produced matrix
metalloproteinase 9 (MMP-9) drives malignant progression and
metastasis of basal-like triple negative breast cancer. Oncotarget.
5:2736–2749. 2014.PubMed/NCBI
|
|
29
|
Kim HS, Kim MH, Jeong M, Hwang YS, Lim SH,
Shin BA, Ahn BW and Jung YD: EGCG blocks tumor promoter-induced
MMP-9 expression via suppression of MAPK and AP-1 activation in
human gastric AGS cells. Anticancer Res. 24:747–753.
2004.PubMed/NCBI
|
|
30
|
Yeh JH, Lecine P, Nunes JA, Spicuglia S,
Ferrier P, Olive D and Imbert J: Novel CD28-responsive enhancer
activated by CREB/ATF and AP-1 families in the human interleukin-2
receptor alpha-chain locus. Mol Cell Biol. 21:4515–4527. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Jendrysik MA, Vasilevsky S, Yi L, Wood A,
Zhu N, Zhao Y, Koontz SM and Jackson SH: NADPH oxidase-2 derived
ROS dictates murine DC cytokine-mediated cell fate decisions during
CD4 T helper-cell commitment. PLoS One. 6:e281982011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kuo FC, Tseng YT, Wu SR, Wu MT and Lo YC:
Melamine activates NF-κB/COX-2/PGE2 pathway and increases NADPH
oxidase-dependent ROS production in macrophages and human embryonic
kidney cells. Toxicol In Vitro. 27:1603–1611. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jiang Q, Zhou C, Healey S, Chu W, Kouttab
N, Bi Z and Wan Y: UV radiation down-regulates Dsg-2 via Rac/NADPH
oxidase-mediated generation of ROS in human lens epithelial cells.
Int J Mol Med. 18:381–387. 2006.PubMed/NCBI
|
|
34
|
Remijsen QF, Fontayne A, Verdonck F,
Clynen E, Schoofs L and Willems J: The antimicrobial peptide
parabutoporin competes with p47(phox) as a PKC-substrate and
inhibits NADPH oxidase in human neutrophils. FEBS Lett.
580:6206–6210. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Anstrom DM, Zhou X, Kalk CN, Song B and
Lan Q: Mosquitocidal properties of natural product compounds
isolated from Chinese herbs and synthetic analogs of curcumin. J
Med Entomol. 49:350–355. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yılmaz Savcun G, Ozkan E, Dulundu E,
Topaloğlu U, Sehirli AO, Tok OE, Ercan F and Sener G: Antioxidant
and anti-inflammatory effects of curcumin against hepatorenal
oxidative injury in an experimental sepsis model in rats. Ulus
Travma Acil Cerrahi Derg. 19:507–515. 2013. View Article : Google Scholar
|
|
37
|
Meng B, Li J and Cao H: Antioxidant and
antiinflammatory activities of curcumin on diabetes mellitus and
its complications. Curr Pharm Des. 19:2101–2113. 2013.
|
|
38
|
Mo N, Li ZQ, Li J and Cao YD: Curcumin
inhibits TGF-β1-induced MMP-9 and invasion through ERK and Smad
signaling in breast cancer MDA-MB-231 cells. Asian Pac J Cancer
Prev. 13:5709–5714. 2012. View Article : Google Scholar
|
|
39
|
Pan Y, Zhang X, Wang Y, Cai L, Ren L, Tang
L, Wang J, Zhao Y, Wang Y, Liu Q, et al: Targeting JNK by a new
curcumin analog to inhibit NF-κB-mediated expression of cell
adhesion molecules attenuates renal macrophage infiltration and
injury in diabetic mice. PLoS One. 8:e790842013. View Article : Google Scholar
|
|
40
|
Bimonte S, Barbieri A, Palma G, Luciano A,
Rea D and Arra C: Curcumin inhibits tumor growth and angiogenesis
in an orthotopic mouse model of human pancreatic cancer. Biomed Res
Int. 2013:8104232013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Vishvakarma NK, Kumar A and Singh SM: Role
of curcumin- dependent modulation of tumor microenvironment of a
murine T cell lymphoma in altered regulation of tumor cell
survival. Toxicol Appl Pharmacol. 252:298–306. 2011. View Article : Google Scholar : PubMed/NCBI
|